
Abstract
Studies suggest that high-intensity physical exercise can cause damage to skeletal muscles, resulting in muscle soreness, fatigue, inflammatory processes and cell apoptosis. The aim of this study was to investigate the effects of low-level laser therapy (LLLT) on a decrease in creatine kinase (CK) levels and cell apoptosis. Twenty male Wistar rats were randomly divided into two equal groups: group 1 (control), resistance swimming; group 2 (LLLT), resistance swimming with LLLT. They were subjected to a single application of indium gallium aluminum phosphide (InGaAlP) laser immediately following the exercise for 40 s at an output power of 100 mW, wavelength 660 nm and 133.3 J/cm2. The groups were subdivided according to sample collection time: 24 h and 48 h. CK was measured before and both 24 h and 48 h after the test. Samples of the gastrocnemius muscle were processed to determine the presence of apoptosis using terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) nick end labeling. (There was a significant difference in CK levels between groups (P < 0.0001) as well as between the 24 h and 48 h levels in the control group, whereas there was no significant intra-group difference in the LLLT group at the same evaluation times. In the LLLT group there were 66.3 ± 13.2 apoptotic cells after 24 h and 39.0 ± 6.8 apoptotic cells after 48 h. The results suggest that LLLT influences the metabolic profile of animals subjected to fatigue by lowering serum levels of CK. This demonstrates that LLLT can act as a preventive tool against cell apoptosis experienced during high-intensity physical exercise.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Muscle damage in response to strenuous exercise has been well documented in humans and in animal models. Exercise-induced muscle damage is characterized by a delayed onset of muscle soreness, muscle fiber disarrangement, muscle protein release into the plasma, acute-phase immune response and decreased muscle performance. Regional adenosine triphosphate (ATP) depletion, disruptions in calcium homeostasis, and oxygen free radicals have all been implicated in the etiology of muscle fiber damage and necrosis [1–3].
Rossini et al. state that inflammation and interstitial apoptosis occur during intense short-term exercise [4]. One explanation is that the phenomenon is due to an increased generation of reactive oxygen species during sustained hyperemia, which accompanies and follows physical muscle activity. When there is a decrease or interruption in physical activity, hyperemia is present, but the increased oxygen flow is not used in metabolic processes.
The serum creatine kinase (CK) level can be raised as a consequence of muscle tissue damage due to prolonged intensive training. This may be a consequence of both metabolic and mechanical causes. Indeed, metabolically exhausted muscle fibers exhibit decreased membrane resistance following an increase in the internal free calcium ions, which promotes the activation of the potassium channel. Another mechanism may be local tissue damage with sarcomeric degeneration from Z-disk fragmentation. CK is an indicator of muscle necrosis and increases with the extent of the necrosis [5].
Apoptosis is a process of individual cell death regulated by the activation of specific genes. Characteristic morphological features of apoptosis include nuclear and cytoplasmatic condensation, fragmentation of the cell into apoptotic bodies (which become engulfed by phagocytes) and local absence of inflammation [4, 6, 7].
Low-level laser therapy (LLLT) is a form of phototherapy that involves the application of low-power, monochromatic, coherent light to injuries and lesions. It has been used successfully to induce wound healing in cases of healing defects by inducing increases in mitotic activity, numbers of fibroblasts, collagen synthesis and neovascularization. Wounds such as burns, amputations, skin grafts and infected injuries are treated with lasers [7]. However, the literature on the effects of laser radiation on these processes is very contradictory, as a large number of variables modify its effect on tissue [8]. The mechanisms of laser radiation interaction at the molecular level are described by Karu, who suggests different action mechanisms for wavelengths emitted in the visible and infrared range [8].
Photobiological reactions involve the absorption of a specific wavelength of light by the functioning photo-acceptor molecule. The photobiological nature of the LLLT effects means that a molecule (photo-acceptor) must first absorb the light used for irradiation. After the promotion of electronically excited states, primary molecular processes from these states may lead to a measurable biological effect at the cell level [9]. Studies have indicated that local LLLT mechanisms can prevent ischemic muscle injury by reducing the release of reactive oxygen species (ROS) as well as creatine phosphokinase activity [10–12].
