
Abstract
Among clinically isolated β-hemolytic streptococci, Streptococcus pyogenes and S. agalactiae were considered the main pathogens in humans until recently. In 1996, S. dysgalactiae subsp. equisimilis (SDSE) was proposed as a novel taxon among human-derived streptococcal isolates. SDSE has Lancefield group C or G antigens, exhibits strong β-hemolysis, and exerts streptokinase activity upon human plasminogen and proteolytic activity upon human fibrin. Similarly to group A streptococci, SDSE possesses virulence factors including M protein, streptolysin O, streptolysin S, streptokinase, hyaluronidase, C5a peptidase, and others. SDSE may exist among the normal flora of the skin, oropharynx, and gastrointestinal and genitourinary tracts. In the twenty-first century, invasive SDSE infection (i.e., cellulitis, urosepsis, and pneumonia) leading to various disseminated diseases is being diagnosed increasingly in Japan, elsewhere in Asia, in Europe, and in America. Particularly, among elderly patients, these invasive diseases are encountered increasingly in Japanese hospital emergency departments. Analysis of the part of the emm gene encoding the amino acid sequence at the N-terminal end of the M protein is used to determine the molecular epidemiology of SDSE. The distribution of emm types from patients with invasive or noninvasive infections differs between surveillance results from different countries. In this review, we summarize the characteristics of phenotypes and virulence factors in SDSE strains; the review also focuses on emerging SDSE infectious disease and future vaccination research.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
The β-hemolytic streptococci are common pathogens causing community-acquired infections. To distinguish between species of β-hemolytic streptococci, clinical laboratories usually use Lancefield typing, which is based on sero-agglutination involving C-polysaccharide antigens on the bacterial cell surface. In general, β-hemolytic streptococci isolated from humans may possess any of Lancefield group A, B, C, G, F, or L antigens. Of these isolates, only group A streptococci (GAS; Streptococcus pyogenes) [1] and group B streptococci (GBS; S. agalactiae) [2, 3] were considered important pathogens in clinical settings until recently. Other streptococci belonging to groups C or G were thought to show extremely low pathogenicity, except in patients with certain predisposing medical conditions.
In 1996, Vandamme et al. [4] proposed that a novel subspecies, S. dysgalactiae subsp. equisimilis (SDSE), was a clinical pathogen. This microorganism possesses group C or G antigens (rarely, A antigen), and exhibits strong β-hemolysis. Distinct categories of C or G streptococci include the S. anginosus group, consisting of S. anginosus, S. constellatus subspecies constellatus, S. constellatus subsp. pharyngis, and S. intermedius. The S. anginosus group is easily distinguished from SDSE on the basis of biologic characteristics and colony size on sheep blood agar plates.
Beginning around 2000, invasive infections such as bacteremia caused by SDSE have been reported increasingly worldwide, as well as those caused by GAS and GBS [5–8].
In this review, we summarize the characteristics of phenotypes and virulence factors in SDSE strains, and describe the emergence of invasive SDSE infection in Japan and other countries. Our review also provides perspectives on SDSE as an emerging infectious disease and on future vaccination research.
The novel taxon and its biologic characteristics
Phenotypic characteristics including Lancefield antigen, hemolysis type, colony size, hosts, and biochemical properties are summarized for the important β-hemolytic streptococci in Table 1 [9]. Lancefield serologic differentiation [10] does not correspond to individual species of streptococci.
For example, streptococci having group C antigen include the following taxa; β-hemolytic Streptococcus dysgalactiae isolated from human specimens, β-hemolytic S. dysgalactiae from porcine specimens, α-hemolytic S. dysgalactiae from bovine specimens [11], 2 β-hemolytic subspecies of Streptococcus equi (subsp. equi and subsp. zooepidemicus) from equine specimens, and a β-hemolytic small-colony Streptococcus anginosus group from human specimens [12, 13].
Analogously, streptococci having group G antigen include S. dysgalactiae isolated from human specimens, an S. anginosus group from human specimens, and β-hemolytic Streptococcus canis from bovine, canine, and feline specimens [14]. The β-hemolytic streptococci with group L antigen also belong to S. dysgalactiae [15]. Thus, S. dysgalactiae consists of at least 5 distinct subgroups based on the findings of serogroup, host, and hemolysis type.
In 1996, two subspecies of S. dysgalactiae were proposed as new taxa by Vandamme and colleagues [4]: SDSE and S. dysgalactiae subspecies dysgalactiae (SDSD). The former, isolated from humans and animals, showed strong β-hemolysis, and the latter, isolated only from animals, showed α-, β-, or no hemolysis.
SDSE commonly possesses group G or C antigens and rarely group A antigen [16–19]. This pathogen forms large, glossy colonies that produce strong β-hemolysis on sheep blood agar plates after CO2 incubation at 37°C, as opposed to hemolysis from GAS. However, rarely, SDSE may show α- or no hemolysis [20–22], similar to GAS.
The main biologic characteristics of SDSE are resistance to bacitracin, negativity to the pyrrolidonyl-arylamidase (PYR) test, negativity to the Voges–Proskauer test, positivity to the β-d-glucuronidase test, and negativity to the β-galactosidase test [23]. The PYR test is very useful for distinguishing SDSE from GAS, which uniformly shows positivity.
Figure 1 shows typical features of colonies of GAS and SDSE isolated from human or porcine specimens, and SDSD isolated from a porcine specimen, as well as S. anginosus from a human specimen.

