
Abstract
We investigated the decomposability of soil organic matter (SOM) along a chronosequence of rainforest sites in Hawaii that form a natural fertility gradient and at two long-term fertilization experiments. To estimate turnover times and pool sizes of organic matter, we used two independent methods: (1) long-term incubations and (2) a three-box soil model constrained by radiocarbon measurements. Turnover times of slow-pool SOM (the intermediate pool between active and passive pools) calculated from incubations ranged from 6 to 20 y in the O horizon and were roughly half as fast in the A horizon. The radiocarbon-based model yielded a similar pattern but slower turnover times. The calculation of the 14C turnover times is sensitive to the lag time between photosynthesis and incorporation of organic C into SOM in a given horizon. By either method, turnover times at the different sites varied two- or threefold in soils with the same climate and vegetation community. Turnover times were fastest at the sites of highest soil fertility and were correlated with litter decay rates and primary productivity. However, experimental fertilization at the two least-fertile sites had only a small and inconsistent effect on turnover, with N slowing turnover and P slightly speeding it at one site. These results support studies of litter decomposition in suggesting that while plant productivity can respond rapidly to nutrient additions, decomposition may respond much more slowly to added nutrients.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
INTRODUCTION
Rates of decomposition of soil organic matter (SOM) vary widely among ecosystems, with much of this variation attributable to the influence of soil temperature, moisture, texture, and disturbance (Singh and Gupta 1977; Gupta and Singh 1981; Linn and Doran 1984; Parton and others 1987; Anderson 1988; Beyer 1991; Nadelhoffer and others 1991; Raich and Schlesinger 1992; Howard and Howard 1993; Trumbore 1993; Biederbeck and others 1994; Hassink 1994; Townsend and others 1995; Trumbore and others 1996). Substantial uncertainties remain in estimates of SOM turnover times in different ecosystems, including how turnover is affected by the nutrient status of ecosystems and by soil mineral composition. We investigate SOM decomposition across gradients of fertility and mineral composition, and compare two methods of estimating SOM turnover time.
The influence of soil nutrient availability on decomposition has two components—direct effects on decomposition and indirect effects on the decomposability of organic material. Both have been investigated for plant litter decomposition. Experimental studies of direct effects on litter have yielded variable responses, with some studies finding stimulation of decomposition by added nutrients, others finding no effect, and a few finding inhibition of decomposition (summarized in Hobbie and Vitousek 2000a). In contrast, the influence of nutrient availability on the decomposability of plant litter (litter quality) is reasonably well-established, at least over the long term. High nutrient availability leads to higher concentrations of nutrients in litter, which are correlated (either directly or through lignin:N ratio) with more decomposable litter (for example, Swift and others 1979; Melillo and others 1982). Common-substrate experiments have found more rapid litter decomposition on more fertile soils (Vitousek and others 1994; Ostertag and Hobbie 1999; Hobbie and Vitousek 2000a) but lower decomposition rates under long-term N additions (Magill and Aber 1998).
Compared to litter studies, much less empirical or theoretical work has been done on the relationship between nutrient availability and decomposition of SOM. The avenues of nutrient influence on SOM decomposability and decomposition rate may include litter quality, nutrient availability to microbes in the soil environment, and long-term adaptation of plants or microbes to persistent nutrient conditions. Soil texture and mineralogy play an important role in long-term stabilization of SOM. For example, in Hawaii, noncrystalline minerals such as allophane stabilize organic carbon for tens of thousands of years in the deep soil (Torn and others 1997). How minerals influence the decomposition of rapidly cycling SOM pools is poorly understood, although isotopic-labeling experiments demonstrate that allophane (in particular) can slow decomposition of fast-cycling carbon (Saggar and others 1994). Other metastable products of weathering, aluminum and iron ions, are also hypothesized to form organometal chelates that stabilize C on decadal time scales (Torn and others 2002). Rates of soil carbon cycling can be modeled by grouping organic matter into three pools: active SOM with annual turnover, slow or intermediate SOM with decadal turnover, and passive SOM with turnover times on the order of centuries or millennia [for example, Century (Parton and others 1987) and Rothamsted (Jenkinson 1990) SOM models]. Organic matter that cycles on intermediate time scales makes up more than half of the organic carbon in the upper meter of most of the world’s soils, including most of our study sites (Schimel and others 1994; Trumbore 1997). As such, it likely represents the largest pool of terrestrial carbon that can respond to environmental changes on decadal time scales (Townsend and others 1995; Trumbore and Torn 2004). For this reason, we focus on this pool of carbon.
There are three methods for estimating the turnover time of intermediate-cycling SOM: (1) by long-term (>6 months) soil incubations (Townsend and others 1997); (2) by modeling the 14C content of the soil, which reflects the incorporation and loss of the atmospheric “bomb” spike (14C produced by aboveground nuclear weapons testing, mostly between 1959 and 1963) by organic matter (Trumbore 2000); and (3) after a change between C3 and C4 vegetation, by using the 13C content of SOM to calculate turnover time under certain equilibrium assumptions (Balesdent and others 1988; Veldkamp 1994). Each of these methods has limitations. Incubations disturb the soil and take place under unrealistic conditions. Radiocarbon-based modeling is limited because of nonunique model solutions and the varying age of carbon inputs to the soil (Trumbore 2000). Vegetation change causes disturbance and nonsteady-state conditions. In addition, these methods capture different types of SOM and time scales of decomposition. Incubations show the relative turnover times of organic matter in different soils under controlled conditions and without plant–soil interactions. The 14C-based turnover times reflect long-term patterns in the residence time of carbon in the plant–soil system in situ. In concert, though, these methods provide useful information on SOM turnover.
This study has two objectives: to determine how turnover times in the surface horizons vary as a function of soil fertility and mineral composition, and to compare incubation and radiocarbon methods of estimating SOM turnover time. We characterized SOM cycling in a set of six sites that vary in soil development and nutrient status. We chose these sites because they differ substantially in nutrient availability and nutrient limitation —our factors of interest—while varying little in climate, parent material, topography, and plant community composition (Crews and others 1995; Vitousek and others 1997; Chadwick and others 1999). We are not interested in soil age or succession per se, but rather in the well-controlled variation in mineralogy and nutrient availability that this gradient provides. In addition, we used long-term fertilization experiments at the youngest and oldest sites (300 y-and 4.1 million y) to test the direct effect of soil nutrients on SOM turnover.
SITE DESCRIPTION
The study was conducted on a chronosequence of six sites of ages 300 y to 4.1 million y, spanning the islands of Hawaii, Molokai, and Kauai (Crews and others 1995). Site ages and names are given in Table 1. The sites are located in cool (16°C mean annual temperature, MAT), wet (2500 mm mean annual precipitation) native rainforest, 1200 m elevation, at locally level areas where erosion and deposition were minimal (Chadwick and others 1999). The same native tree species, Metrosideros polymorpha, dominates live biomass and litterfall at all sites, and all sites share understory species (Kitayama and Mueller–Dombois 1995). The ratio of root-to-shoot C input is fairly consistent among sites, but the depth of rooting varies (Ostertag 2001). Approximately 85% of the fine roots are found within the top 20 cm at the 0.3 ky site, 30 cm at the 20 ky site, and 15 cm at the 4,100 ky site (Ostertag and Hobble 1999). The pH of the O horizon is acidic, ranging from 3.9 (0.3 ky site) to 3.7 (4,100 ky site) (Hobbie and Vitousek 2000a). The A horizon is slightly less acidic.
The chronosequence sites form a gradient in soil development and nutrient availability. Soil nitrogen accumulates with site age. Plant-available phosphorus increases during early soil development but declines at progressively older sites (Crews and others 1995). Metrosideros populations vary across the sites in the efficiency of nutrient resorption during leaf senescence (Herbert and Fownes 1999), and litter quality tends to mirror site nutrient availability. Concentrations of N and P in foliage and litter are lowest at the youngest and oldest sites, as is net primary productivity (NPP) (Table 1; Vitousek and others 1995; Herbert and Fownes 1999). Litter quality, as defined by decomposition rates of litter from all the sites decomposed at a common site, is highest for litter from the intermediate-aged sites and lowest for litter from the oldest site. Litter bags using a common substrate show that the environment for litter decomposition is also most favorable at the two intermediate-aged sites, which have the highest nutrient availability (Crews and others 1995; Hobbie and Vitousek 2000a). The nutrients that are limiting to NPP undergo a transition from N at the youngest site to P at the oldest site (Vitousek and others 1997; Vitousek and Farrington 1997; Herbert and Fownes 1999). The long-term fertilization plots have received semiannual additions of nitrogen, phosphorus, and nitrogen plus phosphorus (100 kg ha−1 y−1 of each nutrient). Fertilization began at the 0.3 ky site in 1985 (Vitousek and others 1993) and at the 4,100 ky site in 1991 (Herbert and Fownes 1995).
