
Abstract
The protective effect of taurine against doxorubicin-induced testicular oxidative stress and apoptosis was investigated in rats. Male rats 8 weeks of age were treated with doxorubicin alone (3 mg/kg, i.p. every other day for 3 doses), taurine alone (150 mg/kg, i.p. every other day for 3 doses) or taurine plus doxorubicin (each dose given 1 day post-taurine). After 28 days, rat testes were collected and analysed. Rats treated with doxorubicin alone displayed reduced body and testicular weights, decreased sperm counts, increased the extent of testicular toxicity (as evident from the decreased activity of testicular marker enzyme, SDH) and oxidative stress (reduced GSH, increased GSSG and MDA levels), decreased antioxidant (SOD, CAT, GST, GPx, GR) and membrane-bound (Na+–K+ and Ca2+ ATPases) enzyme activities as well as plasma testosterone. Reverse transcriptase-PCR analysis revealed that doxorubicin induced a marked decrease in the expression of key enzymes for testicular androgenesis (3β-HSD, 17β-HSD) and testicular steroidogenic acute regulatory (StAR) protein. Western blot analysis showed that doxorubicin administration markedly increased the levels of caspase-9, 3, -8, -12, Fas, Bid and disturbed the Bcl-2 family protein balance. These results suggest that doxorubicin can trigger intrinsic, extrinsic and endoplasmic reticulum-associated apoptotic pathways in testicular pathophysiology. Doxorubicin also triggered activation of JNK, p38MAP kinases and p53. However, taurine could effectively prevent nearly all of these doxorubicin-induced testicular abnormalities, thereby proving to be an effective cytoprotectant.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Doxorubicin (DOX), an anthracycline antibiotic, is a widely used anticancer agent. In spite of its high antitumor efficacy, the use of doxorubicin in clinical chemotherapy is limited due to diverse toxicities, including testicular toxicity. It alters sperm development, production, structural integrity and motility rates in association with increased cellular apoptosis (Herman and Ferrans 1998; Au and Hsu 1980; Lui et al. 1986). The mechanism, responsible for the DOX-induced testicular toxicity is not yet fully clear, but findings from recent studies strongly suggest oxidative stress (Goodman and Hochstein 1977; Atessahin et al. 2006; Yeh et al. 2007) and cellular apoptosis (Shinoda et al. 1999) as being major causes for this pathophysiology. Increased oxidative damage to sperm membranes, proteins and DNA is associated with alterations in signal transduction mechanisms that can be detrimental to male fertility (Sikka 1996). Thus, the combination of the drug delivery together with a potent antioxidant may be the appropriate approach to reduce the toxic side effects of DOX.
As a non-toxic endogenous antioxidant, taurine has become an attractive candidate for attenuating various toxin- and drug-induced pathophysiological conditions (Holloway et al. 1999; Das et al. 2008, 2009a, b, 2010a, b, c, 2011; Ghosh et al. 2009; Manna et al. 2008a, b, 2009; Roy et al. 2009; Sinha et al. 2007, 2008a, b, 2009). Taurine (2-aminoethanesulfonic acid) is a free β-amino acid and is present in high concentrations in several cell types. Two sources of taurine could account for the intracellular levels of this amino acid: a biosynthetic route from cysteine and/or the specific uptake from the extracellular space. Extracellular taurine comes from dietary sources or from some tissues in which it is synthesized (e.g. liver) and released into circulation (Wright et al. 1986; Huxtable 1989, 1992). In the male reproductive system, taurine has been identified as the major free amino acid of sperm cells (Van Der Horst and Grooten 1966). Taurine may act as an antioxidant in preventing sperm lipid peroxidation (Alvarez and Storey 1983) as a capacitating agent (Meizel et al. 1980) and as a sperm motility factor (Fraser 1986). It has been suggested that taurine inhibits plasma membrane Na+, K+-ATPase in membrane homogenates from epididymal hamster sperm cells (Mrsny and Meizel 1985). Taurine could also modify sperm phospholipid methyltransferase activity (Llanos and Ronco 1994). These studies indicate that taurine might also possess protective effects on DOX-induced testicular damage. To test this hypothesis, we established the testes injury model of rats by treating them with DOX and investigated whether administration of taurine could attenuate this injury.
We also investigated the mechanistic aspect of DOX toxicity and its protection by taurine in rat testes. To ascertain the possible mechanism of testicular toxicity following DOX administration, the serum testosterone level and expressions of testicular key androgenic enzymes (like 3β-HSD, 17β-HSD and StAR) were investigated. Depending on the dose of doxorubicin and model system used (i.e. cell culture or animal model), several pathways of apoptosis have been implicated in the induction of cell death (Arola et al. 2000; Muller et al. 1998), including the mitochondrial pathway, the extrinsic receptor-mediated pathway and the endoplasmic/sarcoplasmic reticulum (ER/SR) pathways. In the mitochondrial-dependent pathway, mitochondrial dysfunction can lead to the release of mitochondrial cytochrome c (Pollack and Leeuwenburgh 2001; Green 2000) into the cytoplasm. Cytochrome c, along with Apaf-1, caspase-9, and dATP, may then form a complex called the apoptosome that activates caspase-3, a key cell death protease (Beere et al. 2000). Activation of the extrinsic receptor-mediated pathway leads to cause apoptosis via Fas-mediated activation of caspase-8, which then cleaves and activates procaspase-3 to initiate the caspase cascade (Sun et al. 1999). ER/SR-mediated pathway of apoptosis is associated with the activation of caspase 12, which resides in the ER and is activated on ER stress, including free radicals and disturbances of the intracellular calcium level (Lamkanfi et al. 2004; Nakagawa et al. 2000). In the present study, we, therefore, investigated all the possible pathways through which apoptosis can occur during DOX-induced testicular apoptosis. To evaluate the cell signalling pathways involved in the protective role of taurine against DOX-induced testicular injury, we further investigated the roles of p53 and MAPKinases (p38 and JNK) because these are the well-known proapoptotic signalling molecules responsible for DOX-induced organ-toxicity.
Finally, we conducted histological studies and investigated the mode of cell death by DNA gel electrophoresis in DOX-induced testicular pathophysiology and its protection by taurine.
Materials and methods
Chemicals
Taurine (2-aminoethane sulfonic acid), doxorubicin, anti-JNK, anti-p38, anti-Bcl-2, anti-Bad, anti-caspase 3, anti-caspase 9, anti-caspase 12, anti-caspase 8 and anti-p53 antibodies were purchased from Sigma-Aldrich Chemical Company (St. Louis, USA).
Animals
Swiss albino adult male rats weighing approximately 120–130 g were purchased from M/S Gosh Enterprises, Kolkata, India. Animals were acclimatized under laboratory condition for 4 weeks prior to the experiments. All the experiments with animals were carried out according to the guidelines of the institutional animal ethical committee and full details of the study was approved by the CPCSEA, Ministry of Environment & Forests, New Delhi, India (the permit number is: 95/99/CPCSEA). Eight-week-old rats, equivalent to humans at the age of young adulthood (Quinn 2005), were randomly assigned to four groups as described below according to the pharmacological treatment they received.
