
Abstract
The present study provides data on the insertion of an extra copy of phytochelatin synthase (alr0975) in Anabaena sp. PCC 7120. The recombinant strain (AnFPN-pcs) compared to wild type showed approximately 22.3% increase in growth rate under UV-B, NaCl, heat, CuCl2, carbofuran, and CdCl2. It also registered 2.25-fold enhanced nitrogenase activity and 5-fold higher phytochelatin production. A comparison of the protein profile of wild type with the recombinant strain revealed that recombinant strain accumulated proteins belonging to the following categories: (i) detoxification (nutrient stress induced DNA binding protein, Mn-SOD, Alr0946 (CalA)), (ii) protein folding and modification (molecular chaperone DnaK, FKBP-type peptidyl-prolyl cis-trans isomerase), (iii) nucleotide and amino acid biosynthesis (dihydroorotase and Ketol-acid reductoisomerase), (iv) photosynthesis and respiration (coproporphyrinogen III oxidase, phycocyanin alpha chain, ferredoxin-NADP+ reductase), and (v) transport (sugar transport ATP-binding protein). Thus, it can be concluded that, above category proteins with their respective role in scavenging reactive oxygen species, proper folding of unfolded proteins, and protection of protein from degradation, sustained carbon fixation and energy pool and active transport of sugar together conceivably help the recombinant cyanobacterium (AnFPN-pcs) to cope with abiotic stress employed in the present study. Such recombinant strains have potential for future use as biofertilizer.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Cyanobacteria, an extremely diverse group of photosynthetic prokaryotes, were the sole photosynthesizers for eons and contributed oxygen to the primitive reducing and anaerobic atmosphere (Seckbach and Oren, 2007). Diazotrophic cyanobacteria endowed with the exclusive ability of photosynthesis and nitrogen fixation are an integral component of paddy fields. However, they are inevitably exposed to various stresses such as salinity, drought, heat, heavy metals, and pesticides and display enormous plasticity to adapt under aforesaid stresses (Pandey et al., 2012; Rai et al., 2014; Agrawal et al., 2014; Shrivastava et al., 2015; Singh et al., 2015). Moreover, existence of cyanobacteria under extreme environments (harsh conditions of temperature, pH, harmful radiation, and hypersalinity) makes them an excellent source for mining stress-tolerant genes.
Various reports witnessed the above statement and stress tolerant transgenics in cyanobacteria as well as in plants have been raised utilizing cyanobacterial genes. For example, overexpression of groESL operon (Chaurasia and Apte, 2009), AhpC (Shrivastava et al., 2016), and dps (Narayan et al., 2016) in Anabaena PCC7120 offered tolerance to multiple abiotic stresses. Moreover, the expression of phytoene desaturase (Pds) from Synechococcus sp. PCC7942 and isiB (flavodoxin) from Anabaena sp. in tobacco offered tolerance to various abiotic stresses (Wagner et al., 2002; Tognetti et al. 2006, 2007).
Phytochelatins (PCs) are small cysteine-rich heavy metal-binding peptides, functionally analogous to metallothioneins (Grill et al., 1987, 1989) and display up-accumulation in response to heavy metal stress (Mallick et al., 1994; Tsuji et al., 2004). Metal tolerance through increase in PCs can be accredited to the presence of a thiol group which coordinates with metals (Hall, 2002; Cobbett and Goldsbrough, 2002). However, their role is not constrained to metal-associated stress; various reports suggest their significant role under a variety of stresses such as salinity, heat, UV-B, and drought (Zhang et al. 2005; Bhargava et al. 2005; Seki et al. 2003). PC-mediated tolerance to various abiotic stresses other than heavy metals can be attributed to shielding of thiols through conformational changes of PCs, thus protecting thiols from the damaging effects of stress as thiols are well known for their central role in abiotic stress tolerance (Bhargava et al., 2005).
Extensive work has been done on PCs from organisms other than cyanobacteria. In green alga Stigeoclonium, PCs displayed abundance following heavy metal stress (Pawlik-Skowrońska 2001). Similar results were found in Arabidopsis thaliana (Vatamaniuk et al. 1999), wheat (Ha et al. 1999), marine and fresh water algae (Gekeler et al. 1989; Pawlik-Skowrońska et al. 2007), fungi (Mehra et al. 1988; Kneer et al. 1992; Bolchi et al., 2011), and lichens (Pawlik-Skowrońska et al. 2002). However, the functional importance of cyanobacterial PCs under abiotic stresses has remained confined to only few reports (Bhargava et al., 2005; Chaurasia et al., 2008a, b). Heterologous expression of phytochelatin encoding gene (pcs) of Anabaena PCC7120 in Escherichia coli alleviated damaging effects of high temperature, salinity, carbofuran, cadmium, copper, and UV-B (Chaurasia et al., 2008a, b). In view of the above, to determine the role of PCs in abiotic stresses, an extra copy of phytochelatin-encoding gene phytochelatin synthase (pcs) was constitutively expressed in Anabaena PCC7120. An integrative expression vector pFPN, used for incorporating a cassette of genes for genomic integration and expression into Anabaena sp. PCC 7120 (Chaurasia et al., 2008a, b; Chaurasia and Apte 2009), was employed. Moreover, to address the question as how far the overexpression of pcs (alr0975) affects the performance of other metabolic proteins which might collectively help in the stress tolerance, comparative proteomic studies of the wild-type (Anabaena sp. PCC 7120) and the transformed Anabaena (hereafter AnFPN-pcs) by using 2-DE and MALDI-TOF/MS has also been carried out.
Materials and methods
Conserved domain analysis
The amino acid sequence of Alr0975 was retrieved from cyanobase (http://genome.microbedb.jp/cyanobase/Anabaena). Conserved domain analysis was performed with conserved domain database of NCBI (Marchler-Bauer et al. 2015).