Soil mineral composition varies with site age, and mineral content is always higher in the A horizon than in the O horizon (Table 2). The mineral soil (A horizon and lower) has increasing amounts of noncrystalline minerals up to the 150 ky site [Kohala; referred to as 170 ky in Torn and others 1997) and Chadwick and others (1999) (Crews and others 1995; Vitousek and others 1997)] and of kaolinite and sesquioxide thereafter (Torn and others 1997; Crews and others 1995; Vitousek and others 1997; Chadwick and others 1999). Five of the soils are classified as Andisols and the 4,100 ky site is an Oxisol. Soil texture is not easily determined in Andisols or soils derived from basaltic lava (Nanzyo and others 1993; Nettleton and others 1999). The standard techniques tend to underestimate clay content, although a relative measure of the fine fraction may be approximated as 2.5% × 15-bar water content. Table 2 shows texture, 15-bar water content, and sodium-pyrophosphate-extractable Fe3+ and Al3+ determined by the U.S. Soil Survey, Lincoln, Nebraska, for soils sampled near our transects.
We sampled the O and A horizons—the organic surface horizon and the first mineral horizon below it. The O horizon receives plant inputs from leaf litter and roots, while the A horizon gets carbon inputs from roots and decayed organic matter moving down the soil profile. On average the organic matter of the A horizon is older and in a later stage of decay than at the surface. We expected that the influence of site fertility or added nutrients, if any, would be more important in the O horizon, whereas the A horizon might reflect the influence of minerals on carbon decomposition and stabilization.
METHODS
In the unfertilized soils, the soil O and A horizons were sampled from 10 soil pits dug at 10 m intervals along two transects per site. Approximately 500–700 g of material were collected from each pit. Samples were not composited; there were 10 replicates for each analysis described below except radiocarbon content. In the long-term fertilization experiment, soils were collected by corer (10 cm inner diameter) and divided into O and A horizons in the field as soil was removed from the corer. Samples were analyzed from four replicate plots for each fertilization treatment, with each sample a composite of three soil-core collections. Five of the fertilizer treatment plots at the 4,100 ky site lacked either an O or an A horizon. As a result, n = 2 for A horizon N fertilization; n = 3 for A horizon control, A horizon N+P, and O horizon N+P. The date of collection was 5.5–6 months after the last (semiannual) fertilization. Soil samples from both the main study plots and fertilized plots were collected on Hawaii (site ages = 0.3, 2.1, 20, and 150 ky) in July 1996, and from the 1,400 and 4,100 ky sites in October 1996.
After collection, soils were flown in coolers to California and processed within one week. Field-moist soil was sieved at 4 mm to minimize disturbance of soil aggregates. Fine roots and undecomposed leaves were picked out manually. As a result, most of the Oe layer was removed during sieving and manual extraction, leaving predominantly the Oa horizon. Samples were refrigerated until processing and analysis. Total carbon and nitrogen content of ground soils was measured using a Fisons or Carlo Erba elemental analyzer. The soil mineral and texture analyses described above and bulk density were determined for soils sampled in 1994 from a single soil pit at each site, located near the transects sampled for the current analysis (Torn and others 1997; Chadwick and others 1999).
Incubations
Soils were incubated for 9 months. For incubations, field-moist soil (approximately 6 g dry weight) was placed in a 50 mL plastic beaker with drainage holes at the bottom, lined with a glass filter. The beakers were placed in 1 L glass Mason Ball jars and kept in a growth chamber at 16°C, the average annual site temperature. Soils were brought to and maintained at field capacity with periodic additions of de-ionized water.
Flux-measurement periods ranged from 20 h during the first month to 48 h after 6 months. Based on the CO2 concentrations in the jars, oxygen limitation was never approached (oxygen consumed by CO2 production was never more than 1%). Gas samples were analyzed within 24 h with a Shimadzu gas chromatograph and thermal conductivity detector or a closed-loop Li-Cor LI-6200 portable infrared gas analyzer. Between flux-measurement periods, jars were covered with polyethylene film to prevent moisture loss. Before each measurement period, the jars were aired out and then capped with lids fitted with rubber septa for sampling by syringe. Headspace CO2 samples were collected into glass syringes at the beginning and end of each period.
Extractable and Microbial C, N, P
Microbial C, N, and P were measured by the chloroform fumigation–extraction method (CFEM) on sieved, undried splits of each sample within one week of collection. Fumigated soils were exposed to chloroform in a darkened vacuum dessicator for 36–48 h. Approximately 6 g fumigated or unfumigated (control) soil was extracted with 0.5 M K2SO4 for C and N (Brookes and others 1985; Vance and others 1987; Beck and others 1997) and with Bray’s solution for extractable P (Bray and Kurtz 1945), using 50 or 75 mL extractant. Soil solutions were shaken for 1 h and refrigerated overnight before filtering. Extracts were Kjeldahl-digested and analyzed for total N and total P with an Alpkem analyzer. Extract carbon content was measured with a Shimadzu TOC-5050A. Extractable C, N, and P were determined from the control (unfumigated) soils. Microbial C, N, and P were estimated as the difference between the amount in unfumigated and fumigated soils; no conversion for extraction efficiency was used; hence, values represent the relative content of the different soils.
Radiocarbon Analysis
Radiocarbon measurements and modeling were performed for the main study plots but not for the long-term fertilization experiments. The radiocarbon content of SOM in each horizon was determined for a single composite of the 10 soil pits sampled at each site, using a split of the ground soil used for total carbon and nitrogen analysis. Radiocarbon content was measured by accelerator mass spectrometry (AMS) at Lawrence Livermore National Laboratory (LLNL). Results are reported as Δ14C, the per mil deviation from a standard normalized for 13C content (Stuiver and Polach 1977). AMS analytical precision was reported at 5–7‰ for these samples. Graphite AMS targets were prepared at the LLNL Center for AMS via hydrogen reduction (Vogel and others 1984). Compositing and AMS target preparation were repeated using sealed-tube zinc reduction (Vogel 1992) at the University of California, Irvine, for the 150 ky site A horizon and the 20 ky O and A horizons. Radiocarbon values between replicates differed by only 2‰, 3‰, and 6‰, respectively.
The calculation of Δ14C from raw AMS output requires a correction for the sample’s 13C/12C ratio. The 13C content of an aliquot of the CO2 that was isolated for AMS was analyzed with a dual-inlet isotope ratio mass spectrometer, for all samples except the 20 ky site. For this site, the correction used the average 13C value of the other sites.
Turnover Time
Assuming first-order kinetics, the rate of CO2 production by decomposition of organic matter is the product of the quantity of C and its decomposition constant (k): CO2 respired = kC. Turnover time (τ) is the reciprocal of decomposition rate, and thus CO2 respired = C/τ. The CO2 respired from a soil that is represented by three pools of SOM can be expressed as the sum of fluxes from each pool, or
where R = respired CO2 (g C mineralized d−1 g soil−1), C = soil C (g), and subscripts OM = total SOM, a = active, s = slow, and p = passive.
Passive Soil Organic Matter C
Turnover time for the slow pool of SOM was estimated in two ways: from the incubations and from a box model constrained by soil radiocarbon content. Both approaches required estimating the amount of passive C, that is, carbon with a very long turnover time (hundreds to thousands of years). Most passive C is either stabilized by minerals or chemically recalcitrant. The quantity of mineral-stabilized C was estimated using the regression model for passive C versus noncrystalline mineral content generated for these soils by Torn and others (1997), with data shown in Table 2. Mineral-stabilized C was 2.0%–3.6% of total soil C in the O horizon and 2.7%–7.0% in the A horizon. We arbitrarily assumed total passive C to be twice the amount of mineral-stabilized C. Outcomes were not sensitive to this parameterization: Assigning the amount of passive C as either 1 × or 2 × mineral C had only a small effect on the estimated turnover time of the slow pool. Other approaches to estimating the amount of passive C are not generally applicable to Andisols (Falloon and others 1998).