Determination of dose-dependent activity of taurine by sorbitol dehydrogenase (SDH) assay
For this study, rats were randomly distributed into eight groups each consisting of six animals. The first two groups serving as normal control (received only water as vehicle) and toxin control received doxorubicin (DOX) in three equal doses (3 mg/kg body weight, i.p. on alternate days). Remaining six groups of animals were treated with six different doses of taurine (50, 75, 100, 150, 200 and 250 mg/kg body weight in distilled water, orally on alternate days) followed by doxorubicin (each injection was given 1 day after taurine).
On day 28 after the first dose of normal saline, doxorubicin or taurine, all rats were killed. SDH activities were measured using serum of all experimental rats.
Experimental design for in vivo treatments
Experimental design needed for the present in vivo study has been summarised as follows: 8-week-old rats were randomly assigned to four groups. Rats in “Normal group” received only water as vehicle. Rats in “DOX-treated group” received doxorubicin (DOX) in three equal doses (3 mg/kg body weight, i.p.) (Jahnukainen et al. 2001) on alternate days for a cumulative dose of 9 mg/kg body weight. Rats in “Taurine-treated group” received taurine in three equal doses (150 mg/kg body weight in distilled water, orally) on alternate days for a cumulative dose of 450 mg/kg body weight. Rats in “Taurine–Dox group” received the same dose of taurine as the “Taurine-treated group”, followed by an identical dose of Dox, as the “Dox group” (each dose was given 1 day after taurine). The study design was summarized in Fig. 1.

Schematic diagram of the experimental protocol
On day 28 after the first dose of normal saline, doxorubicin or taurine, the animals were killed under light ether anaesthesia and testes were collected.
Harvest of testicular samples
The rats were killed on day 28 after the first administration of medication (normal water, doxorubicin or taurine). The body weight (taken before) and testes weight were measured and compared between groups. Blood samples were drawn from the caudal vena cava, collected in test tubes containing heparin solution, and centrifuged at 1,500 g for 10 min to obtain serum. The testes tissues were either fixed in 10% formalin for histopathologic examinations or stored at −80°C till later analysis.
Determination of testes weight to body weight ratio
After killing, the testes from experimental animals were quickly excised and weighed. Then the ratio of testes weight to body weight was measured for each.
Sperm abnormality determination, sperm count and motility
For sperm count, sperm abnormality and sperm motility studies, the epididymis was minced with anatomic scissors, a sperm suspension was prepared in phosphate buffer saline (PBS) and centrifuged at 1,000 rpm for 1 min. Sperm smears were drawn on clean and grease-free slides, dried overnight and then stained with 10% Giemsa (diluted with Sorresons buffer, pH 7.0) for 1 h and subsequently observed. Morphologically abnormal sperm was identified and recorded as per Wyrobek and Bruce (1975). The same suspension was used for sperm counting and motility using a haemocytometer.
Preparation of testicular tissue homogenates
The testes were minced, washed, and homogenized in a Dounce glass homogenizer in 10 mM HEPES–KOH/1 mM EGTA buffer (pH 7.5) containing 250 mM sucrose and supplemented with protease and phosphatase inhibitors. The homogenates were spun down for 10 min at 2,000g at 4°C to discard the debris. The supernatant was collected and used for the in vivo experiments.
Determination of protein content
The protein content of the experimental samples was measured by the method of Bradford (1976) using crystalline BSA as standard.
Biochemical analyses
Specific markers related to testicular dysfunction, e.g. sorbitol dehydrogenase (SDH) and Glucose-6-phosphate dehydrogenase (G6PD) levels in the tissues were estimated by the methods of Bergmeyer (1974) and Das et al. (2010a). SDH is an enzyme which catalyses the reversible oxidation–reduction reaction, involving the interconversion of fructose and sorbitol. The rate of oxidation of NADH is directly proportional to the rate of conversion of d-fructose to d-sorbitol. The rate of decrease in absorbance at 340 nm allows measurement of SDH activity.
G6PD is a cytosolic enzyme which maintains the level of NADPH. The NADPH in turn maintains the level of glutathione in cells. G6PD activity was measured by the reduction of 2 mM NADP+ by G6PD, in the presence of glucose 6-phosphate. The enzyme activity was measured by monitoring the increase in absorption at 340 nm.
Lipid peroxidation was estimated according to the method of Das et al. (2009a, b). Lipid peroxidation refers to the oxidative degradation of lipids. Reactive oxygen species degrade polyunsaturated lipids, forming malondialdehyde (MDA). MDA condense with two equivalents of thiobarbituric acid (TBA) to give a fluorescent red derivative that can be assayed spectrophotometrically at 535 nm.
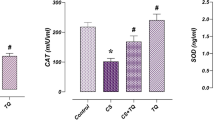
Activities of antioxidant enzymes (SOD, CAT, GST, GR, GPx) in the testes tissue were determined following the methods as described by Das et al. (2009a, b). Superoxide ions (O2−) catalyses the dismutation of superoxide into oxygen and hydrogen peroxide, converts NBT to NBT-diformazan, which absorbs light at 560 nm. SOD reduces the superoxide ion concentration and thereby lowers the rate of NBT-diformazan formation.
CAT catalyzes the decomposition of hydrogen peroxide to water and oxygen; the disappearance of peroxide is followed spectrophotometrically at 240 nm.
GST catalyses the conjugation of 1-chloro-2,4-dinitrobenzene (CDNB) with reduced glutathione. The conjugation is accompanied by an increase in absorbance at 340 nm.
GR reduces glutathione disulfide (GSSG) to the sulfhydryl form GSH, which conjugates with 5,5′-dithiobis(2-nitrobenzoic acid) (DTNB). The conjugation is accompanied by an increase in absorbance at 412 nm.
GPx catalyses the reduction of hydroperoxide, including hydrogen peroxide to water using reduced glutathione. Oxidized glutathione (GSSG), produced upon reduction of hydroperoxide by GPx, is recycled to its reduced state by GR and NADPH. The oxidation of NADPH to NADP+ is accompanied by a decrease in absorbance at 340 nm.
Levels of cellular metabolites (GSH and GSSG) in the testes tissue were also determined following the methods as described by Das et al. (2009a, b). Intracellular glutathione (GSH) was measured by precipitating the protein content first with 5% TCA. Then DTNB solution (Ellman’s reagent) was added to the non-protein part and the absorbance was measured at 412 nm.
Testicular taurine level detection
Testicular taurine level was measured according to the method of Ferreira et al. (1997). In brief, sulphosalicylic acid solution was added to the homogenate and allowed to stand for 10 min. Then it was filtered through W42 paper and derivatised with o-phthalaldehyde and 2-mercaptoethanol. The derivative was then analysed by HPLC using an UV absorbance detector at 350 nm.
Determination of Na+–K+ and Ca2+ ATPase activities
Na+–K+, Ca2+ ATPases are membrane-bound enzymes that control the transport of Na+, K+, Ca2+ ions across the cell membrane at the expense of ATP hydrolysis. ATPase activity was determined by measuring the amount of liberated phosphate ions by ammonium molybdate, in the presence of known amount of ATP. Ammonium molybdate forms a blue-coloured complex with phosphate ion and the absorbance was measured at 820 nm. Testes were homogenized in a cold buffer containing 320 mM sucrose, 25 mM imidazole (pH 7.4), and 0.1 mM EGTA, using a glass homogenizer. The homogenate was then centrifuged at 1,000g for 15 min. The supernatant was again centrifuged for an additional 30 min at 30,000g. The final pellet was resuspended in the original homogenization buffer and Na+–K+ and Ca2+ ATPases activities were measured by the method of Patil and Balaraman (2009).