Serum CK is widely accepted as an indicator of muscle damage after endurance exercise, and many investigators have used this enzyme to indicate muscle injury [13, 14]. The biochemical markers of skeletal muscle damage are creatine kinase (CK), aspartate aminotransferase, lactate dehydrogenase, myoglobin and troponin. Creatine kinase is a muscle enzyme of which there is a significant rise in plasma levels following surgical procedures, trauma, myocardial infarction and strenuous exercise [5, 15].
Thus, there is a clear need to investigate whether LLLT can prevent apoptosis and muscular creatine kinase activity. Our study employed the experimental model of resistance swimming to induce fatigue in rats and to test whether LLLT could prevent the anticipated increase in CK levels, inflammation and interstitial apoptosis.
Materials and methods
Animals
Twenty adult male Wistar rats (body weight between 300 g and 350 g) were used. The animals came from the central vivarium at the University for the Development of the State and Pantanal Region (UNIDERP) and were kept in two cages (ten animals per cage) under controlled lighting and temperature with standard food and water available ad libitum.
All the experimental procedures were carried out in accordance with the norms of the Brazilian College for Animal Experimentation (COBEA). The animals were handled in compliance with the national guidelines for the humane treatment of laboratory animals. The study received approval from the Research Ethics Committee of the Universidade Federal do Mato Grosso do Sul (UFMS), under protocol number 16/2007/CEP/UFMS.
The animals were randomly divided into two experimental groups according to the procedure to be carried out: control group (n = 10), animals induced by resistance to swim until exhaustion, followed by sham LLLT irradiation; LLLT group (n = 10), animals induced by resistance to swim until exhaustion, followed by LLLT irradiation. Each of these groups was further subdivided into two groups for evaluation 24 h and 48 h after exercise (n = 5).
Swimming-induced muscle fatigue
A load-resistance swimming test was used, as described by Wu et al., with modifications [16]. The test forced the animals to swim until exhaustion. The rats were loaded with lead rings weighing 8% of their body weight, attached to their tails, and were placed in a swim tank filled with fresh water (approximately 120 cm in depth). Water temperature was maintained at 25 ± 0.5°C. We determined exhaustion by observing the loss of coordinated movements and the animal’s failure to return to the surface within 10 s. This 10 s criterion was considered to correlate with exhaustion and was used as an indication of the maximum swimming capacity of the animal. The rats were removed before they drowned.
Laser irradiation
An indium gallium aluminum phosphide (InGaAlP) diode laser (Photon Laser III DMC®, São Carlos, SP, Brazil) with a continuous output power of 100 mW and wavelength (λ) of 660 nm (visible red) was used. The optical power was calibrated with a Newport multifunction optical meter (model 1835C). The spot size was 0.03 cm2 and power density was 3.3 W/cm2.
The gastrocnemius muscle was irradiated with 133.3 J/cm2 for 40 s, by a single application immediately following the weight-loaded forced swimming. The animals were placed in the decubitus position, and the paw to be treated was placed in extension and external rotation; the laser illumination spot was placed in contact with the central prominence of the gastrocnemius muscle. Irradiated animals were given irradiation on this single point.
Biochemical parameter: CK activity
Blood samples (1.8 ml) from all experimental groups (n = 20) were collected directly into heparinized tubes by cardiac puncture 24 h and 48 h following the exercise test. Immediately after collection, the blood samples were centrifuged (2,000 rpm/25 min) for plasma separation and colorimetric determination of CK activity (EC 2.7.3.2) with a CK-NAK kit (Wienner Lab., Rosario, Argentina). The CK value was expressed in units/liter (U/l), 1 U being defined as the amount of enzyme phosphorylation need to produce 1 μmol of nicotinamide adenine dinucleotide reduced form (NADH) per minute at 25°C.