Active Soil Organic Matter C (Active SOM)
We estimated active SOM (substrate that is decomposable in less than one year) in two ways. First, we estimated active SOM from CO2 respired in the incubations. Typically in incubations, the initial respiration rate drops off quickly to a steady baseline level that reflects decomposition of slow-pool carbon (Townsend and others 1997). The C mineralized in the initial pulse above the baseline level is identified as the active SOM. The size of active SOM calculated in this way does not depend on the rate of decline as long as nearly all the active C is mineralized within the time of integration. Operationally, Ca was calculated from the cumulative CO2 respired during the first two months of the incubation:
where t = time (d); R = CO2 respired (g C d−1 g soil−1); and subscripts for C pool are OM = total SOM, a = active, and s = slow. R OM was measured at each time point, and Rs was estimated as described below (that is, we assumed that the baseline level of R, the respiration rate measured after the respiration pulse of active C, was Rs). We selected two months as the integration time because the decay rates for all sites had leveled off by that time. Second, we estimated the minimum amount of active SOM as the sum of microbial carbon plus extractable carbon, since microbial carbon and extractable carbon are included in the active pool of soil models (for example, Parton and others 1987). To estimate microbial biomass, we assumed that chloroform-labile C represented 0.38 of total microbial C (Voroney and others 1993).
Incubation Approach to Modeling Turnover Time and Proportion of Slow C
Theoretically, a time series of respiration rate from an incubation can be fit by a sum of exponential decay terms, where each term represents one of the carbon pools and its turnover time. In practice, although the incubation data from some of the sites could be fit smoothly by this model, this approach yielded no statistical improvement in fit between a one- and a multi-term exponential model. For this reason, we adopted the approach used by Townsend and others (1997) to solve for τ S analytically, by estimating Rs and Cs. Townsend defined Rs as the respiration rate in the latter period of the incubation when the respiration rate has leveled off, which occurs when the active C has burned off and the remaining respiration results from decomposition of the slow-cycling organic matter (Townsend and others 1997). The respiration rate followed this model, with a relatively constant rate (Rs) after 60 days. We calculated Cs by subtracting active and passive C from total C. CP and Ca were approximated as described above.
14C Approach for Modeling Turnover Time of Slow C
The model equations and a full description of the 14C-based modeling approach are given in the Appendix. We estimated the turnover time of slow-pool SOM with a stock-flow model of passive, slow, and active C pools that kept track of C and 14C in annual time steps (Trumbore 1993). The radiocarbon content of SOM is a function of the 14C added in new C inputs and the 14C lost because of decomposition and radioactive decay. Because we can measure or closely estimate the input and radioactive decay terms, it is possible to solve for the decomposition rate, or turnover time. For each sample, we solved for the turnover time of the slow pool at which the model-predicted Δ14C for the whole soil matched what we measured. The model requires parameterization of the amounts and turnover times of active and passive carbon. As described above, we estimated the amount of active C from the microbial biomass and incubation, and the amount of passive C from the mineral content.
Radiocarbon modeling also requires estimating the 14C content of C inputs to the horizon, determined by the year the C was fixed by plants in relation to the year that C entered the soil horizon of interest (the “age of inputs”) (Perruchoud and others 1999). In most published 14C soil studies, the age of inputs is assumed to be the same for all soil horizons and is based on the lifetime of leaves, usually 1–2 y (for example, Townsend and others 1995; Harrison 1996; Trumbore and others 1996).
In fact, C inputs to soil horizons may be significantly older than previous studies have assumed. Inputs to the O horizon are older because (1) leaves must partially decay before being included in the Oa horizon; (2) fine roots live longer than leaves in many forest ecosystems (Ostertag and Hobbie 1999; Gaudinski and others 2000); and (3) twigs, branches, and other long-lived material make an episodic but potentially significant contribution to plant litter. Deeper horizons receive C from roots as well as material leached from above that may be decades old (Trumbore and others 1992).
In the 14C modeling, we took into account the lag time between photosynthesis of C and its incorporation into SOM by calculating the flux-weighted average age of C input, including leaf, root, and dissolved organic carbon (DOC) input sources (see equations in Appendix). For the O horizon, a lag time could be calculated from the root and leaf lifetimes, decay times, and relative input rates. For the A horizon, the age of inputs was not well constrained: Assuming 30% DOC input and the slowest plausible O horizon turnover time, input ages were on the order of 20 years, whereas assuming only 10% input from DOC and fast O horizon turnover, input ages to the A horizon ranged from 8 to 11 years, depending on the site. We chose a “best estimate” scenario, which yielded intermediate ages, to use in the modeling results presented here (Appendix and Table 3).
RESULTS AND DISCUSSION
Total Soil Carbon and Nitrogen
The O horizon had high carbon content, ranging from 15% to 20% at the youngest sites to over 40% at the four oldest sites (Table 4).
In the A horizon, carbon content was more variable but followed roughly the same pattern, with a large increase in carbon concentration between 2.1 ky and 20 ky sites. Compared to O horizon samples, the A horizon had lower concentrations of C and N, and the organic matter had a narrower range of C:N ratios (Table 4).
CO2 Respiration
Over the first two months of the incubation, the daily respiration rate decreased sharply. After 60 days, the respiration rate of nearly all replicates had leveled off and would change very little over the subsequent 7 months. The short-term drop in respiration rate is usually attributed to the exhaustion of the active carbon pool, which is not replenished in incubations (for example, by root exudates). Disturbance of soil structure during collection and processing may also create a pulse of labile C that contributes to the short-term spike in respiration (T. Fries personal communication; Baisden and others 1996). The initial respiration rates in the incubations were higher than soil respiration rates measured in the field (Herbert and Fownes 1999; Torn, data not shown) or ecosystem NPP (Vitousek and others 1997) at each site (multiplying the initial incubation respiration rate by the carbon content per square meter in the surface horizons). The radiocarbon-predicted fluxes (based on turnover times given below) are more commensurate with, but still higher than, in situ soil respiration.
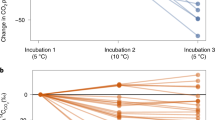
When integrated over the time-course of the incubation, the small but consistent differences in respiration rates among sites added up to large differences in cumulative CO2 evolution (Figure 1). The data are presented per gram of soil C, so that differences in flux rates reflect differences in SOM decomposability, not the amount of organic carbon in each soil. In the O horizon, an intermediate-aged site (150 ky) had greater cumulative emissions than all the other sites, and the oldest site had less respiration than all the other sites (Tukey’s post hoc comparison, P < 0.05). The same relationships held for A horizon soils, except that the 4,100 ky site shared the bottom position with the 20 ky site (P < 0.05). At all sites, the A horizon had much slower emission rates than the O horizon (P < 0.01) except at the youngest site, where this difference was barely significant (P < 0.1) due to one outlier.

Cumulative CO2 evolution from soils of the (a) O horizon and (b) A horizon. Error bars are ± 1 standard error, n = 10. Differences in emissions (g C emitted per g soil C) reflect differences in decomposability rather than in amount of organic carbon. In both horizons, the 150 ky site had greater cumulative respiration than the other sites and the oldest site had the lowest respiration rate. At each site, the A horizon had lower emission rates than the O horizon.
Microbial and Extractable C, N, P
At each site, microbial C, N, and P were higher in the O horizon than in the A horizon (Table 5a and Figure 2a). However, microbial biomass was the same proportion of total SOM in the A and O horizons (Figure 2b). Across sites, microbial biomass C, N, and P were positively correlated with SOM content. Thus, looking across sites or between horizons, total microbial biomass did not vary with availability of mineralizable C or decomposition rate. Table 5a also shows results for extractable C, N, and P from the unfertilized soils.

Microbial biomass C (chloroform labile C) per (a) gram soil and (b) gram soil carbon for O and A horizons. Error bars are ± 1 standard error. Only positive differences between fumigated and unfumigated soils are included in average, n = 5–10. Across horizons and most sites, microbial biomass was a relatively constant proportion of total soil C. Accordingly, total microbial biomass was highest at sites with most SOM and higher in the O than in the A horizon.
Fraction Active Carbon
We estimated the quantity of active C as (1) the sum of microbial and extractable C or (2) the C evolved during the initial period of incubation. The two measures gave different quantities and, in the O horizon, different patterns. Microbial + extractable C ranged from 0.5% to 2% of soil C or 0.1% to 0.6% of total soil (Table 5a). It was distributed in proportion to the total carbon content of the soil, except that active C was a smaller proportion of total C in the 4,100 ky site A horizon compared to other sites. Active carbon estimated via incubation ranged from 0.8% to 7.6% of soil C or 0.2% to 3.5% of soil (Figure 3). In the O horizon, it was distributed in roughly constant proportion to total soil carbon, whereas in the A horizon the total amount of active C (that is, per gram soil) was similar at all sites.