Intracellular ATP determination
Cellular ATP levels in testes of all sets of experimental animals were estimated using standard kits (Luciferase/Luciferin Reagent, Promega, USA).
Determination of plasma testosterone level
Plasma testosterone level in all experimental rats was determined by an enzyme immunoassay kit (Assay Designs, Ann Arbor, MI, USA).
Isolation of mitochondria and determination of mitochondrial membrane potential (Δψm)
The testes were minced, washed and homogenized in a Dounce glass homogenizer in 10 mM HEPES–KOH/1 mM EGTA buffer (pH 7.5) containing 250 mM sucrose and supplemented with protease and phosphatase inhibitors. The homogenates were spun down for 10 min at 2,000g at 4°C to discard the debris. The supernatant was overlaid on 0.75 (M) sucrose in HEPES buffer and centrifuged for 30 min at 10,000g. The supernatant was discarded and the mitochondria pellets were resuspended in HEPES buffer and recentrifuged for 10 min at 10,000g. This supernatant was also discarded and the final mitochondrial pellet was resuspended in PBS. It was stored at −80°C until use. Analytic flow cytometric measurements for the membrane potential (Δψm) of isolated mitochondria were performed according to the procedure as described by Ghosh et al. (2010) using a FACScan flow cytometer with an argon laser excitation at 488 nm. Mitochondrial membrane potential (Δψm) was estimated on the basis of cell retention of the fluorescent cationic probe rhodamine 123.
Measurement of intracellular free Ca2+
Testicular cells were isolated by the method of Yeh et al. (2009). Testes were taken from the experimental rats and cut into small pieces, then transferred to a conical flask (100 mL) and incubated in Dulbecco’s Modified Eagle’s Medium (DMEM) medium (containing 0.25% trypsin, 1% glucose, 0.75 mg mL−1 collagenase, proteinase inhibitor cocktail (1:100), 5 mg mL−1 deoxyribonuclease I and 100 IU penicillium-streptomycin) for 1 h at 37°C under stirring condition. The testes sample was then washed with DMEM medium and filtered through 70- and 40-μm nylon filters. The filtrate (containing approximately 2 × 106 testicular cells/mL) was again centrifuged and the pellet was resuspended in PBS.
Intracellular Ca2+ levels were determined using the intracellular Ca2+ probe Fluo 3/acetoxymethyl ester (Molecular Probes, Inc.), which binds Ca2+ with a 1:1 stoichiometry. Cells were incubated in the darkness with 250 nM Fluo-3 for 30 min at 37°C. Fluorescence was measured at FL1 (530 nm) in a BD Biosciences FACScan flow cytometer with excitation at 488 nm, and CellQuest software was employed for subsequent data analysis.
RT-PCR Analysis
RNA was extracted from testes of the rats that were left untreated or treated with DOX alone or treated with taurine using the Trizol method. One microgram of RNA was converted to cDNA using superscript reverse transcriptase. Thermal cycling was performed as follows: 95°C for 105 s (initial denaturation) followed by the set of cycles: 95°C for 15 s (denaturation), primer annealing temperature for 30 s, 72°C for 30 s (primer extension). After 25 cycles the time of DNA extension was 7 min at 72°C. The PCR amplification products were then cooled at 4°C. The PCR-amplified products were then subjected to electrophoresis on 1.5% agarose gels. The product size and annealing temperature of the primers used were given in Table 1.
Immunoblotting
An equal amount of protein (50 μg) from each sample was resolved by 10% SDS-PAGE and transferred to PVDF membrane. Membranes were blocked at room temperature for 2 h in blocking buffer containing 5% non-fat dry milk to prevent non-specific binding and then incubated with anti-p-38 (1:1,000 dilution), anti-p-JNK (1:1,000 dilution), anti-Bad (1:1,000 dilution), anti Bcl-2 (1:1,000 dilution), anti-cytochrome c (1:1,000 dilution), anti-caspase 3 (1:1000 dilution), anti-caspase 8 (1:1,000 dilution), anti-caspase 9 (1:1,000 dilution), anti-p53 (1:1,000 dilution) and anti-Fas (1:1,000 dilution) primary antibodies separately at 4°C overnight. The membranes were washed in TBST (50 mmol/L Tris–HCl, pH 7.6, 150 mmol/L NaCl, 0.1% Tween 20) for 30 min and incubated with appropriate HRP-conjugated secondary antibody (1:2,000 dilution) for 2 h at room temperature and developed by the HRP substrate 3,3′-diaminobenzidine tetrahydrochloride (DAB) system (Bangalore Genei, India).
Histological studies
Testes from the normal and experimental rats were fixed in 10% buffered formalin and were processed for paraffin sectioning. Sections of about 5 μm thickness were stained with haematoxylin and eosin to study the histology of testes of all experimental rats.
DNA fragmentation assay
DNA is negatively charged and moves to cathode during agarose gel electrophoresis. During apoptosis, DNA cleavage occurs at ~200-BP intervals which form a “ladder” pattern at ~200-BP intervals. On the other hand, necrosis is associated with random DNA fragmentation which forms a “smear” on agarose gels. The DNA fragmentation has been assayed by electrophoresing genomic DNA samples, isolated from normal as well as experimental rat testes, on agarose/EtBr gel by the procedure described by Das et al. (2010a).
Statistical analysis
All the values are expressed as mean ± SD (n = 6). Significant differences between the groups were determined with SPSS 10.0 software (SPSS Inc., Chicago, IL, USA) for Windows using one-way analysis of variance (ANOVA) and the group means were compared by Duncan’s multiple range test (DMRT). A difference was considered significant at the P < 0.05 level.
Results
Dose-dependent study of taurine by SDH assay
We used the sorbitol dehydrogenase (SDH) assay to determine the optimum dose necessary for taurine for the protection of rat testes against DOX-induced testicular toxicity, because SDH has been reported to be an important marker enzyme of the testis, and its activity is associated with the development of the germinal epithelium (Mills and Means 1972). Experimental results suggest that DOX intoxication (3 mg/kg body weight, i.p. on alternate days) decreased the SDH activity and that it could be prevented by the taurine treatment up to a dose of 150 mg/kg body weight in distilled water, orally, 1 day before each dose of doxorubicin (Fig. 2). However, higher dose of taurine provided no additional benefit to SDH activity, so the dose of 150 mg/kg body weight was selected for subsequent in vivo experiments.

