
Abstract
Variability and future alterations in regional and global climate patterns may exert a strong control on the carbon dioxide (CO2) exchange of grassland ecosystems. We used 6 years of eddy-covariance measurements to evaluate the impacts of seasonal and inter-annual variations in environmental conditions on the net ecosystem CO2 exchange (NEE), gross ecosystem production (GEP), and ecosystem respiration (ER) of an intensively managed grassland in the humid temperate climate of southern Ireland. In all the years of the study period, considerable uptake of atmospheric CO2 occurred in this grassland with a narrow range in the annual NEE from −245 to −284 g C m−2 y−1, with the exception of 2008 in which the NEE reached −352 g C m−2 y−1. None of the measured environmental variables (air temperature (Ta), soil moisture, photosynthetically active radiation, vapor pressure deficit (VPD), precipitation (PPT), and so on) correlated with NEE on a seasonal or annual scale because of the equal responses from the component fluxes GEP and ER to variances in these variables. Pronounced reduction of summer PPT in two out of the six studied years correlated with decreases in both GEP and ER, but not with NEE. Thus, the stable annual NEE was primarily achieved through a strong coupling of ER and GEP on seasonal and annual scales. Limited inter-annual variations in Ta (±0.5°C) and generally sufficient soil moisture availability may have further favored a stable annual NEE. Monthly ecosystem carbon use efficiency (CUE; as the ratio of NEE:GEP) during the main growing season (April 1–September 30) was negatively correlated with temperature and VPD, but positively correlated with soil moisture, whereas the annual CUE correlated negatively with annual NEE. Thus, although drier and warmer summers may mildly reduce the uptake potential, the annual uptake of atmospheric CO2, in this intensively managed grassland, may be expected to continue even under predicted future climatic changes in the humid temperate climate region.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Grasslands cover about 40% of the Earth’s terrestrial ice-free area and constitute an important component in the Earth climate system and global carbon (C) balance, storing approximately one third of the global terrestrial C stock (Scurlock and Hall 1998; White and others 2000; IPCC 2007). In addition, grassland ecosystems are important ecological and socio-economical resources through their contribution to biodiversity and their use for food, forage, and livestock production (Sala and Paruelo 1997; White and others 2000). Thus, understanding grassland ecosystem dynamics is imperative to secure resource sustainability and to evaluate the role of grasslands in the global C cycle under future climate and management regimes.
Temperate grasslands are commonly considered to be close to C-neutral or a weak C sink, with mean C sequestration rates for North American and European grasslands ranging from 0.2 to 0.6 Mg C ha−1 y−1 (Jones and Donnelly 2004; Janssens and others 2005). In general, intensively managed grasslands may show enhanced uptake of atmospheric carbon dioxide (CO2) (Allard and others 2007; Gilmanov and others 2010). In a recent review of global grasslands, Gilmanov and others (2010) reported mean annual ecosystem CO2 uptake rates of 1.9 Mg C ha−1 y−1 for intensively managed grasslands, compared to 0.7 Mg C ha−1 y−1 for extensively managed grasslands. In addition to the net ecosystem CO2 exchange (NEE), however, the net C sequestration rate of managed grasslands is highly affected by additional C fluxes due to management activities. For European grasslands, the net biome production (NBP) adjusting NEE by additional C inputs (for example, manure, feed concentrate, and so on) and outputs (for example, biomass and milk export, C losses via dissolved organic C, methane (CH4), and respiration from cattle in stables) was recently estimated to average from 0.74 to 1.04 Mg C ha−1 y−1 on the farm scale (with the majority being intensively managed) (Soussana and others 2007; Ciais and others 2010), and at 101 Tg C y−1 on the European scale (Janssens and others 2003).
In the past, several studies have shown that the annual CO2 exchange of grasslands may vary in response to climate variability. Variations in precipitation (PPT) patterns, drought, and heat stress have been reported to result in prolonged suppression of both net primary and net ecosystem production (Heitschmidt and others 2005; Arnone and others 2008), and to cause a net release of CO2 from grassland to the atmosphere (Kim and others 1992; Flanagan and others 2002; Gilmanov and others 2006, 2007; Ma and others 2007; Nagy and others 2007; Zhang and others 2010). Thus, alterations in temperature and PPT regimes due to climatic changes may considerably affect the grassland C balance.
Variations in NEE result from differences in its two component fluxes: gross ecosystem production (GEP) and ecosystem respiration (ER). GEP is mainly controlled by the amount of photosynthetically active radiation (PAR), modified by effects from temperature and water availability, and further dependent on biotic controls such as vegetation biomass and leaf area, whereas ER is primarily driven by soil microbial activity and plant growth which themselves depend on temperature, moisture availability, and the supply of assimilates from photosynthetic activity (Bond-Lamberty and others 2004; Bahn and others 2008; Gilmanov and others 2010). Thus, ER is linked to GEP through its dependency on production and belowground allocation of photosynthetic assimilates. Considering these multiple controls, it is important to understand responses of the individual component fluxes GEP and ER to changes in environmental parameters to estimate the impacts of climatic conditions on the NEE.
To date, a limited number of multi-year (>5 years) studies of grassland ecosystem CO2 exchange exist, with the majority of them conducted in regions characterized by pronounced seasonal droughts and/or extensive grassland management. These multi-year studies altogether reported large inter-annual variability as well as the potential for the same site to exhibit annual net uptake and emission of CO2 depending on climatic conditions (Frank 2004; Gilmanov and others 2006; Ma and others 2007; Wohlfahrt and others 2008). Meanwhile, less information is available on the long-term dynamics of grassland CO2 exchange and response to climate variation in humid temperate regions. However, to evaluate the long-term CO2 exchange capacity of grassland ecosystems and to predict its potential modification under a changing climate, multi-year observations are necessary to separate natural inter-annual variability in climate and ecosystem CO2 exchange from the additional effects due to a changing climate.
In this study, we used 6 years of eddy-covariance (EC) data to determine the effects of environmental variables on the NEE, GEP, and ER of intensively managed grassland in the humid temperate climate of southern Ireland. Our primary objectives were (i) to determine the long-term annual CO2 uptake/emission potential and (ii) to identify the main environmental drivers of inter-annual variability in CO2 exchange.
