
Abstract
Extensive high-severity fires are creating large shrubfields in many dry conifer forests of the interior western USA, raising concerns about forest-to-shrub conversion. This study evaluates the role of disturbance in shrubfield formation, maintenance and succession in the Jemez Mountains, New Mexico. We compared the environmental conditions of extant Gambel oak (Quercus gambelii) shrubfields with adjoining dry conifer forests and used dendroecological methods to determine the multi-century fire history and successional dynamics of five of the largest shrubfields (76–340 ha). Across the study area, 349 shrubfields (5–368 ha) occur in similar topographic and climate settings as dry conifer forests. This suggests disturbance, rather than other biophysical factors, may explain their origins and persistence. Gambel oak ages and tree-ring fire scars in our sampled shrubfields indicate they historically (1664–1899) burned concurrently with adjoining conifer forests and have persisted for over 115 years in the absence of fire. Aerial imagery from 1935 confirmed almost no change in sampled shrubfield patch sizes or boundaries over the twentieth century. The largest shrubfield we identified is less than 4% the size of the largest conifer-depleted and substantially shrub-dominated area recently formed in the Jemez following extensive high-severity wildfires, indicating considerable departure from historical patterns and processes. Projected hotter droughts and increasingly large high-severity fires could trigger more forest-to-shrub transitions and maintain existing shrubfields, inhibiting conifer forest recovery. Restoration of surface fire regimes and associated historical forest structures likely could reduce the rate and patch size of dry conifer forests being converted to shrubfields.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Across the interior western USA, megafires (> 10,000 ha) driven by climate change and land-use history have significantly affected fire-prone ecosystems (Stephens and others 2013, 2014). The dry conifer forests of the interior west, which include mid-elevation forests dominated by ponderosa pine (Pinus ponderosa) and other dry mixed-conifer species, can persist in a stable state maintained by frequent, low-severity fires; however, an increasing occurrence of moderate- to high-severity fires in some regions have resulted in high rates of widespread conifer mortality and dramatic changes in post-fire vegetation structure (Savage and Mast 2005; Collins and Roller 2013; Rother and Veblen 2016). One consequence of large patches of high-severity fire is an abrupt transition to resprouting shrub species (Figure 1), which commonly includes Gambel oak (Quercus gambelii) in the southwestern USA (Savage and Mast 2005; Haire and McGarigal 2008; Roccaforte and others 2012; Guiterman and others 2015; Coop and others 2016). Gambel oak proliferation post-fire is well documented (Kaufmann and others 2016), but what remains uncertain is whether the forest-to-shrub transitions following high-severity fires are persistent alternative stable states or a seral stage in dry conifer ecosystems (Falk 2013).

Gambel oak shrubfields in New Mexico. Images at top are recent fire-origin shrubfields while those at bottom are older, tree-ring sampled shrubfields in the Jemez Mountains. Top left Sugarite Canyon State Park: 2011 Track Fire; top right Mescalero Apache Reservation, Sacramento Mountains: 1996 Chino Well Fire; bottom left Mesa Pinabetosa site (MPB), northern Jemez Mountains; bottom-right San Pablo Canyon site (SPC), western Jemez Mountains. Bottom-right photo by N. Kessler, all others by E. Margolis.
Gambel oak is a common and ecologically important species of the Rocky Mountains and Sky Islands ecosystems of the interior western USA (Kaufmann and others 2016). It ranges from northern Utah and southern Wyoming, where spring frosts limit its growth, to the mountains of Sonora, Mexico, where drought and increased competition from Madrean oaks and chaparral shrubs limit it to cooler, wetter aspects and elevations (Little 1971; Rehfeldt and others 2006). Its growth character varies along this latitudinal gradient from tree-like single stems in the south to dense clonal shrubfields toward its northern limit (Harper and others 1985; Neilson and Wullstein 1986; Clary and Tiedemann 1992). In much of the central area of its distribution, including northern Arizona and New Mexico, shading from mature conifers and frequent surface fires are likely to have historically suppressed the growth and cover of Gambel oak, maintaining it as an understory shrub with occasional tall, single-stem codominants in ponderosa pine-dominated forests (Fulé and others 1997; Abella 2008). Following high-severity fires, Gambel oak rapidly and prolifically resprouts to form dense shrubfields regardless of its historic architecture. Widespread, newly formed shrubfields following recent megafires raise the possibility that Gambel oak is increasing its distribution and becoming more dominant across the landscape (Van Auken 2000; Abella and Fulé 2008; Kaufmann and others 2016).
Two main hypotheses exist regarding the long-term persistence of post-fire Gambel oak shrubfields. One suggests they are a seral stage in the succession of conifers (Hanks and Dick-Peddie 1974; Moir and others 1997; Haire and McGarigal 2008; Van Auken and Bush 2017), while the other suggests they can persist as an alternative stable state (Strom and Fulé 2007; Feddema and others 2013; Savage and others 2013). Demonstrating the existence of alternative stable states can be challenging because several key traits must be satisfied, including (1) they occur in the same environment, (2) the switch is associated with a loss of resilience and potentially triggered by a catastrophic disturbance, and (3) they persist beyond one generation of the dominant vegetation (Connell and Sousa 1983; Petraitis and Latham 1999; Scheffer and others 2001). This final criterion is the most difficult to document, but several ecological traits of resprouting shrubs support their potential ability to persist long term (Clarke and others 2013; Zeppel and others 2015; Pausas and others 2016). In the case of Gambel oak shrubfields, they likely can persist through drought and fire better than conifer stands (Savage and others 2013; Coop and others 2016), and their tendency to resprout in dense clumps confers a competitive advantage over conifer seedlings (Oliver 1990; Minor and others 2017; Van Auken and Bush 2017). Additionally, recolonization by seed-obligate conifers, like ponderosa pine, in large high-severity patches (100s to 1000s of hectares) can be hampered by a lack of nearby seed trees (Haire and McGarigal 2010; Chambers and others 2016) and drought stress to seedlings (Feddema and others 2013; Rother and others 2015).
The idea that fire-origin oak shrub communities are an early seral stage that eventually transition back to dry conifer forest is analogous to the dynamics of some higher-elevation conifer–aspen (Populus tremuloides) ecosystems. Following high-severity fire, even-aged seral aspen stems that resprout from long-lived clones can dominate for decades until being overtopped by conifers (Kashian and others 2007; Margolis and others 2007). Documented succession of clonal resprouting shrubfields to dry conifer forest is rare, but the natural co-occurrence of conifer and shrub species in the southwestern USA may indicate metastability of shrub-dominated patches, in that there could be some ability to change between dominant species at long-time scales (Falk 2013).
In more mesic environments with shade-tolerant conifers [for example, white fir (Abies concolor)], succession may be accelerated (Van Auken and Bush 2017). For example, in northern California, conifer forests have encroached upon montane chaparral, attributed to over a century of fire exclusion (Nagel and Taylor 2005; Airey Lauvaux and others 2016). Shrub-to-conifer succession may be more common across the western USA than is recognized, given that the evidence for shrub dominance would become obscured as conifers overtop the shrub canopy. However, results from several studies clearly demonstrate the potential for long-term post-fire persistence of Gambel oak, or other oak shrub species (Floyd and others 2000; Ryniker and others 2006; Iniguez and others 2009).
The objectives of this study are to (1) identify Gambel oak-dominated shrubfields in the Jemez Mountains, New Mexico, and compare their landscape distribution and environmental conditions to local dry conifer forests; (2) reconstruct the fire histories and age structures of five of the largest shrubfields via newly validated dendroecological methods (Guiterman and others 2015); (3) use these reconstructions in a “multiple lines of evidence” approach to determine the timing and extent of the high-severity fires that likely initiated the shrubfields; and (4) compare the historical fire regimes of shrubfields to the surrounding dry conifer forests of the Jemez Mountains. We then consider the implications of these findings for the successional trajectories of recent fire-origin shrubfields.