Morphologic and hemolytic features of colonies of Streptococcus pyogenes from a human specimen (a), S. dysgalactiae subsp. equisimilis from a human specimen (b), S. dysgalactiae subsp. equisimilis from a porcine specimen (c), S. dysgalactiae subsp. dysgalactiae from a porcine specimen (d), and S. anginosus from a human specimen (e). These microorganisms were cultured on sheep blood agar plates in a 5% CO2 atmosphere at 37°C for 20 h
Molecular characteristics of SDSE
SDSE possesses many virulence factors that are similar to those of GAS [24]. These factors may be divided into 3 categories, including bacterial products contributing to cell attachment or escape from host immunity (i.e., resistance to phagocytosis), toxins and proteases, and regulatory factors controlling expression of the products.
Factors contributing to cell attachment or escape from host immunity include M protein (encoded by the emm gene), streptokinase A (ska), streptococcal C5a peptidase (scpA), hyaluronidase (hyl), fibronectin binding protein (fbp), collagen binding protein, laminin binding protein (lmb), and others. Streptolysin S (sagA) [25, 26], streptolysin O (slo), and hemolysin (hlyIII, helA1) represent toxins or proteases, while M protein transacting positive regulation (mga) [27], CovR (covR), CovS (covS), and others are regulators of product expression.
Among these virulence factors, sequence analysis of the emm gene, which encodes the M protein on the cell surface, is often used in molecular epidemiologic studies concerning outbreaks of invasive or noninvasive SDSE infections. The first 300 highly variable bases of the 5′ end of emm gene sequences for the SDSE strains were sent online to the Centers for Disease Control (CDC) for comparison with those in the CDC emm sequence database (http://www.cdc.gov/ncidod/biotech/strep/strepblast.htm) [3, 28, 29]. An emm genotype showing more than 98% homology with the CDC reference strain was identified as that particular emm type.
Streptokinase, which dissolves human fibrin [30], and C5a peptidase, which dissolves the complement component C5a, and also inhibits neutrophil migration [30], are the major factors associated with invasive SDSE infection.
Miyoshi-Akiyama and Murayama recently completed whole-genome analyses of the original 2 SDSE isolates [GGS_124 (GenBank accession no. AP010935) and RE378], demonstrating a 61 to 63% overlap between this subspecies and GAS genomes; overlap between the subspecies and GBS genomes was 15% (unpublished data). Ahmad and colleagues [31] have deduced genetic relationships from emm genotyping and multilocus sequence typing of SDSE from isolates collected in the United States, concluding that phylogenetic relationships between SDSE and GAS alleles reflected a history of interspecies recombination, with either species frequently serving as the genetic donor. Accordingly, the genomic characteristics of SDSE appear to differ from those of GBS, being somewhat more similar to those of GAS. In further investigations, virulence factors specific to SDSE as opposed to GAS should be determined to clarify the pathogenesis of SDSE infection.
Emergence of invasive SDSE infections
SDSE may sometimes be isolated as normal flora from the skin, oropharynx, and gastrointestinal and genitourinary tracts, also having been identified in respiratory tract specimens from patients with noninvasive SDSE diseases [29]. Invasive infection is defined as the isolation of SDSE from a normally sterile site (i.e., blood, cerebrospinal fluid, joint fluid, ascites, or pleural effusion) [3, 23, 29]. Since 2003, the prevalence of invasive SDSE infections, including streptococcal toxic shock syndrome (STSS) and severe soft tissue infection [26, 30, 32], has increased gradually each year in Japan. Several SDSE cases including thoracic empyema with gas formation [33], an apparently organizing pneumonia [34], gas gangrene in both legs resulting in amputation of the right leg [35], and acute peritonitis with salpingitis associated with STSS [22], have been reported in Japan.
Our group (Ubukata and colleagues) [3] conducted active laboratory-based surveillance for invasive SDSE, GAS, and GBS infections from August 2006 to July 2007, including 142 medical institutions participating in the Invasive Streptococcal Disease Working Group established at the 19th Annual Meeting of the Japanese Society for Clinical Microbiology. Clinical isolates (n = 286) from patients with invasive infections were identified as SDSE strains, while GAS (n = 116) and GBS (n = 183) isolates were also collected until the present time (end of 2009).
Figure 2 shows age distribution according to our original data, showing a difference between patients with invasive SDSE disease and those with GAS or GBS infections. All patients with invasive SDSE infection were adults, who often were elderly, while GBS infected some patients 4 months old or younger, in addition to adults. Another investigation [29] also indicated that invasive SDSE infection (n = 42) occurred mostly in patients at least 50 years old, especially in elderly adults (60–80 years).

Numbers of patients with invasive infections caused by Streptococcus dysgalactiae subsp. equisimilis (n = 281), S. pyogenes (n = 112), and S. agalactiae (n = 180), shown by age group. Patients with insufficient data were excluded. Data were modified from those of Ref. [3]
Underlying medical conditions in subjects with GAS infection (60.3%) were less frequent than in patients with SDSE (78.8%) or GBS (88.2%) [3]. In another study [29], severe underlying conditions (i.e., diabetes mellitus, liver or renal dysfunction, and others) were associated with 85.7% of invasive SDSE infections.
Figure 3 indicates the hospital departments at which patients with invasive SDSE, GAS, or GBS diseases presented. Some patients with SDSE infection presented to the department of orthopedics as well as to the emergency and internal medicine departments [3].