Active C as estimated by fluxes during the initial phase of the incubation. Error bars are ± 1 standard error, n = 10. In the O horizon, active C was distributed in roughly constant proportion to total soil carbon, whereas in the A horizon the total amount of active C (that is, per gram soil) was similar at all sites.
Pool Sizes: Proportions of Active, Slow, and Passive C
Using the active pool as estimated by the incubations, the O horizon on average consisted of about 5% active C, 89% slow C, and 6% passive C. The range in values for slow C across sites was narrow, 88%–91%. The A horizon consisted of about 3% active, 82%–90% slow, and 7%–15% passive C, where the amount of passive C was estimated from the mineral content.
Turnover Time Based on Incubation
Turnover time of the slow pool of SOM varied greatly across the age gradient and between soil horizons (Figure 4). In the O horizon, turnover times ranged from 6.5 to 20 y. Cycling was slower in the A horizon, ranging from 16 to 40 y, but rates in the O and A horizons were positively correlated (r2 = 0.65, P = 0.05). Turnover time was fastest at the 150 ky site in both horizons (Tukey’s post hoc comparison, P < 0.05).

Turnover times of the slow (intermediate) pool organic matter for (a) O horizon and (b) A horizon. Solid circles (•) are incubations, values are average (n = 10) ± 1 standard error. Open circles (○) are for 14C model, with values based on one composite of ten soil pits sampled in 1996. Turnover times varied by threefold across the gradient, with fastest cycling at the sites of highest fertility. Turnover times in the O and A horizons were positively correlated, with slower rates in the A horizon. Both methods predicted a similar pattern across sites and between horizons, but the 14C turnover times were slower at the three youngest sites.
Turnover Time Based on 14C Model
The radiocarbon content of the SOM ranged from 40‰ to 210‰. The positive Δ14C values indicate that all horizons had incorporated “bomb” carbon and were dominated by carbon cycling on decadal-to-100 y time scales (Table 4a). The slow-pool turnover time varied four fold across the gradient, with fastest turnover at sites of highest fertility. Turnover times were 8–41 y (O horizons) and 22–92 y (A horizons) across the gradient sites, based on 14C (Figure 4 and Table 8).
The 14C model can be forced to produce intermediate-pool turnover times as fast as those in the incubations if we increase the amount of passive C. For a sensitivity analysis, we calculated how much passive C the soils would need to make the 14C model match the incubation outcome. In that case, the A horizon soils would contain 12–22% passive C and 75–85% slow C (the range of values coming from the range of sites).
Turnover Times Compared to Other Sites
The results for the chronosequence incubations are similar to turnover times reported by other studies of moist tropical forest. For example, near Manaus, Brazil, the A horizon SOM turnover time in two intermediate cycling pools is on the order of 5–10 and 70–100 y (Telles and others 2003) [turnover time was originally reported to be 6 y (Trumbore 1993) but has been recalculated with an additional time point of 14C data and using a lag time for litter inputs]. Pastures and forests near our 20 ky site have turnover times of 4–9 and 15–30 y in the top 20 cm, respectively (Townsend and others 1995).
Trumbore and others (1996) proposed that C turnover times of well-drained forest soils vary exponentially with MAT. The incubation-based turnover times at the two most productive chronosequence sites fit their model very well (MAT = 16°C, predicted turnover time = 18 y). Our study’s turnover times were slower at the less productive sites or when estimated by radiocarbon. The average turnover time of SOM in our study sites was faster than in cooler temperate and boreal forests (for comparable, well-drained, mineral soils, intermediate pool; for example, O’Brien and Stout 1978; Trumbore and Harden 1997; Perruchoud and others 1999; Gaudinski and others 2000) and similar to that at Calhoun Forest, a temperate forest site with the same temperature (MAT 16°C, turnover time 12–25 y; Harrison and others 1995; Richter and others 1999).
The Age of C Inputs to Progressively Deeper Soil Horizons
An assessment of reasonable root and DOC inputs suggests that C inputs to soil horizons may be much older than the leaf lifetime or 2 y value that most previous isotope studies have used (Feng and others 1999). Improving the ability to estimate input age is important because this parameter has a large influence on modeling results, and incorporating more realistic estimates of C input ages improved the fit of the model to empirical data.
The main control of—and the main uncertainty in estimating—input age to the O horizon is the ratio of inputs from woody litter versus leaf litter. The main determinants of input age to the A horizon are the amount of DOC entering the A horizon compared to root inputs and the sources of DOC. Unfortunately, very little is known about DOC fluxes in most soils.
The age of inputs had a large effect on turnover times, particularly at the two youngest sites. There, our best estimate of A horizon input age was 20–31 y, and, using these input ages, the radiocarbon model produced turnover times that were 2–3 times faster than if inputs were assumed to be 2 y old (Table 6). A recent study of two temperate forests also found that taking into account the residence time of C in foliar and twig litter (before it entered the SOM pools) reduced the radiocarbon-modeled turnover times of SOM by a factor of 2.5 (Perruchoud and others 1999).
For the 14C and incubation models to predict the same turnover time, the age of C inputs to the A horizon would have to be about 30 y (specifically, a lag time of 33 y would make the turnover times of the 14C model match the incubations for A horizons of five of the six sites). The lag time could be this long if DOC were derived mainly from SOM rather than from litter, and/or if woody material were a significant source of C inputs to soil. In fact, annual production of twigs and stem wood is greater than annual production of leaves in these forests (Herbert and Fownes 1999). If there were no accumulation of forest biomass (that is, if woody litterfall were equal to woody productivity), even the inputs to the Oa horizon would be decades old.
Turnover times in the incubations were about half as fast in the A horizon compared to the O horizon at each site, but rates in the two horizons were positively correlated (r2 = 0.65, P = 0.05; Figure 4). We ascribe slower turnover with depth to a combination of two factors. First, the A horizon has more of a mineral matrix to protect organic matter (Oades 1988). Second, recent leaf litter is chemically more labile than the roots and later-stage products that migrate to the A horizon at these sites (Ostertag and Hobbie 1999). Several 14C studies have observed that soil C is older with depth. These results show that both slower turnover times and progressively older inputs (Feng and others 1999) contribute to this trend.
Comparing the Turnover Times Estimated by Incubations and Radiocarbon-Based Modeling
Both incubation and 14C methods for determining turnover time predicted a similar pattern across sites and between horizons (Figure 4), but the 14C model estimated slower turnover times than did the incubations for the youngest sites (correlation between radiocarbon- and incubation-based turnover times for the six sites: A horizon: r2 = 0.79, P = 0.02; O horizon: r2 = 0.42, P = 0.17). In estimating SOM turnover in pastures, Townsend and others (1995) also found that a 14C-based model predicted slower turnover times than did incubations. The divergence occurs in some measure because the two approaches are surveying different facets of ecosystem carbon cycling. Unlike incubations, radiocarbon content reflects the total residence time of carbon in the ecosystem. There are also several methodological reasons why we would expect incubations to yield faster turnover times than a radiocarbon model.
First, sieving soils for incubation may increase the decomposability of SOM (for example, by breaking up aggregates). However, the incubation flux rates follow the same pattern as NPP, suggesting that incubation artifacts are not the dominant source of differences in flux rates among sites. Second, radiocarbon content reflects the decomposition rate under in situ climate and biological conditions, whereas these incubations occurred under artificial and relatively constant moisture and constant temperature conditions. Third, soils are more heterogeneous than we have modeled them. Using only one pool to represent all slow-pool SOM (cycling at intermediate time scales of decades to centuries) in the radiocarbon model yields turnover times that poorly represent the more dynamic fraction of the SOM. Unfortunately, available data are often not adequate to constrain a model that divides the intermediate-cycling material into two or more pools.