Methods
Study Area
This study was conducted in the Jemez Mountains of north-central New Mexico (Figure 2). The climate of the area is continental and semiarid, with bimodal precipitation that includes regional-scale frontal storms in winter and localized convective storms in summer (Sheppard and others 2002). Average annual precipitation across the Jemez totals 496 mm, with 41% occurring in July–September, based on 1981–2010 climate normals from the precipitation–elevation regression on independent slopes model (PRISM; Daly and others 2008). Average monthly temperatures range from a low of 0.4°C to a high of 16.0°C.

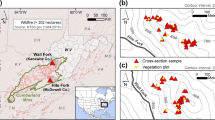
Tree-ring collections in Gambel oak shrubfields of the Jemez Mountains. Site maps have equal scales covering 2.6 km across; site names are provided in Table 1. Notations for sample types are FS, fire scar; DD, death date; GC, growth change; AGE, age structure plot. The center-top map shows the spatial distribution of dry conifer forests and late twentieth-century Gambel oak shrubfields. Insets include a regional map with the range of Gambel oak in blue (Little 1971) and the study area location, and the frequency-size distribution of Gambel oak shrubfields in the Jemez Mountains (Color figure online).
The Jemez Mountains are among the most sampled landscapes in the world for tree-ring fire history research, including about 1400 dated fire-scarred trees (Swetnam and others 2016; Margolis and others 2017). The historical fire regime was dominated by frequent, widespread, low-severity fires, with winter precipitation patterns exerting the greatest control over fire synchrony, and the North American Monsoon limiting and synchronizing warm-season fires (Margolis and others 2017). The collapse of the fire regime around 1860–1900 (Touchan and others 1996) is attributed to the arrival of the transcontinental railroad, which enabled intensive grazing that initiated twentieth-century fire exclusion (Allen 2007). In recent decades, fires have returned to the landscape, with several large, high-severity fires creating conifer-depleted patches of 1000 to more than 10,000 ha that are now mostly dominated by Gambel oak, New Mexico locust (Robinia neomexicana), and/or herbaceous vegetation (Savage and Mast 2005; Haire and McGarigal 2008; Allen 2016; Coop and others 2016).
Identification and Evaluation of Extant Shrubfields
We mapped all Gambel oak shrubfields larger than 5 ha that are embedded in dry conifer forests in the Jemez Mountains to evaluate their landscape distribution and environmental conditions in comparison with local conifer forests, and for the selection of tree-ring sampling sites (Figure 2). We used a combination of vegetation map datasets from the USDA Santa Fe National Forest, Southwest Regional Gap Analysis (Lowry and others 2007), Valles Caldera National Preserve (Muldavin and Tonne 2003), and Bandelier National Monument (Muldavin and others 2011). Visual analyses of aerial photographic imagery, comparison with other GIS layers, and field surveys were conducted to ensure that our final mapped shrubfields are dominated by Gambel oak and not misclassified. The vegetation layers we used are current to 2004, but do not include some Gambel oak-dominated shrubfields generated by the 2000 Cerro Grande Fire and the 1996 Dome Fire in the eastern Jemez. The shrubfield distribution also predates the 2011 Las Conchas Fire. Thus, we refer to the distribution of Gambel oak-dominated shrubfields as representing the Jemez landscape of the late twentieth century.
We compared the environmental conditions of shrubfields to local dry conifer forests to evaluate whether shrubfields occupy distinct landscape positions or climatic niches. Similar physiographic and climate settings would suggest the ability of Gambel oak shrubfields to occupy nearly any area within the dry conifer-dominated landscape, and for the current shrubfields to have formerly been forests, or the potential to return to forests. Dry conifer forests are defined as those mapped in SWReGAP as dominated by ponderosa pine (“Southern Rocky Mountain Ponderosa Pine Woodland”) or dry mixed-conifer (“Southern Rocky Mountain Dry-Mesic Montane Mixed-Conifer Forest and Woodland”). Because the latter forests “often have ponderosa pine” (SWReGAP database), it provides accurate coverage of dry mixed-conifer forests in the study area as opposed to mesic mixed-conifer forests where ponderosa pine is usually absent (Romme and others 2009). The comparison was done using principal components analysis (PCA) on the correlation matrix of ten variables from 20,000 randomly located GIS points divided between shrubfields and dry conifer forests. We included the physiographic variables of elevation, slope, aspect (transformed according to Beers and others 1966), curvature, topographic relative moisture index (TRMI; Parker 1982), and topographic position index (TPI; Jenness 2006). Climate variables came from an 800-m grid of annual precipitation and temperature (max, min, and mean) averaged for the 1981–2010 period (Daly and others 2008).
Site Selection and Field Sampling
We selected five of the largest shrubfields from the late twentieth-century shrubfield distribution for tree-ring sampling (Figure 2). The selection process included field surveys to evaluate the potential for these sites to be fire origin and to have originated prior to approximately 1935. This time period includes the earliest aerial imagery for the landscape and is covered by the local fire atlas (Snyderman and Allen 1997; data updated through 2014 by K. Beeley, Bandelier National Monument). Evidence of fire origin included distinct forest-shrub edges with no clear topographic component, large old Gambel oak clumps (genets), potentially fire-killed trees preserved as logs, snags, and/or occasional stumps, and living trees with elevated crowns and/or fire-scarred boles or branches (see Figure 2 in Guiterman and others 2015). Furthermore, sampling locations did not have known fire origins from fires documented in the local fire atlas. Each site had soils derived from local bedrock, which at four of five sites was Permian sandstone. One site, Redondo Creek (RDC), was located on the lower slopes of Redondo Peak in the Valles Caldera, and had bedrock composed of Pleistocene and Holocene rhyolite and basalt, intermixed with tuff.
The five selected sites range in elevation from 2450 to 2800 m. Understory composition of the shrubfields varies from grasses at lower slope positions to small shrubs [for example, gooseberry (Ribes leptanthum)] in the middle and upper slopes. New Mexico locust and aspen are present in mesic areas. Conifers are present as scattered individuals or in small clumps within shrubfields, usually close to the forest edge, and the species are typically ponderosa pine, Rocky Mountain Juniper (Juniperus scopulorum), Douglas-fir (Pseudotsuga menziesii), white fir, and piñon (Pinus edulis). Forests immediately adjacent to the shrubfields are dominated by ponderosa pine, but on cooler and wetter sites and aspects, forest composition includes Douglas-fir, southwestern white pine (Pinus strobiformis), and white fir.
At each site, we collected multiple lines of tree-ring evidence to characterize stand age and fire history. Our methods were previously validated in the study area, where we reconstructed a 1993 high-severity fire that generated a Gambel oak shrubfield in a former ponderosa pine-dominated forest (Guiterman and others 2015). Analogous methods have been used to reconstruct high-severity fire patches in upper-elevation mixed-conifer–aspen forests (Margolis and others 2007). The technique employs several sampling strategies to collect (1) shrub regeneration dates, (2) fire scars, (3) death dates, and (4) tree-ring growth changes or injuries. For regeneration (sprouting) dates, we installed at least ten circular age structure plots (0.10 ha) spatially distributed across each site. At each plot, we sampled at least two of the largest diameter Gambel oak, or other resprouting species [for example, New Mexico locust, Rocky Mountain maple (Acer glabrum), and quaking aspen], at ground level with a chainsaw or increment borer. We used a targeted sampling approach to collect fire-scarred logs, stumps, or trees within and on the edges of the shrubfield to obtain a complete inventory of all widespread fires for the site over the maximum time period encompassed by the tree-ring material (Farris and others 2013). For the death-date line of evidence, we targeted all potentially fire-killed trees, especially those with well-preserved outside (bark) surfaces. We cored living trees with potential fire-related crown damage in and along the edges of shrubfields to assess for tree-ring growth changes or anomalies related to disturbance.
