
Abstract
We have evaluated the impact of aluminum (Al) on germination, relative root growth, Al accumulation in roots tips, H2O2 levels, plasma membrane integrity, pigment levels, protein content, and the activities of superoxide dismutase (SOD) and catalase (CAT) in seedlings of the endangered Portuguese species Plantago algarbiensis and Plantago almogravensis. We found that up to 400 μM Al had no impact on the germination percentage in either species but inhibited root growth in a concentration-dependent manner (more severely in P. algarbiensis). Al accumulation in the root tips of both species was concentration dependent up to 200 μM but declined thereafter despite the absence of membrane damage. We observed a concentration-dependent induction of SOD activity but no change in CAT activity resulting in the accumulation of H2O2 (a known growth inhibitor), although its impact in P. almogravensis may be partially ameliorated by the accumulation of carotenoid pigments. Our data suggest an association between Al uptake, H2O2 production, and the inhibition of root growth during early seedling development in P. algarbiensis and P. almogravensis, although the latter is more tolerant towards higher concentrations of the metal.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Germination is a complex process involving the coordination of physiological, biochemical, and molecular mechanisms that are particularly susceptible to adverse environmental factors (Bewley 1997). Although the impact of metal toxicity on the growth of vegetative tissues such as shoots and roots has been well documented, there have been few studies focusing on metal toxicity and germination (Kranner and Colville 2011). Physiological metal tolerance strategies in plants depend on age and the nature of the metal ions, but metal-tolerant species are thought to deploy their adaptive mechanisms throughout development, particularly during germination when the young plant is most vulnerable (Lefèvre et al. 2009).
Aluminum (Al) is a major metal component in soils and is solubilized as phytotoxic ions (predominantly Al3+) which inhibit plant growth under low-pH conditions (Tahara et al. 2008). Both germination and seedling growth can be inhibited if the medium surrounding the seed is contaminated with Al (Marciano et al. 2010; Gui et al. 2011). Germination and root growth tests have been used to evaluate the effect of Al in several plant species, particularly crops (Marciano et al. 2010; Zhang et al. 2010). Furthermore, Ezaki et al. (2007, 2008) used germination and relative root growth tests to measure Al tolerance in 49 wild species and in panels of Arabidopsis thaliana mutants.
Plantago almogravensis Franco is an endemic species that grows along the southwest coast of Portugal in acidic, podzolic soils naturally enriched in bioavailable Al and iron (Pimentel et al. 1996; Buurman and Jongmans 2005; Serrano et al. 2011). This species is considered an Al hyperaccumulator plant based on the results of field testing (Branquinho et al. 2007; Serrano et al. 2011). Plantago algarbiensis Samp. is an endemic species from the West-Central Algarve region that occurs in clay-rich soils preferably downstream from small springs or clearings containing acidophilic brushes. Given that both species are currently endangered, micropropagation protocols were established for these species (Gonçalves et al. 2009) in order to provide enough plant material for experiments, allowing us to confirm their ability to grow normally in low-pH medium (Martins et al. 2011). Also, we observed that micropropagated shoots and plantlets of both species accumulate considerable amounts of Al and are moderately tolerant to this metal (Martins et al. 2013a, b, c). These two wild plants may conserve Al resistance genetic information, and therefore, will be useful sources to understand the Al resistance mechanisms and to discover and isolate tolerant genes that can be applied in breeding strategies for crops growing in acidic soils. In addition, due to the endangered nature of these species, the knowledge of their physiological and biochemical response to Al is important for the ecological understanding and for the definition of conservation strategies. The aim of this work was to evaluate the effect of Al on germination and seedling development in P. algarbiensis and P. almogravensis to gain insight into their Al toxicity and tolerance mechanisms during a stage when the young plants are most vulnerable. Although the requirements for germination in both species have been studied (Martins et al. 2012) the behavior of germinating seeds in the presence of Al has not been investigated.
Materials and methods
Plant material
P. algarbiensis and P. almogravensis seeds were collected in June 2010 from wild plants growing among natural populations in Algoz (Algarve Region, Portugal) and Vila Nova de Milfontes (Portugal), respectively. Several plants from both species were selected to ensure the seeds were genetically diverse. The seeds were stored in hermetic glass jars inside paper bags and were kept in the dark under laboratory conditions at 25 ± 2 °C for 2 months.
