
Abstract
Since the thermoacidophilic raw-starch hydrolyzing α-amylase Gt-amy can effectively hydrolyze corn starch under starch liquefaction conditions, it has potential for many industrial applications. To identify the raw starch-binding domain of Gt-amy, a C-terminal domain (CTD)-truncated mutant (Gt-amy-T) was constructed, and its enzymatic properties were compared with Gt-amy. In comparison to CTD of Gt-amy, which could effectively bind corn starch, the Gt-amy-T could not bind to and hydrolyze corn starch under similar conditions. In addition, Gt-amy-T showed significantly lower thermal activity and thermal stability. Using soluble starch as the substrate, the kcat of Gt-amy-T at 80 °C was approximately 77.9% of that of Gt-amy. The half-life of Gt-amy at 80 °C was 3 h, while that of Gt-amy-T was 2 h. These results reveal that the CTD plays a vital role in raw starch binding and degradation by Gt-amy and helps Gt-amy maintain thermal activity and stability.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Natural raw starch is found in granular form with a complex and dense structure that needs to be destroyed with a strong acid, strong alkali, high temperature, or enzymatic treatment before application (Ai and Jane, 2015; Robertson et al., 2006). Enzymatic degradation of raw starch can simplify the pretreatment process in the modern fermentation industry, which is more environmentally friendly and energy-saving. Therefore, research on raw starch-degrading enzymes has received great attention. Raw starch-degrading amylase is an enzyme that exhibits strong hydrolytic activity on raw starch granules that are not gelatinized (Božić et al., 2017; Cinelli et al., 2015; Sun et al. 2009, 2010). Some enzymes in the α-amylase, β-amylase, glucoamylase, and isoamylase families can hydrolyze raw starch. Raw starch-degrading α-amylase can directly act on raw starch granules at a temperature lower than the gelatinization temperature, so that the raw starch-gelatinization procedure can be omitted during starch liquefaction, which is beneficial in terms of energy savings and simplifying the process. Therefore, raw starch α-amylase has great application potential in fields such as brewing, food, paper making, and textiles (Božić et al., 2017; Cinelli et al., 2015; Homaei et al., 2016; Janecek et al., 2014; Rana et al., 2013; Sun et al., 2009; 2010; Zhang et al., 2017).
Most raw amylases possess a starch-binding domain (SBD) that binds to the surface of starch granules, which is key for their ability to directly hydrolyze raw starch granules without gelatinization (Christiansen et al., 2009; Machovic and Janecek, 2006; Majzlova and Janecek, 2014). The SBD belongs to the carbohydrate-binding modules (CBMs). As a natural component of the raw starch amylase, the SBD has the primary effects of enabling amylase molecules to bind insoluble substrates (starch granules) in solution, transporting the substrate to the active site of the catalytic domain, and rupturing the surface of the starch granules. The SBD domain of the microbial-derived raw starch α-amylase is generally located at its C-terminal domain (CTD) (Machovic and Janecek, 2006).
The thermoacidophilic raw starch-hydrolyzing α-amylase Gt-amy derived from the thermophilic bacteria Geobacillus thermoleovorans is a Ca2+-independent raw starch-degrading α-amylase with excellent thermal activity and thermal stability, and can effectively hydrolyze corn starch under starch-liquefaction conditions (30% corn starch, 3-h reaction time, pH of 5.0) (de Souza and de Oliveira Magalhães, 2010; Mehta and Satyanarayana, 2013). Therefore, Gt-amy has excellent potential for application in starch-liquefaction processes. The simulated molecular structure of Gt-amy shows that it consists of three domains, namely domains A, B, and C (Mehta and Satyanarayana, 2013). The conserved domain of Gt-amy was analyzed using the Conserved Domain Database on the National Center for Biotechnology Information (NCBI) website. The results showed that the function of the CTD (Lys470–Phe549) in domain C is unknown (DUF1939, CDD286264) (Marchler-Bauer et al., 2011). The CTD is composed of a β-sheet structure, which is consistent with the structural characteristics of type-B CBMs (Armenta et al., 2017). However, the amino acid sequence of the CTD shares no sequence similarities with other known CBMs (such as CBM20, CBM25, CBM26, CBM41, CBM48, and CBM69), so its function cannot be inferred by sequence analysis. In this study, the function of the CTD in Gt-amy was determined through deletion analysis. The results showed that the CTD mediates Gt-amy adsorption to raw starch, is essential for raw starch degradation by Gt-amy and affects the thermal activity and thermal stability of Gt-amy.
