
Abstract
In arid environments, shade provided by vegetation forms the crux of many facilitation pathways by providing other organisms with relief from high levels of solar radiation and extreme temperatures. Shade is an important determinant that structures arid ecosystem processes and functioning. While shade is considered an essential refuge for many organisms in arid environments, few studies have assessed how parasitic plants such as aerial mistletoes can influence the quality of shade provided by its host, and how this can subsequently influence species interactions. Here, we investigate how mistletoe influences the microclimate and daytime use of sub-canopy habitat by a large herbivore that seeks out shady micro-habitats to avoid daytime heat in Australia’s Strzelecki Desert. We experimentally removed the pale-leaved mistletoe (Amyema maidenii) from mulga trees (Acacia aneura) and monitored sub-canopy ambient temperatures and the daytime use of sub-canopy habitat by red kangaroos (Osphranter rufus). Results showed that average hourly temperature during the daytime and daily maximum temperature were cooler and that kangaroos were approximately three times more likely to shelter beneath trees where mistletoe was present (control) compared to where mistletoes had been removed (treatment). By showing that mistletoe can improve the quality of shade provided by their host trees, our study offers a novel insight into how mistletoes can facilitate the surrounding biota through modifying the abiotic environment beyond its physical structure. More broadly, the results of our study emphasise the prevalence of positive interactions within ecological communities and how they can occur in unexpected ways and indirectly between organisms that never come into direct contact.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Highlights
-
Mistletoes can modify the microclimate at the scale of its host canopy
-
This microclimate effect can indirectly facilitate kangaroo resting behaviour
-
Mistletoes can facilitate a broader range of organisms than previously anticipated
Introduction
Historically much of the focus on the role that inter-specific interactions play in the organisation and functioning of ecosystems has been on understanding negative interactions such as competition and predation (Bruno and others 2003). However, facilitative (positive) interactions occur ubiquitously and play a fundamental role in ecosystem functioning (Michalet and Pugnaire 2016). Facilitation occurs when organisms directly or indirectly interact and at least one benefits while causing no harm to any of the organisms involved (Bertness and Callaway 1994; Bruno and others 2003; Bronstein 2009). For example, in many ecosystems foundation species, such as trees, coral and reef forming mollusks, provide the physical structure which facilitates occupancy by an array of other species (Stachowicz 2001; Angelini and others 2011).
In arid systems, shade is an essential but limited resource that is the crux of many facilitative interactions (Dean and others 1999; Cain and others 2006; Callaway 2007). This is because shade offers refuge from high levels of solar radiation and ameliorates high daytime temperatures that often occur in arid environments. Shade is typically provided by larger shrubs and trees, where even sparse canopy foliage can screen a significant proportion of incoming solar radiation and function as a thermal refuge (Dawson and Denny 1969). For example, trees often facilitate large terrestrial mammals that seek the shade they create in order to reduce heat adsorption and their thermal load (Terrien 2011; Fuller and others 2014; Hetem and others 2014).
In recent decades, there has been an increasing acknowledgement of the role that mistletoes play in ecosystems through a range of facilitative interactions (Watson 2001, 2016; Mathiasen and others 2008). One key mechanism by which mistletoes facilitate other organisms is by creating resource-rich patches through redistributing nutrients acquired from their host. For example, many mistletoes possess nutrient rich leaves that have short lifespans and do not resorb nutrients prior to leaf-fall (March and Watson 2007). Consequently, mistletoes contribute to both an increase in quantity and quality of litter where the understorey soil is more fertile and can support a greater diversity of non-host plant species (Ndagurwa and others 2016a). The increase in litterfall also has indirect flow-on effects where litter-dwelling arthropods increase in abundance but also insectivorous birds that prey on these arthropods (Watson 2015; Mellado and others 2019). Indeed, there is increasing support for the theory that mistletoes function as keystone species by directly and indirectly facilitating multiple trophic levels (Watson 2009).