Dose-dependent effect of taurine on SDH activities. Cont: SDH activity in normal rats; DOX: SDH activity in rats that received doxorubicin (DOX) in three equal doses (3 mg/kg body weight, i.p.) on alternate days; TAU-50, TAU-100, TAU-150, TAU-200 and TAU-250: SDH activities in rats received taurine in three equal doses (50, 100, 150, 200 and 250 mg/kg body weight, respectively, in distilled water, orally) on alternate days followed by DOX exposure. Each column represents mean ± SD, n = 6. a indicates the significant difference between the normal control and DOX-exposed groups and b indicates the significant difference between taurine-treated and DOX-exposed groups (a P < 0.05, b P < 0.05)
Effect of taurine and doxorubicin on body growth, testicular weight, and sperm abnormality
There was no death recorded in any of the experimental groups during the study period. Table 2 shows the effect of DOX and taurine on body and testicular weights. The body weight, testicular weight and the testes weight to body weight ratio were significantly lowered in DOX-exposed rats compared with the controls. Treatment with taurine in DOX-exposed rats restored the body weight, testicular weight and testes weight to body weight ratio to the control levels.
Abnormal sperm count and sperm motility indicate testicular dysfunction. Exposure of male rats to DOX caused a significant decrease in the sperm concentration and motility (%) as compared with those of control animals (Table 2). Treatment with taurine, however, caused an increase in semen quality (involves both sperm quantity and quality is a measure of the ability of semen to accomplish fertilization) and minimized the toxic effects of DOX.
Taurine protected against doxorubicin-induced upregulation of testicular toxicity and oxidative stress
In the present study, we measured several biochemical parameters related to testicular toxicity and oxidative stress in the testes tissue. The decreased activities of SDH and G6PD (Table 3) were notable manifestations of DOX-induced toxicity. However, treatment with taurine maintained the activities of these enzymes almost close to normal.
The MDA (a product of lipid peroxidation) and intracellular ROS levels, activity of antioxidant enzymes and GSH/GSSG ratio reflect the state of oxidative stress within a tissue. DOX-exposed testes contained significantly higher levels of MDA and intracellular ROS (vs. control, Table 3), lower level of GSH as well as GSH/GSSG ratio (vs. control, Table 3) and lower the activities of antioxidant enzymes, namely, SOD, CAT, GST, GR, GPx (vs. control, Table 4), indicating the potent pro-oxidative actions of DOX on testicular tissues. However, taurine treatment neutralized these abnormalities in MDA, GSH and intracellular ROS levels (Table 3). Taurine also normalized the antioxidant enzyme activities (Table 4), showing the protective effect of taurine against DOX-induced oxidative stress in testes.
Effect of doxorubicin on testicular taurine level
The effect of DOX on testicular taurine level has been shown in Table 5. We observed that DOX caused a significant reduction in the taurine level in testicular tissues and that it could be significantly prevented by dietary taurine supplementation.
Effect of taurine on doxorubicin-induced reduction of membrane-bound enzymes
Next we have investigated the activities of two membrane-bound enzymes, Na+–K+ and Ca2+ ATPases. Reduction of these enzymes’ activities clearly reflects the occurrence of membrane damage. In our present study, we observed that DOX exposure significantly decreased the activities of Na+–K+ and Ca2+ ATPases in testes (Fig. 3a). However, taurine treatment restored the activities of these enzymes close to normal.

Effect of taurine on DOX-induced decrease in Na+–K+, Ca2+ ATPases activity (a) and cellular ATP level (b). CON: Na+–K+ and Ca2+ ATPases activity/ATP level in control rats; TAU: Na+–K+ and Ca2+ ATPases activity/ATP level in rats treated with taurine only; DOX: Na+–K+ and Ca2+ ATPases activity/ATP level in DOX-intoxicated rats; TAU + ADOX: Na+–K+ and Ca2+ ATPases activity/ATP level in rats treated with taurine along with DOX. Each column represents mean ± SD, n = 6. a indicates the significant difference between the normal control and DOX-exposed groups and b indicates the significant difference between taurine-treated and DOX-exposed groups (a P < 0.05, b P < 0.05)
Effect of doxorubicin and taurine on cellular ATP level
We have also investigated the effect of DOX on cellular ATP level and found that this level was significantly lower in DOX-exposed animals than in untreated ones (Fig. 3b). However, treatment with taurine could prevent this DOX-induced lowering of ATP content.
Effect of doxorubicin and taurine on the plasma testosterone level
Exposure of DOX significantly decreased the plasma testosterone level in comparison with the control (Fig. 4), suggesting the inhibitory role of DOX on testicular androgenesis. However, treatment with taurine significantly augmented this parameter with respect to the animals treated with only DOX.

Effect of taurine on DOX-induced decrease in plasma testosterone levels. CON: plasma testosterone level; TAU: plasma testosterone level in rats treated with Taurine only; DOX: plasma testosterone level in DOX-intoxicated rats; TAU + ADOX: plasma testosterone level in rats treated with taurine along with DOX. Each column represents mean ± SD, n = 6. a indicates the significant difference between the normal control and DOX-exposed groups and b indicates the significant difference between taurine-treated and DOX-exposed groups (a P < 0.05, b P < 0.05)
Effect on testicular 3β-HSD, 17β-HSD and StAR gene expressions and modulation via taurine
DOX is known to interfere with the testosterone production and decrease the testosterone level (Atessahin et al. 2006). In the present study, we also observed DOX-induced inhibition of testosterone production. To ascertain the possible mechanism of the suppressed testosterone production following DOX administration, the expressions of testicular key androgenic enzymes like 3β-HSD and 17β-HSD along with StAR, a prime regulatory protein for testosterone biosynthesis in testis were studied. The mRNA expressions of 3β-HSD, 17β-HSD and StAR in testicular tissue were diminished significantly following DOX exposure with respect to the control (Fig. 5). However, taurine administration restored the testicular mRNA expressions of all these parameters completely indicating the protective role of taurine on testicular androgenic disorders of DOX exposure.

RT-PCR analysis of 3β-HSD, 17β-HSD and StAR in absence (DOX) and presence of taurine (TAU + DOX) in testes. a Tissue expression of mRNAs for 3β-HSD, b tissue expression of mRNAs for 17β-HSD, c tissue expression of mRNAs for StAR in testes isolated from normal as well as DOX and taurine-treated rats. β actin was taken as an internal control. Data represent the average ± SD of six separate experiments in each group. a indicates the significant difference between the normal control and As-treated groups, b indicates the significant difference between the As-treated and taurine-treated groups (a P < 0.05, b P < 0.05)
Effect of taurine against doxorubicin-induced mitochondria-dependent (intrinsic) cell death pathway
Apoptosis is a well-known cellular action of doxorubicin. Imbalance of Bcl-2 family proteins, loss of mitochondrial membrane potential (Δψm), release of cytochrome c from mitochondria and subsequent activation of caspase-9 and 3 represent a key step in the mitochondrion-dependent apoptotic pathway. To determine whether taurine exerts its antiapoptotic action against doxorubicin via this mechanism, Bad and Bcl-2 protein levels, mitochondrial membrane potential, mitochondrial and cytosolic cytochrome c abundance, cleaved caspase-3 and caspase-9 protein levels were examined. We observed that DOX exposure significantly upregulated the expression of proapoptotic protein (Bad), decreased the abundance of antiapoptotic factor (Bcl-2) and mitochondrial membrane potential (Δψm), increased cytosolic cytochrome c abundance as well as cleavage of caspase-9 and 3 (Fig. 6a–g). However, treatment with taurine could largely inhibit DOX-induced alterations of these parameters.