Organism and growth conditions
Anabaena sp. PCC 7120 was grown photoautotrophically in BG-11 medium (Rippka et al. 1979) without nitrogen, buffered with Tris/HCl at 24 ± 2 °C under day light fluorescent tubes emitting 72 μmol photon m−2 s−1 photosynthetically active radiation (PAR) light intensity with a photoperiod of 14:10 h at pH 7.5. The cultures were shaken by hand two to three times daily. E. coli strain DH5α was used as a host for cloning. E. coli cultures were stored as 10% (v/v) glycerol stocks at −80 °C and maintained on Luria-Bertani (LB) plates at 37 °C containing 1.4% (w/v) agar. Cells harboring recombinant plasmids were grown and maintained on LB medium supplemented with neomycin (Sambrook and Russell, 2001).
DNA isolation, PCR amplification, and cloning of alr0975 gene into the pFPN vector
Genomic DNA from An7120 was isolated as described by Srivastava et al. 2007. An open-reading frame alr0975, putatively encoding phytochelatin synthase (pcs) (Fig. S1), was amplified by polymerase chain reaction using genomic DNA as the template with a pair of primers: Pf (5′-GAATTCCATATGGAATTCATGGCTCTCCGTCTTGGT-3′) and Pr (5′-′CGGGATCCCGTTACTTGTTAGGTTGAG GAGT-3′). The underlined bases are NdeI and BamHI recognition sites, respectively. The PCR was done in a reaction mixture of 25 μl for 30 cycles at 94 °C for 1.5 min, 56 °C for 1 min, and 72 °C for 2 min using standard PCR conditions (100 ng of DNA, 2.5 μl of 10× PCR buffer with 15 mM MgCl2, 200 μM dNTPs, 10 pmol of each primer, and 0.2 U Taq DNA polymerase in an Icycler (Bio-Rad, USA)). The amplified product was purified using a Qiagen gel extraction kit.
Generation of recombinant DNA construct for overexpression of alr0975
The purified PCR product was digested with NdeI and BamHI(NEB), and the resultant DNA fragment was cloned into an integrative expression vector pFPN that had been digested with the same restriction enzymes. To construct the recombinant plasmid, pFPN-alr0975 was introduced into E. coli DH5α and transformed colonies were grown in LB medium. The plasmid was then isolated and the sequence of alr0975 was confirmed by sequencing and double digestion of recombinant plasmid to confirm that no mutations had occurred during PCR amplification.
Electroporation of recombinant plasmid into Anabaena sp. PCC 7120
The pcs gene alr0975 amplicon was cloned into plasmid vector pFPN, which allows integration at a defined undisruptive site in the Anabaena sp. PCC 7120 genome and expression from a strong cyanobacterial PpsbA1 promoter. The recombinant plasmids, pFPN-alr0975, were individually introduced into Anabaena by electroporation as demonstrated by Chaurasia et al., 2008a, b. The electrotransformants were selected on BG11, N+ neomycin (25 μg ml−1) plates by repeated subculturing for at least 24 weeks to achieve complete segregation and were designated as AnFPN-pcs.
Expression analysis using semiquantitative RT-PCR, western blotting, and enzymatic assay
Anabaena strain harboring the pFPN-pcs plasmid was grown in BG-11 medium supplemented with 100 μg ml−1neomycin in an orbital shaker (200 rpm) at 37 °C. Mid-exponential phase of this culture was used to isolate RNA using TRIzol reagent as per the manufacturer’s protocol (Invitrogen, USA). Reverse transcriptase (RT)-PCR was performed as per Mishra et al., 2009 using a Bio-Rad cDNA synthesis kit (Bio-Rad, Singapore), and gene-specific primers sets were designed using primer3 software. For RT-PCR of alr0975, forward primer 5′TAATCGCGCCCGAAGTAGTA3′ and reverse primer 5′TCTTGCCCAATTTCCTTACG3′ were used while for 16SrDNA (as an internal control), Pf 5′CACACTGGGACTGAGACAC3′ and Pr 5′CTGCTGGCACGGAGTTAG3′ were used. First-strand cDNA synthesis was performed using 1 μg of DNase I treated RNA in a 20-μl reaction volume according to the manufacturer’s protocol.
Western blot analysis was done as described previously (Chaurasia et al., 2008a, b). Briefly, samples containing 30 μg of protein both from wild-type and overexpression strain were mixed in 1:1 (v/v) ratio with the gel loading buffer and run at a constant current of 15 mA until the samples entered the resolving gels followed by increasing the current to 25 mA for 4–5 h SDS-PAGE. The gels were transferred to a PVDF membrane (Millipore Immobilon-P), using a dual mini-electroblot system (Precision Instruments, Varanasi, India). The gel cassette was kept in transfer buffer (3.03 g/l Tris base, 14.4 g/l glycine, and 200 ml/l methanol (99% v/v pure) for 12 h at 50 V and 4°C. The membrane was blocked for 4 h in Tris buffer saline containing 0.1% Tween 20 (TTBS) and 5% (w/v) non-fat dried milk. The primary antibody was diluted (1: 2000 times) as per the instructions of the donors. The membrane incubated overnight at 4 °C in the diluted solution of the primary antibody was washed five times for 5 min each in TTBS. This was then incubated in a goat anti-rabbit IgG horseradish peroxidase (HRP)-conjugated secondary antibody (1:5000) (Genei, India) for 4 h. Following four consecutive 5-min washes in TTBS, the membrane was developed with DAB/NiCl2 visualization solution. The reaction was terminated by washing the PVDF membrane with deionized distilled water. The blots were dried between filter papers, which considerably reduced the background staining. Polyclonal antibodies against phytochelatin synthase for detection by immunoblotting were obtained as generous gift from Dr. Stephan Cuine (CEA Cadarache, France). SDS-PAGE was performed with the same concentration of protein used in western blotting both for wild-type and recombinant strain which was taken as internal control.