Mistletoes are also thought to facilitate habitat use of other organisms through the distinct microclimate they create (Watson 2001; Press and Phoenix 2005; Cooney and others 2006). It is theorised that the shade offered by the dense foliage of mistletoes coupled with evaporative cooling due to the high transpiration rates of mistletoes (Glatzel 1983; Ullmann and others 1985; Davidson and others 1989) could ameliorate harsh environmental conditions and in so doing create habitat for other species (Watson 2001; Press and Phoenix 2005; Cooney and others 2006). However, the effects that mistletoe has on the surrounding climate has rarely been measured (Ndagurwa and others 2016b), and its effect on animal behaviour is either anecdotal or deduced post hoc (Watson 2001; Cooney and others 2006). Furthermore, studies that have assumed the importance of a microclimate effect, have been limited to animals that come into direct contact with mistletoe by eating it, seeking shelter in mistletoe or consuming species that live in mistletoe (Watson 2001; Cooney and others 2006). Although it is conceivable that a mistletoe microclimate effect may also indirectly influence a broader range of animals that do not come into contact with mistletoe (Hartley and others 2015; Mellado and others 2019), this has yet to be studied.
In this study, we investigate whether the presence of mistletoe modulates the microclimate beneath its host and whether this indirectly facilitates a large vertebrate in a hot, shade limited arid environment. In our Strzelecki Desert study area, red kangaroos (Osphranter rufus, hereafter kangaroos) frequently use the shade (Figure 1A) provided by mulga trees (Acacia aneura) as resting sites where they avoid daytime heat (Dawson and Denny 1969; Dawson 1973; Roberts and others 2016). Mulga trees are often infected by mistletoes, which significantly augment the canopy of their hosts (Figure 1B, C). Consequently, we hypothesised that the presence of mistletoe modifies the microclimate under mulga trees by improving the quality of shade and thus influence kangaroos’ choice of resting sites. We tested our hypothesis by experimentally removing mistletoe from mulga trees and then measuring the daily maximum temperature under the mulga canopy and daytime use of mulga trees by kangaroos over a 4-month period. Our predictions were: (1) that daily maximum temperature would be greater under trees where mistletoe was removed and (2) that daytime kangaroo activity would be lower under trees where mistletoe was removed.

A Pale-leaf mistletoe (Amyema maidenii) infection on mulga (Acacia aneura), B pale-leaf mistletoe clumps within the canopy of a mulga tree, C red kangaroo (Osphranter rufus) resting beneath one of our procedural control trees where mistletoe was present in the canopy, D dead red kangaroo beneath a mulga tree.
Methods
Study Site
This study was conducted in Sturt National Park (29° 24′ S, 141° 2′ E) in the Strzelecki Desert in north-western New South Wales. The vegetation of the study area is classified as Sand Plain Mulga Shrubland that constitutes sparse overstorey shrubs and ephemeral grasses (Ludwig and others 1996). Mean annual rainfall of the nearest weather station (Fort Grey: 29° 5′ S, 141° 12′ E) study area is 173.8 mm.
In our study site, mulga is the most common tree and rarely exceeds 4 metres in height (Keith 2004). Within the canopy of mulga trees in our study area, two mistletoe species occur—Amyema maidenii and Lysiana murrayi. However, A. maidenii is significantly more abundant than L. murrayi.
Experimental Design
Our tree-scale manipulation study involved three treatment conditions that were grouped together within groves of trees situated on separate sand dunes for a total of 24 trees. The groves of trees selected for experimentation were situated a minimum of 1 km apart. Within each grove, trees were initially shortlisted according to two conditions: that the mistletoe A. maidenii was present and that trees were a similar size and had similar levels of mistletoe infection.
Within each grove, trees were allocated an identity and then randomly allocated a treatment condition (mistletoe removal, control, procedural control). For the mistletoe removal treatment, all mistletoe was removed from the tree using extendable loppers and pruning saws. To ensure that there was no potential for mistletoe to resprout, cutting was conducted close to the haustorium (Watson 2019). To ensure that the removal treatment was successful, trees were periodically checked for mistletoe resprouting during the study. No resprouting mistletoe was found. As the mistletoe removal condition involved the modification of the canopy structure, a procedural control was used to ensure that the results of our study were due to the removal of mistletoe and not due to the modification of the canopy structure. For the procedural control, mistletoe was left undisturbed, but we disturbed the canopy structure by removing a volume of mulga foliage from the northern aspect of the canopy, similar to the volume of mistletoe removed in the mistletoe removal tree within the same grove. For the control trees, the mulga canopy and mistletoe were left undisturbed.