Materials and methods
Bioinformatic analysis
A homology model of Gt-amy (residues 35–516) was created with Swiss-Model (https://swissmodel.expasy.org/interactive) using the 1hvxA template (Waterhouse et al., 2018). The structure file in pdb format was viewed using PyMOL. Conserved domain searches were performed using the NCBI Conserved Domain Database (https://www.ncbi.nlm.nih.gov/cdd) (Marchler-Bauer et al., 2011). SBD sequences were selected from the Carbohydrate-Active Enzymes Database (http://www.cazy.org/Carbohydrate-Binding-Modules.html) (Lombard et al., 2014). All sequence alignments were performed using the CLUSTAL X program, and phylogenetic analysis was performed with MEGA software, version 5.2 (Tamura et al., 2011).
Strains and growth conditions
Escherichia coli JM109 and Bacillus subtilis WB600 were used as hosts for cloning and protein expression. Bacteria were grown at 37 °C in Luria–Bertani (LB) medium containing kanamycin (30 μg/ml), as needed.
Plasmid construction
The sequence of the gtamy gene (GenBank ID JQ409473) encoding Gt-amy was obtained from NCBI’s Entrez Nucleotide database (https://www.ncbi.nlm.nih.gov/nucleotide). The gene gtamyh (α-amylase Gt-amy with a 6×His tail) was synthesized by Shanghai BioYare Co., Ltd (Shanghai, China). The structural gene gtamyhds (containing no signal peptide) was obtained through polymerase chain reaction (PCR)-based amplification using gtamyh as the template and P1 and P2 as the primers (P1: 5′-CGGAGATCTGCCGCACCGTTTAACGGCAC-3′; P2: 5′-CGCGCATGCTCAGTGGTGGTGGTGGTGGTGTCAAGGCCATG-3′). The amplified product was digested with BglII and SphI and ligated into the pSTOP1622 vector to construct the recombinant plasmid, pSTOP1622-gtamyhds. The ligation product was transformed into E. coli JM109 competent cells. Positive clones were confirmed by DNA sequencing. Then, the pSTOP1622-gtamyhds plasmid was used as a PCR template. A DNA fragment corresponding to the N-terminal 396 amino acids of Gt-amy, named Gt-amy-T, was generated using the primers P1 and P3 (P3: 5′-CGCGCATGCTCAGTGGTGGTGGTGGTGGTGGCTTCCTCCCG-3′). The CTD (80 amino acid residues) was amplified using primers P2 and P4 (P4: 5′-CGGAGATCTAAATGGATGTACGTTGGCAAAC-3′). The resulting PCR products were digested with BglII and SphI and cloned into the pSTOP1622 vector to generate the recombinant plasmids, pSTOP1622-gtamy-Th and pSTOP1622-ctdh.
Gene expression and purification
The recombinant plasmid was transformed into B. subtilis WB600. The preparation and transformation of B. subtilis WB600 competent cells were performed with the modified Spizizen method (Anagnostopoulos and Spizizen, 1961). Clones of recombinant B. subtilis were inoculated into liquid LB medium and cultured in a 250-ml Erlenmeyer flask (medium volume: 20 ml) at 37 °C and 200 rotations per min (rpm) for 10 h. Then, the culture was transferred to fresh liquid LB medium and cultured in a 250-ml Erlenmeyer flask (medium volume, 25 ml; inoculum size, 3%) at 37 °C and 200 rpm. When the bacteria OD600nm reached 1, xylose was added at a final concentration of 0.5% to induce expression of the recombinant protein, with an induction duration of 30 h. Cells were harvested and lysed by ultrasonication. After centrifugation at 13,000×g for 10 min, the supernatant containing the crude protein extract was collected. For purification of the recombinant α-amylase, the supernatant was subjected to affinity chromatography on a HiTrap chelating HP column (GE Healthcare BioScience, Boston, USA), equilibrated with 50 mM 2-morpholinoethanesulfonic acid (MES) buffer at pH 5.0. After washing the column with 50 mM MES buffer containing 40 mM imidazole, the bound protein was eluted with 50 mM MES buffer containing 250 mM imidazole. The eluted sample was then dialyzed against 50 mM MES buffer at pH 5.0, to remove the imidazole. The purity of recombinant α-amylase was detected by sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE), and the concentration of recombinant α-amylase was determined by the Bradford method (Michael and Joseph, 2014).
