
Abstract
While disoriented humans and animals use both landmarks and environmental geometry to guide their navigation, it is not clear what kinds of cognitive mechanisms underlie these behaviors. Because traditional tests of trained navigation behavior in environments containing both landmarks and geometric information may cloud our insight into the nature of these processes, the present study tested the spontaneous use of landmarks and environmental shape by two species of fish—Redtail Splitfins (Xenotoca eiseni) and Zebrafish (Danio rerio). The results suggest that while geometry is spontaneously used by both species and both sexes to compute relative position or direction, the spontaneous use of landmarks is limited to direct beaconing and complicated by attraction to features and variability across species and sex. These findings support the view that while multiple cues may ultimately guide behavior, the computation of orientation and relative positions is specified by geometric input and is independent from other navigation processes such as beaconing.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
A highly debated idea in both human and animal navigation concerns the selectivity of environmental cues by which orientation is computed. When an animal loses its way and is disoriented, for example, instead of combining information from all available external cues to regain positions and heading, cognitive processes underlying reorientation may be more specific and limited such that the animal’s computation of direction is guided by only a subset of the environmental inputs. The debate is fueled by studies of disoriented navigation that show reliance on both environmental geometry (e.g., room shape, terrain) and features (e.g., objects, colors, 2D patterns), on one hand, and by studies that show selective uses of environmental geometry for reorientation, on the other.
Currently, there are three main views of reorientation that vary on their characterization of the nature of the processes involved in the computation of direction. The first is the geometric specificity view of reorientation. The specificity view states that while various types of cues may be used to directly mark locations, only the 3D surface layout is used to compute the relative positions and directions of these locations (Cheng and Gallistel 1984; Cheng 1986; Gallistel 1990). This view is supported by evidence of a sharp contrast in disoriented subjects’ ability to reorient by environmental layout geometry (e.g., room shape) from their lack of ability to use nongeometric features as relative positioning cues, even when those same features are successfully encoded and used as beacons or direct markers to location (Lee and Spelke 2011; Lee et al. 2012b; for review, see Lee and Spelke 2010a).
A second view of reorientation is the combination learning view. This domain-general view states that while many types of cues can be used for reorientation, the degree of influence that a particular cue has on the computation of orientation is determined by the reinforced association or experienced usefulness of each cue (Miller and Shettleworth 2007; Newcombe and Ratliff 2007). For instance, if large, stable, and salient parts of the environment (e.g., walls or salient landmarks) are previously experienced as useful cues to orientation, individuals learn to give more weight to these types of cues than others when computing direction. The combination learning view is supported by the greater influence of large, salient landmarks (compared with smaller, less salient ones) on disoriented behavior (e.g., Learmonth et al. 2002).
A third view of reorientation is panoramic image matching. The image-matching view states that when an animal loses its way, it regains heading by matching stored 2D retinal image representations of the goal to its present visual input. According to this view, there is no distinction between environmental surfaces and landmark features; whatever provides the most salient retinal images in that environment dominates the navigation behavior of the disoriented animal (Cheng 2008; Stürzl et al. 2008). The image-matching view is supported by behavioral research showing that various species of animals learn goal locations effectively using view-point-specific representations (bees: Cartwright and Collett 1982; chicks: Pecchia and Vallortigara 2010; Pecchia et al. 2011; ants: Wystrach and Beugnon 2009) and by computational models of image-matching navigation (rats: Sheynikhovich et al. 2009; ants: Wystrach et al. 2011).
While there is much evidence put forth in support of these contrasting theoretical views, making two distinctions can bring some order to the literature on the reorientation research: (1) the distinction between “disoriented navigation” and “reorientation” and (2) the distinction between spontaneous navigation behavior and reinforced, learned behavior. We describe these distinctions below and apply them in the present study to test spontaneous reorientation in two species of fish.
Disoriented navigation versus reorientation
When rats are shown food hidden in a corner of a rectangular arena and then are disoriented, they spontaneously rely only on the shape of the arena to navigate back to the food source, while ignoring other cues such as distinctive odors and local landmarks (Cheng 1986). Nevertheless, given training, the rats learn to incorporate various featural cues into their disoriented search behavior. From these experiments, Cheng (1986) hypothesized that a modular, domain-specific cognitive process allows the rats to spontaneously re-establish their heading and position with respect to the shape of the surrounding environment. In addition, nongeometric features were learned over time, through associative learning mechanisms.
Cheng’s seminal findings motivated many studies across various species and environments (for review, see Cheng and Newcombe 2005; Vallortigara 2009). Nevertheless, the nature of the mechanisms underlying disoriented navigation still remains unclear. One reason for this confusion is the lack of distinction between disoriented behavior and reorientation. Despite Cheng’s initial characterization of independent processes that guide disoriented navigation behavior, much of the literature confounds disoriented use of landmarks with reorientation by landmarks. This confound has been most clearly demonstrated and addressed in studies of reorientation in human children.