Temperature Loggers
Onset® HOBO U22-001 data loggers were used to measure the temperature beneath the canopy of each tree. Because the sun traverses across the northern sky in the southern hemisphere and our initial observations suggested that kangaroos predominantly rest on the south side of trees to maximise shading each logger was placed on the south side of the tree. Loggers were suspended within the canopy approximately 75 cm from the ground and 1.5 m from the base of the tree. The temperature measurement range of each data logger is from − 40° to 70 °C with an accuracy of ± 0.21 °C. Each HOBO logger was programmed to record the temperature once every hour. Due to one malfunctioning data logger, data from 23 trees were used in this study (control: n = 7, procedural control n = 8 and mistletoe removal, n = 8).
Kangaroo Activity Under Trees
We used time-lapse photography to provide an index of the amount of time kangaroos spent in the shade of our experimental mulga trees (n = 24 trees). The cameras were programmed to take a photograph every hour. The rationale for using time-lapse photography as opposed to motion-triggered photography was because kangaroos tend to be immobile for long periods while they rest under trees, then the time they spend under trees could be reliably quantified by taking periodic photographs. An additional rationale was that we wanted to optimise detections of kangaroos versus the risk of filling the camera memory cards with false triggers due to movement of vegetation and litter by wind. This subsequently ensured that the battery life of the cameras would last longer, reducing the need for human replacing batteries at our remote study site.
We deployed a RECONYX HC600 Hyperfire (RECONYX, Holmen, WI, USA) trail-camera at each of our experimental trees. Cameras were each mounted to a stake placed approximately 10 m from the tree base. Cameras were placed on the south side of the trees because our initial observations suggested that kangaroos predominantly rest on the south side of trees to maximise shading as the sun traverses across the northern sky in the southern hemisphere. All photographs were manually reviewed, and any daytime observations of kangaroos were logged. Camera traps were deployed from June 2018 until March 2019. However, due to drought related mortality, the population of kangaroos had severely declined by October 2018 such that no photographs of kangaroos were recorded between November 2018 and March 2019. Therefore, for analysis, the number of daytime observations for each camera was calculated as the number of kangaroo sightings divided by the numbers of days that the camera was operational between June 2018 and October 2018.
Data Analysis
Generalised estimating equations are an extension of generalised linear models that are suited to the analysis of data with repeated measurements of the same individuals (Zuur 2009). We used generalised estimating equations with a Gaussian distribution to investigate the effect that the mistletoe removal treatment had on maximum daily temperatures under the tree canopy. Day of measurement was treated as a repeated measure with an independent error structure. Planned post hoc comparisons were made using Fisher’s least difference test to further investigate how the treatments differed from one another. SPSS 24.0 software (IBM Corp 2016) was used for this analysis.
Using the software environment R4.0.0 (R Core Team 2020), a generalised linear model (GLM) was used to analyse the difference in kangaroo usage of trees that had mistletoe and trees without mistletoe. Count data were analysed with a negative binomial distribution, using the “mass” package (Venables and Ripley 2002). The number of days that cameras were operational was used as an offset in the model as not all cameras functioned equally. Planned post hoc comparisons were conducted to further investigate how the treatments differed from one another. Tukey′s honest significant difference test were conducted using the “multcomp” package (Hothorn and others 2008).
Results
Temperature Loggers
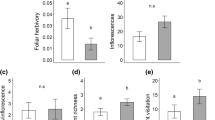
During the daytime, the understorey microclimate was cooler where mistletoe was present compared to where mistletoe was removed. This was evidenced by the average hourly temperature during daytime hours 0900–1700 h being consistently cooler where mistletoe was present (Figure 2).

Diel fluctuation of ambient temperature (°C) recorded in the sub-canopy of mulga trees (Acacia aneura) with mistletoe (control, n = 7), with mistletoe but a proportion of mulga foliage removed (procedural control, n = 8) and mistletoe removed (mistletoe treatment, n = 8). Each value represents the mean temperature for trees in the same condition, across all days of the study period (16 June 2018 until 31 October 2018) and at that time (± 95% CI).