α-Amylase assay
Ten microliters of enzyme solution were mixed with 490 μl of 50 mM MES containing 1% (w/v) soluble starch and a buffer solution (pH 5.0). The mixture was reacted at 80 °C for 30 min and afterward was quickly put into an ice water bath to terminate the reaction. Then, the 3,5-dinitrosalicylic acid (DNS) method was applied to measure the reducing sugar content in the reaction system (Miller, 1959). An enzyme-activity unit (U) was defined as follows: one unit of enzyme activity (U) refers to the amount of enzyme that catalyzes the production of 1 μmol reducing sugar per min.
Raw corn starch adsorption
The ability of proteins to adsorb to insoluble raw corn starch was determined. Raw corn starch was prewashed three times with Milli-Q water. Fifty microliters of Gt-amy (0.6 μmol), Gt-amy-T (0.6 μmol), or CTD (0.6 μmol) was added to 0.5 ml of 50 mM MES buffer (pH 5.0) containing 0, 25, 50, 75, 100, 125, or 150 mg of raw corn starch. The reactions were incubated at 20 °C for 3 h under gentle shaking and then centrifuged at 12,000×g for 5 min to pellet the starch and bound protein. The residual unbound protein was determined using the Bradford method with bovine serum albumin as a standard. The percent adsorption was calculated using the following formula: % adsorption = (original protein − unbound protein)/original protein × 100.
Applicability of the recombinant α-amylases in raw corn starch hydrolysis
Thirty percent raw corn starch slurry prepared in 50 mM MES buffer (pH 5.0) was mixed with 0.1 U of recombinant α-amylases at 60 °C for 3 h. Aliquots were withdrawn at the indicated intervals, and reducing sugars in the supernatants were determined using the DNS reagent. The extent of raw corn starch hydrolysis was calculated using the formula: % starch saccharification = [reducing sugars (mg/ml)/initial weight of starch (mg/ml)] × 0.9 × 100.
Scanning electron microscopy
Thirty percent raw corn starch slurry prepared in 50 mM MES buffer (pH 5.0) was mixed with or without 0.1 U of recombinant α-amylases at 60 °C for 3 h. The mixture was centrifuged at 8000×g, and the pellet was washed three times with pure ethanol, followed by drying at 30 °C. Starch granules were attached to a scanning electron microscopy (SEM) holder and then coated for 40 s with platinum using Ion Sputter E-1010 (Hitachi, Tokyo, Japan) at 5.0 kV and 20 mA. Samples were inserted into the SEM specimen chamber (VEGA3-TESCAN, Brno, Czech), and pictures were taken.
Biochemical characterization of the recombinant α-amylases
The optimum temperature for the enzyme activity was determined by incubating the reaction mixture (pH 5.0) at temperatures ranging from 30 to 100 °C. The optimum pH for the enzyme activity was determined by incubating the reaction mixture at 80 °C in the presence of buffers of varying pH [50 mM MES buffer (pH 3.0–7.0), 50 mM 3-morpholinopropanesulfonic acid (MOPS) buffer (pH 7.0–11.0)]. The thermal stability of the recombinant α-amylase was determined by incubating the enzyme at 80 °C, after which the assay was performed at the indicated time intervals.
Enzyme kinetics
Soluble starch solutions of varying concentrations (0.1–5.0 mg/mL) and diluted enzyme (1.4 × 10−7 M) were prepared in 50 mM MES buffer (pH 5.0). Reaction mixtures containing substrate and enzyme were used to determine enzyme activities at 80 °C and the Km and kcat values. For kinetic analysis of the recombinant α-amylase, the initial rates of the pure enzyme on soluble starch were calculated following the reducing sugar concentration, using the DNS method at different initial starch concentrations (0.1–5.0 mg/mL). The Km and kcat values were obtained using Lineweaver–Burk plots.
Results and discussion
Classification and domain analysis
Raw starch-degrading α-amylases can effectively hydrolyze raw starch, and their molecular structures usually contain a raw SBD. The SBDs of raw starch-degrading α-amylases derived from microorganisms are mostly located in the CTD (Božić et al., 2017). The raw starch-degrading α-amylase Gt-amy used in this study can effectively hydrolyze wheat starch and corn starch (Mehta and Satyanarayana, 2013). The presence of a putative C-terminal SBD in Gt-amy was analyzed by searching in the NCBI Conserved Domain Database (Marchler-Bauer et al., 2011). Conserved domain search results revealed no potential SBD in Gt-amy and that its CTD (Lys470–Phe549) comprises a domain of unknown function (DUF1939, CDD286264). We attempted to determine the role of the Gt-amy CTD in starch adsorption based on structure prediction and phylogenetic analysis.