The activity of gene in terms of enzyme phytochelatin synthase was determined by the level of phytochelatin inside the cells as per the methods of Bhargava et al. (2005). Protein concentration was estimated following the method of Bradford (1976).
Growth behavior analysis under different abiotic stresses
In order to investigate the role of alr0957 in abiotic stress tolerance, a comparative growth behavior of An7120 and AnFPN-pcs was performed under different abiotic stresses. Both wild-type and overexpressed strains at their exponential phase were treated with LC50 dozes of Cd2+ (10 μM) (Singh et al. 2015), Cu2+ (20 μM) (Bhargava et al., 2008), temperature (42 °C) (Rajaram and Apte 2008), UV-B (g 2.34 × 106 μE·cm − 2) (Shrivastava et al. 2015), NaCl (100 mM) (Rai et al. 2014), and carbofuran (452 μM) (Padhy and Mohapatra 2001). Growth was estimated by measuring the optical density of the cyanobacterial cultures at 663 nm in a UV-Vis spectrophotometer (CECIL, England) on every 2nd day up to the 11th day by using a reference blank of basal culture medium. Specific growth rate was calculated by using the following equation: m = [ln (n 2/n 1)]/[t 2 − t 1], where m stands for specific growth rate and n 1 and n 2 are absorbance of culture suspension at the beginning (t 1) and the end (t 2) of the selected time interval.
Estimation of nitrogenase activity
The nitrogenase activity of Anabaena was determined by acetylene reduction assay (Stewart et al. 1968). For acetylene reduction, both the cultures of An7120 and AnFPN-pcs in the exponential phase were transferred to 13-mL glass vials and sealed with Suba seals. The sealed vials were evacuated and flushed with argon twice and then sparged with argon for10–15 min. For the nitrogenase assay, acetylene was injected at 10% concentration by volume and incubated for 2 h under continuous irradiance of 72 μmol photon m−2 s−1 PAR. The reaction was terminated by injecting 0.8 mL of 15% TCA. The ethylene formed was measured in a Varian (Palo Alto, CA, USA) CP-3800 gas chromatograph equipped with a Porapak R column (Varian) and a flame ionization detector. Activity was expressed in terms of nmol C2H2 formed mg−1 protein h−1.
Total protein extraction and two-dimensional gel electrophoresis
Protein isolation and two-dimensional gel electrophoresis were done according to a modified protocol of Wagner et al. (2002). Cultures of An7120 and AnFPN-pcs in the exponential phase were harvested by centrifugation (10,000×g, 10 min.), washed with Tris buffer (pH 8.0), and suspended in 2-ml extraction buffer containing 10 mM Tris–HCl (pH 8.0), 1.5 mM MgCl2, and 10 mM KCl. Cell pellet was ground using liquid nitrogen followed by centrifugation at 9200 rpm for 1 h. Furthermore, the supernatant was precipitated with 10% TCA in acetone, left overnight for complete precipitation, and centrifuged at 6,000×g for 15 min to recover protein pellet. Protein pellet was washed three to four times with ice-cold acetone to remove traces of TCA. Then, the pellet was air dried and dissolved in solubilization buffer containing 7 M urea, 2 M thiourea, 4% CHAPS, 40 mM DTT, and 1.0% IPG buffer (4–7). Protein content was estimated according to Bradford using BSA as standard. Before loading for 2DE, traces of bromophenol blue was added and centrifuged at 19,000×g for 10 min. Sample entry was made through in-gel rehydration. A total of 300 μl of solubilization buffer containing 250 μg protein sample was incubated with the dry immobilized pH gradient (IPG) gel strips (pH 4–7) linear gradients (13 cm; GE healthcare) at 200 °C for 16 h. The first dimension separation was conducted at 20 °C with an EttanIPGphor system (GE Healthcare BioSciences). Focusing was performed in seven steps: linear 30 V for 00.30 h, 150 for 2:00 h, 300 V for 00:40 h, 500 V for 4 h, gradient 1000 V for 1 h, gradient 8000 V for 2 h, and finally 8000 V for 13:00 h. Focused IPG strips were then equilibrated by first incubating them in an equilibration solution (6 M urea, 30% v/v glycerol, 2% w/v SDS, 50 mM Tris–HCl, pH 8.8, and 1% w/v DTT and a trace amount of bromophenol blue) for 15 min, followed by incubation in 2.5% w/v iodoacetamide in the same equilibration solution instead of 1% DTT for 15 min. The strip was placed on the top of 12% SDS-polyacrylamide gels and electrophoresis was performed at 15 mA for 30 min followed by 25 mA for 5 h using a Hoefer SE 600 apparatus (Amersham Biosciences). The gel was stained with Coomassie Brilliant Blue R-250.
Data analysis
PDQuest™ version 7.1 (Bio-Rad) was used for 2D gel image analysis. Protein spot detection, quantification, background subtraction, and spotmatching between multiple gels were performed. K-nearest neighbor (KNN) algorithm [MATLAB] was used for performing missing value imputation. Normalization of spot intensities (% of individual spot intensity/Σ% spot intensity of each gel) was done with total spot intensity per gel in order to reimburse variations between the replicates. Protein spots with p values less than 0.05 were noted as statistically significant, and protein spots greater than minimum abundance ratio of 1.5-fold were selected and subjected to MALDI-TOF LCMS analysis followed by homology search using MASCOT.
In addition, a heat map visualizing proteomics responses of the An7120 and AnFPN-pcs was prepared with TIGR Multiple Experiment Viewer (MEV) 4.2 software (Saeed et al. 2003).