Hospital departments at which patients with invasive disease caused by Streptococcus dysgalactiae subsp. equisimilis (n = 287), S. pyogenes (n = 116), and S. agalactiae (n = 183) presented. Patients with insufficient data were excluded. Data were modified from those of Ref. [3]
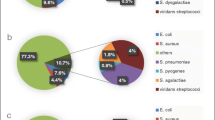
Invasive SDSE infections included sepsis with no known focus, cellulitis, septic arthritis, pneumonia, necrotizing fasciitis, meningitis, infectious endocarditis, STSS, abscesses at sites other than skin, osteomyelitis, and others [3]. Interestingly, sepsis with no known focus was more frequent among GBS-infected patients than among SDSE-infected or GAS-infected patients, while cellulitis was less frequent among GBS-infected patients than among SDSE-infected or GAS-infected patients [3] (Fig. 4). Patients infected with SDSE presented more often with septic arthritis than those infected with GBS, while patients with GAS infection had abscesses involving sites deeper than the skin more often than did patients with SDSE infection [3].

Percentages of clinical syndromes among patients at least 15 years old with invasive infection caused by Streptococcus dysgalactiae subsp. equisimilis (n = 263), S. pyogenes (n = 87), and S. agalactiae (n = 141). Patients with insufficient data were excluded. STSS, Streptococcal toxic shock syndrome. Data were modified from those of Ref. [3]
Concerning the disease outcome of SDSE infection, rates of mortality and post-infection sequelae were 12.7 and 5.3%, respectively [3]. No difference in the frequency of poor outcome (death or post-infective sequelae) was evident between SDSE, GAS, and GBS infections. In addition, poor leukocytic responses (<5000 cells/μL) and thrombocytopenia on admission were associated with a high risk of poor outcome of invasive SDSE infection [3].
The emm type stG6792 confirmed most frequently in SDSE isolates (n = 65; 22.7%), was more strongly related to poor outcome of SDSE disease than were other SDSE emm types [3]. We should continue to survey invasive SDSE infections to further clarify relationships between outcome and clinical laboratory data or emm types in the Japanese population.
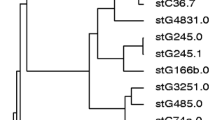
As for emm types of SDSE in our investigation, most strains showed stG6792.3 (n = 54) and displayed similar DNA profiles with pulsed-field gel electrophoresis (PFGE), suggesting the clonal expansion of a specific subpopulation of strains rather than the spread of distinct strains [3] (Fig. 5). Judging from the CDC database concerning emm type sequences, the stG6792.3 reference strain appeared to be derived from a streptococcal isolate in India, suggesting that this strain might have spread from India to Japan. In another study [29], 3 emm types, stG6792, stG485, and stG2078, predominated among the 42 invasive strains; strains with the same emm type showed uniform DNA profiles by PFGE. On the other hand, previous reports described emm types and the DNA profiles by PFGE as variable among SDSE strains [26, 30]. Person-to-person transmission routes of SDSE should be determined in detail by future surveillance.

Distributions of emm genotypes for Streptococcus dysgalactiae subsp. equisimilis strains compared between Japan (n = 286) and the United States (n = 212). Inset shows similar DNA profiles (ApaI digests) of stG6792.3 (n = 27) and stG6792.0 (n = 1, right) strains (Japan) according to pulsed-field gel electrophoresis. NT Non-typeable. Data were modified from those of Refs. [3, 5]
The prevalence of invasive SDSE infection has increased over the years elsewhere in Asia [6, 36], in Europe [37–43], and in America [5, 31, 44], as well as in Japan. As for age distribution, population-based surveillance carried out by Broyles et al. (n = 212) [5] found that most patients with invasive diseases (59.0%) were adults less than 65 years old, while the majority in our study (74.0%) were at least 65 years old [3]. Regarding underlying diseases in United States surveillance reports, 96.2% of patients with invasive infection possessed underlying medical conditions [5], while in Japan 78.8% had an underlying illness [3]. As for the specific clinical syndromes representing invasive infection, the largest category in the United States was cellulitis (41.0%) [5], while in Japan, sepsis with no focus, at 41.4%, was the largest category [3]. Prospective surveillance studies of invasive infection in both countries carried out during the same period will be needed to better define associations between host factors and the pathogenesis of SDSE.
Concerning SDSE disease outcome, the mortality rate in our Japanese observation (12.7%) was similar to those previously described in Hong Kong and the United States (12 and 15%) [3, 5, 45]. Liao et al. [6] reported several recurrent cases (8 patients, with 1 having 4 episodes and 3 having 3 episodes) of invasive SDSE disease (primary bacteremia or cellulitis). PFGE performed upon all 12 available strains from recurrent cases indicated that 9 were identical to that responsible for the initial episode [6]. We also experienced a patient with recurrence of cellulitis caused by SDSE [46]. General practitioners should be aware of the possible recurrence of invasive SDSE infection in their patients.
The distribution of emm types in patients with invasive or noninvasive infections has differed between several surveillance studies in different countries. For instance, we confirmed emm type stG643 in 2 SDSE isolates from a newborn and mother in our case of neonatal STSS [47]. The frequency of this detected emm type differed between 4 reports from different countries; prevalence was 17.2% in western Norway [48], 9.9% in the United States [5], 4.4% in Portugal [39], and 0.7% in Japan. Figure 5 compares emm typing patterns in Japan with those in the United States. Interestingly, the dominant emm types in Japan (stG6792 and stG485) were different from those in the United States (stG6, stG245, stG2078, and stG643), although the study periods differed between these studies (2002–2004 vs. 2006–2009) [3, 5]. These results illustrate the importance of conducting comparative molecular epidemiologic investigations of SDSE strains in the United States and Japan by emm type over the same study period.