Seed germination and seedling growth
Seeds were sown in 9-cm glass Petri dishes on 1 mM CaCl2 solution (pH 4.5) solidified with 0.5 % (w/v) bactoagar (Difco, USA) supplemented with 0, 100, 200, or 400 μM AlCl3, corresponding to 0, 78, 158, or 317 μM Al3+, respectively, as estimated by Geochem-EZ (Shaff et al. 2010). Seeds were incubated under optimal germination conditions (15 ± 2 °C, 16-h photoperiod, 69 μmol m−2 s−1) as previously described (Martins et al. 2012). The seeds were monitored every 2 days over 30 days and germination was scored on the basis of visible radical protrusion.
For each treatment, we recorded the final germination percentage (in percent) and the mean germination time (MGT), which was determined using the formula MGT = ΣDN / ΣN (Ellis and Roberts 1981), where D is the number of days from sowing and N is the number of seeds that have germinated by day D. Root length was measured for each treatment after 30 days of germination and relative root growth was estimated by calculating the root lengths of Al-treated seedlings as a percentage of the root length of untreated seedlings (Ezaki et al. 2007).
Aluminum uptake
The content was determined by hematoxylin staining (Polle et al. 1978). Roots were washed in distilled water for 30 min at room temperature, stained in 0.2 % (w/v) hematoxylin solution containing 0.02 % (w/v) KIO3 for 30 min and washed again as above. Stained roots were photographed using a Moticam 350 (Motic, China) under a stereomicroscope (Olympus SZ40, Japan). For quantitation, 5-mm root tips were excised and soaked in 1 M HCl for 1 h to release the hematoxylin, and the concentration was determined by measuring absorbance at 490 nm.
H2O2 content
The H2O2 content of the seedlings was determined as described by Loreto and Velikova (2001). Fresh plant material (100 mg) was homogenized in 1 ml 0.1 % (w/v) trichloroacetic acid at 4 °C and centrifuged at 12,000×g for 15 min. We then mixed 0.2 ml of the supernatant with 0.2 ml 10 mM sodium phosphate buffer (pH 7.0) and 0.4 ml 1 M KI. The reaction was developed for 30 min in darkness and the H2O2 content was determined by measuring absorbance at 390 nm against a set of H2O2 standards, and subtracting a blank sample lacking the plant extract. The results were expressed as micromole per gram fresh weight (FW).
Plasma membrane integrity
Plasma membrane integrity was evaluated using Evans blue as described by Baker and Mock (1994). Intact roots were stained with 0.25 % (w/v) Evans blue solution for 10 min at room temperature, washed three times with distilled water for 10 min and photographed under a stereomicroscope as above. For quantitation, 5-mm root tips were excised and soaked in N,N-dimethylformamide for 1 h at room temperature to release the stain, and the concentration was determined by measuring absorbance at 600 nm.
Photosynthetic pigments
Photosynthetic pigments were extracted with 100 % acetone from fresh plant material (25 mg) and quantified by measuring the absorbance at 661.6, 644.8, and 470 nm (Lichtenthaler 1987).
SOD and CAT activities and soluble protein levels
Superoxide dismutase (SOD; EC 1.15.1.1) and catalase (CAT; EC 1.11.1.6) activities were evaluated in extracts from seedlings after 30 days of germination. Fresh tissue (100 mg) was ground in a pre-chilled mortar containing 50 mM sodium phosphate buffer (pH 7.0), 0.1 mM ethylenediaminetetraacetic acid, 1 % (w/v) polyvinylpolypyrrolidone, and 2.5 mM dithiothreitol. The homogenate was centrifuged at 20,000×g for 10 min at 4 °C and the supernatant was used for subsequent enzyme assays. The specific activity for both enzymes was expressed as enzyme units per milligram of protein, based on the measurement of total soluble protein levels according to the method of Bradford (1976) using bovine serum albumin as standard.
SOD activity was measured by the reduction of nitroblue tetrazolium chloride (NBT) as described by Beauchamp and Fridovich (1971). The reaction mixture comprised 50 mM sodium phosphate buffer (pH 7.8), 10 mM methionine, 0.075 mM NBT, 0.2 mM riboflavin and 20 μl of the enzyme extract. The reaction was monitored by measuring absorbance at 560 nm for 6 min. One unit of SOD was defined as the amount of enzyme required to inhibit NBT reduction by 50 %.
CAT activity was measured by the degradation of H2O2 as described by Aebi (1983). The reaction mixture comprised of 50 mM sodium phosphate buffer (pH 7.0), 40 mM H2O2 and 20 μl of the enzyme extract. The reaction was monitored by measuring absorbance at 240 nm (ε = 39.4 mM−1 cm−1). One unit of CAT was defined as the amount of enzyme required to degrade 1 μmol H2O2 per min.