Across the duration of the study, there was a significant effect of treatment (Wald χ2 7.576, df2, P = 0.023) on the daily maximum temperature beneath the tree canopy (Figure 3). On average, the daily maximum temperature was greater beneath trees where mistletoe had been removed (28.9 °C se 0.8 °C) when compared to control trees (26.5 °C se 0.2 °C) and procedural controls (26.7° C se 0.2 °C) where mistletoe was present (Fisher’s LSD removal vs. control P = 0.01, removal vs. procedural P = 0.007). Though, there was no significant difference in the maximum temperature between the control trees and the procedural control trees where the host foliage was partially removed in the procedural control (procedural vs. control P = 0.850).

Time series of the median daily maximum sub-canopy temperature (°C) grouped by treatment condition (control, n = 7; procedural control, n = 8 and mistletoe removal n = 8) and between July and October 2018.
Macropod Activity
In total, there were 64 daytime observations of kangaroos beneath the canopy of mulga trees. Overall, 37.5% of kangaroo observations were beneath control trees, 51.5% beneath procedural trees and 11% beneath treatment trees. There was a significant effect of the mistletoe removal treatment on kangaroo activity under mulga trees (Wald test: z = − 6.029, df2, P < 0.001). Kangaroo activity was greater under control (Tukey’s HSD control vs. treatment P < 0.001) and procedural trees (Tukey’s HSD procedural vs treatment P < 0.001) where mistletoe was present than under the trees from which mistletoe had been removed (Figure 4). Kangaroo activity did not differ between the control and procedural control (control vs. procedural P = 0.113) which indicates that the removal of mulga foliage had no effect on kangaroo activity under the trees. Kangaroo activity did not vary according to canopy volume (P = 0.61) or mulga tree height (P = 0.724).

Mean daytime kangaroo activity beneath mulga trees from which mistletoes had been removed (n = 8), and control trees with mistletoe (n = 8) and procedural control trees with mistletoe but a proportion of mulga foliage removed (n = 8) between July and October 2018.
Discussion
In accord with prediction 1, the presence of mistletoe contributed to a cooler understorey when compared to that of control trees and procedural control trees. In accord with prediction 2, kangaroos spent less time resting beneath trees from which mistletoe had been removed than trees with mistletoe. Taken together, our results show that the shade generated by mistletoe can moderate the understorey microclimate of mulga trees and in doing so facilitates kangaroos’ use of the sub-canopy. More generally, our study shows how mistletoe can moderate the microclimate beyond its physical structure and influence the behaviour of animals that do not directly come into contact with mistletoe.
Our experimental manipulation showed that mistletoe created a cooler sub-canopy microclimate during the daytime. This was reflected in both the daytime average hourly temperatures and the daily maximum temperatures (Figures 2 and 3). This effect was likely due to the difference in morphology between the leaves and branches of pale-leaf mistletoe and mulga, where the presence of mistletoe increases the canopy density of their host. Mulga trees have a relatively sparse canopy because their phyllodes are narrow and orientate vertically to minimise heat adsorption and water loss from leaf transpiration (Miller and others 2002). In contrast, pale-leaf mistletoes form dense clumps of intertwining branches and have larger more horizontally orientated leaves that intercept more light than the leaves of their mulga hosts (Fig. 1B; Watson 2019). Consequently, the presence of mistletoe makes the sub-canopy more shady and cooler because it reduces direct radiation from the sun by increasing the canopy’s absorption and reflectance of incoming solar radiation (Lin and Lin 2010; von Arx and others 2012). Furthermore, the relatively high transpiration rate of mistletoes (Ullmann and others 1985) could also contribute to lowering of the sub-canopy temperature by increasing the rate of evaporative cooling (Griebel and others 2017).
Although mulga trees only offer sparse shade, kangaroos frequently spend the daytime resting beneath mulga trees (Dawson 1973). This is because there are few other sources of shade in our desert system and the shade provided by mulga trees is still sufficient to enable kangaroos to reduce their thermal load (Dawson and Denny 1969; Roberts and others 2016). However, our results showing that kangaroos spent more time resting under trees with mistletoe whose average maximum sub-canopy temperature was approximately 2 °C lower than that under trees without mistletoe suggest that kangaroos actively select trees with mistletoe because they benefit from the cooler sub-canopy. We are confident that kangaroos selected trees with mistletoe for resting as they are not known to graze on mulga or mistletoe species (Pahl 2019) and there was also no evidence of any grazing damage to the foliage of mulga or mistletoe on our study trees.