While disoriented human toddlers fail to use one uniquely colored wall to distinguish between the two geometrically identical corners of a small rectangular space (Hermer and Spelke 1994, 1996), they are much better at doing so in a large space with large landmarks (Learmonth et al. 2002, 2008) or in situations in which the salience or relevance of landmarks is highlighted (Twyman et al. 2007; Shusterman et al. 2011). While some researchers have interpreted this result as evidence that children reorient by landmarks—that children can use a landmark as a directional cue (e.g., “the toy is in the corner 30-degrees to the left of the blue wall”)—it is equally possible that children use landmarks to guide their disoriented behavior, but only as an associative beaconing cue rather than a directional cue. In other words, reorientation by geometry would narrow down their choices to the two geometrically correct corners, and children could use the landmark as an associative cue to choose the correct corner over its geometric twin. Because it is impossible to distinguish between these two possibilities in traditionally used rectangular environments with both geometric and featural cues, further research tested children in environments that isolated the landmarks from surface layout geometry.
Subsequent experiments conducted in circular or square environments that eliminate competition between landmarks and informative room shape information showed that while disoriented children use landmarks as direct markers to location, they do not use them as relative directional cues for reorientation (Gouteux and Spelke 2001; Lee et al. 2006; Wang et al. 1999), even when they were large and visually salient (Lee and Spelke 2010b, 2011). In contrast, when presented with 3D surface perturbations (i.e., bumps on the floor or walls), children successfully used the distance and direction relationships between them to distinguish between featurally identical locations, even when the surface layouts were small in size and low in visual salience (Wang et al. 1999; Lee and Spelke 2008, 2010b, 2011; Lee et al. 2012a). These studies support the geometric sensitivity view—that there is a selective sensitivity to 3D surface layouts for reorientation.
Spontaneous disoriented navigation behavior in human children consists of directional orientation by 3D surface layouts along with direct beaconing by landmark features, but do these processes also underlie disoriented navigation in nonhuman animals? While the distinction between disoriented behavior and reorientation has been explored in detail in studies of human children, it has been investigated to a much lesser degree in studies of nonhuman animals. There are indeed studies that show that disoriented animals can make use of both room shape and landmark features (for review, see Cheng and Newcombe 2005). For instance, in past studies of fish (Xenotoca eiseni) and chicks (Gallus gallus), subjects were trained to approach one corner of a rectangular arena with one blue wall or distinctively patterned corner panels and then tested following the translation or removal of the landmarks (chicks: Vallortigara et al. 1990; pigeons: Kelly et al. 1998; fish: Sovrano et al. 2003). The results showed while all species showed reliable use of environmental geometry and direct, local landmarks, the use of more distal landmarks varied. Because both landmarks and geometry were available in these and many other experiments, however, it is not clear whether these differences are caused by differences in cue competition, in learning, or in reorientation capacities. Direct tests of landmark use, therefore, must be conducted in the absence of competition with geometric information.
Trained versus spontaneous navigation
Another challenge in the comparison of navigation behavior across various species and tasks is due to the substantial differences in the methods applied in establishing a goal location. As in the studies described above, many studies of nonhuman animals use a learning paradigm, in which animals are trained over multiple sessions of reinforced learning to navigate to a goal and then are tested in altered environments. Some studies of animal navigation (e.g., Cheng 1986) and most studies of navigation in human children, however, involve a spontaneous task in which subjects are shown, without prior training, the goal at a particular location and then are allowed to approach the goal after immediate inertial disorientation. It is in these spontaneous tasks in which differences between the use of features and geometry are most robustly demonstrated.
Various studies, from controlled rearing experiments of young animals (fish: Brown et al. 2007; chicks: Chiandetti and Vallortigara 2008, 2010) to studies of human adult navigation in virtual-reality environments (Doeller and Burgess 2008), suggest that navigation by 3D surface layouts is automatic and occurs in the absence of experience-based learning, while landmark use is governed by principles of associative, reinforced learning. Similarly, Cheng’s (1986) rats, which only relied on environmental geometry before training, successfully learned to incorporate featural landmarks into their disoriented searches over reinforced trials.
Training may have several possible effects on disoriented navigation behavior. For example, reinforced learning may strengthen the association between the goal and the feature or landmark. Over training, for instance, Cheng’s rats may have learned that the goal was adjacent to the striped panel and adopted a behavioral response to the landmark. Training and experience may also led to the construction of visual snapshot-like representations of the environment. Through training, animals may store more detailed, higher resolution snapshots of the environment and use image-matching strategies to recognize goal locations.
Because these experience-dependent mechanisms may exist independently of automatic, geometrically specific orientation, it is crucial to test for these processes in disoriented animals’ spontaneous navigation behavior.
The present study
Because past studies of reorientation in fish have been limited to training tasks in environments with both geometry and landmarks, the present study implements a spontaneous task of reorientation in environments designed to independently test shape and landmark information. Two species of fish were tested: Redtail Splitfins (X. eiseni) were chosen because of their past reports of successful learning of landmark and geometry (Sovrano et al. 2002, 2003, 2005, 2007), and Zebrafish (D. rerio) were chosen because of their wide usage and importance in biological and behavioral research. Furthermore, the present study tested both males and females to investigate possible sex differences.