Immunoblot analysis on Mitochondrion-dependent pathway and Δψm in absence (DOX) and presence of taurine (TAU + DOX) in testes. a Mitochondrial cytochrome c, complex 4/IV was taken as an internal control. b Mitochondrial membrane potential (Δψm) was measured using a fluorescent cationic probe rhodamine-123 by flow cytometer with FL-1 filter. Results represent one of the six independent experiments. c Cytosolic cytochrome c, d Bad, e Bcl-2, f cleaved caspase 9, g cleaved caspase 3, β actin was used as an internal control. Data represent the average ± SD of six separate experiments in each group. a indicates the significant difference between the normal control and DOX-exposed groups, b indicates the significant difference between the DOX-treated and taurine-treated groups (a P < 0.05, b P < 0.05)
Involvement of extrinsic apoptotic pathway
To determine whether the extrinsic apoptotic pathway was also involved in the action of DOX and taurine on testicular cells, crucial factors mediating the extrinsic apoptotic pathway, including death receptor Fas, caspase 8 and Bid were examined. Results show that DOX exposure significantly increased cellular level of Fas (protein), caspase-8 (protein) and t-Bid (protein), suggesting this apoptotic signalling pathway was also involved in the pathogenic action of DOX (Fig. 7a–c). However, taurine treatment modulated these increases in the protein levels of Fas, caspase-8 and Bid.

Investigation of Receptor-mediated pathway in absence (DOX) and presence of taurine (TAU + DOX) in testes. a Bid, b protein level of Fas, c protein level of caspase 8, β actin was used as an internal control. Data represent the average ± SD of six separate experiments in each group. a indicates the significant difference between the normal control and DOX-exposed groups, b indicates the significant difference between the DOX-treated and taurine-treated groups (a P < 0.05, b P < 0.05)
Involvement of sarcoplasmic reticulum-mediated apoptosis
To investigate the involvement of ER/SR-mediated pathway of apoptosis, we analysed the intracellular Ca2+ level as well as the expressions of calpain and caspase-12, because increased intracellular Ca2+ concentrations significantly increase susceptibility to apoptosis via activation of calpain and caspase-12. As depicted in the histogram in Fig. 8a, testicular cells isolated from DOX-exposed rats, produced an increase in FL1 fluorescence, indicating an increased concentration of intracellular Ca2+. DOX exposure also significantly increased the cellular level of calpain 1 (protein) and caspase-12 (protein) (Fig. 8b, c), indicating the involvement of ER/SR-mediated apoptosis. However, taurine treatment could reduce intracellular Ca2+ concentrations and protein levels of calpain and caspase-12.

Investigation of sarcoplasmic reticulum-mediated pathway in absence (DOX) and presence of taurine (TAU + DOX) in testes. a Intracellular Ca2+ levels were monitored using the fluorescent probe Fluo-3/acetoxymethyl by flow cytometer with FL-1 filter. Results represent one of the six independent experiments. b Protein level of calpain 1, c protein level of caspase 12, β actin was used as an internal control. Data represent the average ± SD of six separate experiments in each group. a indicates the significant difference between the normal control and DOX-exposed groups, b indicates the significant difference between the DOX-treated and taurine-treated groups (a P < 0.05, b P < 0.05)
Involvement of other proapoptotic signaling molecules; p53 and MAPKinases, protection via taurine
To further delineate the signalling pathway of DOX-induced apoptosis, cellular p53 and MAPKinases were examined. The expression of p53 (protein) was markedly upregulated in testicular tissues (Fig. 9a). Besides, DOX significantly increased both phospho JNK and phospho p38 protein levels (Fig. 9b, c). However, treatment with taurine could counteract all these DOX-induced alterations.

Investigation of the involvement of p53 and MAPKinase family proteins in response to DOX and taurine (TAU + DOX). a Protein level of p53, b protein levels of phospho and total JNK, c protein levels of phospho and total p38, β actin was used as an internal control. Data represent the average ± SD of six separate experiments in each group. a indicates the significant difference between the normal control and DOX-exposed groups, b indicates the significant difference between the DOX-treated and taurine-treated groups (a P < 0.05, b P < 0.05)
Taurine protected against doxorubicin-induced testicular apoptosis
Finally, we investigated the mode of cell death using DNA gel electrophoresis analysis. A DNA ladder was formed (a hallmark of apoptosis) in the genomic DNA isolated from DOX-exposed animals (Fig. 10). Taurine treatment effectively reduced the DNA laddering of the DOX-exposed animals.

Impact of DOX and taurine on DNA fragmentation pattern on agarose/EtBr gel. DNA isolated from testes was loaded onto 1% (w/v) agarose gels. Lane A Marker (1 kb DNA ladder); lane B DNA isolated from normal testes; lane C DNA isolated from DOX-intoxicated testes; lane D DNA isolated from taurine-treated testes, Arrows indicate ladder formation in DNA isolated from DOX-intoxicated testes
Histological assessment
Figure 11 represents the histological findings of the testes tissue of normal and experimental group of rats. Histological examination revealed atrophy of seminiferous tubules with significant loss of spermatogenic cell layers. Absence of the centrally located spermatozoa was also observed in the DOX-exposed tissues. Treatment with taurine could, however, prevent the DOX-induced toxicity and maintain the normalcy of the testicular architecture.

Haematoxylin and eosin stained testes section of normal rats testes (×100), DOX-intoxicated testes, showing marked degeneration in the seminiferous tubules (×100) taurine-treated testes (×100) showing almost normal morphology
Discussion
The present study was conducted to evaluate whether taurine protects against doxorubicin (DOX)-induced testicular toxicity in rats. Male rats received doxorubicin (DOX) in three equal doses (3 mg/kg body weight, i.p.) on alternate days for a cumulative dose of 9 mg/kg body weight. Results show that doxorubicin retards testicular growth and impairs spermatogenic function mainly through eliciting oxidative stress and cell apoptosis, while treatment with taurine, 150 mg/kg body weight in distilled water, orally, on alternate days for a cumulative dose of 450 mg/kg body weight (each dose was given 1 day before DOX administration) could effectively prevent these toxic effects. Testicular dysfunction is the most common side effect of cytotoxic chemotherapy. DOX is one of the widely used cytotoxic agents for the treatment of various cancers. In the present study, DOX-exposed animals showed a significant decrease in the body weight and testicular weight, sperm count and sperm motility, which are indicative of the impending toxicity. The present findings also show decreased activities of SDH and G6PD in DOX-exposed rats, which are indicative of testicular toxicity. SDH is responsible for providing energy to sperm cells by converting sorbitol to fructose. Decreased activity of this enzyme after DOX exposure suggests the disturbed energy metabolism in sperm cells. G6PD is another key enzyme of the testicular tissue (Prasad et al. 1995) and provides reducing equivalents for the hydroxylation of steroids. G6PD is also directly associated with glutathione metabolism and lowered activity of this enzyme provokes increased oxidative stress to the tissues and may lead to cell death. The oxidative stress in DOX-exposed animals was also evident from the increased level of lipid peroxidation as well as intracellular ROS and decreased level of cellular GSH as well as decreased activity of antioxidant enzymes. DOX also significantly reduced the cellular ATP level and Na+–K+, Ca2+ ATPases’ activity. However, treatment with taurine in DOX-exposed rats could reduce all these DOX-induced alterations and restore the activities of membrane-bound enzymes like Na+–K+ and Ca2+ ATPases due to its membrane-stabilizing effect (Timbrell et al. 1995; Gordon and Heller 1992), thereby proving it to be a cytoprotectant. On the other hand, Tastesen et al. (2010) showed that neither taurine nor GSH could protect EATC and ELA cells against cisplatin-induced apoptosis. Several mechanisms have been proposed for the antioxidant effects of taurine: (a) as a direct antioxidant, taurine could quench and detoxify several reactive free radicals, such as, hypochlorous acid (HOCl) (Timbrell et al. 1995; Huxtable 1992), H2O2 (Cozzi et al. 1995), nitric oxide (NO) (Redmond et al. 1996) and hydroxyl radical (·OH) (Aruoma et al. 1988), (b) as an indirect antioxidant, taurine could prevent the changes in membrane permeability as a result of oxidative injury via intercalating into the membrane and stabilizing it (Timbrell et al. 1995; Gordon and Heller 1992). Besides, taurine also stabilizes GSH-metabolizing enzymes (Hayes et al. 2001) and stimulates glucose-6-phosphate dehydrogenase G6PD which produces NADPH required for the restoration of GSH from GSSG (Tabassum et al. 2006) and thus maintains GSH/GSSG ratio biochemically.