Staining by terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end labeling
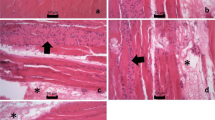
For terminal deoxynucleotidyl transferase (TdT)-mediated deoxyuridine triphosphate (dUTP) nick end labeling (TUNEL) peroxidase staining, we used the an in situ cell death kit (Roche Diagnostics) [17]. The gastrocnemius muscle was placed on a poly(L-lysine)-coated glass-bottomed dish and fixed for 30 min in 4% paraformaldehyde in phosphate-buffered saline (PBS) solution (pH 7.4). After the gastrocnemius muscle had been washed three times with PBS, endogenous peroxidase was inactivated by incubation with 3% hydrogen peroxide (H2O2) in methanol for 30 min at room temperature. The gastrocnemius muscle was then washed with PBS and subsequently permeabilized for 2 min on ice with 0.1% sodium citrate solution containing 0.1% Triton X-100. The muscle was then washed twice with PBS, stained with the TUNEL reaction mixture for 60 min at 37°C, washed twice with PBS and labeled with peroxidase-conjugated goat Ab for 30 min at 37°C. DNA fragmentation was detected by staining with diaminobenzidine. A brown color in the nucleotides was taken as the positive staining of apoptotic gastrocnemius muscle, and we quantified the nucleotides in each group by counting the output of positive cells per eight fields under light microscopy (×400).
Statistical analysis
The original data were tested with the BioEstat 5.0 software. All results are presented as mean ± standard deviation. Two-way analysis of variance (ANOVA) and Tukey’s post-hoc test were used for statistical analysis. All tests were considered to be statistically significant at P < 0.05.
Results
Creatine kinase activity
CK values obtained from animals subjected to the forced-swimming exercise until exhaustion are presented as mean ± average standard deviation. There was a significant increase (P < 0.0001) in creatine kinase release in group 1 (control) in the first 24 h (561.1 ± 38.09 U/l) when the group was compared with the group treated with LLLT (230.0 ± 24.91 U/l). Significant differences were also observed 48 h after exercise (P < 0.0001), with lower CK levels in the group subjected to LLLT. There was a significantly higher value than that in the same group 48 h following exercise (157.14 U/l). There was a very significant difference between the LLLT group and the control group after 24 h as well as after 48 h (P = 0.0002). There was no significant difference between the LLLT subgroups after 24 h and 48 h (P > 0.05) (Fig. 1).

Comparison of mean CK values ± standard deviations between groups and subgroups treated with LLLT and untreated controls 24 h and 48 h after forced swimming. **P < 0.0001 indicates very significant difference from control group after 24 h; *P < 0.0002 indicates very significant difference from control group after 48 h; ## P > 0.05 indicates non-significant difference in LLLT groups
Terminal deoxynucleotidyl transferase-mediated deoxyuridine triphosphate nick end labeling
When the processed slides were submitted for in situ analysis of DNA fragmentation, positive nuclei for TUNEL were detected in the muscle fibers in the sub-sarcolemmal and central positions, thereby confirming the presence of a majority of apoptotic nuclei at the interstitial level. In the rats subjected to exercise and treated with LLLT, there were 66.3 ± 13.2 apoptotic cells after 24 h and 39.0 ± 6.8 apoptotic cells after 48 h (P = 0.0003), In the control group (not subjected to low-level laser), the values were significantly higher after both 24 h (159.4 ± 25.8) and 48 h after the forced swimming (89.2 ± 25.4) (P = 0.0002)
Applying Tukey’s post-hoc test, we obtained the following cross-referenced results: control after 24 h versus LLLT after 24 h, P < 0.0001; control after 48 h versus LLLT after 48 h, P = 0.0002 (Fig. 2).