Three experimental spaces were tested: Experiment 1 tested the use of environmental geometry in a rectangular tank with all-white walls. Experiment 2 tested the use of a landmark in a square tank with three white walls and one blue wall. Experiment 3 tested the simultaneous use of geometry and landmark (as in previous studies involving training) in a rectangular tank with three white walls and one blue wall. Because these fish are social and therefore are motivated to navigate toward conspecifics if isolated, a social attractor was used to indicate the goal location.
General methods
Subjects
Subjects were adult Redtail Splitfins (X. eiseni) and adult Zebrafish (D. rerio). The fish were randomly chosen from a stock maintained in our laboratory within vegetation-rich (Ceratophillum sp.) tanks (55–120 L) provided with artificial illumination 16 h per day. All fish were naïve to experimental testing and were not trained on any type of task prior to testing.
Apparatus
Four small glass jars (diameter, 5 cm; depth, 6 cm) filled with water were placed at the corners of the water-filled experimental tank. The tank was lit from directly above by a single lamp. A video camera was placed above the tank and positioned so as to not cast any shadow onto the apparatus.
Design
For each subject, twelve consecutive trials were administered (all in a single session) with one unreinforced goal location held constant throughout. Approaches to the four corner jars were recorded for 30 s after the release of the disoriented fish back into the tank. The very first approach made by the fish, as well as the total frequency of approaches made during the first 30 s following its release, was analyzed. Navigation to a corner to a distance of 1 cm to a glass jar was coded as an approach.
Procedures
After its removal from the home tank, the subject was first released into the testing tank for 5 min of initial familiarization with the new environment. The fish was then captured and placed in a glass jar at the center of the arena. A female conspecific was placed into one of the corner jars; the subject, while captive in the center of the tank, was given 2 min of exposure to the conspecific in the target corner. The jar containing the subject was then covered, gently removed from the tank, and rotated slowly on a turntable, 360 degrees clockwise and 360 degrees counterclockwise. Meanwhile, the jar containing the conspecific female was replaced with an empty water-filled jar, and the whole tank was rotated 90 degrees to eliminate the possible influence of external cues. The subject was gently released into the center of the tank, and it was allowed to swim freely for 2 min, of which the first 30 s were later analyzed. After 2 min, the fish was placed back in the glass jar with the conspecific social cue in one corner for the start of the next trial.
Experiment 1
In Experiment 1, we tested the spontaneous use of surface layout geometry using a uniformly white rectangular tank (20 cm × 30 cm; depth, 10 cm). Subjects were 11 mature Redtail Splitfins (5 males and 6 females) and 11 mature Zebrafish (5 males and 6 females).
Results
First approach proportions
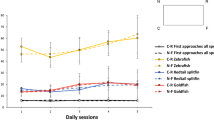
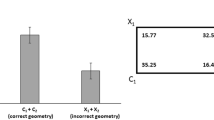
Mean proportions of searches at the Correct (C), Near Incorrect-Geometry (X1), Diagonal Correct-Geometry (D), and Far Incorrect-Geometry (X2) corners are shown in Fig. 1. A univariate ANOVA was performed with Species and Sex as between-subjects measures and Proportion of Geometrically Correct Search as the dependent measure. Because no effects or interactions of factors were found (F values <1, n.s.), the remaining analyses collapsed across both species and sex. Proportions of first approaches to the geometrically correct corners (searches at C and D, mean = 0.626) were significantly higher than a chance level of 0.5, t(21) = 5.608, P < 0.001. There were no differences between the approaches to the correct corner (C) and its geometric twin (D), t(21) = 1.284, P = 0.213, indicating that subjects were disoriented. Finally, no significant changes in geometric search was found across the twelve test trials (β = 0.085, F < 1.3, n.s.), indicating no improvement in performance with repeated exposures to the same goal location.

Mean proportions of first searches with SEM in parentheses. For Exp. 1, corners labeled Correct (C), Near Incorrect-Geometry (X 1), Diagonal Correct-Geometry (D), and Far Incorrect-Geometry (X 2). For Exp. 2, corners labeled Correct (C), Correct-Feature (X 1), Diagonal Incorrect-Feature (D), and Adjacent Incorrect-Feature (X 2). For Exp. 3, corners labeled Correct (C), Correct-Feature (X 1), Correct-Geometry (D), and Incorrect (X 2). For Exp. 2 and 3, goal locations near and far from landmark are shown separately. All data have been rotated so that shown locations for a given condition are aligned
Total approaches in 30 s
The total number of approaches in the 30 s of free exploration following disorientation confirms the above findings. A repeated measures ANOVA with Species and Sex as the between-subjects measure and Geometry of Corner Searched as within-subjects measures shows a significant main effect of geometry (F(1,18) = 9.538, P < 0.006), as well as a between-subjects main effect of Species (F(1,18) = 34.488, P < 0.001). The number of approaches to the two geometrically correct corners was significantly higher than the number of approaches to the geometrically incorrect corners (t(21) = 3.110, P = 0.005; see Fig. 2). Nevertheless, the number of approaches to the correct corner was not different from the number of approaches to its geometric twin (t(21) = 1.170, P = 0.255), indicating successful disorientation. A closer look at the between-subjects effect of species showed that the total number of approaches made during the 30 s period was higher for Zebrafish than for the Redtail Splitfins (t(20) = 5.813, P < 0.001). A complete breakdown of the first approaches and 30-s frequencies is provided in Table 1.