Fertilization Treatment
At the 0.3 ky site, where productivity is N-limited, adding N slowed decomposition and adding P increased the rate of decomposition (Figure 5; only the O horizon had statistically significant treatment effects, with P increasing slow-pool turnover and N decreasing it, P < 0.004 and P < 0.06 respectively). The amount of active carbon was lower in the fertilized plots (A horizon: P < 0.01; O horizon: P < 0.06; Figure 5). It also appears that adding phosphorus alone at the 0.3 ky site decreased the amount of active C compared to treatments in which nitrogen was added. An incubation experiment with Oxisol soils from a tropical forest in Costa Rica found that microbial activity was stimulated by P additions, leading to increased utilization of labile C (Cleveland and others 2002), The effect of fertilization on C, N, and P in total organic, extractable, and microbial biomass pools is shown in Tables 5 4b and 5b. At the 4,100 ky site, where P limits plant growth, the N + P treatment had slower turnover time in the A horizon and N reduced the mean amount of active C, but there was no statistically significant effect of fertilization.

Effect of long-term fertilization at the 0.3 and 4,100 ky sites on turnover time of slow C and amount of active C, based on incubations. Error bars are ± 1 standard error, n = 10. Dark bars are O horizon and light bars are A horizon. Fertilization did not affect turnover at the 4,100 ky site, where P limits plant growth. In contrast, at the 0.3 ky site where productivity is N-limited, added N slowed decomposition and P increased it (treatment effects significant in O horizon). The amount of active C in the mineral soil was lower under fertilization.
Mineral Composition and Soil Texture
Soil mineralogy was weakly associated with turnover times in these surface horizons. Total mineral content was positively correlated with turnover time of slow-pool carbon estimated by 14C, significantly for the difference between O and A horizons (r2 = 0.2, P = 0.07 for both horizons; r2 = 0.51, P = 0.11 for O horizon). Noncrystalline mineral content was positively but not significantly correlated with turnover time, in part because our model for calculating turnover time had a prescribed positive relationship between mineral content and passive C. This relationship acted to make the slow-pool turnover time faster with increased noncrystalline content.
Texture, as estimated by traditional sand:silt:clay fractions or 15-bar water content as a proxy for clay-like particle content (with the latter more reliable in Andisols), was not correlated with carbon density or turnover time. Likewise, sodium-pyrophosphate-extractable (free) Fe3+ and Al3+ were not correlated with carbon content or turnover time (Table 2). Turnover times in the O horizon were not correlated with any of these attributes of A horizon mineralogy. One caveat is that the mineral analyses were performed on different soil samples than were used for the incubations or radiocarbon modeling reported here.
IMPLICATIONS
One common model of decomposition depicts transformation of litter to a few ubiquitous classes of organic compounds by carbon-limited microbial communities. Litter degradation is influenced by litter chemistry in this model, but the process of degradation filters all litters to a common set of organic compounds that have less intrinsic variability in turnover time than did the litter substrates (Melillo and others 1989). In this case, litter quality would have only minor influence on the subsequent decomposability of the organic matter that derives from it. In contrast, others hypothesize that SOM chemistry should reflect litter chemistry more closely. For example, Berg (1986) found that the decay rate of SOM depended on litter quality and nutrient availability, especially availability of nitrogen. To investigate these hypotheses, we used the turnover times from the incubation experiment as indicative of the intrinsic decomposability of SOM. The decomposition rate of litter in situ was highly correlated with SOM decomposition in the O horizon (r2 = 0.94, P < 0.03; Figure 6) and moderately correlated in the A horizon [(r2 = 0.53, P < 0.27; litter decay constants taken from Crews and others 1995) and Hobbie and Vitousek (2000)]. The same was true of litter N and P concentrations (Crews and others 1995), which were tightly correlated with SOM decomposition in the O horizon and less so in the A horizon (for N and P, respectively: O horizon: r2 = 0.64, P < 0.20; r2 = 0.75, P < 0.14; and A horizon: r2 = 0.17, P < 0.59; r2 = 0.28, P < 0.47). The small sample size (n = 4 sites) for these analyses may limit the ability to identify statistical significance of the apparent trends. Higher NPP was significantly correlated with faster decomposition rates in both the O and the A horizons (Figure 7)(Vitousek and others 1997).

Turnover time of the slow (intermediate) C pool versus in situ turnover time for surface litter bags (Crews and others 1995; Hobbie and Vitousek 2000b). Solid circles (•) are O horizon and open circles (○) are A horizon. The decomposition rates of litter and SOM were highly correlated in the O horizon and moderately correlated in the A horizon, supporting the model that the quality of litter influences the decomposability of the organic matter it forms.

Turnover time of the slow (intermediate) C pool versus net primary productivity (Vitousek and others 1997). Solid circles (•) are O horizon and open circles (○) are A horizon. Higher NPP was significantly correlated with faster decomposition rates in both horizons. Faster turnover may promote plant growth by increasing nutrient mineralization. The inverse possibility, that higher NPP causes faster turnover, is not supported by the fertilization results.
The strong correlation between litter quality and SOM decay rates supports the model that litter quality influences the decomposability of the organic matter formed from that litter (Berg and others 1995). Possible mechanisms may include the persistence of recalcitrant plant compounds or effects on decomposition pathways. However, we cannot rule out the confounding influence of soil chemistry, fauna, and the microbial community on decomposition of both litter and SOM.
Response of Decomposition Rate to Fertilization
In contrast to the clear pattern in decomposition rate across the fertility gradient, the effect of nutrient additions was weak and inconsistent. Fertilization affected rates of SOM decomposition at the N-limited (0.3 ky) site but not at the P-limited (4,100 ky site. At the N-limited site, fertilization with nitrogen slightly decreased the rate of decomposition, even though productivity was stimulated by nitrogen at this site. An inhibitory effect of nitrogen on litter decay has been observed in several studies reviewed by Fog (1988), and there are some findings of an inhibitory effect on decomposition and soil respiration as well (for example, Kowalenko and others 1978; De Jong and others 1974). The lack of an N or P effect at the 4,100 ky site where P is limiting to NPP may reflect the fact that the site had been receiving fertilizer for only 5 y. In terms of feedbacks, the long-term fertilization experiments gave no evidence that the same nutrient that limits NPP also limits SOM decomposition at the N- or P-limited sites, just as Hobbie and Vitousek (2000a) observed for litter decomposition.
The failure of fertilization to produce the same pattern in decomposition as did the natural fertility gradient is important. This result converges with studies of the effects of nutrient additions on litter decomposition here and elsewhere (Fog 1988; Ostertag and Hobbie 1999; Hobbie and Vitousek 2000a). These point to a fundamental asynchrony in timing and often to an asymmetry in direction and magnitude in the responses of primary productivity and decomposition to nutrient additions. Although plant productivity can respond directly and positively to fertilization within a growing season (Binkley and Hogberg 1997; Shaver and Chapin 1995; Vitousek and Farrington 1997; Tanner and others 1998), most responses of decomposition take much longer to manifest (Berg and Matzner 1997; Perruchoud and others 1999). Moreover, there is often a qualitative difference in that the same nutrient that limits productivity may have a small, neutral, or even a negative effect on decomposition of litter and SOM (Fog 1988; Magill and Aber 1998; Berg 2000; Hobbie and Vitousek 2000a; Michel and Matzner 2002).
On longer time scales, soil fertility may influence decomposition and productivity through changes in species or genotype (Chapin and others 1986; Hobbie 1992). In parallel, plant composition may limit the decomposition response to changes in soil nutrients or CO2 levels (Scholes and others 1997; McMurtrie and others 2000; Sariyildiz and Anderson 2003). The lack of response to fertilization at our sites, for example, may be due to genotypic adaptation to nutrient conditions. A common-garden study found that the different chronosequence sites support genetically different Metrosideros populations with genotypic traits that constrain the responsiveness of plant chemistry, phenol production, and litter quality to fertilization, and in particular to N fertilization (Treseder and Vitousek 2001; Hattenschwiler and others 2003). To the degree that the observed correlations, for example, between litter and SOM decomposition rates, are causative, the response of SOM decomposition to nutrient additions may be limited by genotype or species composition.