Comparison of mean apoptosis values ± standard deviations (obtained by TUNEL) between groups and subgroups treated with LLLT and untreated controls 24 h and 48 h after forced swimming. ** P < 0.0001 indicates very significant difference from control group after 24 h; ## P = 0.002 indicates very significant difference from control group after 48 h; *P = 0.002 indicates very significant difference from control after 48 h versus LLLT after 48 h
Discussion
Lopes-Martins et al. investigated the effects of LLLT in an experimental model of electrical nerve stimulation used to induce fatigue through tetanic tibial muscle contraction in rats and test whether LLLT could prevent the anticipated reduction in strength muscle or the anticipated increase in CK levels [12]. The authors concluded that LLLT was able to reduce the debilitating influence of local fatigue on muscle strength and possibly reduce muscle damage following strenuous exercise.
According to Fillipin et al., LLLT reduced histological abnormalities, collagen concentration and oxidative stress in an experimental model of Achilles tendon injury [18]. The reduction of fibrosis may be mediated by the beneficial effects on the oxidant/antioxidant balance.
In a study in human muscle fatigue, Leal Junior et al. reported that LLLT may delay the perceived onset of muscle fatigue and exhaustion, likely through local mechanisms, including a reduction in oxidative stress or a decrease in the production of ROS [19]. Blood lactate concentrations were similar in both groups immediately after the muscle performance test. Moreover, the accumulation of lactate may have been slightly delayed by LLLT.
The results of our study confirm the aforementioned studies reporting that LLLT is capable of reducing CK levels. However, our study allowed broader investigation by associating cell apoptosis with high-intensity physical exercise. Apoptosis is well accepted as a type of cell death during the development of mammalian muscles, but the death of adult myofibers in neuromuscular disorders and exercise-induced muscle damage is usually explained in terms of muscle necrosis [20]. Apoptosis occurs during intense exercise. One explanation is that the phenomenon is due to the increased generation of reactive oxygen species during the sustained hyperemia that accompanies and follows physical activity. When there is a decrease or interruption of physical activity, hyperemia is present, but the increased oxygen flow is not used in metabolic processes [4, 21–23]. In our study a significant reduction was found in the number of apoptotic cells in the group subjected to LLLT when compared with the control group.
Rizzi et al. found similar results [11]. The redox state of cells is a consequence of ROS, oxidants and reducing equivalents resulting from strenuous exercise. An elevation in ROS levels above the plugging capacity and enzyme activity used to modulate these levels results in potentially cytotoxic oxidative stress. Although a number of macromolecules are subjected to attack from ROS, attacks on free nucleotides or DNA can cause permanent damage due to the harmful effects of these species [24].
Our results corroborate conclusions by Carnevalli et al., who stated that LLLT promotes positive biomodulation, functioning as a prevention factor for cell apoptosis, even under conditions of nutritional stress, in Chinese Hamster Ovary (CHO) K-1 cells [8]. Our findings also corroborate those described by Shefer et al., who obtained a reduction in cell apoptosis in cultures of muscle satellite cells [25].
Gao et al. found that low-level laser irradiation induced anti-apoptosis in lung adenocarcinoma (ASTC-a-1) cells and that 0.8 J/cm2 induced cell proliferation [26]. However, Wu and Liu state that the data Gao et al. obtained were insufficient to support such statements [27]. Wu et al. found that high doses of LLLT (120 J) did not activate caspase-8, indicating that apoptosis is initiated directly from the generation of mitochondrial ROS and a decrease in DeltaPsim, regardless of caspase-8 activation [28].
In a recent study on cell apoptosis during the tissue repair process in skin injuries, Rocha Júnior et al. reported that LLLT induced an increase in apoptotic cells [29]. This finding contradicts the results of our study, but their study employed different energy densities and exposure times, which leads us to believe that there is a dose-dependent relationship in the inhibition or induction of LLLT-induced cell apoptosis.