Total frequencies of approaches in 30 s according to geometry (C + D vs. X 1 + X 2) in Experiment 1, according to local landmark (C + X 1 vs. D + X 2) in Experiment 2, and according to both landmark and geometry in Experiment 3. For Exp. 2 and 3, goal locations near and far from landmark are shown separately
Discussion
The results of Experiment 1 show that both species and both sexes are equally good at spontaneously using geometry to reorient. These results do not accord with the slight male advantage for geometric reorientation found in Sovrano et al. (2003); one possible explanation for the difference may be due to the fact that the present study tests spontaneous navigation, while Sovrano et al. (2003) tested trained navigation. Regarding the difference in behavior between the two species, Zebrafish were generally more active than Redtail Splitfins in that they moved at faster speeds and made more approaches to the corners than the Redtail Splitfins; nevertheless, the pattern of results was identical in both species.
Experiment 2
In Experiment 2, we tested Redtail Splitfins (8 males and 8 females) and Zebrafish (8 males and 8 females) in a square tank (25 cm × 25 cm × 10 cm) with three white walls and one blue wall. Equal numbers of males and females were tested at each goal location. By eliminating the possibility of cue competition or overshadowing by environmental geometry, this experiment directly tested for the ability to use a landmark to guide disoriented navigation. If fish spontaneously reorient by the blue wall landmark or apply a panoramic image-matching process, they should search the correct goal location significantly more than the others, regardless of whether the goal is near or far from the blue wall. If fish are able to use the landmark as a local beacon, but are unable to spontaneously reorient by it (i.e., encode relative position with respect to the landmark), they should limit their searches to the corners near the landmark (but choose indiscriminately between them) and/or limit their searches to the two all-white corners when the goal corner is at one of them. Finally, if fish are unable to use the blue wall landmark to guide their disoriented search in any way, they should approach the four corners of the tank randomly.
Results
First approach proportions
Searches at the four corners were coded as Correct (C), Correct-Feature (X1), Diagonal Incorrect-Feature (D), and Adjacent Incorrect-Feature (X2) (see Fig. 1). A univariate ANOVA with Species, Sex, and Distance of Goal from Landmark as the independent measures and Proportion of Correct Search as the dependent measure revealed a significant main effect of Distance of Goal from Landmark (F(1,24) = 5.607, P = 0.026). When accuracy at goal locations near the landmark was compared with accuracy at goal locations far from the landmark, the proportion of correct searches was higher when the goal was near the landmark (t(30) = 2.230, P = 0.033); however, neither of these proportions was significantly different from a chance level of 0.25 (near landmark: t(15) = 1.616, P = 0.127; across from landmark: t(15) = 1.542, P = 0.144). Furthermore, the correct corner (C) was chosen no more often than its mirror image location (X1), both when the goal was near the landmark or across from it (t values <1, n.s.), indicating a failure to use the blue wall to reorient in both cases (Fig. 1).
To explore the use of the landmark as a local feature, the ANOVA was performed after collapsing across the corners on the same side with respect to the blue wall, for a dependent measure of Proportion of Search by Distance of Goal from Landmark (searches at C and X1), to again find a significant main effect of Distance of Goal from Landmark (F(1,24) = 26.770, P < 0.001). Comparisons to a chance level of 0.5 showed that when the goal was near the blue wall, the fish approached the blue wall corners more often than chance (mean = 0.662, t(15) = 4.691, P < 0.001). When the goal was far from the blue wall, the fish approached the white corners less often than chance (mean = 0.375, t(15) = 2.818, P = 0.013) and approached the blue wall corners instead (see Fig. 1). Finally, no differences in search accuracy was found across the twelve test trials (β = 0.014, F < 1, n.s.).
Total approaches in 30 s
The 30-s frequency measures confirm the first choice results. A repeated measures ANOVA with Species, Sex, and Distance of Goal from Landmark as the between-subjects measures and Corner Searched (C vs. X1 vs. D vs. X2) as the within-subjects measure revealed only a significant interaction between Corner Searched and Distance of Goal from Landmark (F(3,72) = 17.488, P < 0.001; Greenhouse–Geisser and Huynh–Feldt corrections applied due to a significant violation of the sphericity assumption according to Mauchly’s test). Goal locations near the landmark blue wall were approached more frequently than goal locations across from the blue wall (t(30) = 3.215, P = 0.003); nevertheless, the fish did not distinguish between the correct blue wall corner (C) and its mirror image corner (X1) (t(15) = 1.138, P = 0.273; see Fig. 1). As in the first search analysis, fish approached the two corners near the blue wall more than the all-white corners whether the actual goal location was near the blue wall (t(15) = 4.470, P < 0.001) or across from the blue wall (t(15) = 2.847, P = 0.012; see Fig. 2).