References
DW Anderson (1988) ArticleTitleThe effect of parent material and soil development on nutrient cycling in temperate ecosystems Biogeochemistry 5 71–97
W Baisden Y Wang TC Balser RG Amundson (1996) ArticleTitleMeasurement of natural abundance 14C in respired CO2 from laboratory incubations of natural and cultivated grassland soils Bull Ecol Soc Am 77 21
J Balesdent GH Wagner A Mariotti (1988) ArticleTitleSoil organic matter turnover in long-term field experiments as revealed by carbon-13 natural abundance Soil Sci Soc Am J 52 118–124
T Beck RG Joergensen E Kandeler F Makeschin E Nuss HR Oberholzer S Scheu (1997) ArticleTitleAn inter-laboratory comparison of ten different ways of measuring soil microbial biomass C Soil Biol Biochem 29 1023–1032 Occurrence Handle10.1016/S0038-0717(97)00030-8
B Berg (1986) ArticleTitleNutrient release from litter and humus in coniferous forest soils—a mini review Scand J For Res 1 359–69
B Berg (2000) ArticleTitleLitter decomposition and organic matter turnover in northern forest soils For Ecol Manage 133 13–22
B Berg E Matzner (1997) ArticleTitleEffect of N deposition on decomposition of plant litter and soil organic matter in forest systems Environ Rev 5 1–25 Occurrence Handle10.1139/er-5-1-1
B Berg CA McClaugherty A Virzo Santo ParticleDe M–B Johansson G Ekbohm (1995) ArticleTitleDecomposition of litter and soil organic matter—can we distinguish a mechanism for soil organic matter buildup? Scand J For Res 10 108–19
L Beyer (1991) ArticleTitleIntersite characterization and variability of soil respiration in different arable and forest soils Biol Fertil Soils 12 122–6 Occurrence Handle10.1007/BF00341487
VO Biederbeck HH Janzen CA Campbell RP Zentner (1994) ArticleTitleLabile organic matter as influenced by cropping patterns in an arid environment Soil Bio Biochem 26 1647–56 Occurrence Handle10.1016/0038-0717(94)90317-4
D Binkley P Hogberg (1997) ArticleTitleDoes atmospheric deposition of nitrogen threaten Swedish forests? For Ecol Manage 92 119–52
RH Bray LT Kurtz (1945) ArticleTitleDetermination of total, organic, and available forms of phosphorus in soils Soil Sci 59 39–45
PC Brookes A Landman G Pruden DS Jenkinson (1985) ArticleTitleChloroform fumigation and the release of soil nitrogen: a rapid direct extraction method to measure microbial biomass nitrogen in soil Soil Biol Biochem 17 837–42 Occurrence Handle10.1016/0038-0717(85)90144-0
OA Chadwick LA Derry PM Vitousek BJ Huebert LO Hedin (1999) ArticleTitleChanging sources of nutrients during four million years of ecosystem development Nature 397 491–7 Occurrence Handle10.1038/17276
FS Chapin SuffixIII PM Vitousek K VanCleve (1986) ArticleTitleThe nature of nutrient limitation in plant communities Am Naturalist 127 48–58 Occurrence Handle10.1086/284466
C Cleveland AR Townsend SK Schmidt (2002) ArticleTitlePhosphorus limitation of microbial processes in moist tropical forests: Evidence from short-term laboratory incubations and field studies Ecosystems 5 680–91
TE Crews K Kitayama JH Fownes RH Riley DA Herbert D Mueller–Dombois PM Vitousek (1995) ArticleTitleChanges in soil phosphorus fractions and ecosystem dynamics across a long chronosequence in Hawaii Ecology 76 1407–24
E Jong ParticleDe HJV Schappert KB MacDonald (1974) ArticleTitleCarbon dioxide evolution from virgin and cultivated soil as affected by management practices and climate Can. J. Soil Sci 54 299–307
P Falloon P Smith K Coleman S Marshall (1998) ArticleTitleEstimating the size of the inert organic matter pool from total soil organic carbon content for use in the Rothemsted carbon model Soil Biol Biochem 30 1207–11 Occurrence Handle10.1016/S0038-0717(97)00256-3
XH Feng JC Peterson SA Quideau RA Virginia RC Graham LJ Sonder OA Chadwick (1999) ArticleTitleDistribution, accumulation, and fluxes of soil carbon in four monoculture lysimeters at San Dimas Experimental Forest, California Geochim Cosmochim Acta 63 1319–33 Occurrence Handle10.1016/S0016-7037(99)00048-4
K Fog (1988) ArticleTitleThe effect of added nitrogen on the rate of decomposition of organic matter Biol Rev 63 433–62
J Gaudinski SE Trumbore EA Davidson SH Zheng (2000) ArticleTitleSoil carbon cycling in a temperate forest: radiocarbon-based estimates of residence times, sequestration rates and partitioning of fluxes Biogeochemistry 51 33–69 Occurrence Handle10.1023/A:1006301010014
SR Gupta JS Singh (1981) ArticleTitleSoil respiration in a tropical grassland Soil Biol Biochem 13 261–8 Occurrence Handle10.1016/0038-0717(81)90060-2
KG Harrison (1996) ArticleTitleUsing bulk soil radiocarbon measurements to estimate soil organic matter turnover times: Implications for atmospheric CO2 levels Radiocarbon 38 181–90
KG Harrison WS Broecker G Bonani (1993) ArticleTitleThe effect of changing land use on soil Radiocarbon Science 262 725–9
KG Harrison WM Post DD Richter (1995) ArticleTitleSoil carbon turnover in a recovering temperate forest Global Biogeochem Cycles 9 449–54 Occurrence Handle10.1029/95GB02380
J Hassink (1994) ArticleTitleEffect of soil texture on the size of the microbial biomass and on the amount of C and N mineralized per unit of microbial biomass in Dutch grassland soils Soil Biol Biochem 26 1573–82 Occurrence Handle10.1016/0038-0717(94)90100-7
S Hattenschwiler AE Hagerman PM Vitousek (2003) ArticleTitlePolyphenols in litter from tropical montane forests across a wide range in soil fertility Biogeochemistry 64 129–148 Occurrence Handle10.1023/A:1024966026225
D Herbert JH Fownes (1995) ArticleTitlePhosphorus limitation of forest leaf area and net primary production on a highly weathered soil Biogeochemistry 29 223–35
DA Herbert JH Fownes (1999) ArticleTitleForest productivity and efficiency of resource use across a chronosequence of tropical montane soils Ecosystems 2 242–54 Occurrence Handle10.1007/s100219900072
SE Hobbie (1992) ArticleTitleEffects of plant species on nutrient cycling Trends Ecol Evol 7 336–9 Occurrence Handle10.1016/0169-5347(92)90126-V
SE Hobbie PM Vitousek (2000a) ArticleTitleNutrient limitation of litter decomposition in Hawaiian forests Ecology 81 1867–77
SE Hobbie PM Vitousek (2000b) ArticleTitleNutrient regulation of litter decomposition in Hawaiian forests Ecology 81 1867–77
DM Howard PJA Howard (1993) ArticleTitleRelationships between CO2 evolution, moisture content and temperature for a range of soil types Soil Biol Biochem 25 1537–46 Occurrence Handle10.1016/0038-0717(93)90008-Y
DS Jenkinson (1990) ArticleTitleThe turnover of organic carbon and nitrogen in soil Philos Trans R Soc Lond B Biol Sci 329 361–8
K Kitayama D Mueller–Dombois (1995) ArticleTitleVegetation changes along gradients of long-term soil development in the Hawaiian montane rainforest zone Vegetatio 120 1–20
CG Kowalenko KC Ivarson DR Cameron (1978) ArticleTitleEffect of moisture content, temperature and nitrogen fertilization on carbon dioxide evolution from field soils Soil Boil. Biochem 10 417–423 Occurrence Handle10.1016/0038-0717(78)90068-8
DM Linn JW Doran (1984) ArticleTitleEffect of water-filled pore space on carbon dioxide and nitrous oxide production in tilled and non-tilled soils Soil Sci Soc Am J 48 1267–72
AH Magill JD Aber (1998) ArticleTitleLong-term effects of experimental nitrogen additions on foliar litter decay and humus formation in forest ecosystems Plant Soil 203 301–11 Occurrence Handle10.1023/A:1004367000041
RE McMurtrie RC Dewar BE Medlyn MP Jeffreys (2000) ArticleTitleEffects of elevated [CO2] on forest growth and carbon storage: A modelling analysis of the consequences of changes in litter quality/quantity and root exudation Plant Soil 224 135–52 Occurrence Handle10.1023/A:1004711707787
JM Melillo JD Aber AE Linkins A Ricca B Fry KJ Nadelhoffer (1989) ArticleTitleCarbon and nitrogen dynamics along the decay continuum: plant litter to soil organic matter Plant Soil 115 189–98
JM Melillo JD Aber JF Muratore (1982) ArticleTitleNitrogen and lignin control of hardwood leaf litter decomposition dynamics Ecology 63 621–6
K Michel E Matzner (2002) ArticleTitleNitrogen content of forest floor Oa layers affects carbon pathways and nitrogen mineralization Soil Biol Biochem 34 1807–13 Occurrence Handle10.1016/S0038-0717(02)00170-0
KJ Nadelhoffer AE Giblin GR Shaver JA Laundre (1991) ArticleTitleEffects of temperature and substrate quality on element mineralization in six arctic soils Ecology 72 242–53
M Nanzyo R Dahlgren S Shoji (1993) Physical characteristics of volcanic ash soils S Shoji (Eds) Volcanic Ash Soils, Genesis, Properties, and Utilization Elsevier New York 189–207
WD Nettleton SH Brownfield R Burt EC Benhem SL Baird K Hipple CL McGrath HR Sinclair (1999) ArticleTitleReliability of andisol field texture clay estimates Soil Survey Horizons 40 36–49