Materials and Methods
Site Description
This study was conducted at the Dripsey grassland research station which is located about 25 km northwest of Cork City in southern Ireland (51°59′N; 8°45′W; 195 m above sea level). The regional 30-year normal for annual mean air temperature (Ta) and total PPT are 9.4°C and 1207 mm, respectively (Met Eireann, 1960–1990 climate norms at Cork Airport Meteorological Station). Although, throughout the study period, annual PPT measured at the Dripsey grassland exceeded the annual amount measured at the Cork Airport Station by approximately 150 mm. Growing season length may extend from mid-March to mid-October; however, we define the main growing season in this study as from April 1 to September 30 based on measurements and observations of grass growth and production rates. The prevailing wind direction (Wd) is from the southwest.
The grassland is classified as a high quality pasture and meadow, with perennial ryegrass (Lolium perenne L.) being the dominant plant species, mixed with smaller amounts of Meadow foxtail (Alopecurus pratensis L.) and Yorkshire-fog (Holcus lanatus L.). The soil type is a gleysol with loamy soil texture (42% sand, 41% silt, 17% clay) (Lawton and others 2006). The soil organic C (SOC) concentration of the 0–20 cm soil layer was estimated at 5.9% in 2004 (Byrne and others 2005). The plant available soil moisture content ranges from 0.12 m3 m−3 (wilting point) to 0.32 m3 m−3 (field capacity), with the saturation level being estimated at 0.46 m3 m−3. The leaf area index (LAI) ranges between 0.5 and 1.0 m2 m−2 during the autumn and winter months and reaches 2.0–2.5 m2 m−2 during the summer months, with maximum grass heights in the summer recording approximately 20 and 45 cm in pasture and meadow fields, respectively (Byrne and others 2005; Byrne and Kiely 2006).
The grassland surrounding the EC flux tower consists of a mosaic of individual paddocks (the EC footprint may include ~15–50 fields depending on sensor height and atmospheric conditions) which vary in size between 1 and 3 ha and belong to three (2004–2009) or eight different farmers (2003). Thin wire-and-post fences separate most of these paddocks and thus exert no major obstruction to the wind flow. These paddocks differ in their management regimes but are all intensively managed for either silage/hay (1 or 2 cuts per year, typically in June and August, with an annual yield of 7–10 t dry matter ha−1 y−1) or grazing of dairy and beef cattle (stocking density ranges between 1.7 and 2.2 livestock units ha−1). Occasionally, some fields get mowed before grazing, and the cut grass is left on the field. From late March to early October, grazing events occur at different times for individual fields, with weekly to monthly rotational grazing cycles among fields belonging to the same farm, which introduces spatial and temporal heterogeneity in the footprint during this period. Cattle are housed for the remainder of the year (November–February). Fertilizer, including a variety of industrial inorganic fertilizers, slurry, and manure, was applied at various times to fields throughout the year totalling about 300–350 kg nitrogen (N) ha−1 y−1 between 2003 and 2004 and about 175–250 N ha−1 y−1 since 2005, with inorganic N fertilizer contributing about 60–90%. Although the spatial and temporal variability of these management activities causes considerable heterogeneity of the footprint within each year, averaged over the year, the management intensity and the proportion of grazed versus harvested fields was similar for all years. Furthermore, to evaluate the functioning of this farming system as a whole, our flux measurements represent an estimate of average CO2 exchange of a managed grassland landscape (rather than of a single paddock ecosystem) which in this region is typically characterized by a combination of several small and individually managed paddock ecosystems.
In early 2005, a sector (5.25 ha in size) located southwest of the flux tower was ploughed and afforested with broadleaf trees. In this study we therefore disregarded fluxes if the mean Wd originated from this sector (230°–270°) for all years, including 2003 and 2004 for consistency.
Instrumentation for Micrometeorological Measurements
The net ecosystem exchange (NEE) of CO2 was determined with the EC technique using a LI-7500 open path infrared gas analyzer (IRGA) (LICOR Inc., Lincoln, Nebraska, USA) in combination with a RM Young 3-D sonic anemometer (RM Young, model 81000, Traverse City, Michigan, USA) up to 2005 and with a Campbell Scientific (CS) 3-D sonic anemometer (CSAT3; CS, Logan, Utah, USA) in the following years. High frequency data (10 Hz) were averaged to 30 min averages.
The EC sensor height varied among years with 10, 3, and 6 m in 2003, 2004, and 2005–2009, respectively. For the three different measurement heights, the mean distances for the peak flux source area and the 75% fetch were 191, 65, and 104 m and 1330, 449, and 724 m respectively, based on the Hsieh footprint model (Hsieh and others 2000).
Measurements of meteorological variables included Ta and relative humidity (RH) (HMP45A; Vaisala, Helsinki, Finland), PAR (PAR LITE sensor, Kipp & Zonen, Delft, The Netherlands), 2-dimensional wind speed (Ws) and Wd (RM Young). Vapor pressure deficit (VPD) was calculated based on Ta and RH. PPT was measured using a CS-ARG100 rain gauge. Soil temperature was determined with CS 107 temperature probes at 2.5, 5, and 7.5 cm depth. Volumetric soil water content (SWC) was measured at depths of 5, 10, 25, and 50 cm with CS 615 time domain reflectometers set horizontally in the soil, and for an average of 0–30-cm depth (root zone) with a vertically installed CS 615 probe. All meteorological measurements were sampled at 1 min intervals and averaged over 30 min. Gaps in the meteorological data due to power failure were filled with data from a nearby weather station (Met Eireann, Cork Airport Meteorological Station). Further details of instrument set up for micrometeorological measurements were previously given by Jaksic and others (2006).
Data Processing, Gap-Filling, and Flux Partitioning
Data processing followed standard protocols outlined by Aubinet and others (2000) and was previously described in detail by Jaksic and others (2006). In brief, we removed outliers in the dataset following Papale and others (2006), double rotated the wind vectors to align the coordinate system into the mean wind streamlines (Wilczak and others 2001), and applied a correction for variations in air density (Webb and others 1980). Data collected during and up to 1 h following rain events were rejected. Fluxes were also discarded for periods when friction velocity (u *) was below the critical threshold of 0.2 m s−1 which was determined based on a regression of night time fluxes against u * (Massman and Lee 2002).
Sensor malfunctioning resulted in an extended data gap in 2007. Owing to the large uncertainty associated with the gap-filling of longer periods during the growing season as a result of the spatial and temporal variability in management activities, we excluded the entire year of 2007 from our further analysis. Gaps of 3 to 4 weeks occurred in late March/early April 2005 and December 2009. These gaps however were gap-filled (as described below) and included in the annual sum because interfering effects from management practices (that is, harvest) were minimal during these periods. In total, data gaps as a result of sensor malfunctioning, exclusion of data from the afforested sector, and data filtering during post-processing steps required gap-filling of 64, 58, 74, 68, 56, and 63% of the annual datasets in 2003, 2004, 2005, 2006, 2008, and 2009, respectively.