Results
The extra copy of alr0975 in An7120 induced phytochelatin synthesis, enhanced nitrogen fixation, and conferred better growth against the wild type under different abiotic stresses
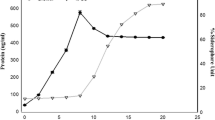
Transfer of pFPN-pcs to An7120 integrated an extra copy of pcs (PpsbA1:pcs) was confirmed and selected on BG-11, N+, Neo25 agar plates which restricted growth of wild type (Fig. 1a). The presence of an extra copy of pcs gene in An7120 was further confirmed by RT-PCR and western blotting approaches. Differential expression of RNA isolated from overexpressed and wild-type strains using RT-PCR revealed increment in the transcript levels of pcs in transformed Anabaena over wild-type strain using 16S as internal control (Fig. 1b). In addition, western blotting and phytochelatin production indicated higher level of protein and its activity respectively, in AnFPN-pcs over An7120 under normal growth conditions (Fig. 1c, d). The overexpressed strain of Anabaena (AnFPN-pcs) showed similar growth behavior as An7120 under ambient growth condition (Fig. 2a). Nitrogenase activity in terms of acetylene reduction was found higher in recombinant strain as compared to wild type. AnFPN-pcs registered 2.25-fold enhanced nitrogenase activity as compared to wild type, suggesting development of elevated nitrogen fixation capacity in the transformed cyanobacterium (Fig. 2b). The enzymatic activity reveals 5-fold higher phytochelatin synthase activity in recombinant strain as compared to wild type. Comparative growth behavior of wild type and overexpressed strain under LC50 doses of UV-B, salt, heat, copper, carbofuran, and cadmium resulted in decrement in specific growth rate by 26, 21, 27, 13, 30, and 17%, respectively, in An7120 compared to AnFPN-pcs thereby suggesting that the transformed cyanobacterium AnFPN-pcs has developed tolerance and hence showed better growth performances under different abiotic stresses (Fig. 3a–f).

a Colonies corresponding to wild-type Anabaena 7120 (An7120) and recombinant Anabaena AnFPN-pcs strains. b Reverse transcriptase (RT)-PCR of transcript fromAn7120 and AnFPN-pcs cells. Lane 1 (M) shows marker, lane 2 (N) negative control, Lane 3 transcript from An7120, and lane 4 transcript from AnFPN-pcs. c I Western blot image using antibody against phytochelatin synthase. Lane 1 marker, Lane 2 An7120, and Lane 3 AnFPN-pcs. c II SDS PAGE image with equal loading of protein in wild type as well as recombinant strain. d Phytochelatin synthase activity measurements in terms of phytochelatin production in n mole mg protein−1

a Comparison of growth behavior of An7120 and AnFPN-pcs under ambient condition. b Nitrogenase activity assessment in nanomoles C2H2 mg−1 protein−1 h from An7120 and AnFPN-pcs cells under ambient condition
Comparison of differentially expressed proteins in An7120 and AnFPN-pcs
To investigate the effect of pcs gene on cytosolic protein of An7120, 2-DE was performed. On analyzing the electrophoretic profiles of transformed and wild-type strain (as revealed by PDQuest), 389 and 431 spots were found respectively (Fig. 4). A total of 17 proteins depicted significant and reproducible changes in all the gels of An7120 and AnFPN-pcs (Fig. 4). Further, the analysis of the proteins using MALDI-TOF MS and MASCOT showed close homology to molecular chaperone DnaK (spot1), coproporphyrinogen III oxidase (spot2), hypothetical protein Alr4979 (spot3) with conserved domain belonging to DUF3386 superfamily (Fig. S2), manganese superoxide dismutase (spot4), sugar transport ATP-binding protein (spot5), phycocyanin alpha chain (spot6), FKBP-type peptidyl-prolyl cis-trans isomerase (spot7), hypothetical protein All0457(spot8) with conserved domain belonging to ChaB superfamily (Fig. S3), dihydroorotase (spot9), carboxyl-terminal processing protease (spot10), hypothetical protein Alr0946 (spot11) with conserved domain as AbrB superfamily (Fig. S4) and asl0060 (spot12) with conserved domain as DUF427 superfamily (Fig. S5), elongation factor Ts (spot13), acyl carrier protein phosphodiesterase (spot14), ferredoxin-NADP+ reductase (spot15), nutrient stress-induced DNA-binding protein (spot16), and ketol-acid reductoisomerase (spot17) (Table 1).
Dynamics of differentially expressed proteins in An7120 and AnFPN-pcs
The analysis of the MALDI-TOF MS results characterized 17 protein spots in the form of heat map (Fig. 5). When the altered proteomic responses were analyzed, wild-type and transformed cyanobacteria formed two separate clusters. The significantly affected proteins also showed two clear major patterns, one group majorly contains up-accumulated proteins comprising molecular chaperone DnaK, coproporphyrinogen III oxidase, hypothetical protein Alr4979, manganese superoxide dismutase, sugar transport ATP-binding protein, phycocyanin alpha chain, FKBP-type peptidyl-prolyl cis-trans isomerase, dihydroorotase, acyl carrier protein phosphodiesterase, ferredoxin-NADP+ reductase, nutrient stress-induced DNA-binding protein, and ketol-acid reductoisomerase in AnFPN-pcs (at the top of the heat map) while a second group showed the up-accumulated proteins in An7120 including elongation factor Ts, carboxy terminal processing protease, hypothetical protein All0457, and Asl0060 at the bottom of the heat map. The first group represents the overexpression-dependent changes in the proteome of the transformed cyanobacterium (Fig. 5).