O’Brien BJ, Stout JD. 1978. Movement and turnover of soil organic matter as indicated by carbon isotope measurements. 10: 309 7
JM Oades (1988) ArticleTitleThe retention of organic matter in soils Biogeochemistry 5 35–70
R Ostertag (2001) ArticleTitleThe effects of nitrogen and phosphorus availability on fine root dynamics in Hawaiian montane forests Ecology 82 485–99
R Ostertag SE Hobbie (1999) ArticleTitleEarly stages of root and leaf decomposition in Hawaiian forests: effects of nutrient availability Oecologia 121 564–73 Occurrence Handle10.1007/s004420050963
W Parton DS Schimel CV Cole DS Ojima (1987) ArticleTitleAnalysis of factors controlling soil organic matter levels in Great Plains grasslands Soil Sci Am J 51 1173–9
D Perruchoud F Joos A Fischlin I Hajdas G Bonani (1999) ArticleTitleEvaluating timescales of carbon turnover in temperate forest soils with radiocarbon data Global Biogeochem Cycles 13 555–73 Occurrence Handle10.1029/1999GB900003
JW Raich WH Schlesinger (1992) ArticleTitleThe global carbon dioxide flux in soil respiration and its relationship to vegetation and climate Tellus 44 81–99 Occurrence Handle10.1034/j.1600-0889.1992.t01-1-00001.x
DD Richter D Markewitz SE Trumbore CG Wells (1999) ArticleTitleRapid accumulation and turnover of soil carbon in a re-establishing forest Nature 6739 56–8 Occurrence Handle10.1038/21867
S Saggar KR Tate CW Feltham CW Childs A Parshotam (1994) ArticleTitleCarbon turnover in a range of allophanic soils amended with 14C-labelled glucose Soil Biol Biochem 26 1263–71 Occurrence Handle10.1016/0038-0717(94)90152-X
T Sariyildiz JM Anderson (2003) ArticleTitleInteractions between litter quality, decomposition and soil fertility: A laboratory study Soil Biol Biochem 35 391–9 Occurrence Handle10.1016/S0038-0717(02)00290-0
DS Schimel BH Brasswell E Holland R McKeown S Ojima TH Painter WJ Parton AR Townsend (1994) ArticleTitleClimatic, edaphic, and biotic controls over storage and turnover of carbon in soils Global Biogeochem Cycles 8 279–94 Occurrence Handle10.1029/94GB00993
MC Scholes D Powlson G Tian (1997) ArticleTitleInput control of organic matter dynamics Geoderma 79 25–47 Occurrence Handle10.1016/S0016-7061(97)00037-2
GR Shaver FS Chapin SuffixIII (1995) ArticleTitleLong-term responses to factorial, NPK fertilizer treatment by Alaskan wet and moist tundra sedge species Ecography 18 259–75
JS Singh SR Gupta (1977) ArticleTitlePlant decomposition and soil respiration in terrestrial ecosystems 3 449–528
M Stuiver H Polach (1977) ArticleTitleReporting of 14C data Radiocarbon 19 355–63
MJ Swift OW Heal JM Anderson (1979) Decomposition in terrestrial ecosystems Blackwell Scientific Oxford 372
EVJ Tanner PM Vitousek E Cuevas (1998) ArticleTitleExperimental investigation of nutrient limitation of forest growth on wet tropical mountains Ecology 79 10–22
EDC Telles PB Camargo Particlede LA Martinelli SE Trumbore ES da Costa J Santos N Higuchi RC Oliveira (2003) ArticleTitleInfluence of soil texture on carbon dynamics and storage potential in tropical forest soils of Amazonia Global Biogeochem Cycles 17 1040 Occurrence Handle10.1029/2002GB001953
MS Torn SE Trumbore OA Chadwick PM Vitousek DM Hendricks (1997) ArticleTitleMineral control of soil organic carbon cycling Nature 389 170–3 Occurrence Handle10.1038/38260
MS Torn C Masiello I Basile–Doelsch N Bijoor R Sutton (2002) Mineral control of carbon storage in Andisols: Case studies and applications to other soils O Arnalds (Eds) Soil Resources of European Volcanic Systems: Mineralogy related features and processes common to European Andosols European Cooperation in the field of Scientific and Technical Research (COST) Manderscheid, Germany 7–8
AR Townsend PM Vitousek SE Trumbore (1995) ArticleTitleSoil organic matter dynamics along gradients in temperature and land-use Ecology 76 721–733
AR Townsend PM Vitousek DJ DesMarais A Tharpe (1997) ArticleTitleSoil carbon pool structure and temperature sensitivity inferred using CO2 and 13CO2 incubation fluxes from five Hawaiian soils Biogeochemistry 38 1–17 Occurrence Handle10.1023/A:1017942918708
KK Treseder PM Vitousek (2001) ArticleTitlePotential ecosystem-level effects of genetic variation among populations of Metrosideros polymorpha from a soil fertility gradient in Hawaii Oecologia 126 266–75 Occurrence Handle10.1007/s004420000523
SE Trumbore (1993) ArticleTitleComparison of carbon dynamics in tropical and temperate soils using radiocarbon measurements Global Biogeochem Cycles 7 275–90
Trumbore SE. 1997. Potential responses of soil organic carbon to global environmental change. In: Keeling D, Eds. Carbon Dioxide and Climate Change. Proc Nat Acad Sci USA 94:8284–91
SE Trumbore (2000) ArticleTitleAge of soil organic matter and soil respiration: Radiocarbon constraints on belowground C dynamics Ecol Appl 10 399–411
SE Trumbore JW Harden (1997) ArticleTitleAccumulation and turnover of carbon in organic and mineral soils of the BOREAS northern study area J Geophys Res 102 28817–30 Occurrence Handle10.1029/97JD02231
SE Trumbore SL Schiff R Aravena R Elgood (1992) ArticleTitleSources and transformation of dissolved organic carbon in the Harp Lake forested catchment: the role of soils Radiocarbon 34 626–35
SE Trumbore OA Chadwick R Amundson (1996) ArticleTitleRapid exchange between soil carbon and atmospheric carbon dioxide driven by temperature change Science 272 393–6
Trumbore SE, Torn MS. 2004. Soils and the global carbon cycle.In: Holland EA, Eds,. Notes from Underground: Soils and Global Change. NATO Advanced Study Institute http://esd.lbl.gov/ESD_staff/torn/nato_soilcarbon.pdf
ED Vance PC Brookes DS Jenkinson (1987) ArticleTitleAn extraction method for measuring soil microbial biomass C Soil Biol Biochem 19 703–7 Occurrence Handle10.1016/0038-0717(87)90052-6
E Veldkamp (1994) ArticleTitleOrganic carbon turnover in 3 tropical soils under pasture after deforestation Soil Sci Am J 58 175–80
PM Vitousek H Farrington (1997) ArticleTitleNutrient limitation and soil development: Experimental test of a biogeochemical theory Biogeochemistry 37 63–75 Occurrence Handle10.1023/A:1005757218475
PM Vitousek OA Chadwick TE Crews JH Fownes DM Hendricks D Herbert (1997) ArticleTitleSoil and ecosystem development across the Hawaiian Islands GSA Today 7 1–7 Occurrence Handle11541665
PM Vitousek LR Walker LD Whiteaker PA Matson (1993) ArticleTitleNutrient limitations to plant growth during primary succession in Hawaii Volcanoes National Park Biogeochemistry 23 197–215 Occurrence Handle10.1007/BF00023752
PM Vitousek DR Turner WJ Parton RL Sanford (1994) ArticleTitleLitter decomposition on the Mauna Loa environmental matrix, Hawai’i: patterns, mechanisms, and models Ecology 75 418–29
PM Vitousek DR Turner K Kitayama (1995) ArticleTitleFoliar nutrients during long soil development in Hawaiian montane rain forest Ecology 76 712–20
JS Vogel (1992) ArticleTitleA rapid method for preparation of biomedical targets for AMS Radiocarbon 34 344–50
JS Vogel JR Southon DE Nelson TA Brown (1984) ArticleTitlePerformance of catalytically condensed carbon for use in Accelerator Mass Spectrometry Nucl Instrum Meth B5 289–93
RP Voroney JP Winter RP Beyaert (1993) Soil microbial biomass C and N MR Carter (Eds) Soil Sampling and Methods of Analysis Lewis Publishers Boca Raton FL 277–86
Acknowledgments
We thank D. Turner, T. Macomber, J.H. Williams, and F.S. Chapin III for help in the laboratory and access to lab equipment; H. Farrington, J. Moen, J. Gaudinski, and O. Chadwick for help with field sampling; USDA, Mellon Foundation, National Science Foundation Grant No. 9618803, U.S. Department of Energy Office of Science under Contract No. DE-AC03-765F00098, and Joan Irvine Trust for financial support; Lawrence Livermore National Laboratory’s Center for Accelerator Mass Spectrometry for radiocarbon analysis; and The Nature Conservancy, U.S. National Park Service, Parker Ranch, Hawaii Division of Forestry and Wildlife, Hawaii Division of State Parks, and the Joseph Souza Center for access to field sites. We thank C. Castanha, S.E. Hobbie, J.C. Neff, R. Ostertag, and W.J. Riley for helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Appendix: 14C Approach for Modeling the Turnover Time of the Slow (Intermediate) C Pool
Appendix: 14C Approach for Modeling the Turnover Time of the Slow (Intermediate) C Pool
We estimated the turnover time of slow-pool SOM with a stock-flow model of passive, slow, and active C pools that kept track of C and 14C in annual time steps (Trumbore 1993). In Equation, (A1) the radiocarbon content of SOM in a given year is the content in the previous year plus the difference between the 14C added in plant inputs and the 14C lost in decomposition and radioactive decay.