Antibiotic susceptibility
Table 2 summarizes the minimum inhibitory concentration (MIC) ranges and MIC90 values of oral and parenteral antimicrobial agents for SDSE, GAS, and GBS isolates in Japan obtained from a normally sterile site [3]. Susceptibility testing was performed by dilution methods using agar plates supplemented with 5% sheep blood agar, as described previously [2].
No penicillin or cephalosporin resistance was observed in 231 SDSE strains collected by the Invasive Streptococcal Disease Working Group from 2006 to 2007.
Regarding macrolide (ML) resistance, some SDSE isolates possessed 1 of 3 resistance genes: 1.7% had the mef(A) gene; 5.6%, the erm(A) gene; and 2.6%, the erm(B) gene. Strains with the mef(A) gene showed an M phenotype involving an active efflux pump system for 14- and 15-membered MLs. Strains with the erm(A) gene showed an inducible ML/lincosamide/streptogramin B resistance phenotype, while those with the erm(B) gene showed a constitutive ML/lincosamide/streptogramin B resistance phenotype arising from methylation of 23S rRNA.
Fluoroquinolone-resistant strains first were reported in North America and Europe [49]; recently in Portugal, resistance was reported to have increased gradually to 12% [50]. The 2 resistant strains (0.9%) in our study [3] showed high resistance to levofloxacin, a fluoroquinolone agent (MIC >32 μg/ml). The strains possessed amino acid substitutions changing Ser81 to Phe or Tyr in GyrA and Ser79 to Tyr in ParC, and also the ML resistance genes erm(B) or mef(A); all showed emm type stG10. Similar results for ML- and levofloxacin-resistant strains have been reported by Sunaoshi et al. [29] for isolates collected from noninvasive and invasive infections from 2003 through 2005 in Japan.
SDSE isolates (n = 212) collected in a multicenter surveillance study by Broyles et al. [5] in the United States showed resistance rates of 28.8% for erythromycin, 4.2% for clindamycin, and 0.9% for fluoroquinolones, according to standard biologic methods [51]. In Korea [52], a high frequency of the tetracycline resistance-mediating tet(S) gene was demonstrated (68.8%), while erythromycin, clindamycin, and chloramphenicol resistance rates were low (9.4, 3.1, and 9.4%, respectively) [53].
Failure of β-lactam treatment was described in 2 patients with pharyngitis caused by SDSE [54]. In the future, continuous multicenter surveillance will be needed to elucidate the prevalence of antibiotic resistance among SDSE isolates and to establish clinical guidelines for selecting the most appropriate antimicrobial agent.
Perspectives concerning emerging infectious disease
SDSE is being identified increasingly as a pathogen responsible for invasive and noninvasive infections worldwide. Our investigation in Japan found the median patient age for invasive diseases to be 75 years [3]. Age population reported by the Japanese government in October 2008 (http://www.stat.go.jp/data/jinsui/2008np/index.htm) indicated that the general age distribution ratios of people aged >65 and >75 years were 22.1 and 10.4%, respectively. Because the elderly Japanese population is expected to increase steadily, the possibility of invasive SDSE infection should be considered by primary-care doctors treating elderly patients with fever or respiratory distress, especially in emergency departments [23, 46]. To detect SDSE isolates in elderly subjects, at least two sets of blood cultures should be obtained before the administration of antimicrobials, even when symptoms are limited.
Recently, fatal or near-fatal transfusion-transmitted infections involving platelet concentrates have been reported from some countries other than Japan [55–57]. SDSE was isolated from both the residual blood component bag and the recipient. A donor throat swab specimen collected 20 days after donation still contained SDSE [55]. These observations call for the implementation of improved safety measures for platelet concentrates. We have obtained an SDSE isolation rate of 19% in adult patients with pharyngitis in Japan (unpublished data). We, therefore, may need to determine SDSE colonization rates for skin, oropharynx, and gastrointestinal and genitourinary tracts in healthy volunteers.
Eleven isolates of SDSE from pigs slaughtered in Japan with endocarditis, arthritis, or lymphadenitis were characterized on the basis of nucleotide sequences of 16S rRNA and 23S rRNA genes [58]. Our colleagues have completed whole-genome analysis of the pig-derived SDSE L1 strain, finding high homology between this L1 strain and human SDSE strains (GGS_124 and RE378; unpublished data). Notably, many virulence-associated genes were found in SDSE strains isolated from both humans and pigs. Currently circulating SDSE strains in humans might have been formed by genetic recombination after the horizontal transfer of genes between SDSE originating in animals [59–61] and human streptococci, including GAS, GBS, or oral streptococci.
Vaccination trials
In a surveillance study in Mumbai, the SDSE carrier rate in children attending 7 public schools from 2006 to 2008 [36] was 11%, 8 times higher than the GAS carrier rate of 1.5% in the same population, suggesting that high rates of SDSE colonization in the throat make this organism an important cause of pharyngitis. In another study, in the United States, children 6 months to 18 years old who presented with pharyngitis at a children’s hospital emergency department and two outpatient offices were enrolled in a cross-sectional study [62]; controls were children who presented at the same facilities during the same period with nonrespiratory symptoms, or those who presented at the orthopedic cast clinic. Sixty-five SDSE strains (3%) were obtained from the patients with pharyngitis, while 3 (1.5%) were obtained from the smaller control group. Thus, further research and development efforts directed to the vaccination of children against SDSE infection are needed.