Discussion
Even when a colored landmark was the only source of information available for navigation (with no competing geometric cues), the fish did not use the landmark to reorient to the correct corner. Interestingly, the fish did not use the local features to guide remember the goal location and seemed simply to be attracted to the landmark, approaching the two blue wall corners significantly more often, regardless of the actual goal location (i.e., even when the goal was on the other side of the blue wall). This highly attractive influence of the landmark makes it difficult to address whether landmarks can be used to guide disoriented navigation. Did the fish approach the blue wall corners because they were more attracted to it than they were to the conspecific? Or, alternatively, did the fish approach the blue wall because they did not have a representation of the conspecific’s location with respect to the environmental features and therefore just approached any attractive or salient cue? It is impossible to know from the results of Experiment 2 whether the fish could not reorient by the blue wall despite its motivation to find the conspecific, or whether the fish were not motivated to find the conspecific, despite their capacity to reorient by the blue wall. The final experiment, which provided the fish with both geometric and landmark information, distinguishes between these two interpretations.
Experiment 3
The results of Experiment 1 show that the fish were clearly motivated and able to use the rectangular shape of the opaque tank to reorient to the observed location of a conspecific. Yet, the results of Experiment 2 do not distinguish the possibility that the fish were motivated to find the conspecific but could not use the landmark effectively from the possibility that the fish were not motivated to search for the conspecific in the presence of a more attractive object in the environment (the blue wall).
Redtail Splitfins (8 males and 8 females) and Zebrafish (8 males and 8 females) were tested in a rectangular tank (same as in Experiment 1) with three white walls and one blue wall. Equal numbers of males and females were tested at each goal location. If fish are indeed motivated to search for the conspecific even in the presence of an attractive landmark, they should base their approaches on the geometric properties of the goal location, choosing the geometrically correct corners over the geometrically incorrect corners, despite the fact that one of the geometrically incorrect corners is near the landmark. On the other hand, if the fish simply do not search for the conspecific in the presence of an attractive landmark, they should disregard the geometry of the observed location of the conspecific and instead approach the two blue wall corners without distinguishing between them, as they did in Experiment 2.
Results
First approach proportions
Searches at the four corners were coded as Correct (C), Correct-Feature (X1), Correct-Geometry (D), and Incorrect (X2). A univariate ANOVA with Species, Sex, and Distance of Goal from Landmark as the independent measures and Proportion of Correct Search as the dependent measure showed a significant effect of Species, F(1,25) = 6.731, P = 0.016. When analyzed separately, we found that Redtail Splitfins chose the correct corner over both its geometric twin (C vs. D, t(16) = 3.445, P = 0.003) and its mirror image corner (C vs. X1, t(16) = 5.418, P < 0.001). Zebrafish, on the other hand, showed no preference for C over X1, or C over D (both t’s < 1). No differences in search accuracy were found across the twelve test trials (β = 0.013, F < 1, n.s.).
To explore the use of geometry, the ANOVA was performed using Proportion of Geometrically Correct Search (searches at C and D) as the dependent measure, revealing no significant differences between the sexes and species on their use of geometry (F values <2, n.s.). The mean proportions of geometrically correct first search were significantly higher than chance of 0.5 for both males (searches at C and D = 0.587, t(16) = 2.589, P = 0.020,) and females (searches at C and D = 0.648, t(15) = 5.786, P < 0.001). The ANOVA using Proportion of Search by Distance of Goal from Landmark (searches at C and X1) as the dependent measure, on the other hand, showed a significant effect of Sex, F(1,25) = 4.824, P = 0.038. Males searched at C and X1 more often than chance of 0.50 (mean = 0.649, t(16) = 3.898, P = 0.001), while females searched at C and X1 equal to chance (mean = 0.517, t(15) = 0.386, P = 0.705).
Total approaches in 30 s
Looking at the total approaches during the 30-s observation period, a repeated measures ANOVA was performed with Species, Sex, and Distance of Goal from Landmark as the between-subjects measures and Corner Searched (C vs. X1 vs. D vs. X2) as the within-subjects measure. A main effect of Corner Searched was found (F(3,75) = 12.543, P < 0.001), along with significant interactions of Corner Searched × Sex (F(3,75) = 13.721, P < 0.001) and Corner Searched × Distance of Goal from Landmark (F(3,75) = 8.810, P < 0.001), and a marginally significant interaction of Corner Searched × Species (F(3,75) = 2.639, P = 0.056). Additionally, as in Experiment 1, a significant between-subjects effect of Species was found (F(1,25) = 14.620, P = 0.001), reflecting the higher overall activity (i.e., number of approaches) of the Zebrafish (mean = 65.44) compared to the Redtail Splitfins (mean = 35.12) (see Table 1).