We developed a general exponential relationship between night time ER (above u * threshold) and soil temperature at 5 cm depth based on data from all years. The additional effect of soil moisture was determined through residual analysis and added as a second scalar. To account for seasonal changes in the ER–Ts–SWC relationship, we then applied a time-varying correction factor determined by the comparison of modeled to observed values over a 100-points moving window (using a fixed number of measured (non-missing) data points rather than a fixed period of time) to adjust modeled values based on current conditions following Barr and others (2004). This final model was used to fill gaps in night time fluxes and to estimate daytime ER.
GEP was determined as the difference between daytime estimates of measured NEE and modeled ER. We developed a model that related GEP to environmental parameters including PAR, Ts, VPD, and SWC through residual analysis following Richardson and others (2007). A time-varying correction factor (Barr and others 2004) was also added to the final GEP model to account for seasonal variations in the general environmental response due to changes in phenology and short-term impacts from management events (that is, biomass removal via grazing and cutting; growth stimulation via fertilization) throughout the year. Previous studies used daily to monthly relationships between fluxes and environmental variables to account for these effects (Reichstein and others 2005; Gilmanov and others 2007; Chen and others 2009a; Zeeman and others 2010). However, high spatial and temporal heterogeneity of land management activities within the footprint caused random effects that in combination with fluctuations of the flux source area often hampered the development of robust relationships between component fluxes and environmental variables on daily to monthly scales throughout the summer months. Therefore, using a general robust multi-environmental-parameter relationship adjusted by a high temporal resolution correction factor proved best to account for these complex short-term management effects (model evaluation based on comparison of root mean square errors). The correction factor however was considered unsuitable during the multi-week gaps (see above) and during autumn 2005 when good data coverage was sparse. These periods were therefore filled using the gap-filling models that related ER to Ts and SWC and related GEP to PAR, Ts, SWC, and VPD as described above. Gaps in daytime NEE were filled with the difference of modeled GEP and ER estimates. We use the micrometeorological sign convention in which fluxes from the atmosphere to the ecosystem are negative and emission fluxes are positive. For illustrative purposes however, GEP is presented in absolute values (|GEP|) where appropriate to facilitate the direct comparison with flux magnitudes of ER.
Uncertainty in Gap-Filled Flux Estimates
We estimated the uncertainty in monthly, seasonal and annual NEE, GEP, and ER estimates introduced by the gap-filling models by aggregating their mean absolute error (MAE) and bias error (BE) over the respective number of filled data points. Because MAE and BE varied on a seasonal scale, we derived MAE and BE separately for winter (December–February), spring (March–May), summer (June–August), and autumn (September–November) periods. In addition, MAE was also added to and aggregated over the respective number of observed values to simulate random flux measurement errors (Richardson and others 2008).
Results
Environmental Conditions
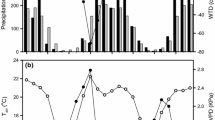
The seasonal patterns of monthly mean air and soil temperatures were similar for all years with the 6-year averages peaking in July/August (Figure 1). The 6-year average of monthly mean PAR peaked in June. On average, monthly PPT during autumn and winter months (October–January) was about 50 mm higher compared to the other months. The greatest deviation in soil moisture dynamics from the 6-year average was observed in 2006 when monthly mean SWC remained exceptionally high and close to the saturation level (~0.46 m3 m−3) throughout the winter and early spring, followed by minimum monthly values of 0.21 m3 m−3 in the summer coinciding with reduced rainfall. It is noteworthy, however, that diurnal SWC never dropped below the wilting point of 0.12 m3 m−3 during this exceptional drought period. The summer of 2006 also included months with highest VPD and Ta among all years.

Monthly averages of air temperature (Ta), soil temperature (Ts) at 5-cm depth, photosynthetically active radiation (PAR), volumetric soil water content (SWC) from 0 to 30 cm depth (root zone), atmospheric water vapor pressure deficit (VPD), and monthly totals of precipitation (PPT) at the Dripsey grassland. Circles connected with dotted line indicate 6-year average for each month.
On a growing season and annual scale, the year 2008 was characterized by lowest mean temperatures (Ta and Ts), PAR, VPD and above average SWC, whereas highest annual mean temperature and VPD combined with lowest SWC were measured in 2006 (Table 1). Overall, the inter-annual variability was small for annual mean Ta and Ts (< ±0.5°C), PAR (212 ± 17 μmol m−2 s−1), VPD (1.26 ± 0.08 kPa), and SWC (0.38 ± 0.03 m3 m−3), whereas annual totals of PPT covered a wide range from 1178 to 1642 mm during 2003–2009.
Seasonal and Inter-Annual Patterns in Ecosystem CO2 Fluxes
Minimum monthly NEE occurred in April and May suggesting the greatest net CO2 uptake during this period (Figure 2A). Continuous CO2 uptake occurred from February to July in all years, whereas net uptake or emission of CO2 alternated for the remaining months among the study years, with the 6-year average suggesting a net CO2 loss from the ecosystem to the atmosphere for the 5 months from September to January.

Monthly totals of A net ecosystem exchange (NEE), B gross ecosystem production (GEP), and C ecosystem respiration (ER) at the Dripsey grassland. Circles connected with dotted line indicate 6-year average for each month. Error bars represent uncertainty estimates as described in the text.
The 6-year average of monthly GEP was the greatest in May and July (Figure 2B). The intermittent reduction of GEP in June was most likely the result of the first harvest cut commonly being carried out during this month. Monthly GEP from May to August was the highest in 2004. In contrast, monthly GEP was considerably reduced in spring (March and April) and late summer (August) of 2006, compared to the other years.
The 6-year average of monthly ER was the greatest from July to September (Figure 2C). The shift in the timing of maximum ER and GEP explains the patterns observed for NEE including maximum net CO2 uptake in spring and net CO2 loss throughout autumn. Similar to GEP, the monthly ER from May to August was the greatest in 2004, whereas ER was considerably lower during those months of 2006 in which GEP was reduced (that is, March, April, and August). This indicates coupling of GEP and ER on a monthly scale.
The annual NEE remained within a narrow range from −245 to −284 g C m−2 y−1 for 5 out of 6 years, given an uncertainty of ±39–52 g C m−2 y−1 (Table 1). The exception was the year 2008 in which NEE reached −352 g C m−2 y−1. Annual ER and GEP were considerably higher in the years 2003 and 2004 compared with other years.