Discussion
The present study evaluates the possible role of expression of an extra copy of pcs (alr0975) in abiotic stress tolerance and its impact on cytosolic proteome of transformed strain AnFPN-pcs. The AnFPN-pcs strain showed 5.0-fold higher expression of pcs and enhanced PCS activity adjudged by increased production of phytochelatin over An7120 (Fig. 1c (i), d), thereby offering functional validation of this gene in AnFPN-pcs. Interestingly, no major differences were noticed in the growth of AnFPN-pcs and An7120 under ambient condition (Fig. 2a). However, the AnFPN-pcs exhibited better nitrogen fixation ability as compared to An7120. Such findings suggested that the integration of an extra copy of pcs improved the nitrogen fixation ability in An7120 (Fig. 2b). The better growth performance of AnFPN-pcs over wild type following exposure to different abiotic stresses (Fig. 3a–f) attested the role of pcs in multiple stress tolerance of Anabaena sp. PCC 7120. This may be attributed to the protection of thiols from the damaging effects of physical stresses such as heat and UV-B (Bhargava et al., 2005) through conformational changes of PCs.

a–f Effect of different abiotic stresses on growth behavior of An7120 and AnFPN-pcs. a UV-B, b heat (45 °C), b carbofuran (pesticide), d salt(NaCl), e copper (CuCl2), and f cadmium (CdCl2) on the growth of An7120 and AnFPN-pcs. The mean of three independent experiments are plotted with error bars indicating standard deviations
To understand how pcs overexpression modulates the proteome of the transformed strain, the proteome of An7120 and AnFPN-pcs were compared (Fig. 4). Qualitative and quantitative proteome profiling of wild-type as well as overexpressed strain revealed 17 differentially expressed proteins. Out of 17, 13 proteins were found to be up-accumulated in AnFPN-pcs compared to An7120 as depicted in the heat map (Fig. 5). The up-accumulated proteins in wild-type compared to recombinant strain belong to different functional categories including stress-inducible proteins, photosynthesis and respiration, protein folding and modification, and nucleotide and amino acid biosynthesis and transport. These proteins appear to play a crucial role in providing tolerance against abiotic stress and maintaining metabolism.

Two-dimensional gel electrophoretic images of total cytosolic protein extracts from An7120 and AnFPN-pcs. All gels were run in triplicate. The gels were visualized by CBB staining. The total 18 protein spots differentially expressed were analyzed by MALDI-TOF MS

Heat map and cluster tree representation of the identified proteins from An7120 and AnFPN-pcs. The changes in protein spots were calculated with PDQuest software. Values are means ± S.E. of protein volumes of gels from three independent experiments
Among the up-accumulated proteins in AnFPN-pcs, one of the important proteins involved in chlorophyll biosynthesis is coproporphyrinogen III oxidase (CPO) which catalyzes oxidative decarboxylation of coproporphyrinogen III to form protoporphyrinogen IX in the porphyrin metabolism of cyanobacteria (Cornah and Smith, 2009). The upregulation of this protein probably helped in continuation of chlorophyll synthesis under stressful conditions. Ferredoxin-NADP+ reductase (FNR) belongs to a family of flavoproteins catalyzing the electron flow from ferredoxin to NADP+ and ATP synthase (ATP-B) for ATP synthesis (Carrillo and Vallejos 1987). Its upregulation in AnFPN-pcs was not only involved in the maintenance of photosynthesis and ATP pool under stressful conditions but also in the maintenance of nitrogen fixation by transfer of electrons to the nitrogenase (Sehrautemeier et al. 1984). Thus, the up-accumulation of FNR supported the enhanced nitrogen fixation ability of AnFPN-pcs compared to An7120 (Fig. 2b).
Dihydroorotase involved in the catalysis of reversible interconversion of carbamoyl aspartate and dihydroorotate in the pathway for the synthesis of pyrimidine nucleotides by catalyzing the reversible interconversion of carbamoyl aspartate and dihydrorotate. Liu et al. (2009) reported the importance of dihydroorotase in stress tolerance strategies of rice against salinity and drought. Hence, upregulation of this enzyme helped AnFPN-pcs to have more tolerance against the damaging effects of different stresses.
Up-accumulation of an important HSP, DnaK, in AnFPN-pcs might be involved in folding of unfolded proteins which otherwise is detrimental for regular cellular functioning under stress. It not only ensures proper folding of the polypeptide chains but prevents misfolding and aggregation of proteins generated during cellular stress (Boston et al. 1996). A high level of Mn-SOD in AnFPN-pcs compared to An7120 suggested that the transformed organism has developed better antioxidative defense system to combat oxidative stress. Such findings support the better growth responses as reflected by growth curves (Fig. 3a–f) of the overexpressed strain compared to wild type. In addition, Mn-SOD has been found to be involved in binding with metals ions (Ciriolo et al. 1994).
The sugar transport ATP-binding proteins are is to be involved in the transport of sugar across the membrane. The upregulation of this protein in AnFPN-pcs provides an important way to maintain sugar transport under stressful conditions. Phycocyanin alpha chain in cyanobacteria constitute a substantial fraction of the total soluble proteins (50%); their degradation can supply amino acids for the synthesis of essential proteins under nutrient-limited conditions (Allen and Smith 1969).
The FKBP is an important signaling molecule (Vallon, 2005) involved in the repair of stress-dented proteins which showed up-accumulation in AnFPN-pcs. This might have been helpful in maintaining signaling events as well as proper functioning of important proteins during stress leading to better growth (Fig. 3a–f). Another interesting protein, i.e., DNA-binding protein, has been reported to protect organism under nutrient-starved condition as well as abiotic stresses (Richaud et al., 2001; Narayan et al. 2010) by binding to the DNA of the host organism. The up-accumulation of this protein in AnFPN-pcs might support a better growth under stressful conditions.