The molecular structure of Gt-amy was simulated using Swiss-Model with the molecular protein structure of the α-amylase BStA (PDB ID: 1hvxA) derived from B. stearothermophilus as the template. As shown in Fig. 1A, the tertiary structure of Gt-amy consists of three domains, namely domain A (residues 35–139 and 241–430), domain B (residues 140–240), and domain C (residues 431–549). The CTD (residues 470–549) in domain C is composed of a β-sheet structure, and the structural features of the CTD are consistent with those of type-B glycan chain-binding CBMs. Type-B glycan chain-binding CBMs are also composed of a β-sandwich or a β-sheet structure. Phylogenetic analysis of the Gt-amy CTD with the known SBDs and domain C of the known α-amylase revealed that the CTD is closely related to domain C of the known α-amylase, rather than CBM20, CBM25, CBM26, CBM41, CBM48, CBM69 families (Fig. 1B). Furthermore, aligning the CTD sequence with known SBDs from the CBM20, CBM25, CBM26, CBM41, CBM48, and CBM69 families revealed that the CTD shares no significant sequence similarity with the known SBDs, and the starch-binding sites in the known SBDs are not present in the CTD (results not shown). These results suggest that the Gt-amy CTD is not a typical SBD and is, in fact, a domain of unknown function. Therefore, a more in-depth study is needed to determine its function and elucidate any possible relation of the Gt-amy CTD with raw starch binding.

Domain analysis of Gt-amy. (A) Tertiary structure display of Gt-amy. Domains A, B, and C are marked in red, green, and yellow, respectively. (B) Phylogenetic analysis of the evolutionary relationship between the CTD of Gt-amy and CBM20, CBM25, CBM26, CBM41, CBM69, and domain C of α-amylases
Construction and purification of truncated derivatives
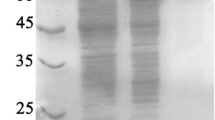
To explore the function of the Gt-amy CTD, we used recombinant DNA techniques to generate the C-terminal-truncated Gt-amy-T and the CTD (Fig. 2A). During the expression in B. subtilis WB600, the target proteins Gt-amy, Gt-amy-T, and CTD were successfully expressed and mainly localized in the soluble compartments of cells. The target proteins were purified to near homogeneity by Ni2+ affinity chromatography. The apparent molecular weights of Gt-amy, Gt-amy-T, and CTD were 56, 47, and 9 kDa (Fig. 2B), respectively, consistent with their theoretical molecular weights.

(A) Schematic representation of Gt-amy. Amino acid numbers corresponding to the module boundaries are shown above the schematic, and the individual module constructs are also shown (B) SDS-PAGE analysis of the purified recombinant proteins. Lane Mr, marker (kDa); lane 1, full-length Gt-amy; lane 2, truncated Gt-amy-T; lane 3, CTD
Adsorption rate and degradation rate of corn starch
To study the role of the Gt-amy-CTD as a raw SBD, the adsorptions of Gt-amy, Gt-amy-T, and the CTD to raw corn starch were assayed at various starch concentrations. The amount of bound protein, expressed as a fraction of the total protein, is plotted in Fig. 3A versus the concentration (%) of raw corn starch. Gt-amy and the CTD displayed similar affinities for raw corn starch, while Gt-amy-T did not adsorb to raw corn starch. Maximum adsorption of both enzymes to raw corn starch was observed at 30% raw corn starch. Gt-amy and the CTD showed 78.8% and 84.5% adsorption to raw corn starch, respectively. These results suggested the role of the CTD in raw starch adsorption, and Gt-amy binding to raw starch is very much dependent on the CTD. The phenomena that the CTD alone showed the highest affinity for raw corn starch may be due to the more facile interactions between a small protein and a large starch granule, compared to Gt-amy. Similar phenomena were also observed with other raw starch α-amylases. For example, the raw starch α-amylase AmyP and its SBD showed similar relative affinities to raw rice starch, and its SBD-deleted mutant AmyP∆SBD did not bind raw rice starch. The adsorption rate of AmyP∆SBD to rice starch was 73%, and that of AmyP was 53% (Peng et al., 2014).