Strong positive relationships were observed between GEP and ER on a seasonal and annual scale (Figure 3). The linear regression slope switched from less than 1 in spring to greater than 1 in summer, while being approximately 1 in autumn. Linear regression slopes close to unity over the period March–August as well as for the entire year demonstrate the strong coupling of GEP and ER on these temporal scales and may explain the small variations in annual NEE. This dependency is further confirmed by similar directions and magnitudes in annual anomalies of GEP and ER in all years, except for 2008 in which anomalies were found diverted in opposite directions (Figure 4). Furthermore, although statistically not significant (P > 0.05), there was an apparent trend of spring GEP and summer ER being positively correlated (coefficient of determination (R 2) = 0.38).

Correlation between totals of ecosystem respiration (ER) and gross ecosystem production (GEP) on A seasonal (spring = March 1–May 31; summer = June 1–August 31; autumn = September 1–October 31) and B early growing season (that is, spring and summer) and annual scale; dotted line indicates 1:1 relationship; GEP is shown in absolute values (|GEP|) for illustrative purposes. Error bars represent uncertainty estimates as described in the text.

Anomalies in annual gross ecosystem production (GEP) and ecosystem respiration (ER). Double arrow highlights divergence of GEP and ER anomalies.
Environmental Controls on Ecosystem CO2 Fluxes
In general, short-term relationships between fluxes and environmental parameters were weak and most likely masked by grazing and silage cutting events which caused spatial and temporal heterogeneity of biomass and leaf area primarily during the harvesting period (June–August). Owing to the concurrent variations of these management events and the predominant flux source area, a clear signal from environmental variables was, therefore, not always apparent for the entire flux footprint.
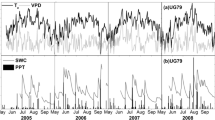
Nevertheless, peaks in the 5-day averages of GEP and NEE commonly coincided with peaks in Ta and PAR (Figure 5, showing the representative years 2003, 2006, and 2008). Furthermore, peaks in GEP were often accompanied by simultaneous or slightly delayed (by 1 or 2 weeks) peaks in ER indicating a dependency of ER on GEP.

Five-day averages for daily totals of net ecosystem exchange (NEE), gross ecosystem production (GEP), ecosystem respiration (ER), and for daily means of air temperature (Ta), photosynthetically active radiation (PAR), soil water content (SWC, 0–30-cm depth) and 5-day totals of precipitation (PPT; shown as horizontal bars) in the selected years 2003, 2006, and 2008. Roman numbers (I and IIa,b,c) indicate various drought phases; note that GEP is shown in absolute values (|GEP|) for illustrative purposes.
There were no related patterns between the time series of CO2 fluxes and PPT events or fluctuations of soil moisture contents (Figure 5). However, residual analysis showed that both ER and GEP started to decrease as SWC either approached values beyond the saturation levels (above ~0.46 m3 m−3) or below about 0.25 m3 m−3 (data not shown). Thus, higher soil moisture levels during the early spring 2006, for instance, may explain lower ER and GEP during this period compared with other years (Figure 5: also recall Figures 1 and 2).
There were two pronounced drought periods occurring within the study period which had varying effects on ecosystem CO2 fluxes. The first drought event in late summer 2003 showed no apparent effect on ecosystem CO2 fluxes (Figure 5, I). The effect of the second drought period in summer 2006, however, can be subdivided into three phases (IIa,b,c). During the first phase (IIa, first week of June to mid-July), ecosystem CO2 fluxes remained initially unaffected by the drop in SWC and continued to be primarily driven by variations in Ta and PAR. The end of this phase was characterized by a drop in GEP, possibly because of a combination of harvesting events and/or drops in Ta and PAR, resulting in a net CO2 loss. In the second phase (IIb; mid-July until mid-August), SWC further decreased to less than 0.20 m3 m−3. During this period, a simultaneous rapid decrease in both GEP and ER occurred, whereas NEE remained unaffected. The third phase terminating this drought period (IIc, mid-August until mid-September) showed large net CO2 losses because the onset of heavy rainfall events resulted in an immediate increase of ER, whereas the recovery of GEP was delayed by 1 or 2 weeks.
Mean spring GEP and ER were positively correlated with mean spring Ta and PAR; however, owing to the simultaneous increase of both GEP and ER with higher TA and PAR, spring NEE did not correlate with these two environmental parameters (Figure 6A, B). Similarly, despite a positive correlation of annual GEP to summer PPT (up to a threshold of ~300 mm) and spring PAR, no effect on annual NEE was apparent because the simultaneous response in ER counter-balanced the differences in GEP (Figure 6C, D). We did not find any statistical relationship between any of the measured environmental controls and NEE on an annual scale.

Correlations of net ecosystem exchange (NEE), gross ecosystem production (GEP), and ecosystem respiration (ER) with mean air temperature (Ta), photosynthetically active radiation (PAR), and totals of precipitation (PPT) on seasonal (spring = March 1–May 31; summer = June 1–August 31) and annual scales; note that GEP is shown in absolute values (|GEP|) for illustrative purposes.
Monthly ecosystem C use efficiency (CUE) defined as the ratio of NEE:GEP was negatively correlated with temperature (Ta and Ts) and VPD, and positively correlated with SWC (Figure 7A–C). Monthly CUE was not related to PAR (Figure 7D). A stepwise regression further suggested that anomalies in monthly CUE may be sufficiently explained by temperature and SWC, whereas the explanatory power of VPD was eliminated because of its correlation with temperature. The correlations of CUE to these environmental parameters explained the highest annual CUE in 2008, which was the year with the lowest annual mean temperatures and VPD, combined with relatively high annual mean SWC (recall Table 1), and produced a negative relationship between annual CUE and NEE (see inset in Figure 7D). Thus, while variations in individual environmental parameters did not directly result in inter-annual differences in NEE, they indirectly affected annual NEE through their control on CUE.

Correlations of monthly ecosystem carbon use efficiency (CUE; =NEE:GEP) with monthly means of A air/soil temperature (Ta; Ts), B volumetric soil water content (SWC, 0–30-cm depth average), C vapor pressure deficit (VPD), and D photosynthetically active radiation (PAR) during the main growing season. Inset figure shows relationship between annual net ecosystem exchange (NEE) and CUE (with 2008 highlighted as unfilled symbol).