Data analysis
Final germination percentages, MGT, and relative root growth values were based on data from ten replicates of ten seeds or seedlings, whereas other values were determined from five replicates of plant material randomly collected from different Petri dishes. The values obtained were expressed as means ± standard errors. The impact of different treatments was tested by one-way analyses of variance using the SPSS statistical package for Windows (release 19.0, SPSS Inc., California). We used SigmaPlot version 11.0 (Systat Software, San Jose, CA, USA) for data presentation. Significant differences between means were identified using Duncan’s New Multiple Range Test or Dunnett’s test. Final germination percentages were arcsine transformed prior to statistical analysis.
Results
Effect of Al on seed germination and seedling growth
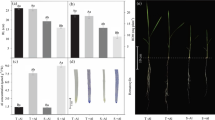
To investigate the effect of Al on germination of P. algarbiensis and P. almogravensis seeds, the final germination percentages, and the mean germination time (MGT) were measured (Table 1). We observed that the final germination percentages and MGT values were not significantly affected (p ≥ 0.05) by the Al loading in both species (Table 1). As shown in the Fig. 1a, there were no visible signs of toxicity such as chlorotic shoots and stubby, brittle, and brown roots in seedlings of both Plantago species exposed to different concentrations of Al even at the highest concentration. The quantitative impact of Al loading was investigated by measuring root growth in Al-treated seedlings relative to root growth in untreated controls. In P. almogravensis seedlings, there was no difference (p ≥ 0.05) in relative root growth at 100 μM Al, a negligible (p ≥ 0.05) reduction at 200 μM Al and even at 400 μM Al the relative root growth was still ∼60 % (Fig. 2). In P. algarbiensis, there was a slight reduction in relative root growth at 100 μM Al, and the values fell (p < 0.05) to approximately 63 and 41 % at 200 and 400 μM Al, respectively.

a Effect of aluminum on P. algarbiensis and P. almogravensis seedling growth (bar = 1 cm). b Aluminum accumulation in roots tips revealed by hematoxylin staining (bar = 1 mm). c Evaluation of plasma membrane integrity in the roots tips using Evans blue (bar = 1 mm). Panels from left to right show seedlings grown in medium containing 0, 100, 200, and 400 μM Al respectively, for 30 days. Images represent at least three seedlings or roots

Effect of aluminum on relative root growth in P. algarbiensis and P. almogravensis seedlings. Values are expressed as the mean ± SE (n = 10). *p < 0.05, significant difference from control (Dunnett’s test)
Al accumulation in root tips
To monitor the accumulation and distribution of Al ions in the two species, we incubated seedling roots with the Al-specific dye hematoxylin. The intensity of hematoxylin staining in the root tips of both species increased in concert with Al loading (Fig. 1b) and the corresponding quantitative data showed that Al accumulates in the roots in a concentration-dependent manner in both species when exposed to media containing up to 200 μM Al (Table 2). However, in both species there was a significant reduction in hematoxylin staining (p < 0.05) between 200 and 400 μM Al (Table 2).
Effect of Al on the H2O2 content
We investigated the H2O2 content of Al-treated seedlings compared to untreated controls, and observed a significant increase (p < 0.05) in both species at the highest Al concentration and also at 200 μM in P. algarbiensis (Fig. 3a).

Effect of aluminum on a H2O2 content, b protein content, c SOD activity, and d CAT activity in P. algarbiensis and P. almogravensis seedlings. Values are expressed as the mean ± SE (n = 5). For each species, mean values are significantly different at p < 0.05, according to Duncan’s test
Effect of Al on the plasma membrane integrity, photosynthetic pigments, and soluble protein levels
To evaluate the oxidative damage caused by Al loading, the plasma membrane integrity, photosynthetic pigments, and soluble protein contents were measured. The integrity of the plasma membrane (measured as Evans blue dye uptake) of roots tips was maintained (p ≥ 0.05) in both species under Al treatment (Fig. 1c; Table 2). We observed no significant differences (p ≥ 0.05) in the total chlorophyll content when we compared Al-treated seedlings and controls, but a significant (p < 0.05) increase in the carotenoid content was observed in Al-treated P. almogravensis seedlings (Table 3). The protein content in seedlings of both species was higher (p < 0.05) in response to Al loading (Fig. 3b).