Looking more closely at the within-subjects comparisons, fish made significantly more approaches to the Correct corner than to both the mirror image corner (t(32) = 4.648, P < 0.001) and the geometric twin corner (t(32) = 3.137, P < 0.004). When the females and males were analyzed separately, we found that the above pattern applied only to males (C vs. X1, t(16) = 3.210, P = 0.005; C vs. D, t(16) = 4.135, P = 0.001). Females approached C more than the geometrically incorrect X1 (t(15) = 3.559, P = 0.003), but did not distinguish between C and its geometric twin D (t(15) = 0.149, P = 0.884).
Further, when goal locations adjacent to the blue wall landmark were analyzed separately from goal locations at all-white corners, we found that only when the goal was at a blue wall corner did fish choose the goal significantly over its neighboring blue wall corner (C vs. X1, t(15) = 5.537, P < 0.001) and over its geometric twin (C vs. D, t(15) = 4.341, P = 0.001). When the goal was across from the landmark, the fish did weakly prefer the correct corner over its geometrically incorrect neighbor (C vs. X1, t(16) = 1.883, P = 0.078), but did not distinguish between the correct corner and its geometric twin near the blue wall (C vs. D, t(16) = 0.143, P = 0.888). Unlike Experiment 2, only when the goal was near the landmark did fish approach C and X1 more than D and X2 (t(15) = 2.812, P = 0.013); when the goal location was at an all-white corner across from landmark, the fish divided their approaches to the two geometrically correct corners and did not show an attraction for the blue wall (t(16) = 0.332, P = 0.744). In contrast to this difference in the influence of the blue wall, fish chose the two geometric corners (C and D) over the geometrically incorrect corners (X1 and X2), both when the goal was at a blue wall corner (t(15) = 3.835, P = 0.002) and when the goal was at an all-white corner (t(16) = 2.293, P = 0.036).
Finally, the marginal species difference was explored further to confirm the results found with first choice measures. Redtail Splitfins chose C over any other corner, demonstrating the combination of both geometry and local landmark features for their approach behavior (C vs. X1: t(16) = 4.621, P < 0.001, C vs. D: t(16) = 2.747, P = 0.014). While Zebrafish also demonstrated the use of geometry by choosing C and D over X1 and X2 (t(15) = 2.371, P = 0.031), they did not robustly distinguish between C and D (t(15) = 2.000, P = 0.064) and showed only a weak use of the landmark in their preference for C and X1 over D and X2 (t(15) = 2.064, P = 0.057).
To test specifically for the use of geometry, a repeated measures ANOVA was performed with Species, Sex, and Distance of Goal from Landmark as the between-subjects measures and Geometry of Corner Searched (C and D vs. X1 and X2) as the within-subjects measure. The only significant within-subjects effect revealed was that of Geometry of Corner Searched, F(1,25) = 17.632, P < 0.001, showing similarly successful use of geometry across the groups and motivation to approach the previous location of the conspecific (see Fig. 2). When the ANOVA was repeated using Distance of Corner Searched from Landmark (C and X1 vs. D and X2) as the within-subjects measure, however, all of the same effects from the ANOVA using Corner Searched (C vs. X1 vs. D vs. X2) were once again found: a main effect of Distance of Corner Searched from Landmark, F(1,25) = 10.551, P = 0.003, an interaction of Distance of Corner Searched from Landmark × Distance of Goal from Landmark, F(1,25) = 14.923, P = 0.001, an interaction of Distance of Corner Searched from Landmark × Sex, F(1,25) = 31.227, P < 0.001, and an interaction of Distance of Corner Searched from Landmark × Species, F(1,25) = 4.493, P = 0.044.
Discussion
While the use of geometry is consistent across species, sex, and landmark distance, as in past studies, the influence of the landmark on disoriented navigation behavior interacted with all three factors. First, while navigation according to geometry was equally good in both sexes, approaches based on both the local landmark and geometry was significant only in males. Females only showed the use of geometry without distinction between the two geometric twins. Nevertheless, the landmark was only helpful in guiding the male fish to the correct corner when the goal was near the attractive local landmark. Importantly, when the goal location was across from the landmark blue wall, the fish did not show an attraction to the blue wall and searched the two geometrically correct corners equally. This is a crucial and illuminating difference from the results of Experiment 2. When there was informative geometry available (Experiment 3), the fish suppressed the attraction to the blue wall (observed in Experiment 2) and searched in the geometrically correct corners, approaching the all-white goal locations as often as their geometric twin near the blue wall.
Finally, there was a weak species difference in the spontaneous use of the two available cues. While Redtail Splitfins showed successful use of both geometry and a local attractive landmark to guide disoriented behavior, Zebrafish showed a tendency to use mainly geometry and landmarks only weakly. It is not clear, however, whether this difference is due to a lower degree of attraction to the blue wall or a lower degree of landmark use overall.