Discussion
Stable Inter-Annual NEE Due to Coupling of GEP and ER
The 6-year averages of NEE, GEP, and ER in this grassland were at the upper end of respective ranges reported for intensively managed grasslands in a recent review of global non-forest ecosystems by Gilmanov and others (2010), indicating its potential for high productivity and C turnover. Compared to previous long-term studies primarily conducted in drier and more extensively managed grasslands, the range of approximately 100 g C m−2 y−1 between minimum and maximum annual NEE observed in this study was less than the range of 88 to −141 g C m−2 y−1 over 6 years in a Californian grassland site (Ma and others 2007), and 170 to −116/236 to −32 g C m−2 y−1 over 6/7 years in two sagebrush–steppe ecosystems (Gilmanov and others 2006), but comparable to the range from 69 to −42 g C m−2 y−1 over a 6-year study period in a temperate mountain meadow (Wohlfahrt and others 2008) and from 4 to 70 g C m−2 y−1 over 6 years in a moderately grazed mixed-grass prairie (Frank 2004). In contrast to these other long-term studies, however, our grassland provided consistent and strong annual CO2 uptake throughout the entire study period. A longer (from mid-March until mid-October) and more humid growing season compared to global grasslands may generally favor the CO2 uptake potential in humid temperate grasslands. Thus, the strong capability for atmospheric CO2 uptake might be a unique and distinguishing feature for intensively managed humid temperate grasslands compared with more extensively managed grasslands in dry temperate, continental, or Mediterranean climates.
Based on our results, we suggest that the stable inter-annual NEE was primarily due to a strong coupling of GEP and ER in this grassland. Numerous studies have previously suggested a tight relationship between GEP and ER in grassland ecosystems. For instance, Bahn and others (2008) found that soil respiration was positively correlated with assimilate supply over a range of European grassland sites. There is evidence in the literature that reduced assimilate supply following clipping events may induce a negative effect on both autotrophic (root) and heterotrophic respiration (Bond-Lamberty and others 2004; Bahn and others 2006, 2008). Kaštovská and Šantrůčková (2007) reported rapid belowground allocation of fixed CO2 and soil microbial biomass turnover in pasture plant–soil systems.
The approximate 1:1 linear GEP–ER response observed on the growing season and annual scale in our study, however, is in contrast to other studies which commonly observed linear response slopes less than 1 (Ma and others 2007; Fu and others 2009; Schmitt and others 2010). A recent synthesis of non-forest ecosystems, however, indicated that a slope of less than 1 occurs primarily for lower productivity ecosystems, whereas the slope for high productive grasslands appeared to be close to 1 (Figure 10b in Gilmanov and others 2010), suggesting that the strength of GEP–ER coupling increases with grassland productivity. To achieve a 1:1 linear GEP–ER response, similar absolute changes in GEP and ER are required. This condition was met in this study, possibly because of greater biomass production during the early growing season resulting in greater heterotrophic respiration from increased availability of decomposable material during the later parts of the growing season. This hypothesis is supported by the positive relationship of spring GEP and summer ER in our study, suggesting that changes in spring productivity lead to corresponding changes in summer ER, thereby maintaining a similar net CO2 balance. Thus, fast C allocation and turnover rates resulting in tight coupling of GEP and ER may favor a stable annual uptake of atmospheric CO2 in this intensively managed grassland.
Besides the coupling mechanism between GEP and ER, limited inter-annual variation in temperature and generally abundant and sufficient soil moisture availability may have been the additional factors contributing to a stable and narrow range of annual NEE in this grassland. Studies in drier regions more prone to soil water stress have reported net CO2 loss during drought events, commonly because of greater reductions in GEP compared with ER during these periods (Kim and others 1992; Novick and others 2004; Nagy and others 2007; Aires and others 2008; Arnone and others 2008; Zhang and others 2010). Thus, a consistent annual CO2 uptake may be a feature more common for grasslands in humid temperate regions.
Environmental Controls on Grassland Ecosystem CO2 Exchange
The lack of an individual environmental driver for inter-annual variability in NEE observed in this study was due to the compensating effects of environmental variables (for example, Ta, PAR, PPT, and so on) on the component fluxes GEP and ER, combined with a masking effect from management practices. Similar observations were previously made by Wohlfahrt and others (2008) for a repeatedly cut temperate mountain meadow.
Interestingly, two prolonged periods with reduced summer PPT showed contrasting effects on short-term CO2 flux dynamics in 2003 versus 2006. We propose that, in this grassland, the effect of reduced summer PPT on CO2 fluxes may depend on (i) the timing of the drought event (mid-summer versus late summer), and/or (ii) SWC falling below a threshold level of less than about 0.20 m3 m−3. In the first case, the combined effects from harvest and drought during mid-summer may have greater impact on ecosystem CO2 fluxes than the sole drought effect during the late summer season (after the main harvesting season). The second possibility implies that soil moisture levels below a SWC threshold of about 0.20 m3 m−3 are necessary in this grassland to observe negative effects on GEP and ER as a result of incipient soil water stress. It is noteworthy, however, that NEE was less affected during this period of minimum SWC. Instead, it was the onset of the rainy period following the drought which led to positive NEE and CO2 losses due to an immediate pulse in ER whereas the recovery of GEP lagged behind by 1 or 2 weeks. The timing and interaction of drought and rain pulse events have been previously highlighted as important controls on grassland CO2 fluxes (Kim and others 1992; Xu and others 2004; Gilmanov and others 2006; Ma and others 2007; Chen and others 2009b).
Despite the apparent lack of a single environmental control on annual NEE, temperature, VPD, and SWC indirectly affected annual NEE through their correlation with CUE in our study. This implies that despite generally coupled component fluxes, environmental variables may affect GEP and ER in a different manner. For instance, higher temperature may increase ER while the concurrent high VPD may constrain GEP, which, therefore, reduces CUE. However, our results suggested that only the continuous combination of low temperature, low VPD, and high SWC or vice versa may cause significant deviations from the otherwise stable and narrow range of annual CUE and NEE in this intensively managed temperate grassland landscape. Thus, drier and warmer summer periods predicted for Ireland by future scenarios (McGrath and Lynch 2008) might possibly result in a moderate reduction of the CO2 uptake potential but are unlikely to lead to a net CO2 loss from this grassland on an annual scale.
Effect of Management Practices on Ecosystem CO2 Exchange
Owing to the spatial and temporal heterogeneity in management practices combined with additional effects from environmental variables (that is, Ta and PAR), it was not possible to clearly isolate the effects of management practices in this study. Nevertheless, our results indicated that management practices have a limited effect on annual NEE at this grassland because of the strong coupling of GEP and ER. For instance, we observed that sudden drops in GEP throughout the summer season, likely as a result of enhanced harvesting and or/grazing intensity were usually accompanied by simultaneous or somewhat delayed decreases in ER. This is in agreement with previous studies that reported significant and immediate impacts of harvest/grazing on soil respiration due to the link between photosynthetic supply and respiratory processes (Bremer and Ham 2002; Bahn and others 2006, 2008). In contrast, studies in mountainous grassland sites reported that management events (harvesting, grazing, and so on) negatively affected NEE primarily via their impact on GEP (Rogiers and others 2005; Wohlfahrt and others 2008; Schmitt and others 2010). Other studies suggested that an initial reduction of NEE following harvest may be restored within the timescale of a week because of the fast recovery of leaf biomass, with weak effects on annual NEE (for example, Novick and others 2004).