Trials of vaccination against GAS infection are now underway, with the M protein selected as a target antigen. The M protein, a cell surface protein, consists of 4 repeat regions (from A to D) that vary in size and amino acid composition, as well as a non-structured amino-terminal segment [63]. The sequence of C- and D-repeat regions in the carboxy-terminal domain is well conserved among the different emm types. The amino-terminal portion, which extends into host cells, consists of a non-helical, non-repeated region, as well as a hypervariable region (A-repeats) and a semivariable region (B-repeats). Vaccine-design strategies select either the hypervariable region (A-repeats) or a well-conserved region (C-repeats) to develop the vaccine. For instance, a multivalent M protein-based vaccine containing type-specific determinants from 26 different M serotypes is undergoing clinical trials [64–67]. These serotypes include types frequently isolated from invasive infections or pharyngitis (e.g., emm1, 12, 28, 3, and others) and others associated with the onset of rheumatic fever (emm5, 3, 6, 18, and others). The efficacy and safety of this 26-valent vaccine have been demonstrated in healthy adult volunteers [66]. However, the predicted coverage with this 26-valent vaccine, formulated for use in the United States, was estimated to be only 60% in Israel [67].
Another vaccine strategy involving the conserved region (C-repeats) is developing [68, 69]. Most adults possess antibodies against the p145 peptide, located within C-repeats; these antibodies act to opsonize multiple GAS strains having sequence variations in C-repeats. Because these antibodies cross-react with human cardiomyocytes, the J14, J14.1, J14-R1, and J14-R2 motifs, which exclude the cross-reactive epitope within p145, have been proposed as components of a future C-repeat vaccine [70].
SDSE also possesses an M surface protein, similar to that of GAS. Vaccination trials against SDSE infection based upon the M protein with A-repeats and C-repeats as target antigens should be initiated to prevent the development of invasive SDSE diseases, rather than to reduce the SDSE carrier rate, because the invasive illnesses may lead to poor outcomes (death or post-infective sequelae).
Conclusions
The characterization of SDSE strains was first proposed by Vandamme and colleagues in 1996 [4]. Based on whole-genome analyses of the original isolates, the genomic features of this pathogen somewhat resemble those of GAS, while differing from those of GBS. Virulence factors specific to SDSE as opposed to GAS should be determined to better understand the pathogenesis of SDSE.
In the present new century, the prevalence of invasive (e.g., STSS, necrotizing fasciitis, cellulitis, urosepsis, and pneumonia) and noninvasive SDSE infections has increased gradually year by year in Japan, other Asian countries, Europe, and America. Our investigation, the first nationwide surveillance concerning 231 invasive SDSE infections over 1 year in Japan, found the infections to be community-acquired in elderly subjects having underlying diseases. Insufficient white blood cell responses and thrombocytopenia on admission each suggested a higher risk of poor outcome (death or post-infective sequelae). Because the elderly Japanese population will continue to increase, invasive SDSE infection will be an important concern for primary-care physicians. In India, however, the disease burden caused by SDSE has been higher than that of GAS among schoolchildren. Therefore, research and development concerning vaccination against SDSE infection are also needed.
References
Wajima T, Murayama SY, Sunaoshi K, Nakayama E, Sunakawa K, Ubukata K. Distribution of emm type and antibiotic susceptibility of group A streptococci causing invasive and noninvasive disease. J Med Microbiol. 2008;57:1383–8.
Murayama SY, Seki C, Sakata H, Sunaoshi K, Nakayama E, Iwata S, et al. Capsular type and antibiotic resistance in Streptococcus agalactiae isolates from patients, ranging from newborns to the elderly, with invasive infections. Antimicrob Agents Chemother. 2009;53:2650–3.
Takahashi T, Sunaoshi K, Sunakawa K, Fujishima S, Watanabe H, Ubukata K. Clinical aspects of invasive infections with Streptococcus dysgalactiae ssp. equisimilis in Japan: differences with respect to Streptococcus pyogenes and Streptococcus agalactiae infections. Clin Microbiol Infect. doi:10.1111/j.1469-0691.2009.03047.x.
Vandamme P, Pot B, Falsen E, Kersters K, Devriese LA. Taxonomic study of Lancefield streptococcal groups C, G, and L (Streptococcus dysgalactiae) and proposal of S. dysgalactiae subsp. equisimilis subsp. nov. Int J Syst Bacteriol. 1996;46:774–81.
Broyles LN, Van Beneden C, Beall B, Facklam R, Shewmaker PL, Malpiedi P, et al. Population-based study of invasive disease due to β-hemolytic streptococci of groups other than A and B. Clin Infect Dis. 2009;48:706–12.
Liao CH, Liu LC, Huang YT, Teng LJ, Hsueh PR. Bacteremia caused by group G streptococci, Taiwan. Emerg Infect Dis. 2008;14:837–40.
Cohen-Poradosu R, Jaffe J, Lavi D, Grisariu-Greenzaid S, Nir-Paz R, Valinsky L, et al. Group G streptococcal bacteremia in Jerusalem. Emerg Infect Dis. 2004;10:1455–60.
Sylvetsky N, Raveh D, Schlesinger Y, Rudensky B, Yinnon AM. Bacteremia due to beta-hemolytic Streptococcus group G: increasing incidence and clinical characteristics of patients. Am J Med. 2002;112:622–6.