Laboratory Procedures
We employed standard dendrochronological procedures (Speer 2010) to prepare and crossdate tree-ring samples. We determined annually accurate and precise dates for regeneration (pith) dates, fire scars, death dates, and growth changes. We looked for and dated tree-ring growth changes across all of our samples, not just those trees cored as suspected growth change trees. Evidence for growth changes included growth suppression or release, multiple missing rings, and the presence of traumatic resin ducts (Arbellay and others 2014). We assessed whether these events could be climate-related by comparing the growth patterns to nearby climatically sensitive tree-ring chronologies at Cat Mesa, Fenton Lake, and Mesa Alta (Swetnam and Lynch 1993; Guiterman and others 2016). If severe growth anomalies were not correlated with climate variability, we attributed synchronous growth changes among trees (and with other lines of fire evidence) to be a response to partial crown or root mortality from high-intensity fire (Guiterman and others 2015).
Analyses
The four lines of evidence were compiled for each site in order to compare the reconstructed fire history, timing of tree mortality, and growth changes to major pulses in regeneration of oak and other sprouting species. Fire history data were analyzed in the R statistical computing environment (R Core Team 2015) using the burnr library (Malevich and others 2016). We calculated fire interval statistics on composite fire chronologies by filtering for fire events recorded by a minimum of two and at least 10% of fire-scarred trees. Overall analysis of fire history was limited to the historical period 1664–1899 when four of five sites were recording fire, but prior to cessation of the historical fire regime after 1899.
Shrubfield fire regimes (fire frequency and fire–climate relationships) were compared to fire regimes of conifer-dominated sites in the Jemez (Touchan and others 1996; Swetnam and others 2016). By filtering the 60 available Jemez conifer fire history sites in the same manner as the shrubfield sites, and only using sites with at least ten widespread fires, we narrowed the overall sample to 39 Jemez conifer-dominated sites. We used superposed epoch analysis (SEA) to compare fire–climate relationships between the conifer and shrubfield fire regimes (Swetnam 1993). Climate data for the SEA included tree-ring reconstructed summer (June through August) Palmer Drought Severity Index (PDSI; Cook and Krusic 2004: gridpoint 133). Although this tree-ring-derived PDSI reconstruction is for summer, it primarily reflects variability in cool-season moisture in the region (St. George and others 2010).
Results
We identified 349 Gambel oak shrubfields larger than 5 ha in the Jemez Mountains, totaling 10,809 ha (Figure 2). There were 21 shrubfields over 100 ha, accounting for roughly one-third of the total shrubfield area. The median shrubfield size was 17 ha and the largest was 368 ha. Gambel oak shrubfields originating prior to recent (1996–2011) fires made up 4.75% of the total dry conifer/shrub area.
Principal components analysis revealed broad similarities in the physiographic and climate spaces of Gambel oak shrubfields and dry conifer forests across the Jemez study area (Figure 3). Our observations in the field support this result, as we detected no obvious physical factor that distinguished shrubfield from conifer forest sites. The first principal component explained 62% of total variance, relating strongly to elevation, precipitation, and temperature. A general lack of shrubfields at the highest elevations of dry conifer forests may relate to increased competition with species such as aspen in mesic environments. Topographic factors including TRMI, TPI, slope, and curvature were important in PC2, with no discernable difference between vegetation types. Although the distribution of Gambel oak shrubfields may be influenced by a variety of edaphic factors that we may have failed to measure or account for, Gambel oak is known to occur across a broad range of environments (Rehfeldt and others 2006; Kaufmann and others 2016).

Principal components analysis of Gambel oak shrubfields versus dry conifer forest in the Jemez Mountains. Comparisons of 10,000 random points within each geographic space (see Figure 2) reveals broadly similar settings for shrubfields and forests, with shrubfields occupying the lower two-thirds of elevation and climate space (PC1). There is no discernible difference between shrubfields and forests by topographic position or physiography (PC2).
At the five tree-ring sites, we collected 336 samples from 301 trees of 11 species, and successfully crossdated samples from 281 trees, or 93% (Figure 2, Table 1). Shrubfield age structures were derived from 158 trees in 72 plots. The majority (85%) of species sampled in age structure plots were Gambel oak, reflecting oak dominance in the shrubfields (Figure 4). Shrub age structures show a common pattern of synchronous recruitment in the late 1890s, with 1899 being the primary cohort initiation year at all but one site, Señorito North (SNN), where oak sprouted over a 7-year period (1894–1900) without evidence for a cause of initiation (no fire, frost, or logging). The relatively old age of the Gambel oak shrubfields demonstrates that they can dominate a site for more than 115 years, even in the absence of recurrent fire disturbance. Indeed, we observed little change in shrubfield area and minimal conifer recruitment within shrubfields in comparisons with aerial imagery from 1935 to 2010 (Figure 5).

Age structure of Gambel oak shrubfields with additional lines of tree-ring fire evidence. Species include PIPO, ponderosa pine; POTR, quaking aspen; QUGA, Gambel oak; RONE, New Mexico locust. Site abbreviations are defined in Table 1.

Aerial photo-comparisons for two shrubfield sites with the most conifer recruitment since 1935.
The sprouting dates of Gambel oak cohorts shown in Figure 4 are generally synchronous with widespread fire events recorded by fire-scarred trees in shrubfields and locally, but most sites lack further evidence of higher severity fire (for example, death dates or growth changes). One exception, RDC, shows all four lines of evidence in the year 1899, indicating a fire event that killed and damaged conifers. This fire, however, did not burn at high severity across the site (76 ha), as the evidences for mortality and growth change were spatially separated, multiple oaks survived the fire, and multiple trees—including oaks—were scarred rather than killed. Another instance of synchrony in multiple lines of evidence is at Señorito South (SNS) in 1870, when growth changes and fire scars were recorded on four trees, and two trees had 1869 death dates (that is, mortality prior to the 1870 growing season). There is no oak cohort associated with this event nor are there any older oaks at the site, although one tree sprouted in 1873, after the next widespread fire event. It is plausible that this fire could have caused some conifer mortality and led to the establishment of part or all of the shrubfield, but this could not be confirmed from our data.
The tree-ring fire histories of Gambel oak shrubfields show frequent fire throughout the historical period (Figure 6). We documented fire events at these sites as early as 1250 CE (at SNN), with the majority of fire scars dating from 1664 to 1899. Median fire intervals at shrubfield sites ranged from 9 to 13 years during the 1664–1899 period (Table 2). Our sampling was sensitive enough to record recent fires documented in the local fire atlas, and an undocumented fire in 1989 across the upper portion of San Pablo Canyon (SPC).

Fire histories of Gambel oak shrubfields. Each colored horizontal line represents the sampled life span of an individual tree (n = 100), with pith and bark years indicated by dots. Solid line segments indicate that the tree is recording fires. Vertical tick marks show fire scars (n = 388) and triangles show growth changes on fire scar trees. At the bottom (“comp”) are filtered composite fire histories for each site. Species include JUSC, Rocky Mountain juniper; PIPO, ponderosa pine; PIST, southwestern white pine; POTR, quaking aspen; QUGA, Gambel oak. Site abbreviations are defined in Table 1 (Color figure online).
Climate variables affecting historical fire occurrence were similar among the shrubfield sites and 39 dry conifer sites in the study area (Figure 7). SEA results indicate that single-year drought was associated with fire occurrence (p < 0.01) at all shrubfield sites and 36 dry conifer sites. In addition, there was generally (sometimes significantly) above average cool-season moisture 1–3 years prior to the year of fire at both the shrub and conifer sites that related to fire occurrence.

Superposed epoch analyses for 39 dry conifer sites (box-and-whisker plots) and five Gambel oak shrubfield sites (triangles). The lag-year zero is the year of fire. Numbers in parentheses indicate the number of sites with a mean PDSI departure beyond the 95% bootstrapped confidence limit for that lag year. For visual aid, we show the approximate bootstrapped confidence limits used in each of the 44 SEA runs depicted here, with dashed and dotted lines for the 95 and 99% limits, respectively. These lines were based on a SEA of 1000 iterations with 19 random event years. There are 13 fire events included at MPB, 16 at RDC, 25 at SNN, 21 at SNS, and 18 at SPC.