General discussion
The present study provides new insight into the independent processes underlying disoriented navigation in animals. While both landmarks and environmental geometry influence spontaneous disoriented behavior, it is clear that in the absence of training, the two types of cues are used in very different ways.
First, geometry is used by both species and both sexes. This outcome is consistent with the findings of past studies (Vallortigara et al. 2009) that suggest that geometric reorientation is governed by widespread, automatic underlying mechanisms, specifically attuned to environmental surface layouts and independent of landmark use.
Second, landmarks are not used as a directional cues (or in combination with sense relations), but as an attractive local beacon, at best. When it is near the goal, it can be somewhat useful, though not as a cue to reorientation; however, when it is far from the goal, it can be a distraction, especially when there is no useful environmental geometry to otherwise inform navigation. Moreover, the influence of the blue wall landmark is variable and inconsistent across species and sex. These results are at a first glance inconsistent with the findings of Sovrano et al. (2003) in which both males and females were equally able to learn to use local landmarks to guide disoriented behavior. This difference may be due to the influence of training or motivation (given that only female conspecifics were used as the social attractors in the present task) on the use of landmarks but not geometry. Nevertheless, it is important to recognize that the present study tested only one color and type of landmark (i.e., one blue wall). It is an open question whether other landmarks or other attractor cues will result in different patterns or interactions of sex and species. Further tests are needed to address these questions.
The use of environmental shape to compute orientation in the absence of experience and training, alongside the limited use of landmarks as local beacons, is consistent with recent findings of spontaneous reorientation in other animals (Lee and Spelke 2011; Lee et al. 2012b) and suggestive of the existence of independent processes and cue specificity that characterize the geometric specificity view of reorientation. The image-matching view, for instance, cannot explain the inability to use wall color when the goal was at an all-white corner. Furthermore, the image-matching view cannot readily explain the inability to distinguish between a blue-white corner (right of the blue wall) and a white-blue corner (left of the blue wall). The combination learning view, on the other hand, cannot explain the reliability of reorientation by geometry and the contrasting failure to reorient by a stable, reliable, and salient landmark in the absence of competing cues (Experiment 2), despite the fact that the fish were motivated in the task and could overcome their attraction to the landmark when provided with geometric cues (Experiment 3). Finally, neither the image-matching view nor the combination learning theory can explain the species and sex differences in the use of the landmark and the lack of differences in the use of geometry, given that all subjects were housed in identical home aquariums.
While it is important to characterize the basic cognitive mechanisms that provide animals with abilities to navigate by geometry in the absence of training, it is also crucial to then develop an account of how learning adds new representations or changes old ones. For instance, while image-matching or domain-general learning processes do not explain spontaneous reorientation behavior, it is possible that experience and reinforced learning in familiar environments give rise to image-based or associative representations. Future studies may reveal that rather than being mutually exclusive, multiple independent processes of representing the environment, some that require more learning than others, exist in parallel with geometrically specific mechanisms for reorientation.
References
Brown AA, Spetch ML, Hurd PL (2007) Growing in circles: rearing environment alters spatial navigation in fish. Psychol Sci 18:569–573
Cartwright BA, Collett TS (1982) How honey bees use landmarks to guide their return to a food source. Nature 295:560–564
Cheng K (1986) A purely geometric module in the rat’s spatial representation. Cognition 23:149–178
Cheng K (2008) Whither geometry? Troubles of the geometric module. TICS 12:355–361
Cheng K, Gallistel CR (1984) Testing the geometric power of an animal’s spatial representation. In: Roitblat HL, Bever TG, Terrace HS (eds) Animal cognition: proceedings of the Harry Frank Guggenheim conference. Erlbaum, Hillsdale
Cheng K, Newcombe NS (2005) Is there a geometric module for spatial orientation? Squaring theory and evidence. Psychon Bull Rev 12:1–23
Chiandetti C, Vallortigara G (2008) Is there an innate geometric module? Effects of experience with angular geometric cues on spatial re-orientation based on the shape of the environment. Anim Cogn 11:139–146
Chiandetti C, Vallortigara G (2010) Experience and geometry: controlled-rearing studies with chicks. Anim Cogn 13:463–470
Doeller CF, Burgess N (2008) Distinct error-correcting and incidental learning of location relative to landmarks and boundaries. PNAS 105:5909–5914
Gallistel CR (1990) The organization of learning. MIT Press, Cambridge