According to Kreisler et al., in the stimulation of photoreceptors by laser light in the respiratory mitochondrial chain, changes in ATP levels are not yet fully established and remain the subject of discussion [30]. It is generally accepted that both the stimulatory and inhibitory power of laser light on cells are dose dependent and wavelength dependent.
The magnitude or efficiency of the biomodulatory effects attributed to low-level laser therapy when interacting with biological tissues is dependent on the prior metabolic conditions of the tissue, such that the response tends to be better when the cell redox potential is changed. As the response in our study was greater in the group that was subjected to physical exercise followed by LLLT, it is likely that the metabolic stress induced by the exercise favored the greater action of the laser and consequently greater effectiveness of cell stimulation.
Conclusion
The results of this study suggest that LLLT influences the metabolic profile of animals subjected to fatigue by lowering the serum CK level. This demonstrates that LLLT may act as a preventive resource for cell apoptosis unleashed by high-intensity physical exercise.
References
Stupka N, Lowther S, Chorneyko K, Bourgeois JM, Hogben C, Tarnopolsky MA, Stupka N, Lowther S, Chorneyko K, Bourgeois JM, Hogben C, Tarnopolsky MA (2000) Gender differences in muscle inflammation after eccentric exercise. J Appl Physiol 89:2325–2332
Close GL, Ashton T, McArdle A, Maclaren DP (2005) The emerging role of free radicals in delayed onset muscle soreness and contraction-induced muscle injury. Comp Biochem Physiol A Mol Integr Physiol 142:257–266
Nakatani K, Komatsu M, Kato T, Yamanaka T, Takekura H, Wagatsuma A, Aoyama K, Xu B, Hirano T, Kasai H, Ando S, Takeuchi T (2005) Habitual exercise induced resistance to oxidative stress. Free Radic Res 39:905–911
Rossini K, Donà A, Sandri M, Destro C, Donà M, Carraro U (2000) Time-course of exercise and apoptosis in dystrophin-deficient muscle of mice. Basic Appl Myol 10:33–38
Brancaccio P, Maffulli N, Limongelli FM (2007) Creatine kinase monitoring in sport medicine. Br Med Bull 81–82:209–230
Koçtürk S, Kayatekin BM, Resmi H, Açikgöz O, Kaynak C, Özer E (2008) The apoptotic response to strenuous exercise of the gastrocnemius and solues muscle fibers in rats. Eur J Appl Physiol 102:515–524
Phaneuf S, Leewenburgh C (2001) Apoptosis and exercise. Med Sci Sports Exerc 33:393–396
Carnevalli CM, Soares CP, Zângaro RA, Pinheiro AL, Silva NS (2003) Laser light prevents apoptosis in Cho K-1 cell line. J Clin Laser Med Surg 21:193–196
Karu T (1989) Photobiology of low-power laser effects. Health Phys 56:691–704
Avni D, Levkovitz S, Maltz L, Oron U (2005) Protection of skeletal muscles from ischemic injury: low-level laser therapy increases antioxidant activity. Photomed. Laser Surg. 23:273–277
Rizzi CF, Mauriz JL, Freitas-Corrêa DS, Moreira AJ, Zettler CG, Filippin LI, Marroni NP, González-Gallego J (2006) Effects of low-level laser therapy (LLLT) on the nuclear factor (NF) kappaB signaling path. Lasers Surg Med 38:704–713
Lopes-Martins RA, Marcos RL, Leonardo PS, Prianti AC Jr, Muscará MN, Aimbire F, Frigo L, Iversen VV, Bjordal JM (2006) Effect of low-level laser (Ga-Al-As 655 nm) on skeletal muscle fatigue induced by electrical stimulation in rats. J Appl Physiol 101:283–288
Gella A, Ponce J, Cussó R, Durany N (2008) Effect of the nucleotides CMP and UMP on exhaustion in exercise rats. J Physiol Biochem 64:9–17