Discussion
Historical Conditions and Possible Origins of Gambel Oak Shrubfields
We found that Gambel oak shrubfields historically maintained a frequent surface fire regime for hundreds of years until it collapsed after 1899 due to livestock grazing (Touchan and others 1996; Allen 2007). Moreover, the shrubfields burned at the same general rates and under similar climate patterns as the highly fire-prone local dry conifer forests in which they are embedded. Relatively wet, antecedent climate conditions associated with fire suggest that short-term fine-fuel production was important in supporting fire regimes in both vegetation types (that is, a fuel-limited system; Figure 7). The evidence of historical surface fuel limitation in oak shrubfields of the Jemez Mountains suggests a distinct historical fuel structure, with a higher proportion of grass and possibly more widely spaced and shorter oak ramets. A more open structure with herbaceous surface fuels could have sustained high fire frequencies, but we also found evidence of historical high-intensity fires, likely supported by the shrubs as fuels. Growth changes on fire-surviving conifers in four of five shrubfields included 3–11 absent rings in the decade or more following some fires (Figure 4). These growth suppressions could have resulted from crown and/or root scorch by intense fire and are not commonly observed on tree-ring samples from forests with frequent-fire regimes that lack shrubs. More work is needed to understand the potential for relatively frequent, but sometimes intense fire regimes in Gambel oak shrubfields.
We used multiple lines of dendroecological evidence to attempt to reconstruct the timing and scale of high-severity fire events that likely initially led to the dominance of Gambel oak at our shrubfield sites. Our results, however, did not match the broader-scale tree-ring fingerprint of high-severity fires in pine–oak that we expected (for example, Guiterman and others 2015). Across the shrubfield sites we sampled, there is no clear evidence of widespread high-severity fire events with high rates of conifer mortality in the period of reconstruction (1664–1899). Instead we found evidence that points to long-term maintenance of the oak-dominated shrubfields through recurrent fire activity. Of the 100 fire-scarred trees we sampled, 45 ponderosa pines recorded fire events and survived for centuries within shrubfields, and 32 oak stems were scarred rather than killed by fires in the late nineteenth century (Figure 6). Thus, it seems unlikely that high-severity fires during the last 350 years initiated the largest pre-1900 shrubfields (> 75 ha) still present in the Jemez Mountains. This supports the long-held understanding that low-severity surface fires predominated during the historical period in ponderosa pine and dry mixed-conifer forests in the Jemez Mountains (Touchan and others 1996) and in many other nearby areas (Swetnam and Baisan 1996; Grissino-Mayer and others 2004; Margolis and Balmat 2009; Bigio and others 2016; Margolis and Malevich 2016). High-severity patches were indeed a component of historical fire regimes in these forest types, but tree-ring-based evidence from the Jemez and similar areas in northern Arizona and New Mexico and southern Colorado demonstrates their relative rarity and constrains patch sizes to less than 100 ha (Margolis and Balmat 2009; Huffman and others 2015; Bigio and others 2016; Margolis and Malevich 2016).
If the large Gambel oak shrubfields we sampled did not originate from high-severity fire in the last 350 years, when and how did they originate? We speculate that it is plausible that shrubfields originated from undocumented (perhaps undocumentable) earlier fires of unknown antiquity. Our tree-ring methods were limited in temporal reach by the frequent burning of shrubfields before 1899 that would have consumed evidence of earlier tree-killing events (that is, death dates on logs or snags). Resprouting Gambel oak clones can almost certainly persist for many centuries (like aspen, Margolis and others 2007), if not much longer (like coppiced oaks in Spain actively resprouting for > 500 years; Salomón and others 2016), making it possible that their origins are pre-historic. Perhaps the numerous and diverse oak shrubfields found in the Jemez Mountains (Figure 2), as well as regionally across the Four Corners states (Arizona, New Mexico, Colorado, and Utah), cumulatively originated from a spectrum of fire sizes and severities over the Holocene, perhaps in pulses during climatic windows that favored more high-severity fires in oak-potential settings. Humans also may have played a role in these dynamics because some tribes inhabited mountain areas such as the Jemez at relatively high population densities for centuries prior to the Spanish arrival, with discernible impacts on forest structures and fire regimes (Liebmann and others 2016; Swetnam and others 2016). Further research that utilizes other paleoecological proxies, such as charcoal (Allen and others 2008) or alluvial sediments (Roos 2008; Fitch and Meyer 2014), may provide deeper-time insights into the timing of the events and the potential for human–fire–climate interactions that could have led to Gambel oak dominance over conifers in some landscape settings.
Dynamics of Gambel Oak Shrubfields as an Alternative Stable State
Gambel oak-dominated shrubfields in the Jemez Mountains share the environmental conditions of the local dry conifer forests in which they are embedded. These similarities indicate the potential for a given site to switch between dominant vegetation types and implicate disturbance as a key process in the origins and maintenance of these different vegetative states (Petraitis and Latham 1999). Indeed, recent high-severity fires have triggered many instances of rapid forest transitions in the Jemez Mountains and across the southwest region, and our study sites may have similar origins, as discussed above. The shrubfields we sampled have persisted for well over a generation of Gambel oak, with centuries of recurrent fires apparently maintaining their general configuration through resprouting response. Even in the absence of fire disturbances, oak has been the dominant vegetation for more than 115 years, underscoring shrubfield resilience to variation in climate and disturbance regimes. Thus, our findings do not support an interpretation that oak shrubfields are a seral stage in dry conifer forest ecosystems, but rather support the hypothesis that large (> 75 ha) Gambel oak-dominated shrubfields, at least in the Jemez Mountains, are an alternative state to dry conifer forest.
Dry conifer forests of the interior western USA (Falk and others 2011), and the Jemez Mountains in particular (Touchan and others 1996; Swetnam and others 2016), historically were subject to relatively frequent surface fire regimes. At landscape scales, these recurrent and extensive low-severity fires stabilized pre-1900 forest stand structures by maintaining low surface fuel accumulations and elevated canopies (Allen and others 2002), reducing the probability of stand-replacing crown fire behavior. As illustrated in our conceptual model of pre-1900 dry pine–oak forest interactions (Figure 8), combinations of high levels of shade from overstory conifers, along with frequent surface fire consumption of small-sized oak resprouts, probably limited the growth and spread of shade-intolerant oaks and other shrubs (Figure 8A; Abella and Fulé 2008; Van Auken and Bush 2017). Thus, under historical frequent-fire regimes and forest stand structures, the potential for fire-induced state transitions from dry conifer forest to shrubfield dominance was probably quite low in most landscape settings.

Conceptual model of alternative stable states in dry conifer–shrub ecosystems. With an active surface fire regime, dry conifer forests remain stable, and conifer seedlings and light-suppressed shrubs are present in the understory (A). Anthropogenic drivers, including fire exclusion that increases forest fuels and climate change leading to greater aridity, push the stand past a tipping point where extensive high-severity fire can trigger a rapid transition (large black arrow). This post-fire state includes shrubs that resprout from surviving roots, fire-killed mature conifers, and potentially, episodic scattered conifer seedling recruitment (B). Once in this highly altered state, there are multiple process pathways that can maintain a variety of shrub-dominated structures (C–E). A lack of fire will allow some of the conifer seedlings to survive amidst the competition from closing shrub canopies, but for decades the conifer seedlings are at high risk of mortality from drought and/or fire (C). These disturbances also foster regeneration and spread of the resprouting clonal shrubs, further reinforcing and stabilizing shrub dominance (D). Climate change is likely to favor shrubs over conifers in many areas, including through the process of more extensive and severe fires. These added pressures will maintain the shrubfield state, with fewer chances for conifer regeneration and survival (E).