Out of the total proteins examined in the present study, 27% were hypothetical proteins with mostly unknown function. One of the hypothetical proteins, Alr0946 (CalA), with conserved domain belonging to an AbrB-like superfamily (Fig. S4), was up-accumulation in the AnFPN-pcs. Proteins with such functional domains are known to be involved in DNA binding and act as a transcriptional regulator of Fe-SOD, a key player of the antioxidative defense system during stress (Agervald et al. 2010a). In addition, Alr0946 (CalA) also interacts with the upstream regions of hypC, a gene encoding a hydrogenase (Agervald et al., 2010b). Such findings argue in favor of efficient hydrogen metabolism of AnFPN-pcs over An7120.
Taken together, the data presented suggests that phytochelatin synthase acts as an intermediate regulator in the complex redox system of the cell by connecting photosynthesis, carbon, and nitrogen assimilation with protection from oxidative stress, maintenance of unfolded proteins, and active transport of nutrient. Owing to the importance of cyanobacteria in nitrogen fixation and adaptation to environmental vagaries and the increasing impact of abiotic stresses in the environment, it can be proposed that stress-induced protein may be used for the engineering of multiple stress-tolerant crop plants capable of growing in variable climate.
References
Agervald Å, Baebprasert W, Zhang X, Incharoensakdi A, Lindblad P, Stensjö K (2010a) The CyAbrB transcription factor CalA regulates the iron superoxide dismutase in Nostoc sp. strain PCC 7120. Environ Microbiol 12:2826–2837. doi:10.1111/j.1462-2920.2010.02255.x
Agervald Å, Zhang X, Stensjo K, Devine E, Lindblad P (2010b) CalA, a cyanobacterial AbrB protein, interacts with the upstream region of hypC and acts as a repressor of its transcription in the cyanobacterium Nostoc sp. strain PCC 7120. Appl Environ Microbiol 76:880–890. doi:10.1111/j.1462-2920.2010.02255.x
Agrawal C, Sen S, Singh S, Rai S, Singh PK, Singh VK, Rai LC (2014) Comparative proteomics reveals association of early accumulated proteins in conferring butachlor tolerance in three N2-fixing Anabaena spp. J Proteomics 96:271–290
Allen MM, Smith AJ (1969) Nitrogen chlorosis in blue-green algae. Arch Microbiol 69:114–120. doi:10.1007/BF00409755
Bhargava P, Mishra Y, Srivastava AK, Narayan OP, Rai LC (2008) Excess copper induces anoxygenic photosynthesis in Anabaena doliolum: a homology based proteomic assessment of its survival strategy. Photosynth Res 96:61–74. doi:10.1007/s11120-007-9285-7
Bhargava P, Srivastava AK, Rai LC, Snigdha U (2005) Phytochelatin plays a role in UV-B tolerance in N2-fixing cyanobacterium Anabaena doliolum. J Plant Physiol 162:1220–1225. doi:10.1016/j.jplph.2004.12.006
Bolchi A, Ruotolo R, Marchini G, Vurro E, Sanità di Toppi L, Kohler A, Tisserant E, Martin F, Ottonello S (2011) Genome-wide inventory of metal homeostasis-related gene products including a functional phytochelatin synthase in the hypogeous mycorrhizal fungus Tuber melanosporum. Fungal Genet Biol 48:573–584. doi:10.1016/j.fgb.2010.11.003
Boston R, Viitanen P, Vierling E (1996) Molecular chaperones and protein folding in plants. Plant Mol Biol 32:191–222. doi:10.1007/BF00039383
Bradford MM (1976) A rapid and sensitive method for the quantification of microgram quantity of proteins utilizing the principle of protein dye binding. Anal Biochem 72:248–254
Carrillo N, Vallejos RH (1987) Ferredoxin-NADP-oxidoreductase. In: Barber J (ed) Topics in photosynthesis 8. Oxford, Elsevier, New York, Amsterdam, pp. 527–560
Chaurasia AK, Apte SK (2009) Overexpression of the groESL operon enhances the heat and salinity stress tolerance of the nitrogen-fixing cyanobacterium Anabaena sp. strain PCC 7120. Appl Environ Microbiol 75:6008–6012. doi:10.1128/AEM.00838-09
Chaurasia AK, Parasnis A, Apte SK (2008a) An integrative expression vector for strain improvement and environmental applications of the nitrogen fixing cyanobacterium, Anabaena sp. strain PCC 7120. J Microbiol Meth 73:133–141. doi:10.1016/j.mimet.2008.01.013
Chaurasia N, Mishra Y, Rai LC (2008b) Cloning, expression and analysis of phytochelatin synthase (pcs) gene from Anabaena sp. PCC7120 offering multiple stress tolerance in Escherichia coli. Biochem Biophys Res Commun 376:225–230. doi:10.1016/j.bbrc.2008.08.129
Ciriolo MR, Civitareale P, Carri MT, De Martino A, Galiazzo F, Rotilio G (1994) Purification and characterization of Ag, Zn-superoxide dismutase from Saccharomyces cerevisiae exposed to silver. J Biol Chem 269:25783–25787
Cobbett C, Goldsbrough P (2002) Phytochelatins and metallothioneins: roles in heavy metal detoxification and homeostasis. Annu Rev Plant Biol 53:159–182. doi:10.1146/annurev.arplant.53.100301.135154
Cornah JE, Smith AG (2009) Transformation of uroporphyrinogen III into protoheme. In:Warren MJ and Smith AG (eds) Tetrapyrroles: birth, life and death, Landes Bioscience and Springer Science Business Media, Austin, pp 74–88.