Volfinger L, Lassourd V, Michaux JM, Braun JP, Toutain PL (1994) Kinetic evaluation of muscle damage during exercise by calculation of amount of creatine kinase released. Am J Physiol 266:R434–R441
Brancaccio P, Maffulli N, Buonauro R, Limongelli FM (2008) Serum enzyme monitoring in sports medicine. Clin Sports Med 27:1–18
Wu JL, Wu QP, Huang JM, Chen R, Cai M, Tan JB (2007) Effects of L-malate on physical stamina and activities of enzymes related to the malate-aspartate shuttle in liver of mice. Physiol Res 56:213–220
Parton M, Dowsett M, Smith I (2001) Studies of apoptosis in breast cancer. BMJ 322:1528–1532
Fillipin LI, Mauriz JL, Vedovelli K, Moreira AJ, Zettler CG, Lech O, Marroni NP, González-Gallego J (2005) Low-level laser therapy (LLLT) prevents oxidative stress and reduces fibrosis in rat traumatized Achilles tendon. Lasers Surg Med 37:293–300
Leal Junior EC, Lopes-Martins RA, Dalan F, Ferrari M, Sbabo FM, Generosi RA, Baroni BM, Penna SC, Iversen VV, Bjordal JM (2008) Effect of 655-nm low-level laser therapy on exercise-induced skeletal muscle fatigue in humans. Photomed Laser Surg 26:419–424
Carraro U (1995) Apoptotic death of dystrophic muscle fibers after exercise: a new hypothesis on the early events of muscle damage. Basic Appl Myol 5:371–374
Podhorska-Okolow M, Sandri M, Zampieri S, Brun B, Rossini K, Carraro U (1998) Apoptosis of myofibres and satellite cells: exercise-induced damage in skeletal muscle of the mouse. Neuropathol Appl Neurobiol 24:518–531
Matsuda R, Nishikawa A, Tanaka H (1995) Visualization of dystrophic muscle fibers in mdx mouse by vital staining with Evans Blue: evidence of apoptosis in dystrophin-deficient muscle. J Biochem 118:959–964
Sandri M, Carraro U (1999) Apoptosis of skeletal muscles during development and disease. Int J Biochem Cell Biol 31:1373–1390
Allen RG, Tresini M (2000) Oxidative stress and gene regulation. Free Radic Biol Med 28:463–499
Shefer G, Partridge TA, Heslop L, Gross JG, Oron U, Halevy O (2002) Low-energy laser irradiation promotes the survival and cell cycle entry of skeletal muscle satellite cells. J Cell Sci 115:1461–1469
Gao X, Chen T, Xing D, Wang F, Pei Y, Wei X (2006) Single cell analysis of PKC activation during proliferation and apoptosis induced by laser irradiation. J Cell Physiol 206:441–448
Wu M, Liu TC-Y (2007) Single-cell analysis of protein kinase C activation during anti-apoptosis and apoptosis induced by laser irradiation. Photomed Laser Surg 25:129–130
Wu S, Xing D, Wang F, Chen T, Chen WR (2007) Mechanistic study of apoptosis induced by high-fluence low-power laser irradiation using fluorescence imaging techniques. J Biomed Opt 12:064015
Rocha Júnior AM, Vieira BJ, de Andrade LC, Aarestrup FM (2009) Low-level laser therapy increases transforming growth factor-beta2 expression and induces apoptosis of epithelial cells during the tissue repair process. Photomed Laser Surg 27:303–307
Kreisler M, Christoffers AB, Willershausen B, Hoedt B (2003) Low-level 809 nm GaAlAs laser irradiation increases the proliferation rate of human laryngeal carcinoma cells in vitro. Lasers Med Sci 18:100–103
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Sussai, D.A., Carvalho, P.T.C., Dourado, D.M. et al. Low-level laser therapy attenuates creatine kinase levels and apoptosis during forced swimming in rats. Lasers Med Sci 25, 115–120 (2010). https://doi.org/10.1007/s10103-009-0697-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10103-009-0697-9