Now, because of human-caused forest structural changes and climate stressors, greater proportions of interior western US landscapes are susceptible to ecosystem tipping points initiated by a change to large patch size, high-severity fires (Falk 2013; Allen 2016; Johnstone and others 2016; Miller and Safford 2017). Whether land-use changes have also led to widespread increases in Gambel oak and other shrubs is unclear (Kaufmann and others 2016), but disturbances associated with human and livestock activities could have encouraged repeated resprouting, dispersal into new sites, and both incremental and episodic expansion of overall shrub coverage (Ryniker and others 2006). Following release from tree dominance by recent high-severity fires, extensive areas of dry conifer forests have transitioned to an altered, shrub-dominated state (Figure 8B). Our data and field observations indicate potential for several different future shrubfield structures (Figure 8C–E), all of which are shrub-dominated and none of which have a clear pathway to return to conifer dominance, especially given likely future climate across the region (as in Coop and others 2016).
At our shrubfield sites, Gambel oak dominance persists after more than 115 years in the absence of fires. Conifer trees are present in the shrubfields, as they have been for centuries, but even favorable climate windows for tree regeneration, such as the 1910s (Savage and others 1996) or the late 1970s to early 1990s (Swetnam and Betancourt 1998), have resulted in little change to the composition of these oak-dominated shrub patches (Figure 5). Conifer seedlings and saplings are present in some areas, but in low densities, even along shrubfield margins where distance to seed sources are minimal. Other studies have found conifer seedlings established in shrubfields are concentrated within about 250 m from the forest edge, and at densities of 11–26 trees ha−1 up to 30 years post-fire (Haire and McGarigal 2010; Haffey 2014; Guiterman and others 2015). These young conifers are often highly vulnerable to mortality by competition, drought, and/or fire because of their delayed recruitment and initial slow growth rates (Feddema and others 2013; Savage and others 2013). Clonal resprouting shrubs have significant competitive advantages over conifer seedlings for space, light, and moisture (Clarke and others 2013), and ponderosa pine, in particular, has much reduced vigor in these circumstances (Shainsky and Radosevich 1986; Oliver 1990; Van Auken and Bush 2017). Therefore, even in the absence of fire or drought, few conifer seedlings are likely to survive to maturity (Figure 8C). We do note, however, that we found one exception to this trajectory. At the 292-ha SNN site, there is an 11-ha conifer island dominated by white fir that established in the 1940s and eventually overtopped and suppressed mature Gambel oak stems (Figure 5, bottom-right image). This occurrence of successful conifer establishment is in accord with the conclusions of Van Auken and Bush (2017), but we emphasize its relative rarity in the Jemez Mountains. The conifer patch is localized and especially mesic and has not burned since the 1890s. In addition, the shade tolerance of white fir likely favored its establishment under shrub cover, whereas ponderosa pine is shade intolerant and may be less likely to regenerate under shrub cover (Van Auken and Bush 2017). Without fire suppression, a twentieth-century fire would have likely killed all or most of the fire-intolerant white fir and regenerated oak sprouts, maintaining the small patch in its historical, oak-dominated configuration.
The effects of fire in a shrubfield are to reset oak dominance through rapid and vigorous post-fire resprouting, and to likely kill growth-suppressed conifers (Figure 8D). Recurrent burns of young shrubfields could be fatal to surviving or regenerating conifers given the potential for high-intensity shrub fires that may be exacerbated by woody fuels from fire-killed trees left by the initializing event (Odion and others 2010; Savage and others 2013; Paritsis and others 2015). Even intense fires, however, generally do little long-term damage to resprouting shrub composition, as root survival is relatively high (Clarke and others 2013; Minor and others 2017; although see Nicholson and others 2017).
Severe drought events may affect shrubfields in some similar ways to reburns by killing many young regenerating conifers already competing for moisture (Figure 8D; Savage and others 2013). Droughts can trigger canopy die-back in shrubs, but rarely exhaust carbohydrate stores in the roots, allowing for new sprouts (Choat and others 2012; Zeppel and others 2015). In the case of Gambel oak, live stems are both more resistant to low moisture levels and less susceptible to periodic drought-induced growth decline than ponderosa pine (Kolb and Stone 2000; Adams and Kolb 2005), giving them a substantial advantage in a future warmer, drier climate.
An example of the amplified effects of reburns and severe drought episodes occurred recently in the southeastern Jemez Mountains. The 2011 Las Conchas Fire burned at high severity through a mosaic of persistent dry conifer stands and shrubfields of oak and locust that emerged after a series of fires since 1977. The Las Conchas Fire led to the transition of 86% of the pre-fire forested area to a different vegetative configuration, with the largest proportion (38%) transitioning to oak shrubfield (Coop and others 2016). In the area of the largely high-severity 1996 Dome Fire, the Las Conchas Fire generated a huge (> 10,000 ha) conifer-depleted area that is now dominated by shrubs and ruderal vegetation (Allen 2016; Coop and others 2016). It is worth noting that this 10,000-ha area is more than 27 times the size of our largest mapped shrubfield (368 ha). Areas already oak-dominated prior to the Las Conchas Fire showed the greatest resilience to reburn (Coop and others 2016).
Warmer, drier climate conditions are projected for the southwest USA in coming decades, which would increase the frequency, severity, and duration of regional drought episodes (Cook and others 2015) and associated forest drought stress and fire activity (Williams and others 2013). High rates of overstory conifer mortality, already attributable to increasing drought and heat stress in many areas of the interior western USA (McDowell and others 2016) and globally (Allen and others 2010), portend future widespread forest die-offs. Hotter droughts are also likely to drive increases in the numbers and sizes of fires, with increases in the proportion of fires burning at moderate to high severity (Williams and others 2013; Abatzoglou and Williams 2016; Westerling 2016; Miller and Safford 2017). This climate trajectory and associated intensified disturbance processes in the southwest USA would severely hamper conifer recruitment on many sites (Rother and others 2015; Rother and Veblen 2016). All of these trends increasingly favor Gambel oak over co-occurring dry conifer forests in the southwest, driving recent and anticipated future transitions of greater proportions of historically forested landscapes in the Jemez Mountains and elsewhere regionally toward dominance by relatively resilient shrubfields (Figure 8E).
Conclusions
Critical questions facing managers of post-fire landscapes in the southwest are whether conifers will return to high-severity burn areas dominated by shrub species, and if so, how long will that take? Or will shrubfields persist, representing a long-term type conversion from forest to shrub dominance? Our results demonstrate that Gambel oak-dominated shrubfields larger than 75 ha in the Jemez Mountains, New Mexico, can survive in their current configurations for more than 115 years without fire in the midst of dry conifer landscapes. Historical aerial imagery comparisons at our sites indicate that only isolated conifer recruitment has occurred and that shrubfield extents have not changed substantially since at least 1935. Tree-ring fire history reconstructions within shrubfields show that they historically (1664–1899) burned as frequently (every 9–13 years) and under similar climate conditions as the fire-prone dry conifer forests in which they are embedded. Resprouting clonal shrubs like Gambel oak have post-fire competitive growth advantages relative to co-occurring seed-based conifer regeneration, and resprouter resilience to drought and fire likely stabilizes shrubfield ecosystems over centuries, and potentially longer (for example, Strom and Fulé 2007). We therefore conclude that large patches of forest-to-oak transitions following high-severity fires are not a seral stage to conifer forest in these mountains, but rather represent metastable landscape features. Consequently, further expansion and long-term persistence of Gambel oak-dominated shrubfields in the southwestern USA can be expected with warmer, drier future climates.
We found that the ecological processes by which Gambel oak shrubfields in the Jemez Mountains previously formed have either changed or dramatically intensified. Up through the late twentieth century, shrubfields made up a relatively small proportion of the Jemez dry conifer landscape (4.75%), with sizes and a spatial distribution that suggest disturbances (probably fire) played an important role in their origins and maintenance. Recent high-severity fire patches in the Jemez Mountains now largely dominated by shrubs are almost 30 times larger than the most extensive late twentieth-century shrubfield that we identified (Allen 2016). This increase is consistent with our understanding of the combined effects of past land management (for example, fire suppression) and amplified drought with climate change on forest and fuel structures and flammability.