Brandt C, Spellerberg B. Streptococcus. In: Murray PR, Baron EJ, Landry ML, Jorgensen JH, Pfaller MA, editors. Manual of clinical microbiology. 9th ed. Washington: American Society for Microbiology; 2007. p. 412–29.
Lancefield RC. A serological differentiation of human and other groups of hemolytic streptococci. J Exp Med. 1933;57:571–95.
Garvie EI, Farrow JAE, Collins MD. Streptococcus dysgalactiae (Diernhofer) nom. rev. Int J Syst Bacteriol. 1983;33:404–5.
Carmeli Y, Ruoff KL. Report of cases of and taxonomic considerations for large-colony-forming Lancefield group C streptococcal bacteremia. J Clin Microbiol. 1995;33:2114–7.
Devriese LA. Streptococcal ecovars associated with different animal species: epidemiological significance of serogroups and biotypes. J Appl Bacteriol. 1991;71:478–83.
Devriese LA, Hommez J, Kilpper-Bälz R, Schleifer KH. Streptococcus canis sp. nov.: a species of group G streptococci from animals. Int J Syst Bacteriol. 1986;36:422–5.
Farrow JAE, Collins MD. Taxonomic studies on streptococci of serological groups C, G, and L and possibly related taxa. Syst Appl Microbiol. 1984;5:483–93.
Brandt CM, Haase G, Schnitzler N, Zbinden R, Lütticken R. Characterization of blood culture isolates of Streptococcus dysgalactiae subsp. equisimilis possessing Lancefield’s group A antigen. J Clin Microbiol. 1999;37:4194–7.
Katsukawa C, Tamaru A, Morikawa Y. Streptococcus dysgalactiae subsp. equisimilis possessing Lancefield’s group A antigen. Kansenshogaku Zasshi. 2002;76:155–60.
Mitsuno N, Hari T, Tamagawa N, Itoi J, Ikeda E, Hamasaki K, et al. Evaluation of rapid diagnostic kits for the detection of group A streptococcus to Streptococcus pyogenes and Streptococcus spp. with Lancefield’s group A antigen. Kansenshogaku Zasshi. 2006;80:665–73.
Tanaka D, Isobe J, Watahiki M, Nagai Y, Katsukawa C, Kawahara R, et al. Genetic features of clinical isolates of Streptococcus dysgalactiae subsp. equisimilis possessing Lancefield’s group A antigen. J Clin Microbiol. 2008;46:1526–9.
Dierksen KP, Tagg JR. Haemolysin-deficient variants of Streptococcus pyogenes and S. dysgalactiae subsp. equisimilis may be overlooked as aetiological agents of pharyngitis. J Med Microbiol. 2000;49:811–6.
Woo PC, Teng JL, Lau SK, Lum PN, Leung KW, Wong KL, et al. Analysis of a viridans group strain reveals a case of bacteremia due to lancefield group G alpha-hemolytic Streptococcus dysgalactiae subsp. equisimilis in a patient with pyomyositis and reactive arthritis. J Clin Microbiol. 2003;41:613–8.
Horii T, Izumida S, Takeuchi K, Tada T, Ishikawa J, Tsuboi K. Acute peritonitis and salpingitis associated with streptococcal toxic shock syndrome caused by Lancefield group G alpha-haemolytic Streptococcus dysgalactiae subsp. equisimilis. J Med Microbiol. 2006;55:953–6.
Takahashi T, Asami R, Tanabe K, Hirono Y, Nozawa Y, Chiba N, et al. Clinical aspects of invasive infection with Streptococcus dysgalactiae subsp. equisimilis in elderly patients. J Infect Chemother. 2010;16:68–71.
Kalia A, Bessen DE. Natural selection and evolution of streptococcal virulence genes involved in tissue-specific adaptations. J Bacteriol. 2004;186:110–21.
Humar D, Datta V, Bast DJ, Beall B, De Azavedo JC, Nizet V. Streptolysin S and necrotising infections produced by group G streptococcus. Lancet. 2002;359:124–9.
Hashikawa S, Iinuma Y, Furushita M, Ohkura T, Nada T, Torii K, et al. Characterization of group C and G streptococcal strains that cause streptococcal toxic shock syndrome. J Clin Microbiol. 2004;42:186–92.
Geyer A, Schmidt KH. Genetic organisation of the M protein region in human isolates of group C and G streptococci: two types of multigene regulator-like (mgrC) regions. Mol Gen Genet. 2000;262:965–76.
Sunaoshi K, Aburahashi H, Kobayashi R, Yamamoto Y, Okuzumi K, Yoshida A, et al. Emm typing by genetic identification of Streptococcus dysgalactiae subsp. equisimilis and susceptibility to oral antibiotics. Kansenshogaku Zasshi. 2006;80:488–95.
Sunaoshi K, Murayama SY, Adachi K, Yagoshi M, Okuzumi K, Chiba N, et al. Molecular emm genotyping and antibiotic susceptibility of Streptococcus dysgalactiae subsp. equisimilis isolated from invasive and non-invasive infections. J Med Microbiol. 2010;59:82–8.
Ikebe T, Murayama S, Saitoh K, Yamai S, Suzuki R, Isobe J, et al. Surveillance of severe invasive group-G streptococcal infections and molecular typing of the isolates in Japan. Epidemiol Infect. 2004;132:145–9.
Ahmad Y, Gertz RE Jr, Li Z, Sakota V, Broyles LN, Van Beneden C, et al. Genetic relationships deduced from emm and multilocus sequence typing of invasive Streptococcus dysgalactiae subsp. equisimilis and S. canis recovered from isolates collected in the United States. J Clin Microbiol. 2009;47:2046–54.