Effect of the CTD on raw corn starch adsorption and hydrolysis. (A) Protein binding to raw corn starch. Fraction of protein bound versus the number of mg of starch. (B) Extent of raw corn starch hydrolysis by Gt-amy and Gt-amy-T. Digestion of 30% raw corn starch by Gt-amy and Gt-amy-T, as measured by the release of reducing sugars. The incubations were carried out at pH 6.5 and 60 °C
The degradation rates of 30% corn starch by Gt-amy and Gt-amy-T were compared to investigate the effect of the CTD on corn starch degradation by Gt-amy. The extent of raw corn starch hydrolysis was expressed as the percent of starch saccharification (Fig. 3B). Considering that the gelatinization starting temperature of corn starch is 64 °C (Robertson et al., 2006), the reaction was performed at 60 °C. As shown in Fig. 3B, the degradation rates of corn starch by Gt-amy and Gt-amy-T increased over time. As the reaction time increased to 3 h, Gt-amy hydrolyzed 65.8% of the raw corn starch, while Gt-amy-T failed to hydrolyze raw corn starch. The starch-saccharification rate of Gt-amy-T was only 2.8% when the reaction was stopped, and the scanning electron micrographs also revealed that Gt-amy hydrolyzed raw corn starch efficiently while Gt-amy-T showed no activity toward raw corn starch (Fig. 4). These results indicated that the CTD plays a functional role in raw corn starch hydrolysis. For amylases containing SBD or starch-binding regions, deletion of this domain would affect the raw starch-binding rate of the amylase. However, deletion of this region had various effects on the raw starch-hydrolysis rate of different amylases. For instance, for the α-amylase derived from Lactobacillus amylovorus (Rodriguez Sanoja et al., 2000) and the α-amylase Gt-amyII derived from G. thermoleovorans (Mehta and Satyanarayana, 2014), the SBD-deletion and domain C-deletion mutants completely lost the ability to hydrolyze raw starch. For the α-amylase SusG, derived from Bacteroides thetaiotaomicron, the specific enzyme activity of its SBD-deleted mutant against raw starch was approximately 30% that of SusG (Koropatkin and Smith, 2010). However, for the α-amylase derived from Bacillus sp. TS-23 (Lo et al., 2002) and the starch pullulanase derived from Thermoanaerobacter ethanolicus 39E (Lin et al., 2008), SBD deletion did not affect raw starch hydrolysis. These findings indicate that for some amylases, the raw SBD is not required for raw starch hydrolysis. Moreover, some special raw amylases do not contain raw SBDs, but can hydrolyze raw starch, probably because of the presence of raw starch-binding sites in their molecular structures (Baroroh et al., 2017; Peng et al., 2015).

SEM images of raw corn starch before and after enzymatic hydrolysis at 60 °C for 3 h. (A1, A2) Untreated raw corn starch. (B1, B2) Gt-amy-treated raw corn starch. (C1, C2) Gt-amy-T-treated raw corn starch
The results above indicate that Gt-amy can effectively bind to and hydrolyze corn starch, while its C-terminal domain-deleted mutant Gt-amy-T cannot. For the first time, the role of the Gt-amy CTD has been experimentally verified to be involved in raw starch binding and hydrolysis, although the CTD is not a typical SBD, as mentioned above. Data from previous studies have shown that domain C of raw starch-degrading α-amylase is involved in binding to raw starch, such as domain C of barley α-amylase (Tibbot et al., 2002) and domain C of Gt-amyII (Mehta and Satyanarayana, 2014). In addition, the α-amylase BLA derived from Bacillus licheniformis has been confirmed to be effective in corn starch hydrolysis (Sokarda Slavic et al., 2016; Wu et al., 2018), and its domain C is closely related to the Gt-amy CTD (Fig. 1B), but whether the domain C of BLA also binds to raw starch remains to be confirmed.
Biochemical characterization of the recombinant Gt-amy and Gt-amy-T
The thermal activity of Gt-amy and its CTD-deleted mutant Gt-amy-T, the optimum reaction pH, and the thermal stability were further compared to determine the effect of the Gt-amy CTD on the other enzymatic properties of Gt-amy. With 1% soluble starch as the substrate, the specific enzyme activity of recombinant α-amylase was determined under different temperature conditions (30–100 °C), as shown in Fig. 5A. The optimal reaction temperature for both recombinant α-amylase Gt-amy and Gt-amy-T was 80 °C, but the specific activity of Gt-amy was significantly higher than that of mutant Gt-amy-T at 40–100 °C. Among them, the specific activity of Gt-amy was 1756.7 U/mg at 80 °C, whereas that of Gt-amy-T was 1424.4 U/mg. Determination of the kinetic parameters of Gt-amy and Gt-amy-T at 80 °C revealed that Gt-amy had a kcat/Km value 1.2-fold higher than that of Gt-amy-T, mainly due to a higher hydrolysis rate constant (kcat) (Table 1). These results demonstrate that the Gt-amy CTD increased the catalytic efficiency of Gt-amy. The SBD of α-amylase AmyP had a significant effect on its catalytic activity towards soluble starch, and the specific activity of the SBD-deleted mutant AmyP∆SBD towards soluble starch decreased by 80% (Peng et al., 2014). Moreover, previous studies showed that SBD deletion did not affect soluble starch hydrolysis by α-amylase derived from Bacillus sp. TS-23 (Lo et al., 2002) and starch pullulanase derived from T. ethanolicus 39E (Lin et al., 2008). However, SBD deletion enhanced soluble starch hydrolysis by certain amylases, such as α-amylase derived from B. thetaiotaomicron (Koropatkin and Smith, 2010) and α-amylase derived from Bacillus sp. 195 (Sumitani et al., 2000). A potential explanation of the different effects of the SBD on soluble amylase activity is that SBD deletion leads to differences in the molecular structures of each amylase involved.