Similar recent forest-to-shrub type conversions across the southwestern USA suggest an ongoing adjustment of conifer forests to new bioclimatic and fire regimes (Turner 2010; Johnstone and others 2016). Such rapid rates of widespread forest transformation to shrub-dominated states, fueled by historical land use, will greatly affect ecosystem functioning and services (Hurteau and others 2014). This underscores the need to restore vital ecosystem processes such as surface fire to enhance the resilience of dry conifer forests to severe disturbance and climate change (Stephens and others 2016) and to maintain the structural and species diversity of dry conifer/shrub ecosystems.
References
Abatzoglou JT, Williams AP. 2016. The impact of anthropogenic climate change on wildfire across western US forests. Proc Natl Acad Sci 113:11770–5.
Abella S. 2008. Managing Gambel oak in southwestern ponderosa pine forests: the status of our knowledge. Gen. Tech. Rep. RMRS-GTR-218. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. p 27.
Abella SR, Fulé PZ. 2008. Changes in Gambel oak densities in southwestern Ponderosa Pine Forests since Euro-American Settlement. Res. Note RMRS-RN-36. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. p 6.
Adams HD, Kolb TE. 2005. Tree growth response to drought and temperature in a mountain landscape in northern Arizona, USA. J Biogeogr 32:1629–40.
Airey Lauvaux C, Skinner CN, Taylor AH. 2016. High severity fire and mixed conifer forest-chaparral dynamics in the southern Cascade Range, USA. For Ecol Manage 363:74–85.
Allen CD. 2007. Interactions Across spatial scales among forest dieback, fire, and erosion in Northern New Mexico landscapes. Ecosystems 10:797–808.
Allen CD. 2016. Forest ecosystem reorganization underway in the Southwestern US: a preview of widespread forest changes in the anthropocene? In: Sample VA, Bixler RP, Miller C, Eds. Forest conservation and management in the anthropocene: adaptation of science, policy and practices. Boulder (CO): University Press of Colorado.
Allen CD, Anderson RS, Jass RB, Toney JL, Baisan CH. 2008. Paired charcoal and tree-ring records of high-frequency Holocene fire from two New Mexico bog sites. Int J Wildland Fire 17:115–30.
Allen CD, Macalady AK, Chenchouni H, Bachelet D, McDowell N, Vennetier M, Kitzberger T, Rigling A, Breshears DD, Hogg EH, Gonzales P, Fensham R, Zhang Z, Castro J, Demidova N, Lim J-H, Allard G, Running SW, Semerci A, Cobb N. 2010. A global overview of drought and heat-induced tree mortality reveals emerging climate change risks for forests. For Ecol Manage 259:660–84.
Allen CD, Savage M, Falk DA, Suckling KF, Swetnam TW, Schulke T, Stacey PB, Morgan P, Hoffman M, Klingel JT. 2002. Ecological restoration of southwestern ponderosa pine ecosystems: a broad perspective. Ecol Appl 12:1418–33.
Arbellay E, Stoffel M, Sutherland EK, Smith KT, Falk DA. 2014. Resin duct size and density as ecophysiological traits in fire scars of Pseudotsuga menziesii and Larix occidentalis. Ann Bot 114:973–80.
Beers TW, Dress PE, Wensel LC. 1966. Notes and observations: aspect transformation in site productivity research. J For 64:691–2.
Bigio ER, Swetnam TW, Baisan CH. 2016. Local-scale and regional climate controls on historical fire regimes in the San Juan Mountains, Colorado. For Ecol Manage 360:311–22.
Chambers ME, Fornwalt PJ, Malone SL, Battaglia MA. 2016. Patterns of conifer regeneration following high severity wildfire in ponderosa pine—dominated forests of the Colorado Front Range. For Ecol Manage 378:57–67.
Choat B, Jansen S, Brodribb TJ, Cochard H, Delzon S, Bhaskar R, Bucci SJ, Feild TS, Gleason SM, Hacke UG, Jacobsen AL, Lens F, Maherali H, Martínez-Vilalta J, Mayr S, Mencuccini M, Mitchell PJ, Nardini A, Pittermann J, Pratt RB, Sperry JS, Westoby M, Wright IJ, Zanne AE. 2012. Global convergence in the vulnerability of forests to drought. Nature 491:752–5.
Clarke PJ, Lawes MJ, Midgley JJ, Lamont BB, Ojeda F, Burrows GE, Enright NJ, Knox KJE. 2013. Resprouting as a key functional trait: how buds, protection and resources drive persistence after fire. New Phytol 197:19–35.
Clary WP, Tiedemann AR. 1992. Ecology and values of Gambel oak woodlands. In: Ffolliott PF, Gottfried GJ, Bennett DA, Hernandez CVM, Ortega-Rubio A, Hamre RH, Eds. Ecology and management of oaks and associated woodlands: perspectives in the southwestern United States and northern Mexico; 1992 April 27–30. Sierra Vista, AZ: Gen. Tech. Rep. RM-218. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. pp 87–95.
Collins BM, Roller GB. 2013. Early forest dynamics in stand-replacing fire patches in the northern Sierra Nevada, California, USA. Landsc Ecol 28:1801–13.
Connell J, Sousa W. 1983. On the evidence needed to judge ecological stability or persistence. Am Nat 121:789–824.
Cook BI, Ault TR, Smerdon JE. 2015. Unprecedented 21st century drought risk in the American Southwest and Central Plains. Sci Adv 1:1–7.
Cook ER, Krusic PJ. 2004. The North American Drought Atlas. (Lamont–Doherty Earth Obs Natl Sci Found New York) Available at http://iridl.ldeo.columbia.edu/SOURCES/.LDEO/.TRL/.NADA2004/.pdsi-atlas.html.
Coop JD, Parks SA, McClernan SR, Holsinger LM. 2016. Influences of prior wildfires on vegetation response to subsequent fire in a reburned southwestern landscape. Ecol Appl 26:346–54.
Daly C, Halbleib M, Smith JI, Gibson WP, Doggett MK, Taylor GH, Curtis J, Pasteris P. 2008. Physiographically sensitive mapping of climatological temperature and precipitation across the conterminous United States. Int J Climatol 28:2031–64.
Falk D. 2013. Are Madrean ecosystems approaching tipping points? Anticipating interactions of landscape disturbance and climate change. In: Gottfried G, Ffolliott P, Gebow B, Eskew L, Collins L, Eds. Merging science and management in a rapidly changing world: biodiversity and management of the Madrean Archipelago III; 2012 May 1–5; Tucson, AZ. Proceedings. RMRS-P-67. Fort Collins, CO. U.S. Department of Agriculture, Forest Service, Rocky Mountain Research Station. pp 40–7.
Falk DA, Heyerdahl EK, Brown PM, Farris C, Fulé PZ, McKenzie D, Swetnam TW, Taylor AH, Van Horne ML. 2011. Multi-scale controls of historical forest-fire regimes: new insights from fire-scar networks. Front Ecol Environ 9:446–54.
Farris C, Baisan C, Falk D, Van Horne M, Fule P, Swetnam T. 2013. A comparison of targeted and systematic fire-scar sampling for estimating historical fire frequency in south-western ponderosa pine forests. Int J Wildland Fire 22:1021–33.
Feddema J, Mast J, Savage M. 2013. Modeling high-severity fire, drought and climate change impacts on ponderosa pine regeneration. Ecol Modell 253:56–69.
Fitch EP, Meyer GA. 2014. Temporal and spatial climatic controls on Holocene fire-related erosion and sedimentation, Jemez Mountains, New Mexico. Quat Res (United States) 85:75–86.
Floyd M, Romme W, Hanna D. 2000. Fire history and vegetation pattern in Mesa Verde national Park, Colorado, USA. Ecol Appl 10:1666–80.
Fulé P, Covington W, Moore M. 1997. Determining reference conditions for ecosystem management of southwestern ponderosa pine forests. Ecol Appl 7:895–908.
Grissino-Mayer H, Romme W, Floyd M, Hanna D. 2004. Climatic and human influences on fire regimes of the southern San Juan Mountains, Colorado, USA. Ecology 85:1708–24.