Misawa Y, Okugawa S, Ubukata K, Okuzumi K, Okada M, Moriya K, et al. A case of severe necrotizing cellulitis caused by group G Streptococcus dysgalactiae subsp. equisimilis. Kansenshogaku Zasshi. 2006;80:436–9.
Ueno K, Kawayama T, Edakuni N, Koga T, Aizawa H. A case of thoracic empyema with gas formation associated with Streptococcus dysgalactiae subsp. equisimilis. Kansenshogaku Zasshi. 2006;80:527–30.
Matsui D, Kitasato Y, Honda S, Ueno K, Tanaka A, Edakuni N, et al. A case of bacterial pneumonia caused by Streptococcus dysgalactiae subsp. equisimilis, showing patchy consolidations resembling organizing pneumonia. Nihon Kokyuki Gakkai Zasshi. 2007;45:36–42.
Horibe M, Sano Y, Himeno T, Ichikawa M, Ban Y, Kano H, et al. Case of gas gangrene in both legs due to Streptococcus dysgalactiae subsp. equisimilis, resulting in amputation of right leg. Nippon Naika Gakkai Zasshi. 2008;97:1879–81.
Bramhachari PV, Kaul SY, McMillan DJ, Shaila MS, Karmarkar MG, Sriprakash KS. Disease burden due to Streptococcus dysgalactiae subsp. equisimilis (group G and C streptococcus) is higher than that due to Streptococcus pyogenes among Mumbai school children. J Med Microbiol. 2010;59:220–3.
Sing A, Trebesius K, Heesemann J. Diagnosis of Streptococcus dysgalactiae subspecies equisimilis (Group C streptococci) associated with deep soft tissue infections using fluorescent in situ hybridization. Eur J Clin Microbiol Infect Dis. 2001;20:146–9.
Kumar A, Sandoe J, Kumar N. Three cases of vertebral osteomyelitis caused by Streptococcus dysgalactiae subsp. equisimilis. J Med Microbiol. 2005;54:1103–5.
Pinho MD, Melo-Cristino J, Ramirez M. Clonal relationships between invasive and noninvasive Lancefield group C and G streptococci and emm-specific differences in invasiveness. J Clin Microbiol. 2006;44:841–6.
Siljander T, Karppelin M, Vähäkuopus S, Syrjänen J, Toropainen M, Kere J, et al. Acute bacterial, nonnecrotizing cellulitis in Finland: microbiological findings. Clin Infect Dis. 2008;46:855–61.
Fernández-Martínez AI, Pascual MR, Cimas D, Esteban J. Septic arthritis due to Streptococcus dysgalactiae ssp. equisimilis. Enferm Infecc Microbiol Clin. 2008;26:670–1.
Lestin F, Mann S, Podbielski A. Spondylodiscitis and paraspinal abscess caused by beta-haemolytic group G streptococci spreading from infected leg ulcers. J Med Microbiol. 2008;57:1157–60.
Kittang BR, Langeland N, Skrede S, Mylvaganam H. Two unusual cases of severe soft tissue infection caused by Streptococcus dysgalactiae subsp. equisimilis. J Clin Microbiol. 2010;48:1484–7.
Lopardo HA, Vidal P, Sparo M, Jeric P, Centron D, Facklam RR, et al. Six-month multicenter study on invasive infections due to Streptococcus pyogenes and Streptococcus dysgalactiae subsp. equisimilis in Argentina. J Clin Microbiol. 2005;43:802–7.
Woo PC, Fung AM, Lau SK, Wong SS, Yuen KY. Group G beta-hemolytic streptococcal bacteremia characterized by 16S ribosomal RNA gene sequencing. J Clin Microbiol. 2001;39:3147–55.
Takahashi T, Yoshino M, Morozumi M, Chiba N, Kishii K, Murayama SY, Ubukata K. Clinical aspects of invasive infection with Lancefield group B and G streptococci in elderly patients. Kansenshogaku Zasshi. 2010;84:135.
Yamaoka S, Ogihara T, Yasui M, Hasegawa M, Hira S, Oue S, et al. Neonatal streptococcal toxic shock syndrome caused by Streptococcus dysgalactiae subsp. equisimilis. Pediatr Infect Dis J. 2010. doi:10.1097/INF.0b013e3181e5292f.
Kittang BR, Langeland N, Mylvaganam H. Distribution of emm types, subtypes among noninvasive group A, C and G streptococcal isolates in western Norway. APMIS. 2008;116:457–64.
Biedenbach DJ, Toleman MA, Walsh TR, Jones RN. Characterization of fluoroquinolone-resistant beta-hemolytic Streptococcus spp. isolated in North America and Europe including the first report of fluoroquinolone-resistant Streptococcus dysgalactiae subspecies equisimilis: report from the SENTRY Antimicrobial Surveillance Program (1997–2004). Diagn Microbiol Infect Dis. 2006;55:119–27.
Pinho MD, Melo-Cristino J, Ramirez M. Fluoroquinolone resistance in Streptococcus dysgalactiae subsp. equisimilis and evidence for a shared global gene pool with Streptoccocus pyogenes. Antimicrob Agents Chemother. doi:10.1128/AAC.01377-09.
Clinical and Laboratory Standards Institute (CLSI). Performance standards for antimicrobial susceptibility testing: seventeenth informational supplement. CLSI document M100-S17. Wayne, PA: CLSI; 2007.