where F = radiocarbon content of soil organic C (14C fraction modern); C = soil organic C (g C); Δt is one year; F I = radiocarbon content of plant inputs (14C fraction modern); I = inputs (g C y−1); τ = turnover time of SOM (y); and λ = 14C radioactive decay constant (0.000121 y−1). Using an explicit formulation (i.e., decay and decomposition are a function of Ft−ΔtCt−Δt rather than F t C t ) had a negligible effect on results.
where the symbols are as defined above with subscripts for C pool as follows: a = active, s = slow, and p = passive. In modeling each site and horizon, we solved for the turnover time of the slow pool at which the model-predicted Δ14C for the whole soil matched what we measured. The model requires parameterization of the amounts and turnover times of active and passive carbon. We used the following parameters: For all sites, the turnover time of the active SOM pool was set to 1 y (Parton and others 1987); the amount of active SOM was set to 3%, which was between the incubation-based and extraction-based estimates. As described for incubations, the amount of passive carbon was estimated as twice the mineral-stabilized C. For the turnover time of passive C, we used the turnover time of the deepest horizon at the site (Harrison and others 1993) estimated at these sites by Torn and others (1997) up to a maximum of 11,000 y. A sensitivity analysis using turnover times of 810, 2,000, and 11,000 y (bracketing the fastest and slowest values among sites) showed only a small effect except at the 20 ky site, which had the highest mineral content and an estimated 14% passive C.
Radiocarbon modeling requires estimating the 14C content of the C inputs to the soil, which is determined by the age of inputs (that is, the year that photosynthesis fixed the C from atmosphere relative to the year the inputs enter the soil). For example, if inputs to the O horizon have an age of 2 y, it means that inputs to that soil horizon in 1996 are assumed to have the 14C value of the atmosphere in 1994. In modeling with 14C, we took into account the lag time between photosynthesis and C input to SOM. We use the average residence time of all inputs to a given horizon, henceforth called the average age of inputs, in modeling.
The average age of C inputs to the O horizon [Eq. (A3)] and the A horizon [Eq. (A4)] were calculated as the mass-weighted average for the C coming from leaf, root, and dissolved organic carbon (DOC) sources.
where A = age (y) of the specified C input (that is, the number of years C spends in the specified C reservoir); I = inputs (g C m2 y−1) of the specified input; O = O horizon, A = A horizon; L = leaves, LL = leaf litter; R = roots, RL = root litter, D = DOC). As shown in Eqs. (A3) and (A4), the age of leaf or root inputs is the sum of the time spent as live tissue plus as litter before decaying into SOM. The amount of time spent as litter (ALL or ARL) before entering SOM is estimated as one-half litter turnover time [where litter turnover time is 1/k, where k is decay rate (y−1)]. We were able to build upon empirical measurements of biomass, lifetime, and decay rates for both leaves and roots for many of the study sites (Crews and others 1995; Herbert and Fownes 1999; Ostertag and Hobbie 1999; Hobbie and Vitousek 2000a). These data and our estimate of input age are summarized in Table 3. The root biomass data comes from cores that sampled the combined O and A horizons (Ostertag and Hobbie 1999). We apportioned root biomass inputs into each horizon in proportion to the horizon’s relative depth interval. The age of leaf litter used in Eq. (A3) is half the litter turnover time in Table 3 (that is, we assume leaf litter is more than half-decayed by the time it is counted as OM rather than litter). For the second and fifth site on the age sequence, we had little of the needed data and interpolated between the closest (in age and location) sites.
For the O horizon of most of the sites, there were two turnover times that would predict SOM with the observed 14C value, because of nonlinearity in the atmospheric 14C record of the past 5 decades (Trumbore 2000). Where this occurred, the two turnover times differed by at least twofold and consequently implied very different C stocks and respiration rates (since flux = stock/τ). By comparing the implied stocks and fluxes to measurements, we were able to rule out one of the possible turnover times (Table 6). First, we compared the measured stock and NPP inputs to the implied stock. Second, we compared the predicted respiration from the O horizon to the measurement of total soil respiration at that site. In all but one case, the comparisons eliminated one of the possible turnover times.
In the A horizon, carbon is input via roots, DOC, and bioturbation. We had no data on the amount or age of DOC moving down from the surface, but we constructed scenarios to explore the implications of reasonable assumptions about the amount and age of DOC inputs. Bioturbation was not treated. The scenarios were as follows:
-
1
DOC inputs to the A horizon were (a) 10% or (b) 30% of C mineralized in the O horizon.
-
2
The age of DOC inputs to A horizon was (a) the age of material entering the O horizon SOM from leaves and roots or (b) the age of material leaving the O horizon (that is, the age of litter inputs plus the turnover time in the O horizon).
-
3
The O horizon turnover time was the (a) faster of the two possible turnover times or (b) slower of the two possible turnover times.
For the O horizon, the lag time could be calculated from empirical estimates of the root and leaf lifetimes, litter decay rates, and input rates.
For the A horizon, the age of inputs was not well constrained; assuming 30% DOC input and the slowest O horizon turnover time yielded input ages on the order of 20 y. Conversely, assuming only 10% input from DOC and fast O horizon turnover, input ages to the A horizon ranged from 8 to 11 y, depending on the site. Considering all scenarios and sites, the estimated age of inputs to the A horizon varied from 4.7 to 32.6 y. We identified an intermediate estimate (“best estimate”) based on a scenario with 1b, 2b, and the O horizon turnover times that gave the best fit to the data, according to the method described two paragraphs above and shown in Table 6. The reported estimate of turnover time for the A horizon (Figure 4 and Table 6) is based on this “best estimate” for age of inputs. We also calculated turnovers times based on an input age of 2 y for a sensitivity analysis and comparison with previously published radiocarbon studies (Table 6).
Using an age calculated as the mass-weighted average of the age of all input sources to estimate the 14C of inputs is a simplification. The complete solution is to convolve, annually, the 14C content and mass flux of each carbon input to the ecosystem. The simplification was adequate for this sensitivity analysis and for scenarios in which the dates of origin are in the period of roughly linear yearly change in atmospheric 14C content (that is, do not extend earlier than about 20 y).
Rights and permissions
About this article
Cite this article
Torn, M.S., Vitousek, P.M. & Trumbore, S.E. The Influence of Nutrient Availability on Soil Organic Matter Turnover Estimated by Incubations and Radiocarbon Modeling. Ecosystems 8, 352–372 (2005). https://doi.org/10.1007/s10021-004-0259-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10021-004-0259-8