Guiterman CH, Margolis EQ, Swetnam TW. 2015. Dendroecological methods for reconstructing high severity fire in pine-oak forests. Tree-Ring Res 71:67–77.
Guiterman CH, Swetnam TW, Dean JS. 2016. Eleventh-century shift in timber procurement areas for the great houses of Chaco Canyon. Proc Natl Acad Sci 113:1186–90.
Haffey CM. 2014. Patterns and predictors of crown fire induced type conversion in dry conifer forests. M.S. Thesis, Northern Arizona University, Flagstaff, AZ. p 91.
Haire S, McGarigal K. 2008. Inhabitants of landscape scars: succession of woody plants after large, severe forest fires in Arizona and New Mexico. Southwest Nat 53:146–61.
Haire SL, McGarigal K. 2010. Effects of landscape patterns of fire severity on regenerating ponderosa pine forests (Pinus ponderosa) in New Mexico and Arizona, USA. Landsc Ecol 25:1055–69.
Hanks JP, Dick-Peddie WA. 1974. Vegetation patterns of the White Mountains, New Mexico. Southwest Nat 18:371–82.
Harper KT, Wagstaff FJ, Kunzler LM. 1985. Biology and management of the Gambel oak vegetative type: a literative review. General technical report/Intermountain Forest and Range Experiment Station. USDA (no. INT-179). p 34.
Huffman D, Zegler T, Fule P. 2015. Fire history of a mixed conifer forest on the Mogollon Rim, northern Arizona, USA. Int J Wildland Fire 24:680–9.
Hurteau MD, Bradford JB, Fule PZ, Taylor AH, Martin KL. 2014. Climate change, fire management, and ecological services in the southwestern US. For Ecol Manage 327:280–9.
Iniguez J, Swetnam T, Baisan C. 2009. Spatially and temporally variable fire regime on Rincon Peak, Arizona, USA. Fire Ecol 5:3–21.
Jenness J. 2006. Topographic Position Index (tpi_jen.avx) extension for ArcView 3.x, v. 1.2. http://www.jennessent.com/arcview/tpi.htm.
Johnstone JF, Allen CD, Franklin JF, Frelich LE, Harvey BJ, Higuera PE, Mack MC, Meentemeyer RK, Metz MR, Perry GLW, Schoennagel T, Turner MG. 2016. Changing disturbance regimes, ecological memory, and forest resilience. Front Ecol Environ 14:369–78.
Kashian DM, Romme WH, Regan CM. 2007. Reconciling divergent interpretations of quaking aspen decline on the northern Colorado Front Range. Ecol Appl 17:1296–311.
Kaufmann MR, Huisjen DW, Kitchen S, Babler M, Abella SR, Gardiner TS, McAvoy D, Howie J, Page DHJ. 2016. Gambel oak ecology and management in the southern rockies: the status of our knowledge. Southern Rockies Fire Science Network Publication 2016-1.
Kolb TE, Stone JE. 2000. Differences in leaf gas exchange and water relations among species and tree sizes in an Arizona pine-oak forest. Tree Physiol 20:1–12.
Liebmann MJ, Farella J, Roos CI, Stack A, Martini S, Swetnam TW. 2016. Native American depopulation, reforestation, and fire regimes in the Southwest U.S., 1492-1900 C.E. Proc Natl Acad Sci 113:E697–704.
Little EL. 1971. Atlas of United States trees, volume 1, conifers and important hardwoods. U.S. Department of Agriculture Miscellaneous Publication 1146. p 9, 200 maps.
Lowry JHJ, Ramsey RD, Thomas KA, Schrupp D, Kepner W, Sajwaj T et al. 2007. Land cover classification and mapping. In: Prior-Magee JS, Boykin K., Bradford DF, Kepner WG, Lowry JH, Schrupp DL, Thomas KA, Thompson BC, Eds. Southwest Regional Gap Analysis Final Report. Moscow (ID): Southwest regional gap analysis final report.
Malevich S, Margolis E, Guiterman C. 2016. burnr: fire history analysis in R https://github.com/ltrr-arizona-edu/burnr.
Margolis EQ, Balmat J. 2009. Fire history and fire–climate relationships along a fire regime gradient in the Santa Fe Municipal Watershed, NM, USA. For Ecol Manage 258:2416–30.
Margolis EQ, Malevich SB. 2016. Historical dominance of low-severity fire in dry and wet mixed-conifer forest habitats of the endangered terrestrial Jemez Mountains salamander (Plethodon neomexicanus). For Ecol Manage 375:12–26.
Margolis EQ, Swetnam TW, Allen CD. 2007. A stand-replacing fire history in upper montane forests of the southern Rocky Mountains. Can J For Res 37:2227–41.
Margolis EQ, Woodhouse CA, Swetnam TW. 2017. Drought, multi-seasonal climate, and wildfire in northern New Mexico. Clim Change 142:433–46.
McDowell NG, Williams AP, Xu C, Pockman WT, Dickman LT, Sevanto S, Pangle RE, Limousin J-M, Plaut JA, Mackay DS, Ogée J, Domec JC, Allen CD, Fisher RA, Jiang X, Muss JD, Breshears DD, Rauscher SA, Koven C. 2016. Multi-scale predictions of massive conifer mortality due to chronic temperature rise. Nat Clim Change 6:295–300.
Miller JD, Safford HD. 2017. Corroborating evidence of a pre-euro-American low-to moderate-severity fire regime in yellow pine–mixed conifer forests of the sierra Nevada, California, USA. Fire Ecol 13:58–90.
Minor J, Falk D, Barron-Gafford G. 2017. Fire severity and regeneration strategy influence shrub patch size and structure following disturbance. Forests 8:221.
Moir WH, Geils B, Benoit MA, Scurlock D. 1997. Ecology of Southwestern Ponderosa Pine Forests. In: Block WM, Finch, Deborah M, Eds. Songbird ecology in south-western ponderosa pine forests: a literature review. Gen. Tech. Rep. RM-292. Fort Collins, CO: U.S. Department of Agriculture, Forest Service, Rocky Mountain Forest and Range Experiment Station. pp 3–27.
Muldavin E, Kennedy A, Jackson C, Neville T, Neville P, Schultz K, Reid M. 2011. Vegetation classification and map: Bandelier National Monument.
Muldavin E, Tonne P. 2003. A vegetation survey and preliminary ecological assessment of Valles Caldera National Preserve, New Mexico.
Nagel TA, Taylor AH. 2005. Fire and persistence of montane chaparral in mixed conifer forest landscapes in the northern Sierra Nevada, Lake Tahoe Basin, California, USA. J Torrey Bot Soc 132:442–57.
Neilson RP, Wullstein LH. 1986. Microhabitat affinities of Gambel oak seedlings. Gt Basin Nat 46:294–8.
Nicholson Á, Prior LD, Perry GLW, Bowman DMJS. 2017. High post-fire mortality of resprouting woody plants in Tasmanian Mediterranean-type vegetation. Int J Wildland Fire 26:532–7.
Odion DC, Moritz MA, Dellasala DA. 2010. Alternative community states maintained by fire in the Klamath Mountains, USA. J Ecol 98:96–105.
Oliver W. 1990. Spacing and shrub competition influence 20-year development of planted ponderosa pine. West J Appl For 5:79–82.
Paritsis J, Veblen TT, Holz A. 2015. Positive fire feedbacks contribute to shifts from Nothofagus pumilio forests to fire-prone shrublands in Patagonia. J Veg Sci 26:89–101.
Parker AJ. 1982. The topographic relative moisture index: an approach to soil-moisture assessment in mountain terrain. Phys Geogr 3:160–8.
Pausas JG, Pratt RB, Keeley JE, Jacobsen AL, Ramirez AR, Vilagrosa A, Paula S, Kaneakua-Pia IN, Davis SD. 2016. Towards understanding resprouting at the global scale. New Phytol 209:945–54.
Petraitis P, Latham R. 1999. The importance of scale in testing the origins of alternative community states. Ecology 80:429–42.