Liu LC, Tsai JC, Hsueh PR, Tseng SP, Hung WC, Chen HJ, et al. Identification of tet(S) gene area in tetracycline-resistant Streptococcus dysgalactiae subsp. equisimilis clinical isolates. J Antimicrob Chemother. 2008;61:453–5.
Uh Y, Hwang GY, Jang IH, Cho HM, Noh SM, Kim HY, et al. Macrolide resistance trends in beta-hemolytic streptococci in a tertiary Korean hospital. Yonsei Med J. 2007;48:773–8.
Savini V, Catavitello C, Talia M, Manna A, Pompetti F, Di Bonaventura G, et al. β-Lactam failure in treatment of two group G Streptococcus dysgalactiae subsp. equisimilis pharyngitis patients. J Clin Microbiol. 2008;46:814–6.
Lessa F, Leparc GF, Benson K, Sanderson R, Van Beneden CA, Shewmaker PL, et al. Fatal group C streptococcal infection due to transfusion of a bacterially contaminated pooled platelet unit despite routine bacterial culture screening. Transfusion. 2008;48:2177–83.
Bay JO, Tournilhac O, Ducher E, Romaszko JP, Ergani A, Bouvet A, et al. A near fatal septic transfusion reaction due to Streptococcus dysgalactiae subspecies equisimilis calls for novel safety measures. Vox Sang. 2009;96:271.
Courbil R, Romaszko JP, Odent-Malaure H, Fabrigli P, Chavarin P, Tournilhac O, et al. Analysis of a severe septic transfusion reaction with standard platelet concentrate. Transfus Clin Biol. 2010;17:9–13.
Kawata K, Minakami T, Mori Y, Katsumi M, Kataoka Y, Ezawa A, et al. rDNA sequence analyses of Streptococcus dysgalactiae subsp. equisimilis isolates from pigs. Int J Syst Evol Microbiol. 2003;53:1941–6.
Laus F, Preziuso S, Spaterna A, Beribè F, Tesei B, Cuteri V. Clinical and epidemiological investigation of chronic upper respiratory diseases caused by beta-haemolytic Streptococci in horses. Comp Immunol Microbiol Infect Dis. 2007;30:247–60.
Imai D, Jang S, Miller M, Conrad PA. Characterization of beta-hemolytic streptococci isolated from southern sea otters (Enhydra lutris nereis) stranded along the California coast. Vet Microbiol. 2009;136:378–81.
Zoric M, Nilsson E, Lundeheim N, Wallgren P. Incidence of lameness and abrasions in piglets in identical farrowing pens with four different types of floor. Acta Vet Scand. 2009;51:23.
Zaoutis T, Attia M, Gross R, Klein J. The role of group C and group G streptococci in acute pharyngitis in children. Clin Microbiol Infect. 2004;10:37–40.
Smeesters PR, Mardulyn P, Vergison A, Leplae R, Van Melderen L. Genetic diversity of Group A Streptococcus M protein: implications for typing and vaccine development. Vaccine. 2008;26:5835–42.
Dale JB, Penfound T, Chiang EY, Long V, Shulman ST, Beall B. Multivalent group A streptococcal vaccine elicits bactericidal antibodies against variant M subtypes. Clin Diagn Lab Immunol. 2005;12:833–6.
Hu MC, Walls MA, Stroop SD, Reddish MA, Beall B, Dale JB. Immunogenicity of a 26-valent group A streptococcal vaccine. Infect Immun. 2002;70:2171–7.
McNeil SA, Halperin SA, Langley JM, Smith B, Warren A, Sharratt GP, et al. Safety and immunogenicity of 26-valent group a streptococcus vaccine in healthy adult volunteers. Clin Infect Dis. 2005;41:1114–22.
Nir-Paz R, Korenman Z, Ron M, Michael-Gayego A, Cohen-Poradosu R, Valinsky L, et al. Streptococcus pyogenes emm and T types within a decade, 1996–2005: implications for epidemiology and future vaccines. Epidemiol Infect. 2010;138:53–60.
Guilherme L, Faé KC, Higa F, Chaves L, Oshiro SE, Freschi de Barros S, et al. Towards a vaccine against rheumatic fever. Clin Dev Immunol. 2006;13:125–32.
Vohra H, Dey N, Gupta S, Sharma AK, Kumar R, McMillan D, et al. M protein conserved region antibodies opsonise multiple strains of Streptococcus pyogenes with sequence variations in C-repeats. Res Microbiol. 2005;156:575–82.
Yoonim N, Olive C, Pruksachatkunakorn C, Pruksakorn S. Bactericidal activity of M protein conserved region antibodies against group A streptococcal isolates from the Northern Thai population. BMC Microbiol. 2006;6:71.
Acknowledgments
Our study was funded in part by a grant under the category, ‘‘Research on Emerging and Re-emerging Infectious Diseases’’ (H19-002 and H22-013), from the Japanese Ministry of Health, Labour and Welfare (to Drs. K. Ubukata and H. Watanabe). The authors thank Katsuhiko Sunaoshi, Keisuke Okada, and Akiko Ono for assistance with manuscript preparation.
Author information
Authors and Affiliations
Corresponding author
About this article
Cite this article
Takahashi, T., Ubukata, K. & Watanabe, H. Invasive infection caused by Streptococcus dysgalactiae subsp. equisimilis: characteristics of strains and clinical features. J Infect Chemother 17, 1–10 (2011). https://doi.org/10.1007/s10156-010-0084-2
Received:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10156-010-0084-2