Effect of Al on SOD and CAT activities
To investigate if the antioxidant system efficiently protects P. algarbiensis and P. almogravensis seedlings from Al-induced oxidative stress we measured the SOD and CAT activities (Fig. 3c, d). In P. almogravensis seedlings, there was a significant increase in SOD activity (p < 0.05) at all three Al concentrations, whereas in P. algarbiensis the significant increase was observed at the two highest concentrations (Fig. 3c). Conversely, Al had no impact (p ≥ 0.05) on CAT activity in either species (Fig. 3d).
Discussion
Germination assays are often used to evaluate the toxicity of exogenous compounds including metals such as Al (Labra et al. 2006; Ezaki et al. 2008). Therefore, we investigated the effect of Al on germination of P. algarbiensis and P. almogravensis seeds. We found no significant differences in either species when we compared the final germination percentages and MGT values between the Al-treated seeds and controls (Table 1). This reflects the behavior of other Al-tolerant plants, which have also been shown to germinate successfully despite high concentrations of Al ions in the environment (Ezaki et al. 2008).
Seedling growth is more sensitive to metal toxicity than germination (Kranner and Colville 2011) and seedling root growth is the most relevant indicator because roots are generally the first point of contact between plants and toxic metals in the environment (Lamhamdi et al. 2011). In this study, there were no overt signs of toxicity in seedlings of both Plantago species exposed to Al (Fig. 1a). However, Al loading inhibited the relative root growth of both species in a concentration-dependent manner, but more severely in P. algarbiensis (Fig. 2). In a previous work, we observed that Al at 400 μM induced root growth inhibition in P. algarbiensis in vitro-grown plantlets but did not affect root growth in P. almogravensis (Martins et al. 2013c).
The extent of Al accumulation in root cells can be easily evaluated through hematoxylin staining that has been widely used to study Al tolerance in several plant species (Polle et al. 1978; Marciano et al. 2010). Our results showed that Al accumulates in the root tips of both species in a concentration-dependent manner up to 200 μM Al (Fig. 1b; Table 2). This suggests that both Plantago species lack effective aluminum exclusion mechanisms and corroborates the Al accumulation capacity already observed in these species (Branquinho et al. 2007; Serrano et al. 2011; Martins et al. 2013a, b, c). However, in both species there was a reduction in hematoxylin staining between 200 and 400 μM Al (Table 2), which could reflect either the induction of Al exclusion mechanisms under high-loading conditions or the consequences of root apex damage caused by metal toxicity.
Excess Al is thought to cause oxidative damage to cells by triggering the production of reactive oxygen species (ROS) that interact with nucleic acids in the embryo as well as stored proteins and lipids in the seed (Pereira et al. 2010). These processes can interfere with germination and seedling development, and also generate potentially harmful mutations (Kranner and Colville 2011). Oxidative damage triggered by Al is primarily thought to result from the production of H2O2 (Yamamoto et al. 2003). The increased H2O2 content observed in seedlings of P. algarbiensis and P. almogravensis grown at the highest Al concentrations (Fig. 3a) seems to be associated with the inhibition of relative root growth (Fig. 2). We also observed Al-induced oxidative damage in P. algarbiensis roots in later stages of development (Martins et al. 2013d). Our results are consistent with previous studies where the H2O2 accumulation increase in plants undergoing Al toxicity (Pereira et al. 2010; Xu et al. 2011) and the resulting oxidative stress may be responsible for the inhibition of root growth (Zhang et al. 2010; Xu et al. 2011).
Membrane lipids and proteins are especially vulnerable to ROS, therefore damaged membranes are considered to be reliable indicators of cell death caused by oxidative stress in plants (Halliwell and Gutteridge 1993). We investigated the potential link between Al loading, the inhibition of root growth, ROS production, and cell death by measuring plasma membrane integrity. The azo dye Evans blue is not taken up by living cells because it cannot cross functional lipid membranes, but it can leak through ruptured membranes and is therefore a sensitive indicator of membrane integrity and cell death (Xu et al. 2011; Zelinová et al. 2011). No differences in dye uptake were observed between Al-treated roots and controls suggesting that the plasma membrane was not damaged to a significant extent by the increased production of ROS in either species (Fig. 1c; Table 2). These findings suggest that ROS production does not inhibit root growth by damaging membranes, although it is possible that other targets including nucleic acids and cytosolic proteins may be involved (Gill and Tuteja 2010).