Taurine is accumulated via the Na+-dependent transporter TauT and long-term exposure to high extracellular concentrations of taurine down regulates TauT expression (Poulsen et al. 2002; Lambert 2004), thereby decreasing the cellular concentration of taurine. Therefore, we have checked the taurine concentration in the testicular tissue of the experimental animals. We have observed that testicular taurine level was decreased in DOX-exposed rats, whereas dietary taurine supplementation significantly increased its level in the testicular tissue.
From the previous studies we observed that DOX significantly decreased serum testosterone level (Atessahin et al. 2006). The germ cell degeneration by DOX exposure may be because of the low intratesticular concentrations of testosterone, as high level of testosterone in testis is essential for the normal spermatogenesis as well as for the maintenance of the structural morphology and the normal physiology of seminiferous tubule (Sharpe et al. 1992). In the present study, we also observed decreased plasma testosterone level in DOX-exposed animals and investigated the possible mechanism of the suppressed testosterone production. We observed that the testicular expression of 3β-HSD and 17β-HSD, the key enzymes for testicular androgenesis (Jana et al. 2006), was decreased following DOX administration. StAR transport of cholesterol across the mitochondrial membrane is generally considered the rate-limiting step in steroidogenesis (Stocco 2001), and expression of StAR mRNA was significantly reduced following DOX exposure. However, taurine treatment significantly reversed all these adverse effects associated with the testicular changes induced by DOX.
Apoptosis is a delicately controlled process of programmed cell death that could be completed through either mitochondrion-dependent or -independent pathway. Doxorubicin is a well-known pro-apoptotic agent. In the present study, our results also demonstrate that DOX induces apoptosis in testicular cells as evident from DNA gel electrophoresis where we obtained a ladder pattern of genomic DNA isolated from DOX-intoxicated testes. Treatment with taurine, however, prevents the DNA ladder formation (apoptosis). Yeh et al. (2009) showed that within a short period of exposure (7 days or at an early stage of testicular toxicity) of DOX at the same dose (Jahnukainen et al. 2001), both intrinsic and extrinsic (without the involvement of FAS) pathways were involved in the pathogenic action of DOX. In the present study, we used the same dose of DOX (Jahnukainen et al. 2001) and investigated the involvement of both intrinsic and extrinsic pathways after 3 weeks from exposure to the last dose of DOX. From immunoblot analyses we observed up-regulation of Bad, cytosolic cytochrome c as well as cleaved caspase-9 and down-regulation of Bcl-2, indicating the involvement of intrinsic pathway. DOX exposure also significantly increased cellular level of Fas (protein), caspase-8 (protein) and Bid, suggesting the involvement of extrinsic pathway. However, the involvement of ER/SR-mediated pathway of apoptosis is still not known in DOX-induced testicular pathophysiology. Therefore, in the present study we investigated the role of ER/SR-mediated pathway. We observed that DOX significantly increased intracellular Ca2+ concentrations as well as activated calpain 1 (protein level) and caspase-12 (protein level). Caspase-12 is an endoplasmic reticulum stress-associated caspase and activation of calpain is required for cleavage of procaspase-12 to generate active caspase-12. Thus, DOX can activate mitochondrial-mediated, receptor-mediated and endoplasmic/sarcoplasmic reticulum-mediated apoptotic signal transduction pathways in testicular tissue under the present pathophysiological condition. However, treatment with taurine could antagonize all these DOX-mediated pro-apoptotic pathways.
The present study is the first to investigate the MAPKinase signalling pathways in DOX-induced testicular pathophysiology. Mitogen-activated protein kinase (MAPK) signalling pathways are the primary intermediator of induction of apoptosis by oxidative stress. Among the MAPKinase family, JNKs and p38-MAPKs are activated by cellular stresses, including oxidative stress and are thought to correlate with cell apoptosis (Kyriakis and Avruch 1996). In the present study, phosphorylation of JNKs and p38-MAPKs were elevated in the testicular tissue exposed to DOX. Another important signalling molecule is p53, which is known to be involved in DOX-induced testicular mitochondriopathy (Yeh et al. 2007). We also observed that protein level of p53 was increased in the testicular tissue exposed to DOX. However, treatment with taurine, a known antioxidant, modulated the activation of the MAPKs and p53, indicating the activation of MAPKs and p53 through oxidative stress.
The testicular toxicity induced by DOX is further confirmed by the abnormal histologic findings in DOX-exposed animals. DOX-induced testicular tissue shows seminiferous tubules with destruction and disruption in spermatogenesis. In our study, taurine-treated animals showed minimal histologic abnormalities, thereby highlighting its protective role in counteracting the cytotoxic injury induced by DOX.
In conclusion, the present study suggests for the first time that taurine is a potential candidate for combating the testicular abnormalities and apoptosis induced by DOX. DOX can stimulate testicular apoptosis via the activation of MAPKs and p53. p53 responds to both DNA damage and oxidative stress, whereas MAPKs respond to mainly oxidative stress to trigger downstream apoptotic signalling. Therefore, the mechanism for DOX-induced apoptosis seems to originate both from the pharmacological consequences of DNA injury and oxidative stress. However, taurine effectively attenuated these oxidative and apoptotic actions of DOX in testes due to its potent antioxidant as well as membrane-stabilizing effect (as indicated in the proposed scheme of Fig. 12). The results further emphasize the potential utilization of taurine in protecting toxic side effects of doxorubicin in clinical chemotherapy.