Kangaroos rely on shade less during winter, but our results show that shade-seeking behaviour still occurs in the cooler months and they do this to reduce their heat load (Roberts and others 2016). Although we intended to run our study through the Austral summer, there was a mass die-off of kangaroos due to drought related starvation during the Austral Spring. Consequently, kangaroos became rare and stopped using our study trees. Had kangaroos been present in the same numbers during summer, we expected that they would have used the shade more and that the effect of our mistletoe removal would have been more marked. Nevertheless, our results showing that kangaroos prefer to rest under trees with mistletoe provide evidence that shade is a resource for kangaroos and that they choose resting sites based on the quality of the shade they provide.
This finding raises the question; do mulga trees or mistletoes gain any benefit from kangaroos resting in the shade they provide? In our study, mulga trees could benefit from the nutrients that kangaroos bring in the form of dung and urine which they deposit while resting (Figure 1D; Eldridge and Rath 2002). In addition, it is not uncommon for kangaroos to die while they shelter under mulga trees and fertilise trees as their carcasses decompose (Figure 1D: Wilson and Read 2003; Fillios and others 2010). However, we think it is unlikely that mulga trees benefit from the presence of mistletoes because mistletoes likely impose a cost on the trees by parasitising water and nutrients, but it is conceivable that mistletoes may benefit from the nutrients that kangaroos transport to trees (Eldridge and Rath 2002). In support of this idea, previous studies show that mistletoes are more abundant on hosts with greater access to water and fertile soils (Dean and others 1994; Norton and Smith 1999).
In desert environments, even small modifications of the climate may make survival through hot and dry periods more likely for animals and thus elicit distinct behavioural preferences for thermal refuges (Dawson and others 2006; Cain and others 2006; Fuller and others 2014). Many small endotherms avoid the heat of the day by being nocturnal and using burrows or cavities as thermal refuges (Chapman 2013; Amat-Valero and others 2014). However, while desert endotherms that are too large to seek shelter underground or lack fossorial adaptations generally have a relatively high thermal tolerance, they must still reduce their thermal load during the day but have relatively few sites available that they can use as thermal shelters (Fuller and others 2016). Our findings show that even a relatively small difference in microclimate (approximately 2 °C) generated by the presence of mistletoe can influence the daytime refuge selection of a relatively large-bodied desert endotherm.
Our study offers two unique insights into how mistletoes can facilitate their biotic environment by creating a distinct microclimate. First, although our results concord with previous research showing that mistletoe clumps are cooler than the surrounding foliage (Ndagurwa and others 2016b), we demonstrate that the influence of a mistletoe microclimate can extend beyond the physical structure of mistletoe clumps to the sub-canopy microclimate. Second, our study shows how this moderation of the sub-canopy microclimate by mistletoe can then indirectly facilitate habitat use by a larger-bodied animal that does not directly contact mistletoe. More broadly, the results of our study emphasise the prevalence of positive interactions within ecological communities and how they can occur in unexpected ways and indirectly between organisms that never come into direct contact.
References
Amat-Valero M, Calero-Torralbo MA, Václav R, Valera F. 2014. Cavity types and microclimate: implications for ecological, evolutionary, and conservation studies. Int J Biometeorol 58:1983–94.
Angelini C, Altieri AH, Silliman BR, Bertness MD. 2011. Interactions among Foundation Species and Their Consequences for Community Organization, Biodiversity, and Conservation. BioScience 61:782–9.
Bertness MD, Callaway R. 1994. Positive interactions in communities. Trends in Ecology and Evolution 9:191–3.
Bronstein JL. 2009. The evolution of facilitation and mutualism. Journal of Ecology 97:1160–70.
Bruno JF, Stachowicz JJ, Bertness MD. 2003. Inclusion of facilitation into ecological theory. Trends in Ecology and Evolution 18:119–25.
Cain JW, Krausman PR, Rosenstock SS, Turner JC. 2006. Mechanisms of Thermoregulation and Water Balance in Desert Ungulates. Wildlife Society Bulletin 34:570–81.
Callaway RM. 2007. Positive Interactions and Interdependence in Plant Communities. Dordrecht: Springer. https://doi.org/10.1007/978-1-4020-6224-7
Chapman TF. 2013. Relic bilby (Macrotis lagotis) refuge burrows: assessment of potential contribution to a rangeland restoration program. Rangel J 35:167–80.