Moreover, greater GEP in 2003 and 2004 was likely the result of higher N fertilizer application rates compared to the following years. However, owing to a simultaneous increase in ER, the net CO2 uptake potential of those 2 years with higher N fertilizer application rates remained similar to that in following years with comparably lower N fertilizer application rates. It is known that intensive N fertilization may, besides increasing gross plant production, also enhance mineralization rates and subsequently lead to greater soil C losses from stimulated respiratory processes (Hatch and others 2000; Jones and Donnelly 2004). In contrast, Shimizu and others (2009) found that relative to a non-fertilized control site, inorganic fertilizer application rates of approximately 175 kg N ha−1 y−1 (thus slightly lower compared to ~175 to 250 kg N ha−1 y−1 applied at our grassland during 2005–2009) led to a greater net ecosystem CO2 uptake in a less intensively managed meadow located in a humid continental climate. Thus, our study indicates that the response of NEE to management practices may diminish for intensively managed (inorganic N fertilization rates > ~200 kg N ha−1 y−1) and highly productive grasslands.
Grassland C Balance
An important characteristic of managed grasslands is the additional C input (via manure fertilization, concentrate feed, and CH4 oxidation) and output (in silage, milk, animal respiration during winter housing, dissolved organic C, and CH4 losses from enteric fermentation and dung/slurry storage tanks) beyond ecosystem boundaries. Accounting for these fluxes provides an estimate of the net C balance on the farm scale, also defined as the NBP (Soussana and others 2007). For the Dripsey grassland, previous studies estimated the additional annual C import and export to be in the magnitude of 125–169 and 40–68 g C m−2 y−1, respectively (Jaksic and others 2006; Byrne and others 2007). Thus, the long-term mean NBP of our grassland based on the 6-year average NEE and these additional farm scale C fluxes (ignoring additional C losses via volatile organic compounds and dissolved inorganic C) is estimated at −184 ± 57 g C m−2 y−1 (negative NBP indicating net C sink). This is more than twice a recent estimate of average NBP of −74 ± 10 g C m−2 y−1 (Ciais and others 2010) and at the upper end of the range from 266 to −231 g C m−2 y−1 reported for intensively managed European grasslands (Soussana and others 2007; Ciais and others 2010). Assuming that most of the C sequestration occurs within the upper 30-cm soil layer, we estimate that C sequestration at that rate would result in an annual increase in the SOC concentration of 0.061%. Unfortunately, it was not possible to confirm such small changes with repeated soil C measurements given the natural spatial variability of SOC. Nevertheless, these findings indicate that under the current management practices, this grassland may also be considered as a considerable and consistent C sink on the farm scale.
Conclusions
This study investigated 6 years of CO2 flux data measured with the EC technique in an intensively managed grassland in the humid temperate climate of southern Ireland. A wide range in annual N fertilization and PPT rates, as well as contrasting amounts of summer PPT occurred throughout the study period. Based on findings from these long-term measurements, we conclude that this grassland landscape provided a consistent and strong annual uptake of CO2 with a 6-year mean of 277 ± 46 g C m−2 y−1. The consistency in the CO2 uptake capacity might be a distinguishing feature of grasslands in humid temperate regions compared to those in drier climate regions. Our findings further imply that the strength of CO2 uptake in this grassland may not be very sensitive to predicted alterations in future climate (that is, warmer and drier summers).
Inter-annual variability in NEE was limited mainly by a strong coupling of the component fluxes GEP and ER, which showed similar responses to changes in environmental variables and/or management practices on a seasonal and annual scale. We propose that enhanced coupling of component fluxes could be an important characteristic for intensively managed grasslands. An estimate of the NBP, in which NEE was adjusted by additional C input and output fluxes because of management activities, indicated that this grassland may also function as a C sink on the farm scale. Thus, under the current management regime, this intensively managed grassland is able to provide long-term beneficial effects within both economical and climate change related context.
References
Aires L, Pio C, Pereira J. 2008. Carbon dioxide exchange above a Mediterranean C3/C4 grassland during two climatologically contrasting years. Glob Chang Biol 14:539–55.
Allard V, Soussana J, Falcimagne R, Berbigier P, Bonnefond J, Ceschia E, D’hour P, Hénault C, Laville P, Martin C, Pinarès-Patino C. 2007. The role of grazing management for the net biome productivity and greenhouse gas budget (CO2, N2O and CH4) of semi-natural grassland. Agric Ecosyst Environ 121:47–58.
Arnone JAIII, Verburg PSJ, Johnson DW, Larsen JD, Jasoni RL, Lucchesi AJ, Batts CM, von Nagy C, Coulombe WG, Schorran DE, Buck PE, Braswell BH, Coleman JS, Sherry RA, Wallace LL, Luo Y, Schimel DS. 2008. Prolonged suppression of ecosystem carbon dioxide uptake after an anomalously warm year. Nature 455:383–6.
Aubinet M, Grelle A, Ibrom A, Rannik U, Moncrieff J, Foken T, Kowalski A, Martin P, Berbigier P, Bernhofer C, Clement R, Elbers J, Granier A, Grunwald T, Morgenstern K, Pilegaard K, Rebmann C, Snijders W, Valentini R, Vesala T. 2000. Estimates of the annual net carbon and water exchange of forests: The EUROFLUX methodology. Adv Ecol Res 30:113–75.
Bahn M, Knapp M, Garajova Z, Pfahringer N, Cernusca A. 2006. Root respiration in temperate mountain grasslands differing in land use. Glob Chang Biol 12:995–1006.
Bahn M, Rodeghiero M, Anderson-Dunn M, Dore S, Gimeno C, Drösler M, Williams M, Ammann C, Berninger F, Flechard C, Jones S, Balzarolo M, Kumar S, Newesely C, Priwitzer T, Raschi A, Siegwolf R, Susiluoto S, Tenhunen J, Wohlfahrt G, Cernusca A. 2008. Soil respiration in European grasslands in relation to climate and assimilate supply. Ecosystems 11:1352–67.