Schematic diagram of the DOX-induced testicular toxicity and its prevention by taurine
Abbreviations
- CAT:
-
Catalase
- DOX:
-
Doxorubicin
- GSH:
-
Glutathione
- GSSG:
-
Glutathione disulfide
- GST:
-
Glutathione S-transferase
- GPx:
-
Glutathione peroxidase
- G6PD:
-
Glutathione 6 peroxidase
- GR:
-
Glutathione reductase
- 3β-HSD:
-
3β-Hydroxysteroid dehydrogenase
- 17β-HSD:
-
17β-Hydroxysteroid dehydrogenase
- MDA:
-
Malonaldehyde
- MAPKs:
-
Mitogen-activated protein kinases
- ROS:
-
Reactive oxygen species
- SDH:
-
Sorbitol dehydrogenase
- SOD:
-
Superoxide dismutase
- StAR protein:
-
Steroidogenic acute regulatory protein
- TAU:
-
Taurine
References
Alvarez JG, Storey BT (1983) Taurine, hypotaurine, epinephrine and albumin inhibit lipid peroxidation in rabbit spermatozoa and protect against loss of motility. Biol Reprod 29:548–555
Arola OJ, Saraste A, Pulkki K, Kallajoki M, Parvinen M, Voipio-Pulkki LM (2000) Acute doxorubicin cardiotoxicity involves cardiomyocyte apoptosis. Cancer Res 60:1789–1792
Aruoma OI, Halliwell B, Hoey BM, Butler J (1988) The antioxidant action of taurine, hypotaurine and their metabolic precursors. Biochem J 256:251–255
Atessahin A, Turk G, Karahan I, Yolmaz S, Ceribasi AO, Bulmus O (2006) Lycopene prevents adriamycin-induced testicular toxicity in rats. Fertil Steril 85:1216–1222
Au WW, Hsu TC (1980) The genotoxic effects of adriamycin in somatic and germinal cells of the mouse. Mutat Res 79:351–361
Beere HM, Wold BB, Caines K, Mosser DD, Mahboubi A, Kuwana T, Tailor P, Morimoto RI, Cohen GM, Green DR (2000) Heat-shock protein 70 inhibits apoptosis by preventing recruitment of procaspase-9 to the Apaf-1 apoptosome. Nat Cell Biol 2:469–475
Bergmeyer HU (1974) Methods of enzymatic analysis. Academic Press, New York, pp 744–761
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Cozzi R, Ricordy R, Bartolini F et al (1995) Taurine and ellagic acid: two differently-acting natural antioxidants. Environ Mol Mutagen 26:248–254
Das J, Ghosh J, Manna P, Sil PC (2008) Taurine provides antioxidant defense against NaF-induced cytotoxicity in murine hepatocytes. Pathophysiology 15:181–190
Das J, Ghosh J, Manna P, Sinha M, Sil PC (2009a) Taurine protects rat testes against NaAsO(2)-induced oxidative stress and apoptosis via mitochondrial dependent and independent pathways. Toxicol Lett 187:201–210
Das J, Ghosh J, Manna P, Sinha M, Sil PC (2009b) Arsenic-induced oxidative cerebral disorders: protection by taurine. Drug Chem Toxicol 32:93–102
Das J, Ghosh J, Manna P, Sil PC (2010a) Protective role of taurine against arsenic-induced mitochondria-dependent hepatic apoptosis via the inhibition of PKCd-JNK pathway. PLoS ONE 5:e12602
Das J, Ghosh J, Manna P, Sil PC (2010b) Acetaminophen induced acute liver failure via oxidative stress and JNK activation: protective role of taurine by the suppression of cytochrome P450 2E1. Free Radic Res 44:340–355
Das J, Ghosh J, Manna P, Sil PC (2010c) Taurine protects acetaminophen-induced oxidative damage in mice kidney through APAP urinary excretion and CYP2E1 inactivation. Toxicology 269:24–34
Das J, Ghosh J, Manna P, Sil PC (2011) Taurine suppresses doxorubicin-triggered oxidative stress and cardiac apoptosis in rat via up-regulation of PI3-K/Akt and inhibition of p53, p38-JNK. Biochem Pharmacol (in press). doi:10.1016/j.bcp.2011.01.008
Ferreira MPLVO, Nunes MV, Mendes E, Remiilo F, Ferreirat IMA (1997) Development of an HPLC-UV method for determination of taurine in infant formulae and breast milk. J Liq Chrom Rel Techno 20:1269–1278
Fraser LR (1986) Both taurine and albumin support mouse sperm motility and fertilizing ability in vitro but there is no obligatory requirement for taurine. J Reprod Fertil 77:271–280
Ghosh J, Das J, Manna P, Sil PC (2009) Taurine prevents arsenic-induced cardiac oxidative stress and apoptotic damage: role of NF-kappaB, p38 and JNK MAPK pathway. Toxicol Appl Pharmacol 240:73–87
Ghosh J, Das J, Manna P, Sil PC (2010) Hepatotoxicity of di-(2-ethylhexyl) phthalate is attributed to calcium aggravation, ROS-mediated mitochondrial depolarization, and ERK/NF-κB pathway activation. Free Radic Biol Med 49:1779–1791
Goodman J, Hochstein P (1977) Generation of free radicals and lipid peroxidation by redox cycling of adriamycin and daunomycin. Biochem Biophys Res Commun 77:797–803
Gordon RE, Heller RF (1992) Taurine protection of lungs in hamster models of oxidant injury: a morphologic time study of paraquat and bleomycin treatment. In: Lombardini JB, Schaffer SW, Azuma J (eds) Taurine: nutritional value and mechanisms of action. Plenum Press, New York, pp 319–323
Green DR (2000) Apoptotic pathways: paper wraps stone blunts scissors. Cell 102:1–4
Hayes J, Tipton KF, Bianchi L, Corte LD (2001) Complexities in the neurotoxic actions of 6-hydroxydopamine in relation to the cytoprotective properties of Taurine. Brain Res Bull 55:239–245
Herman EH, Ferrans VJ (1998) Animal models of anthracycline cardiotoxicity: basic mechanisms and cardioprotective activity. Prog Pediatr Cardiol 8:49–58
Holloway C, Kotsanas G, Wendt I (1999) Acute effects of taurine on intracellular calcium in normal and diabetic cardiac myocytes. Pflugers Arch 438:384–391
Huxtable RJ (1989) Taurine in the central nervous system and the mammalian actions of taurine. Prog Neurobiol 32:471–533
Huxtable RJ (1992) Physiological actions of taurine. Physiol Rev 72:101–163
Jahnukainen K, Jahnukainen T, Salmi TT, Svechnikov K, Eksborg S, Soder O (2001) Amifostine protects against early but not late toxic effects of doxorubicin in infant rats. Cancer Res 61:6423–6427
Jana K, Jana S, Samanta PK (2006) Effects of chronic exposure to sodium arsenite on hypothalamo-pituitary-testicular activities in adult rats: possible an estrogenic mode of action. Reprod Biol Endocrinol 4:9–22
Kyriakis JM, Avruch J (1996) Sounding the alarm: protein kinase cascades activated by stress and inflammation. J Biol Chem 271:24313–24316
Lambert IH (2004) Regulation of the cellular content of the organic osmolyte taurine, in mammalian cells. Neurochem Res 29:27–63