Cooney SJN, Watson DM, Young J. 2006. Mistletoe nesting in Australian birds: a review. Emu - Austral Ornithology 106:1–12.
Davidson NJ, True KC, Pate JS. 1989. Water relations of the parasite: host relationship between the mistletoe Amyema linophyllum (Fenzl) Tieghem and Casuarina obesa Miq. Oecologia 80:321–30.
Dawson TJ. 1973. Thermoregulatory responses of the arid zone kangaroos, Megaleia rufa and Macropus robustus. Comparative Biochemistry and Physiology Part A: Physiology 46:153–69.
Dawson TJ, Denny MJS. 1969. A Bioclimatological Comparison of the Summer Day Microenvironments of Two Species of Arid-Zone Kangaroo. Ecology 50:328–32.
Dawson TJ, McTavish KJ, Munn AJ, Holloway J. 2006. Water use and the thermoregulatory behaviour of kangaroos in arid regions: Insights into the colonisation of arid rangelands in Australia by the Eastern Grey Kangaroo (Macropus giganteus). Journal of Comparative Physiology B 176:45–53.
Dean WRJ, Midgley JJ, Stock WD. 1994. The Distribution of Mistletoes in South Africa: Patterns of Species Richness and Host Choice. Journal of Biogeography 21:503.
Dean WRJ, Milton SJ, Jeltsch F. 1999. Large trees, fertile islands, and birds in arid savanna. Journal of Arid Environments 41:61–78.
Eldridge DJ, Rath D. 2002. Hip holes: kangaroo (Macropus spp.) resting sites modify the physical and chemical environment of woodland soils: KANGAROOS’ HIP HOLES AND SOILS. Austral Ecology 27:527–36.
Fillios M, Gordon C, Koch F, Letnic M. 2010. The effect of a top predator on kangaroo abundance in arid Australia and its implications for archaeological faunal assemblages. Journal of Archaeological Science 37:986–93.
Fuller A, Hetem RS, Maloney SK, Mitchell D. 2014. Adaptation to Heat and Water Shortage in Large, Arid-Zone Mammals. Physiology 29:159–67.
Fuller A, Mitchell D, Maloney SK, Hetem RS. 2016. Towards a mechanistic understanding of the responses of large terrestrial mammals to heat and aridity associated with climate change. Climate Change Responses 3:1–19. https://doi.org/10.1186/s40665-016-0024-1.
Glatzel G. 1983. Mineral nutrition and water relations of hemiparasitic mistletoes: a question of partitioning. Experiments with Loranthus europaeus on Quercus petraea and Quercus robur. Oecologia 56:193–201.
Griebel A, Watson D, Pendall E. 2017. Mistletoe, friend and foe: synthesizing ecosystem implications of mistletoe infection. Environmental Research Letters 12:115012.
Hartley SE, Green JP, Massey FP, Press MCP, Stewart AJA, John EA. 2015. Hemiparasitic plant impacts animal and plant communities across four trophic levels. Ecology 96:2408–16.
Hetem RS, Fuller A, Maloney SK, Mitchell D. 2014. Responses of large mammals to climate change. Temperature 1:115–27.
Hothorn T, Bretz F, Westfall P. 2008. Simultaneous Inference in General Parametric Models. Biometrical Journal 50:346–63.
IBM Corp. 2016. IBM SPSS Statistics for Windows, Version 24.0. Armonk, NY: IBM Corp
Keith DA. 2004. Ocean shores to desert dunes: the native vegetation of New South Wales and the ACT. Hurstville, NSW: Hurstville, NSW: Dept. of Environment and Conservation NSW
Lin B-S, Lin Y-J. 2010. Cooling Effect of Shade Trees with Different Characteristics in a Subtropical Urban Park. HortScience 45:83–6.
Ludwig J, Tongway D, Hodgkinson K, Freudenberger D, Noble J. 1996. Landscape Ecology, Function and Management: Principles from Australia’s Rangelands. CSIRO Publishing https://ebooks.publish.csiro.au/content/ISBN/9780643101159. Last accessed 02/05/2020
March WA, Watson DM. 2007. Parasites boost productivity: effects of mistletoe on litterfall dynamics in a temperate Australian forest. Oecologia 154:339–47.