Gekeler W, Grill E, Winnacker EL, Zenk MH (1989) Survey of the plant kingdom for the ability to bind heavy metals through phytochelatins. Z Naturforsch 44:361–369. doi:10.1093/jxb/erf107
Grill E, Loffler S, Winnacker EL, Zenk MH (1989) Phytochelatins, the heavy metal-binding peptides of plants are synthesized from glutathione by a specific γ-glutamylcysteinedipeptidyltranspeptidase (phytochelatin synthase). Proc Natl Acad Sci U S A 86:6838–6842
Grill E, Winnacker E-L, Zenk MH (1987) Phytochelatins, a class of heavy-metal-binding peptides from plants, are functionally analogous to metallothioneins. Proc Natl Acad Sci U S A 84:439–443
Ha SB, Smith AP, Howden R, Dietrich WM, Bugg S, O’Connell MJ, Goldsbrough PB, Cobbett CS (1999) Phytochelatin synthase genes from Arabidopsis and the yeast, Schizosaccharomyces pombe. Plant Cell 11:1153–1164. doi:10.1105/tpc.11.6.1153
Hall JL (2002) Cellular mechanisms for heavy metal detoxification and tolerance. J Exp Bot 53:1–11
Kneer R, Kutchan TM, Hochberger A, Zenk MH (1992) Saccharomyces cerevisiae and Neurospora crassa contain heavy metal sequestering phytochelatin. Arch Microbiol 157:305–310. doi:10.1007/BF00248673
Liu WY, Wang MM, Huang J, Tang HJ, Lan HX, Zhang HS (2009) The OsDHODH1 gene is involved in salt and drought tolerance in rice. J Integr Plant Biol 51:825–833. doi:10.1111/j.1744-7909.2009.00853.x
Mallick N, Pandey S, Rai LC (1994) Involvement of a cadmium-induced low molecular weight protein in regulating cadmium toxicity in the diazotrophic cyanobacterium Anabaena doliolum. Biometals 7:299–304. doi:10.1007/BF00144125
Marchler-Bauer A, Derbyshire MK, Gonzales NR, Lu S, Chitsaz F, Geer LY, Geer RC, He J, Gwadz M, Hurwitz DI, Lanczycki CJ, Lu F, Marchler GH, Song JS, Thanki N, Wang Z, Yamashita RA, Zhang D, Zheng C, Bryant SH (2015) CDD: NCBI’s conserved domain database. Nucleic Acids Res 43(Database issue):D222–D226. doi:10.1093/nar/gku1221
Mehra RK, Tarbet EB, Gray WR, Winge DR (1988) Metal-specific synthesis of two metallothioneins and γ-glutamil peptides in Candida glabrata. Proc Natl Acad Sci U S A 85:8815–8819
Mishra Y, Chaurasia N, Rai LC (2009) AhpC (alkyl hydroperoxide reductase) from Anabaena sp. PCC 7120 protects Escherichia coli from multiple abiotic stresses. Biochem Biophy Res Commun 381:606–611. doi:10.1016/j.bbrc.2009.02.100
Narayan OP, Kumari N, Bhargava P, Rajaram H, Rai LC (2016) A single gene all3940 (dps) overexpression in Anabaena sp. PCC 7120 confers multiple abiotic stress tolerance via proteomic alterations. Funct Integr Genomics 16:67–78. doi:10.1007/s10142-015-0467-7
Narayan OP, Kumari N, Rai LC (2010) Heterologous expression of Anabaena sp. PCC 7120 all3940 (a dps family gene) protects Escherichia coli from nutrient limitation and abiotic stresses. Biochem Biophy Res Commun 394:163–169. doi:10.1016/j.bbrc.2010.02.135
Padhy RN, Mohapatra K (2001) Toxicity of two carbamate insecticides to the cyanobacterium anabaena PCC 7120 and computations of partial lethal concentrations by the probit method. Microbios 106:81–95
Pandey S, Rai R, Rai LC (2012) Proteomics combines morphological, physiological and biochemical attributes to unravel the survival strategy of Anabaena sp. PCC7120 under arsenic stress. J Proteomics 75:921–937
Pawlik-Skowrońska B (2001) Phytochelatin production in freshwater algae Stigeoclonium in response to heavy metals contained in mining water; effects of some environmental factors. Aquat Toxicol 52(3–4):241–249
Pawlik-Skowrońska B, Pirszel J, Brown MT (2007) Concentrations of phytochelatins and glutathione found in natural assemblages of seaweeds depend on species and metal concentrations of the habitat. Aquat Toxicol 83:190–199. doi:10.1016/j.aquatox.2007.04.003
Pawlik-Skowrońska B, Sanità di Toppi L, Favali MA, Fossati F, Pirszel J, Skowroński T (2002) Lichens respond to heavy metals by phytochelatin synthesis. New Phytol 156:95–102. doi:10.1046/j.1469-8137.2002.00498.x
Rai S, Agrawal C, Shrivastava AK, Singh PK, Rai LC (2014) Comparative proteomics unveils cross species variations in anabaena under salt stress. J Proteomics 98:254–270. doi:10.1016/j.jprot.2013.12.020
Rajaram H, Apte SK (2008) Nitrogen status and heat-stress dependent differential expression of the cpn60 chaperonin gene influences thermotolerance in the cyanobacterium Anabaena. Microbiology 154:317–325. doi:10.1099/mic.0.2007/011064-0
Richaud C, Zabulon G, Jodder A, Thomas JC (2001) Nitrogen and sulphur starvation differentially affects phycobilisomes degradation and expression of the nblA gene in Synechocystis sp. strain PCC 6803. J Bacteriol 183:2989–2994. doi:10.1128/JB.183.10.2989-2994.2001
Rippka R, Deruelles J, Waterbury JB, Herdman M, Stanier RY (1979) Generic assignments, strain histories and properties of pure cultures of cyanobacteria. J Gen Microbiol 111:–61. doi:10.1099/00221287-111-1-1
Saeed AI, Sharov V, White J, Li J, Liang W, Bhagabati N, Braisted J, Klapa M, Currier T, Thiagarajan M, Sturn A, Snuffin M, Rezantsev A, Popov D, Ryltsov A, Kostukovich E, Borisovsky I, Liu Z, Vinsavich A, Trush V, Quackenbush J (2003) TM4: a free, open-source system for microarray data management and analysis. Biotechnol 34:374–378
Sambrook J, Russell DW (2001) Molecular cloning: a laboratory manual. Cold Spring Harbor Laboratory Press. Cold Spring Harbor, New York
Seckbach J and Oren A (2007). Oxygenic photosynthetic microorganisms in extreme environments: possibilities and limitations. Seckbach J (Ed.) Algae and Cyanobacteria in Extreme Environments, pp 3–25, Springer Netherlands.