Gouteux S, Spelke ES (2001) Children’s use of geometry and landmarks to reorient in an open space. Cognition 81:119–148
Hermer L, Spelke ES (1994) A geometric process for spatial reorientation in young children. Nature 370:57–59
Hermer L, Spelke ES (1996) Modularity and development: the case of spatial reorientation. Cognition 61:195–232
Kelly DM, Spetch ML, Heth CD (1998) Pigeons’ (Columba livia) encoding of geometric and featural properties of a spatial environment. J Comp Psychol 112:259–269
Learmonth AE, Nadel L, Newcombe NS (2002) Children’s use of landmarks: implication for modularity theory. Psychol Sci 13:337–341
Learmonth AE, Newcombe NS, Sheridan N, Jones M (2008) Why size counts: children’s spatial reorientation in large and small enclosures. Dev Sci 11:414–426
Lee SA, Spelke ES (2008) Children’s use of geometry for reorientation. Dev Sci 11:743–749
Lee SA, Spelke ES (2010a) Two systems of spatial representation underlying navigation. Exp Brain Res 206:179–188
Lee SA, Spelke ES (2010b) Modular geometric mechanisms for navigation in disoriented children. Cogn Psychol 61:152–176
Lee SA, Spelke ES (2011) Young children reorient by computing layout geometry, not by matching images of the environment. Psychon Bull Rev 18:192–198
Lee SA, Shusterman A, Spelke ES (2006) Reorientation and landmark-guided search by young children: evidence for two systems. Psychol Sci 17:577–582
Lee SA, Sovrano VA, Spelke ES (2012a) Navigation as a source of geometric knowledge: young children’s use of length, angle, distance, and direction in a reorientation task. Cognition 123:144–161
Lee SA, Spelke ES, Vallortigara G (2012b) Chicks, like children, spontaneously reorient by 3D environmental geometry, not by image matching. Biol Lett. doi:10.1098/rsbl.2012.0067
Miller NY, Shettleworth SJ (2007) Learning about environmental geometry: an associative model. J Exp Psychol Anim Beh Process 33:191–212
Newcombe NS, Ratliff KR (2007) Explaining the development of spatial reorientation: modularity-plus-language versus the emergence of adaptive combination. In: Plumert J, Spencer J (eds) The emerging spatial mind. Oxford University Press, New York
Pecchia T, Vallortigara G (2010) View-based strategy for reorientation by geometry. J Exp Biol 213:2987–2996
Pecchia T, Gagliardo A, Vallortigara G (2011) Stable panoramic views facilitate snap-shot like memories for spatial reorientation in homing pigeons. PLoS ONE 6:e22657
Sheynikhovich D, Chavarriaga R, Strösslin T, Arleo A, Gerstner W (2009) Is there a geometric module for spatial orientation? Insights from a rodent navigation model. Psychol Rev 116:540–566
Shusterman A, Lee SA, Spelke ES (2011) Cognitive effects of language on human navigation. Cognition 120:186–201
Sovrano VA, Bisazza A, Vallortigara G (2002) Modularity and spatial reorientation in a simple mind: Encoding of geometric and nongeometric properties of a spatial environment by fish. Cognition 85:B51–B59
Sovrano VA, Bisazza A, Vallortigara G (2003) Modularity as a fish views it: conjoining geometric and non-geometric information for spatial reorientation. J Exp Psych Anim Behav Proc 29:199–210
Sovrano VA, Bisazza A, Vallotigara G (2005) Animals’ use of landmarks and metric information to reorient: Effects of the size of the experimental space. Cognition 97:121–133
Sovrano VA, Bisazza A, Vallortigara G (2007) How fish do geometry in large and in small spaces. Anim Cogn 10:47–54
Stürzl W, Cheung A, Cheng K, Zeil J (2008) The information content of panoramic images I: the rotational errors and the similarity of views in rectangular experimental arenas. J Exp Psych Anim Behav Proc 34:1–14
Twyman A, Friedman A, Spetch ML (2007) Penetrating the geometric module: catalyzing children’s use of landmarks. Dev Psychol 43:1523–1530
Vallortigara G (2009) Animals as natural geometers. In: Tommasi L, Peterson M, Nadel L (eds) The biology of cognition. MIT Press, Cambridge
Vallortigara G, Zanforlin M, Pasti G (1990) Geometric modules in animals’ spatial representations: a test with chicks (Gallus gallus domesticus). J Comp Psychol 104:248–254
Vallortigara G, Sovrano VA, Chiandetti C (2009) Doing Socrates experiment right: controlled-rearing studies of geometrical knowledge in animals. Curr Opin Neurobiol 19:20–26
Wang RF, Hermer L, Spelke ES (1999) Mechanisms of reorientation and object localization by children: a comparison with rats. Behav Neurosci 113:475–485
Wystrach A, Beugnon G (2009) Ants learn geometry and features. Curr Biol 19:61–66
Wystrach A, Cheng K, Sosa S, Beugnon G (2011) Geometry, features, and panoramic views: ants in rectangular arenas. J Exp Psychol Anim Behav Process 37:420–435
Acknowledgments
These experiments comply with the Italian and the European Community law on research with animal. Funding was provided by the Provincia Autonoma di Trento and the Fondazione Cassa di Risparmio di Trento e Rovereto; the authors declare that they have no conflict of interest. We would like to thank Francesco Cerri for the construction of experimental stimuli and maintenance of the aquariums.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Lee, S.A., Vallortigara, G., Ruga, V. et al. Independent effects of geometry and landmark in a spontaneous reorientation task: a study of two species of fish. Anim Cogn 15, 861–870 (2012). https://doi.org/10.1007/s10071-012-0512-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10071-012-0512-z