Mathiasen RL, Nickrent DL, Shaw DC, Watson DM. 2008. Mistletoes: Pathology, Systematics, Ecology, and Management. Plant Disease 92:988–1006.
Mellado A, Hobby A, Lázaro-González A, Watson DM. 2019. Hemiparasites drive heterogeneity in litter arthropods: Implications for woodland insectivorous birds. Austral Ecology 44:777–85.
Michalet R, Pugnaire FI. 2016. Facilitation in communities: underlying mechanisms, community and ecosystem implications. Functional Ecology 30:3–9.
Miller JT, Andrew RA, Maslin BR. 2002. Towards an understanding of variation in the Mulga complex (Acacia aneura and relatives). 17.
Ndagurwa HGT, Ndarevani P, Muvengwi J, Maponga TS. 2016a. Mistletoes via input of nutrient-rich litter increases nutrient supply and enhance plant species composition and growth in a semi-arid savanna, southwest Zimbabwe. Plant Ecology 217:1095–104.
Ndagurwa HGT, Nyawo E, Muvengwi J. 2016b. Use of mistletoes by the Grey Go-away-bird (Corythaixoides concolor, Musophagidae) in a semi-arid savannah, south-west Zimbabwe. African Journal of Ecology 54:336–41.
Norton DA, Smith MS. 1999. Why might roadside mulgas be better mistletoe hosts? Austral Ecology 24:193–8.
Pahl L. 2019. Macropods, feral goats, sheep and cattle. 2. Equivalency in what and where they eat. The Rangeland Journal 41:519.
Press MC, Phoenix GK. 2005. Impacts of parasitic plants on natural communities: Tansley review. New Phytologist 166:737–51.
R Core Team. 2020. R: A language and environment for statistical computing. Vienna, Austria: R Foundation for Statistical Computing https://www.R-project.org/
Roberts JA, Coulson G, Munn AJ, Kearney MR. 2016. A continent-wide analysis of the shade requirements of red and western grey kangaroos. Temperature 3:340–53.
Stachowicz JJ. 2001. Mutualism, Facilitation, and the Structure of Ecological Communities. BioScience 51:235.
Terrien J. 2011. Behavioral thermoregulation in mammals: a review. Frontiers in Bioscience 16:1428.
Ullmann I, Lange OL, Ziegler H, Ehleringer J, Schulze E-D, Cowan IR. 1985. Diurnal courses of leaf conductance and transpiration of mistletoes and their hosts in Central Australia. Oecologia 67:577–87.
Venables WN, Ripley BD. 2002. Modern Applied Statistics with S. 4th edn. New York: Springer.
von Arx G, Dobbertin M, Rebetez M. 2012. Spatio-temporal effects of forest canopy on understory microclimate in a long-term experiment in Switzerland. Agricultural and Forest Meteorology 166–167:144–55.
Watson D M. 2019. Mistletoes of Southern Australia. 2nd ed. CSIRO Publishing
Watson DM. 2001. Mistletoe—A Keystone Resource in Forests and Woodlands Worldwide. Annual Review of Ecology and Systematics 32:219–49.
Watson DM. 2009. Determinants of parasitic plant distribution: the role of host quality. Botany 87:16–21.
Watson DM. 2015. Disproportionate Declines in Ground-Foraging Insectivorous Birds after Mistletoe Removal. Umapathy G, editor. PLOS ONE 10:e0142992.
Watson DM. 2016. Fleshing out facilitation–reframing interaction networks beyond top-down versus bottom-up. New Phytologist 211:803–8.
Wilson D, Read J. 2003. Kangaroo harvesters: fertilising the rangelands. The Rangeland Journal 25:47.
Zuur AF. 2009. Mixed effects models and extensions in ecology with R. New York, NY: Springer.
Acknowledgements
The authors thank all the volunteers and employees that assisted with the fieldwork and those that contributed towards insightful discussions (especially Caitlin Buckle and Max Campbell).
Funding
Funding was provided by Australian Research Council (Grant No. DP180101477).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Chu, N., Cornwell, W. & Letnic, M. Mistletoes Facilitate a Desert Herbivore by Improving the Quality of Shade. Ecosystems 24, 1393–1401 (2021). https://doi.org/10.1007/s10021-020-00590-9
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-020-00590-9