Barr A, Black T, Hogg E, Kljun N, Morgenstern K, Nesic Z. 2004. Inter-annual variability in the leaf area index of a boreal aspen-hazelnut forest in relation to net ecosystem production. Agricult Forest Meteorol 126:237–55.
Bond-Lamberty B, Wang C, Gower ST. 2004. A global relationship between the heterotrophic and autotrophic components of soil respiration? Glob Chang Biol 10:1756–66.
Bremer DJ, Ham JM. 2002. Measurement and modeling of soil CO2 flux in a temperate grassland under mowed and burned regimes. Ecol Appl 12:1318–28.
Byrne KA, Kiely G. 2006. Partitioning of respiration in an intensively managed grassland. Plant Soil 282:281–9.
Byrne KA, Kiely G, Leahy P. 2005. CO2 fluxes in adjacent new and permanent temperate grasslands. Agric For Meteorol 135:82–92.
Byrne K, Kiely G, Leahy P. 2007. Carbon sequestration determined using farm scale carbon balance and eddy covariance. Agric Ecosyst Environ 121:357–64.
Chen J, Shen M, Kato T. 2009a. Diurnal and seasonal variations in light-use efficiency in an alpine meadow ecosystem: causes and implications for remote sensing. J Plant Ecol 2:173–85.
Chen S, Lin G, Huang J, Jenerette GD. 2009b. Dependence of carbon sequestration on the differential responses of ecosystem photosynthesis and respiration to rain pulses in a semiarid steppe. Glob Chang Biol 15:2450–61.
Ciais P, Soussana JF, Vuichard N, Luyssaert S, Don A, Janssens IA, Piao SL, Dechow R, Lathière J, Maignan F, Wattenbach M, Smith P, Ammann C, Freibauer A, Schulze ED, the CARBOEUROPE Synthesis Team. 2010. The greenhouse gas balance of European grasslands. Biogeosci Discuss 7:5997–6050.
Flanagan L, Wever LA, Carlson P. 2002. Seasonal and interannual variation in carbon dioxide exchange and carbon balance in a northern temperate grassland. Glob Chang Biol 8:599–615.
Frank AB. 2004. Six years of CO2 flux measurements for a moderately grazed mixed-grass prairie. Environ Manage 33:S426–31.
Fu Y, Zheng Z, Yu G, Hu Z, Sun X, Shi P, Wang Y, Zhao X. 2009. Environmental influences on carbon dioxide fluxes over three grassland ecosystems in China. Biogeosciences 6:2879–93.
Gilmanov TG, Svejcar TJ, Johnson DA, Angell RF, Saliendra NZ, Wylie BK. 2006. Long-term dynamics of production, respiration, and net CO2 exchange in two sagebrush–steppe ecosystems. Rangeland Ecol Manage 59:585–99.
Gilmanov T, Soussana J, Aires L, Allard V, Ammann C, Balzarolo M, Barcza Z, Bernhofer C, Campbell C, Cernusca A, Cescatti A, Clifton-Brown J, Dirks B, Dore S, Eugster W, Fuhrer J, Gimeno C, Gruenwald T, Haszpra L, Hensen A, Ibrom A, Jacobs A, Jones M, Lanigan G, Laurila T, Lohila A, Manca G, Marcolla B, Nagy Z, Pilegaard K, Pinter K, Pio C, Raschi A, Rogiers N, Sanz M, Stefani P, Sutton M, Tuba Z, Valentini R, Williams M, Wohlfahrt G. 2007. Partitioning European grassland net ecosystem CO2 exchange into gross primary productivity and ecosystem respiration using light response function analysis. Agric Ecosyst Environ 121:93–120.
Gilmanov TG, Aires L, Barcza Z, Baron VS, Belelli L, Beringer J, Billesbach D, Bonal D, Bradford J, Ceschia E, Cook D, Corradi C, Frank A, Gianelle D, Gimeno C, Gruenwald T, Guo H, Hanan N, Haszpra L, Heilman J, Jacobs A, Jones MB, Johnson DA, Kiely G, Li S, Magliulo V, Moors E, Nagy Z, Nasyrov M, Owensby C, Pinter K, Pio C, Reichstein M, Sanz MJ, Scott R, Soussana JF, Stoy PC, Svejcar T, Tuba Z, Zhou G. 2010. Productivity, respiration, and light-response parameters of world grassland and agroecosystems derived from flux-tower measurements. Rangeland Ecol Manage 63:16–39.
Hatch DJ, Lovell RD, Antil RS, Jarvis SC, Owen PM. 2000. Nitrogen mineralization and microbial activity in permanent pastures amended with nitrogen fertilizer or dung. Biol Fertil Soils 30:288–93.
Heitschmidt RK, Klement KD, Haferkamp MR. 2005. Interactive effects of drought and grazing on northern Great Plains rangelands. Rangeland Ecol Manage 58:11–19.
Hsieh CI, Katul G, Chi T. 2000. An approximate analytical model for footprint estimation of scaler fluxes in thermally stratified atmospheric flows. Adv Water Resour 23:765–72.
IPCC. 2007. Climate change 2007: the physical science basis. Contribution of Working Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate Change. Cambridge, UK and New York, NY, USA: Cambridge University Press. 881 pp.
Jaksic V, Kiely G, Albertson J, Oren R, Katul G, Leahy P, Byrne K. 2006. Net ecosystem exchange of grassland in contrasting wet and dry years. Agric For Meteorol 139:323–34.
Janssens IA, Freibauer A, Ciais P, Smith P, Nabuurs GJ, Folberth G, Schlamadinger B, Hutjes RWA, Ceulemans R, Schulze ED, Valentini R, Dolman AJ. 2003. Europe’s terrestrial biosphere absorbs 7 to 12% of European anthropogenic CO2 emissions. Science 300:1538–42.
Janssens IA, Freibauer A, Schlamadinger B, Ceulemans R, Ciais P, Dolman AJ, Heimann M, Nabuurs GJ, Smith P, Valentini R et al. 2005. The carbon budget of terrestrial ecosystems at country-scale—a European case study. Biogeosciences 2:15–26.
Jones MB, Donnelly A. 2004. Carbon sequestration in temperate grassland ecosystems and the influence of management, climate and elevated CO2. New Phytol 164:423–39.
Kaštovská E, Šantrůčková H. 2007. Fate and dynamics of recently fixed C in pasture plant–soil system under field conditions. Plant Soil 300:61–9.
Kim J, Verma SB, Clement RJ. 1992. Carbon dioxide budget in a temperate grassland ecosystem. J Geophys Res 97:6057–63.