R Core Team. 2015. R: a language and environment for statistical computing.
Rehfeldt GE, Crookston NL, Warwell MV, Evans JS. 2006. Empirical analyses of plant-climate relationships for the western United States. Int J Plant Sci 167:1123–50.
Roccaforte J, Fule P, Chancellor W, Laughlin D. 2012. Woody debris and tree regeneration dynamics following severe wildfires in Arizona ponderosa pine forests. Can J For Res 42:593–604.
Romme WH, Floyd ML, Hanna D. 2009. Historical Range of variability and current landscape condition analysis: south central highlands section, southwestern Colorado and northwestern New Mexico. Fort Collins (CO): Colorado Forest Restoration Institute at Colorado State University and USDA Forest Service.
Roos CI. 2008. Fire, climate, and social-ecological systems in the ancient southwest: alluvial geoarchaeology and applied historical ecology. Ph.D. Thesis. University of Arizona.
Rother MT, Veblen TT. 2016. Limited conifer regeneration following wildfires in dry ponderosa pine forests of the Colorado Front Range. Ecosphere 7:e01594.
Rother MT, Veblen TT, Furman LG. 2015. A field experiment informs expected patterns of conifer regeneration after disturbance under changing climate conditions. Can J For Res 45:1607–16.
Ryniker KA, Bush JK, Van Auken OW. 2006. Structure of Quercus gambelii communities in the Lincoln National Forest, New Mexico, USA. For Ecol Manage 233:69–77.
Salomón R, Rodríguez-Calcerrada J, Zafra E, Morales-Molino C, Rodríguez-García A, González-Doncel I, Oleksyn J, Zytkowiak R, López R, Miranda JC, Gil L, Valbuena-Carabaña M. 2016. Unearthing the roots of degradation of Quercus pyrenaica coppices: A root-to-shoot imbalance caused by historical management? For Ecol Manage 363:200–11.
Savage M, Brown P, Feddema J. 1996. The role of climate in a pine forest regeneration pulse in the southwestern United States. Ecoscience 3:310–18.
Savage M, Mast JN. 2005. How resilient are southwestern ponderosa pine forests after crown fires? Can J For Res 977:967–77.
Savage M, Mast JN, Feddema JJ. 2013. Double whammy: high-severity fire and drought in ponderosa pine forests of the Southwest. Can J For Res 43:570–83.
Scheffer M, Carpenter S, Foley JA, Folke C, Walker B. 2001. Catastrophic shifts in ecosystems. Nature 413:591–6.
Shainsky L, Radosevich S. 1986. Growth and water relations of Pinus ponderosa seedlings in competitive regimes with Arctostaphylos patula seedlings. J Appl Ecol 23:957–66.
Sheppard P, Comrie A, Packin G, Angersbach K, Hughes M. 2002. The climate of the US Southwest. Clim Res 21:219–38.
Snyderman DA, Allen CD. 1997. Fire in the mountains: analysis of historical fires for Bandelier National Monument, Santa Fe National Forest and Surrounding Areas 1909 through 1996. Los Alamos, New Mexico.
Speer JH. 2010. Fundamentals of tree-ring research. Tucson: The University of Arizona Press.
St. George S, Meko DM, Cook ER. 2010. The seasonality of precipitation signals embedded within the North American Drought Atlas. The Holocene 20:983–8.
Stephens SL, Agee JK, Fulé PZ, North MP, Romme WH, Swetnam TW, Turner MG. 2013. Managing forests and fire in changing climates. Science 342:41–2.
Stephens SL, Burrows N, Buyantuyev A, Gray RW, Keane RE, Kubian R, Liu S, Seijo F, Shu L, Tolhurst KG, Van Wagtendonk JW. 2014. Temperate and boreal forest mega-fires: characteristics and challenges. Front Ecol Environ 12:115–22.
Stephens SL, Collins BM, Biber E, Fulé PZ. 2016. U.S. federal fire and forest policy: emphasizing resilience in dry forests. Ecosphere 7:1–19.
Strom BA, Fulé PZ. 2007. Pre-wildfire fuel treatments affect long-term ponderosa pine forest dynamics. Int J Wildland Fire 16:128.
Swetnam T. 1993. Fire history and climate change in giant sequoia groves. Science 262:885–9.
Swetnam T, Baisan C. 1996. Historical fire regime patterns in the southwestern United States since AD 1700. In: Allen CD, Ed. Fire effects in southwestern forests: Proceedings of the second La Mesa Fire Symposium. Los Alamos, New Mexico: USDA Forest Service General Technical Report RM-GTR-286. pp 11–32.
Swetnam TW, Betancourt JL. 1998. Mesoscale disturbance and ecological response to decadal climatic variability in the American Southwest. J Clim 11:3128–47.
Swetnam TW, Farella J, Roos CI, Liebmann MJ, Falk DA, Allen CD. 2016. Multi-scale perspectives of fire, climate and humans in western North America and the Jemez Mountains, USA. Philos Trans R Soc B 371:20150168.
Swetnam TW, Lynch AM. 1993. Multicentury, regional-scale patterns of western spruce budworm outbreaks. Ecol Monogr 63:399–424.
Touchan R, Allen C, Swetnam T. 1996. Fire history and climatic patterns in ponderosa pine and mixed-conifer forests of the Jemez Mountains, northern New Mexico. In: Allen CD, Ed. Fire effects in southwestern forests, Proceedings of the Second La Mesa Fire Symposium, Los Alamos, New Mexico, March 29–31, 1994. USDA Forest Service General Technical Report RM-GTR-286. pp 33–46.
Turner M. 2010. Disturbance and landscape dynamics in a changing world. Ecology 91:2833–49.
Van Auken OW. 2000. Shrub invasions of North American semiarid grasslands. Annu Rev Ecol Syst 31:197–215.
Van Auken OW, Bush JK. 2017. Succession in Quercus gambelii (Gambel’s Oak) woodlands. Am J Plant Sci 8:96–112.
Westerling AL. 2016. Increasing western US forest wildfire activity: sensitivity to changes in the timing of spring. Philos Trans R Soc B 371:20150178.
Williams AP, Allen CD, Macalady AK, Griffin D, Woodhouse CA, Meko DM, Swetnam TW, Rauscher SA, Seager R, Grissino-Mayer HD, Dean JS, Cook ER, Gangodagamage C, Cai M, McDowell NG. 2013. Temperature as a potent driver of regional forest drought stress and tree mortality. Nat Clim Change 3:292–7.
Zeppel MJB, Harrison SP, Adams HD, Kelley DI, Li G, Tissue DT, Dawson TE, Fensham R, Belinda E, Palmer A, West AG, Mcdowell NG. 2015. Drought and resprouting plants. New Phytol 206:583–589.
Acknowledgements
This study was funded by the Western Mountains Initiative and Bandelier National Monument (CESU Agreement #UADS-402), the US Geological Survey (Cooperative Agreement G13AC00247), and an EPA STAR Fellowship (#F13F51318 to C.H.G.). Additional support was provided by Bandelier National Monument, the USGS New Mexico Landscapes Field Station, and the Laboratory of Tree-Ring Research. We were aided in the field and lab by Eze Ahanonu, Chris Baisan, Andrew Bieg, John Danloe, Galen Gudenkauf, Collin Haffey, Nick Kessler, and Ben Olimpio. We thank Chris Baisan, Kay Beeley, Erica Bigio, and Collin Haffey for helpful discussions. Comments by Brandon Bestelmeyer, Jonathan Coop, and two anonymous reviewers helped to improve this manuscript. EPA has not officially endorsed this publication and the views expressed herein may not reflect the views of the EPA. Any use of trade, firm, or product names is for descriptive purposes only and does not imply endorsement by the US Government.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Guiterman, C.H., Margolis, E.Q., Allen, C.D. et al. Long-Term Persistence and Fire Resilience of Oak Shrubfields in Dry Conifer Forests of Northern New Mexico. Ecosystems 21, 943–959 (2018). https://doi.org/10.1007/s10021-017-0192-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-017-0192-2