Lamkanfi M, Kalai M, Vandenabeele P (2004) Caspase-12: an overview. Cell Death Differ 11:365–368
Llanos MN, Ronco AM (1994) Sperm phospholipid methyltransferase activity during preparation for exocytosis. Cell Biochem Funct 12:289–296
Lui RC, Laregina MC, Herbold DR, Johnson FE (1986) Testicular cytotoxicity of intravenous doxorubicin in rats. J Urol 136:940–943
Manna P, Sinha M, Sil PC (2008a) Amelioration of cadmium-induced cardiac impairment by taurine. Chem Biol Interact 174:88–97
Manna P, Sinha M, Sil PC (2008b) Taurine triggers a chemoprevention against cadmium induced testicular oxidative injury. Reprod Toxicol 26:282–291
Manna P, Sinha M, Sil PC (2009) Taurine plays a beneficial role against cadmium-induced oxidative renal dysfunction. Amino Acids 36:417–428
Meizel S, Lui CW, Working PK, Mrsny RJ (1980) Taurine and hypotaurine: their effects on motility, capacitation and the acrosome reaction of hamster sperm in vitro and their presence in sperm and reproductive tract fluids of several mammals. Dev Growth Differ 22:483–494
Mills NC, Means AR (1972) Sorbitol dehydrogenase of rat testis, changes of activity during development after hypophysectomy and following gonadotrophic hormone administration. Endocrinology 91:47–56
Mrsny RJ, Meizel S (1985) Inhibition of hamster sperm Na+, K+-ATPase activity by taurine and hypotaurine. Life Sci 36:271–275
Muller I, Niethammer D, Bruchelt G (1998) Anthracycline-derived chemotherapeutics in apoptosis and free radical cytotoxicity (review). Int J Mol Med 1:491–494
Nakagawa T, Zhu H, Morishima N, Li E, Xu J, Yankner BA, Yuan J (2000) Caspase-12 mediates endoplasmic-reticulum-specific apoptosis and cytotoxicity by amyloid. Nature 403:98–103
Patil L, Balaraman R (2009) Effect of melatonin on doxorubicin induced testicular damage in rats. Int J Pharm Tech Res 1:879–884
Pollack M, Leeuwenburgh C (2001) Apoptosis and aging: role of the mitochondria. J Gerontol A Biol Sci Med Sci 56:475–482
Poulsen KA, Litman T, Eriksen J, Mollerup J, Lambert IH (2002) Downregulation of taurine uptake in multidrug resistant Ehrlich ascites tumor cells. Amino Acids 22:333–350
Prasad AK, Pant N, Srivastava SC, Kumar R, Srivastava SP (1995) Effect of dermal application of hexachlorocyclohexane (HCH) on male reproductive system of rat. Hum Exp Toxicol 14:484–488
Quinn R (2005) Comparing rat’s to human’s age: how old is my rat in people years? Nutrition 21:775–777
Redmond HP, Wang JH, Bouchier-Hayes D (1996) Taurine attenuates nitric oxide- and reactive oxygen intermediate-dependent hepatocyte injury. Arch Surg 131:1287–1288
Roy A, Manna P, Sil PC (2009) Prophylactic role of taurine on arsenic mediated oxidative renal dysfunction via MAPKs/NF-kappaB and mitochondria dependent pathways. Free Radic Res 43:995–1007
Sharpe RM, Maddocks S, Millar M, Saunders PTK, Kerr JB, McKinnell C (1992) Testosterone and spermatogenesis: identification of stage dependent, androgen-regulated proteins secreted by adult rat seminiferous tubules. J Androl 13:172–184
Shinoda K, Mitsumori K, Yasuhara K, Uneyama C, Onodera H, Hirose M, Uehara M (1999) Doxorubicin induces male germ cell apoptosis in rats. Arch Toxicol 73:274–281
Sikka SC (1996) Oxidative stress and role of antioxidants in normal and abnormal sperm function. Front Biosci 1:e78–e86
Sinha M, Manna P, Sil PC (2007) Taurine, a conditionally essential amino acid, ameliorates arsenic-induced cytotoxicity in murine hepatocytes. Toxicol In Vitro 21:1419–1428
Sinha M, Manna P, Sil PC (2008a) Taurine protects antioxidant defense system in the erythrocytes of cadmium treated mice. BMB Rep 41:657–663
Sinha M, Manna P, Sil PC (2008b) Cadmium induced neurological disorders: prophylactic role of taurine. J Appl Toxicol 28:974–986
Sinha M, Manna P, Sil PC (2009) Induction of necrosis in cadmium-induced hepatic oxidative stress and its prevention by the prophylactic properties of taurine. J Trace Elem Med Biol 23:300–313
Stocco DM (2001) StAR protein and the regulation of steroid hormone biosynthesis. Annu Rev Physiol 63:193–213
Sun XM, MacFarlane M, Zhuang J, Wolf BB, Green DR, Cohen GM (1999) Distinct caspase cascades are initiated in receptor-mediated and chemical-induced apoptosis. J Biol Chem 274:5053–5060
Tabassum H, Rehman H, Banerjee BD, Raisuddin S, Parvez S (2006) Attenuation of tamoxifen-induced hepatotoxicity by taurine in mice. Clin Chim Acta 370:129–136
Tastesen HS, Holm JB, Møller J, Poulsen KA, Møller C, Stürup S, Hoffmann EK, Lambert IH (2010) Pinpointing differences in cisplatin-induced apoptosis in adherent and non-adherent cancer cells. Cell Physiol Biochem 26:809–820
Timbrell JA, Seabra V, Watereld CJ (1995) The in vivo and in vitro protective properties of taurine. Gen Pharmacol 26:453–462
Van Der Horst CJG, Grooten HJG (1966) The occurrence of hypotaurine and other sulfur containing amino acids in seminal plasma and spermatozoa of boar, bull and dog. Biochim Biophys Acta 117:495–497
Wright CE, Tallan HH, Lin YY (1986) Taurine: biological update. Annu Rev Biochem 55:427–453
Wyrobek AJ, Bruce WR (1975) Chemical induction of sperm abnormalities in mice. Proc Natl Acad Sci USA 72:4425–4429
Yeh YC, Lai HC, Ting CT, Lee WL, Wang LC, Wang KY, Lai HC, Liu TJ (2007) Protection by doxycycline against doxorubicin-induced oxidative stress and apoptosis in mouse testes. Biochem Pharmacol 74:969–980
Yeh YC, Liu TJ, Wang LC, Lee HW, Ting CT, Lee WL, Hung CJ, Wang KY, Lai HC, Lai HC (2009) A standardized extract of Ginkgo biloba suppresses doxorubicin-induced oxidative stress and p53-mediated mitochondrial apoptosis in rat testes. Br J Pharmacol 156:48–61
Acknowledgments
The authors are grateful to Mr. Prasanta Pal for excellent technical assistance for the study.
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Das, J., Ghosh, J., Manna, P. et al. Taurine protects rat testes against doxorubicin-induced oxidative stress as well as p53, Fas and caspase 12-mediated apoptosis. Amino Acids 42, 1839–1855 (2012). https://doi.org/10.1007/s00726-011-0904-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00726-011-0904-4