Lawton D, Leahy P, Kiely G, Byrne KA, Calanca P. 2006. Modeling of net ecosystem exchange and its components for a humid grassland ecosystem. J Geophys Res 111.
Ma S, Baldocchi DD, Xu L, Hehn T. 2007. Inter-annual variability in carbon dioxide exchange of an oak/grass savanna and open grassland in California. Agric For Meteorol 147:157–71.
Massman W, Lee X. 2002. Eddy covariance flux corrections and uncertainties in long-term studies of carbon and energy exchanges. Agric For Meteorol 113:121–44.
McGrath R, Lynch P. 2008. Ireland in a warmer world: scientific predictions of the Irish climate in the twenty-first century. Report of the C4I Project, Met Éireann.
Nagy Z, Pintér K, Czóbel S, Balogh J, Horváth L, Fóti S, Barcza Z, Weidinger T, Csintalan Z, Dinh N, Grosz B, Tuba Z. 2007. The carbon budget of semi-arid grassland in a wet and a dry year in Hungary. Agric Ecosyst Environ 121:21–9.
Novick KA, Stoy PC, Katul GG, Ellsworth DS, Siqueira MBS, Juang J, Oren R. 2004. Carbon dioxide and water vapor exchange in a warm temperate grassland. Oecologia 138:259–74.
Papale D, Reichstein M, Aubinet M, Canfora E, Bernhofer C, Kutsch W, Longdoz B, Rambal S, Valentini R, Vesala T, Yakir D. 2006. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences 3:571–83.
Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grunwald T, Havrankova K, Ilvesniemi H, Janous D, Knohl A, Laurila T, Lohila A, Loustau D, Matteucci G, Meyers T, Miglietta F, Ourcival J, Pumpanen J, Rambal S, Rotenberg E, Sanz M, Tenhunen J, Seufert G, Vaccari F, Vesala T, Yakir D, Valentini R. 2005. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Glob Chang Biol 11:1424–39.
Richardson AD, Hollinger DY, Aber JD, Ollinger SV, Braswell BH. 2007. Environmental variation is directly responsible for short- but not long-term variation in forest–atmosphere carbon exchange. Glob Chang Biol 13:788–803.
Richardson AD, Mahecha MD, Falge E, Kattge J, Moffat AM, Papale D, Reichstein M, Stauch VJ, Braswell BH, Churkina G et al. 2008. Statistical properties of random CO2 flux measurement uncertainty inferred from model residuals. Agric For Meteorol 148:38–50.
Rogiers N, Eugster W, Furger M, Siegwolf R. 2005. Effect of land management on ecosystem carbon fluxes at a subalpine grassland site in the Swiss Alps. Theor Appl Climatol 80:187–203.
Sala O, Paruelo J. 1997. Ecosystem services in grasslands. In: Daily GC, Ed. Nature’s services: societal dependence on natural ecosystems. Washington, DC: Island Press. p 237–52.
Schmitt M, Bahn M, Wohlfahrt G, Tappeiner U, Cernusca A. 2010. Land use affects the net ecosystem CO2 exchange and its components in mountain grasslands. Biogeosciences 7:2297–309.
Scurlock JMO, Hall DO. 1998. The global carbon sink: a grassland perspective. Glob Chang Biol 4:229–33.
Shimizu M, Marutani S, Desyatkin A, Jin T, Hata H, Hatano R. 2009. The effect of manure application on carbon dynamics and budgets in a managed grassland of Southern Hokkaido, Japan. Agric Ecosyst Environ 130:31–40.
Soussana J, Allard V, Pilegaard K, Ambus P, Amman C, Campbell C, Ceschia E, Clifton-Brown J, Czobel S, Domingues R, Flechard C, Fuhrer J, Hensen A, Horvath L, Jones M, Kasper G, Martin C, Nagy Z, Neftel A, Raschi A, Baronti S, Rees R, Skiba U, Stefani P, Manca G, Sutton M, Tubaf Z, Valentini R. 2007. Full accounting of the greenhouse gas (CO2, N2O, CH4) budget of nine European grassland sites. Agric Ecosyst Environ 121:121–34.
Webb E, Pearman G, Leuning R. 1980. Correction of flux measurements for density effects due to heat and water-vapor transfer. Q J R Meteorol Soc 106:85–100.
White R, Murray S, Rohweder M. 2000. Pilot analysis of global ecosystems: grassland ecosystems. Washington, DC: World Resources Institute.
Wilczak J, Oncley S, Stage S. 2001. Sonic anemometer tilt correction algorithms. Boundary Layer Meteorol 99:127–50.
Wohlfahrt G, Hammerle A, Haslwanter A, Bahn M, Tappeiner U, Cernusca A. 2008. Seasonal and inter-annual variability of the net ecosystem CO2 exchange of a temperate mountain grassland: effects of weather and management. J Geophys Res Atmos 113.
Xu L, Baldocchi DD, Tang J. 2004. How soil moisture, rain pulses, and growth alter the response of ecosystem respiration to temperature. Global Biogeochem Cycles 18.
Zeeman MJ, Hiller R, Gilgen AK, Michna P, Plüss P, Buchmann N, Eugster W. 2010. Management and climate impacts on net CO2 fluxes and carbon budgets of three grasslands along an elevational gradient in Switzerland. Agric For Meteorol 150:519–30.
Zhang L, Wylie BK, Ji L, Gilmanov TG, Tieszen LL. 2010. Climate-driven interannual variability in net ecosystem exchange in the northern Great Plains grasslands. Rangeland Ecol Manage 63:40–50.
Acknowledgments
This study was financed by the Irish Government under the National Development Plan 2000–2006 (Grant No. 2001-CC/CD-(5/7)), and the STRIVE program 2007–2013 (Grant No. 2008-CCRP-1.1A) which is managed by the Environmental Protection Agency. We would like to thank Kilian Murphy and Nelius Foley for their support in instrument maintenance and data collection. We also express our gratitude to the local farmers for granting access to their private land to conduct this study. Helpful comments from the reviewers are greatly appreciated.
Author information
Authors and Affiliations
Corresponding author
Additional information
Author Contributions
MP wrote the paper, analyzed and interpreted the data. PL contributed to data analysis, commented on paper. GK conceived and designed the study, assisted in writing of paper, supported the research.
Rights and permissions
About this article
Cite this article
Peichl, M., Leahy, P. & Kiely, G. Six-year Stable Annual Uptake of Carbon Dioxide in Intensively Managed Humid Temperate Grassland. Ecosystems 14, 112–126 (2011). https://doi.org/10.1007/s10021-010-9398-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-010-9398-2