Sehrautemeier B, Bohme H, Boger P (1984) In vitro studies on pathways and regulation of electron transport to nitrogenase with a cell-free extract from, heterocysts of Anabaena variabilis. Arch Microbiol 137:14–20. doi:10.1007/BF00425801
Seki M, Kamei A, Yamaguchi-Shinozaki K, Shinozaki K (2003) Molecular responses to drought, salinity and frost: common and different paths for plant protection. Curr Opin Biotechnol 14:194–199. doi:10.1016/S0958-1669(03)00030-2
Shrivastava AK, Chatterjee A, Yadav S, Singh PK, Singh S, Rai LC (2015) UV-B stress induced metabolic rearrangements explored with comparative proteomics in three Anabaena species. J Proteomics 127:122–133. doi:10.1016/j.jprot.2015.05.014
Shrivastava AK, Pandey S, Yadav S, Mishra Y, Singh PK, Rai R, Singh S, Rai S, Rai LC (2016) Comparative proteomics of wild type, An+ahpC and An∆ahpC strains of Anabaena sp. PCC7120 demonstrates AhpC mediated augmentation of photosynthesis, N2-fixation and modulation of regulatory network of antioxidative proteins. J Proteomics 140:81–99. doi:10.1016/j.jprot.2016.04.004
Singh PK, Shrivastava AK, Chatterjee A, Pandey S, Rai S, Singh S, Rai C (2015) Cadmium toxicity in diazotrophic Anabaena spp. adjudged by hasty up-accumulation of transporter and signaling and severe down-accumulation of nitrogen metabolism proteins. J Proteomics 127:134–146. doi:10.1016/j.jprot.2015.05.019
Srivastava AK, Ara A, Bhargava P, Mishra Y, Rai SP, Rai LC (2007) A rapid and cost-effective method of genomic DNA isolation from cyanobacterial culture, mat and soil suitable for genomic fingerprinting and community analysis. J Appl Phycol 19:373–382. doi:10.1007/s10811-006-9144-5
Stewart WDP, Fitzgerald GP, Burris RH (1968) Acetylene reduction by nitrogen-fixing blue-green algae. Arch Mikrobiol 62(4):336–348
Tognetti VB, Palatnik JF, Fillat MF, Melzer M, Hajirezaei MR, Valle EM, Carrillo N (2006) Functional replacement of ferredoxin by a cyanobacterial flavodoxin in tobacco confers broad-range stress tolerance. Plant Cell 18:2035–2050
Tognetti VB, Zurbriggen MD, Morandi EN, Fillat MF, Valle EM, Hajirezaei MR, Carrillo N (2007) Enhanced plant tolerance to iron starvation by functional substitution of chloroplast ferredoxin with a bacterial flavodoxin. Proc Natl Acad Sci USA 104 (27):11495–11500
Tsuji N, Nishikori S, Iwabe O, Shiraki K, Miyasaka H, Takagi M, Hirata K, Miyamoto K (2004) Characterization of phytochelatin synthase-like protein encoded by alr0975 from a prokaryote, Nostoc sp. PCC 7120. Biochem Biophys Res Commun 315:751–755. doi:10.1016/j.bbrc.2004.01.122
Vallon O (2005) Chlamydomonas Immunophilins and Parvulins: survey and critical assessment of gene models. Eukary cell 4:230–241. doi:10.1128/EC.4.4.836.2005
Vatamaniuk OK, Mari S, Lu YP, Rea PA (1999) AtPCS1, a phytochelatin synthase from Arabidopsis thaliana: isolation and in vitro reconstitution. Proceedings of the National Academy of Sciences, USA 96:7110–7115. doi:10.1073/pnas.96.12.7110
Wagner T, Windhövel U, Römer S (2002) Transformation of tobacco with a mutated cyanobacterial phytoene desaturase gene confers resistance to bleaching herbicides. Z Natuforsch 57c:671–679
Zhang H, Xu W, Guo J, He Z, Ma M (2005) Coordinated responses of phytochelatins and metallothioneins to heavy metals in garlic seedlings. Plant Sc 169(6):1059–1065
Acknowledgments
L. C. Rai is thankful to CSIR, ICAR, for the financial support; DST, for J C Bose National Fellowship; and DAE for Rajaramanna Fellowship. Neha Chaurasia, Yogesh Mishra, Antra Chatterjee, and Shivam Yadav are thankful to CSIR-UGC JRF. We thank Professor S. K. Apte from BARC Mumbai for providing the pFPN vector. We also thank the Head and Program Coordinator, CAS in Botany, BHU, for the facilities.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Bhumi Nath Tripathi
Electronic supplementary material
ESM 1
(DOCX 1265 kb)
Rights and permissions
About this article

Cite this article
Chaurasia, N., Mishra, Y., Chatterjee, A. et al. Overexpression of phytochelatin synthase (pcs) enhances abiotic stress tolerance by altering the proteome of transformed Anabaena sp. PCC 7120. Protoplasma 254, 1715–1724 (2017). https://doi.org/10.1007/s00709-016-1059-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00709-016-1059-7