Biochemical characterization of Gt-amy and Gt-amy-T. (A) Influence of temperature on α-amylase activity. Activity assays were performed for 30 min using 1% soluble starch as the substrate at pH 6.5 over a temperature range of 30 to 100 °C. (B) Influence of pH on α-amylase activity. Activity assays were performed at the indicated pH for 30 min using 1% soluble starch as the substrate. (C) Heat inactivation of the enzymes. The enzymes were incubated at 80 °C for the indicated time intervals and then subjected to α-amylase activity assays. The relative residual activities were calculated by defining the non-heat-treated sample as 100%. (D) Influence of Ca2+ and EDTA on α-amylase activity. Enzyme activity was measured at 80 °C for 30 min using 1% soluble starch as the substrate with different concentrations of Ca2+ or EDTA. (E) Influence of Ca2+ and EDTA on α-amylase stability. The half-lives were measured at 80 °C with different concentrations of Ca2+ or EDTA. G and GT represent Gt-amy and Gt-amy-T, respectively. G[Ca ~ (2 +) 1] and GT[Ca ~ (2 +) 1] represent Gt-amy and Gt-amy-T with 1 mM Ca2+, respectively. G[Ca ~ (2 +) 5] and GT[Ca ~ (2 +) 5] represent Gt-amy and Gt-amy-T with 5 mM Ca2+, respectively. G[EDTA 5] and GT[EDTA 5] represent Gt-amy and Gt-amy-T with 5 mM EDTA, respectively. G[EDTA 10] and GT[EDTA 10] represent Gt-amy and Gt-amy-T with 10 mM EDTA, respectively. The values are expressed as means ± standard deviations (bars) of the results of three independent experiments
The optimum reaction pH for both the recombinant α-amylases Gt-amy and Gt-amy-T was approximately 5.0, and they had similar relative enzyme activities over a pH range of 3.0 to 11.0 (Fig. 5B). The thermal stability of the recombinant α-amylase at 80 °C is shown in Fig. 5C. The half-life of Gt-amy at 80 °C was approximately 3 h, and that of Gt-amy-T at 80 °C was approximately 2 h. The investigation on α-amylase Gt-amyII also showed that the C-terminal SBD contributes to its thermostability (Mehta and Satyanarayana, 2014), although the associated mechanism remains unknown.
The simulated molecular structure of Gt-amy showed that there is a Ca2+-binding site adjacent to CTD. As mentioned before, Gt-amy-T displayed lower thermal activity and stability as compared to Gt-amy. To further investigate whether CTD or the Ca2+ binding is a critical factor for the thermal activity and stability of Gt-amy, the activity and stability of Gt-amy and Gt-amy-T were measured with or without Ca2+. As shown in Fig. 5D, E, Gt-amy displayed decreased thermal activity and stability in the presence of Ca2+. These results were consistent with those of a previous study in which the thermal activity and stability of Gt-amy were reportedly inhibited by supplemental Ca2+ (Mehta and Satyanarayana, 2013). A similar influence of Ca2+ on the thermal activity and stability was also observed for Gt-amy-T. Moreover, the thermal activity and stability of Gt-amy and Gt-amy-T seemingly decreased in the presence of EDTA. This result indicates that the binding of Ca2+ is essential for the thermal activity and stability of Gt-amy and Gt-amy-T. Altogether, the above results show that the Ca2+ influences both the thermal activity and stability of Gt-amy and Gt-amy-T in a similar way. Besides, Gt-amy displayed higher thermal activity and stability than Gt-amy-T in the presence of EDTA. One reasonable explanation for this observation is that CTD truncation may disrupt the compact structure of the enzyme, thus decreasing the thermal activity and stability of Gt-amy. In conclusion, CTD is important for the thermal activity and stability of Gt-amy, although CTD is not the leading factor that affects its thermal activity and stability.
In summary, deletion mutagenesis performed in this study demonstrated the function of the C-terminal domain in Gt-amy. Gt-amy can bind to raw starch through the CTD, the CTD-deleted mutant Gt-amy-T cannot bind to raw starch, and the CTD can bind to raw starch. The CTD affected the raw starch-hydrolysis activity of Gt-amy, and Gt-amy-T lost the ability of raw corn starch hydrolysis. The CTD helped Gt-amy maintain thermal activity and thermal stability. The Gt-amy CTD is not a typical SBD but is capable of raw starch binding and influences the enzymatic properties of Gt-amy. This study not only expands our understanding of the structure and function of the raw SBD, but also provides a new theoretical basis and design ideas for constructing α-amylase mutants with enhanced performance.
References
Ai Y, Jane J. Gelatinization and rheological properties of starch. Starch 67: 213-224 (2015)
Anagnostopoulos C, Spizizen J. Requirements for transformation in Bacillus subtilis. J. Bacteriol. 81: 741-746 (1961)
Armenta S, Moreno-Mendieta S, Sánchez-Cuapio Z, Sánchez S, Rodríguez-Sanoja R. Advances in molecular engineering of carbohydrate-binding modules. Proteins 85: 1602-1617 (2017)
Baroroh U, Yusuf M, Rachman SD, Ishmayana S, Syamsunarno MRAA, Levita J, Subroto T. The importance of surface-binding site towards starch-adsorptivity level in α-amylase: a review on structural point of view. Enzyme Res. 2017: 4086845 (2017)
Božić N, Lončar N, Slavić MŠ, Vujčić Z. Raw starch degrading α-amylases: an unsolved riddle. Amylase 1: 12-25 (2017)
Christiansen C, Abou Hachem M, Janecek S, Viksø-Nielsen A, Blennow A, Svensson B. The carbohydrate-binding module family 20-diversity, structure, and function. FEBS J. 276: 5006-5029 (2009)
Cinelli BA, Castilho LR, Freire DMG, Castro AM. A brief review on the emerging technology of ethanol production by cold hydrolysis of raw starch. Fuel 150: 721-729 (2015)
de Souza PM, de Oliveira Magalhães P. Application of microbial α-amylase in industry – a review. Braz. J. Microbiol. 41: 850-861 (2010)
Homaei A, Ghanbarzadeh M, Monsef F. Biochemical features and kinetic properties of α-amylases from marine organisms. Int. J. Biol. Macromol. 83: 306-314 (2016)
Janecek S, Svensson B, MacGregor EA. α-Amylase: an enzyme specificity found in various families of glycoside hydrolases. Cell. Mol. Life Sci. 71: 1149-1170 (2014)
Koropatkin NM, Smith TJ. SusG: a unique cell-membrane-associated α-amylase from a prominent human gut symbiont targets complex starch molecules. Structure 18: 200-215 (2010)
Lin HY, Chuang HH, Lin FP. Biochemical characterization of engineered amylopullulanase from Thermoanaerobacter ethanolicus 39E-implicating the non-necessity of its 100 C-terminal amino acid residues. Extremophiles 12: 641-650 (2008)
Lo HF, Lin LL, Chiang WY, Chie MC, Hsu WH, Chang CT. Deletion analysis of the C-terminal region of the α-amylase of Bacillus sp. strain TS-23. Arch. Microbiol. 178: 115-123 (2002)
Lombard V, Golaconda Ramulu H, Drula E, Coutinho PM, Henrissat B. The carbohydrate-active enzymes database (CAZy) in 2013. Nucleic Acids Res. 42: D490-D495 (2014)
Machovic M, Janecek S. The evolution of putative starch-binding domains. FEBS Lett. 580: 6349-6356 (2006)
Majzlova K, Janecek S. Two structurally related starch-binding domain families CBM25 and CBM26. Biologia 69: 1087-1096 (2014)
Marchler-Bauer A, Lu S, Anderson JB, Chitsaz F, Derbyshire MK, DeWeese-Scott C, Fong JH, Geer LY, Geer RC, Gonzales NR, Gwadz M, Hurwitz DI, Jackson JD, Ke Z, Lanczycki CJ, Lu F, Marchler GH, Mullokandov M, Omelchenko MV, Robertson CL, Song JS, Thanki N, Yamashita RA, Zhang D, Zhang N, Zheng C, Bryant SH. CDD: a Conserved Domain Database for the functional annotation of proteins. Nucleic Acids Res. 39: D225-D229 (2011)
Mehta D, Satyanarayana T. Biochemical and molecular characterization of recombinant acidic and thermostable raw-starch hydrolysing α-amylase from an extreme thermophile Geobacillus thermoleovorans. J. Mol. Catal. B Enzym. 85-86: 229-238 (2013)
Mehta D, Satyanarayana T. Domain C of thermostable α-amylase of Geobacillus thermoleovorans mediates raw starch adsorption. Appl. Microbiol. Biotechnol. 98: 4503-4519 (2014)
Michael RG, Joseph S. Molecular Cloning: A Laboratory Manual. 4th ed. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, USA. pp. 350-361 (2014)
Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 31: 426-428 (1959)
Peng H, Chen M, Yi L, Zhang X, Wang M, Xiao Y, Zhang N. Identification and characterization of a novel raw-starch-degrading a-amylase (AmyASS) from the marine fish pathogen Aeromonas salmonicida ssp. salmonicida. J. Mol. Catal B Enzym. 119: 71-77 (2015)
Peng H, Zheng Y, Chen M, Wang Y, Xiao Y, Gao Y. A starch-binding domain identified in α-amylase (AmyP) represents a new family of carbohydrate-binding modules that contribute to enzymatic hydrolysis of soluble starch. FEBS Lett. 588: 1161-1167 (2014)
Rana N, Walia A, Gaur A. α-Amylases from microbial sources and its potential applications in various industries. Natl. Acad. Sci. Lett. 36: 9-17 (2013)
Robertson GH, Wong DW, Lee CC, Wagschal K, Smith MR, Orts WJ. Native or raw starch digestion: a key step in energy efficient biorefining of grain. J. Agric. Food Chem. 54: 353-365 (2006)
Rodriguez Sanoja R, Morlon-Guyot J, Jore J, Pintado J, Juge N, Guyot JP. Comparative characterization of complete and truncated forms of Lactobacillus amylovorus α-amylase and role of the C-terminal direct repeats in raw-starch binding. Appl. Environ. Microbiol. 66: 3350-3356 (2000)
Sokarda Slavic M, Pešić M, Vujčić Z, Božić N. Overcoming hydrolysis of raw corn starch under industrial conditions with Bacillus licheniformis ATCC 9945a α-amylase. Appl. Microbiol. Biotechnol. 100: 2709-2719 (2016)
Sumitani J, Tottori T, Kawaguchi T, Arai M. New type of starch-binding domain: the direct repeat motif in the C-terminal region of Bacillus sp. no. 195 α-amylase contributes to starch binding and raw starch degrading. Biochem. J. 350: 477-484 (2000)
Sun H, Ge X, Wang L, Zhao P, Peng M. Microbial production of raw starch digesting enzymes. Af. J. Biotechnol. 8: 1734-1739 (2009)
Sun HY, Zhao P, Ge X, Xia Y, Hao Z, Liu J, Peng M. Recent advances in microbial raw starch degrading enzymes. Appl. Biochem. Biotechnol. 2010. 160: 988-1003 (2010)
Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 28(10): 2731-2739 (2011)
Tibbot BK, Wong DWS, Robertson GH. Studies on the C-terminal region of barley α-amylase 1 with emphasis on raw starch-binding. Biologia 57: 229-238 (2002)
Waterhouse A, Bertoni M, Bienert S, Studer G, Tauriello G, Gumienny R, Heer FT, de Beer Tap, Rempfer C, Bordoli L, Lepore R, Schwede T. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46: W296-W303 (2018)
Wu XR, Wang Y, Tong B, Chen X, Chen J. Purification and biochemical characterization of a thermostable and acid-stable α-amylase from Bacillus licheniformis B4-423. Int. J. Biol. Macromol. 109: 329-337 (2018)
Zhang Q, Han GY, Xiao HZ. Microbial α-amylase: a biomolecular overview. Process Biochem. 53: 88-101 (2017)
Acknowledgments
This work was financially supported by grants from the National Natural Science Foundation of China (Grant Number 31501422) and the Jiangxi Natural Science Foundation (Grant Numbers 20151BAB214001 and 20171BAB214003).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Zeng, J., Guo, J., Tu, Y. et al. Functional study of C-terminal domain of the thermoacidophilic raw starch-hydrolyzing α-amylase Gt-amy. Food Sci Biotechnol 29, 409–418 (2020). https://doi.org/10.1007/s10068-019-00673-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10068-019-00673-x