Although several studies have shown that photosynthetic pigment levels decline in plants exposed to Al (Yadav and Mohanpuria 2009; Pereira et al. 2010), our results showed no significant differences in the total chlorophyll content when we compared Al-treated plants and controls of either species (Table 3) suggesting that Al does not interfere with early photosynthetic activity. In contrast, the carotenoid contents were higher in Al-treated P. almogravensis seedlings (Table 3) indicating that the production of antioxidant pigments may be induced to prevent oxidative damage (Larson 1988).
The soluble protein content of plants is an important indicator of metabolic changes which responds to a wide variety of stresses (Singh and Tewari 2003) and predicts the physiological status of seedlings (Nataraj and Parmar 2008). It is likely that the increased protein content observed in the seedlings of both species in response to Al (Fig. 3b) reflects the de novo synthesis of stress-induced proteins that mediate the adaptive physiological mechanisms of metal tolerance (Verma and Dubey 2003). Al may also be detoxified through the formation of complexes with cytosolic Al-binding proteins (Basu et al. 1999).
The oxidative damage caused by ROS can be prevented by the induction of both enzymatic and non-enzymatic antioxidant systems in plants (Apel and Hirt 2004). These systems are thought to play a substantial role during the germination and development of seedlings exposed to metals (Kranner and Colville 2011), although the precise roles of antioxidant enzymes in Al tolerance have not been investigated in detail. We therefore studied the activities of two pivotal antioxidant enzymes, namely superoxide dismutase (SOD) and catalase (CAT). SOD represents the first step in the detoxifying process, catalyzing the dismutation of O2 ·− to H2O2 and O2, whereas CAT converts H2O2 into harmless oxygen and water. The complementary activities of SOD and CAT are therefore necessary to mitigate oxidative stress caused by superoxides (Benavides et al. 2005). In our study, the increased H2O2 generation observed in P. algarbiensis and P. almogravensis seedlings exposed to Al (Fig. 3a) reflects a parallel increase in SOD activity (Fig. 3c) but the maintenance of CAT activity at normal levels (Fig. 3d).
Our results indicate that both species may be able to tolerate the presence of Al and germinate in the acidic Al-rich soils colonized by them. However, the Al induced H2O2 accumulation and root growth inhibition observed in both species during the initial seedling development, indicate that high Al amounts (≥ 400 μM) may compromise plants establishment in their natural habitats. Moreover, P. almogravensis demonstrated to be more tolerant in this early stage than P. algarbiensis, which is in agreement with our previous results (Martins et al. 2013c, d) and its predominance in geochemical islands rich in Al and their inexistence outside these areas with the surrounding vegetation, reflecting an obligate metallophyte behavior (Serrano et al. 2011). The concentration-dependent inhibition of root growth we observed was associated with a quantitative increase in H2O2 levels resulting from the induction of SOD with no compensatory increase in the level of CAT activity. However, the higher levels of H2O2 were not sufficient to induce membrane damage suggesting that the inhibition of root growth reflects the impact of oxidative stress on intracellular targets, such as cytosolic proteins and nucleic acids, although this issue should be clarified in future works.
Abbreviations
- CAT:
-
Catalase
- FW:
-
Fresh weight
- MGT:
-
Mean germination time
- NBT:
-
Nitroblue tetrazolium
- ROS:
-
Reactive oxygen species
- SOD:
-
Superoxide dismutase
References
Aebi HE (1983) Catalase. In: Bergmeyer HU (ed) Methods of enzymatic analysis. Verlag Chemie, Weinhern, pp 273–286
Apel K, Hirt H (2004) Reactive oxygen species: metabolism, oxidative stress, and signal transduction. Annu Rev Plant Biol 55:373–399
Baker CJ, Mock NM (1994) An improved method for monitoring cell death in cell suspension and leaf disc assays using Evan’s blue. Plant Cell Tissue Organ Cult 39:7–12
Basu U, Good AG, Aung T, Slaski JJ, Basu A, Briggs KG, Taylor GJ (1999) A 23-kDa, root exudate polypeptide co-segregates with aluminum resistance in Triticum aestivum. Physiol Plant 106:53–61
Beauchamp CO, Fridovich I (1971) Superoxide dismutase: improved assays and assays applicable to acrylamide gels. Anal Biochem 44:276–287
Benavides MP, Gallego SM, Tomaro ML (2005) Cadmium toxicity in plants. Braz J Plant Physiol 17:21–34
Bewley DJ (1997) Seed germination and dormancy. Plant Cell 9:1055–1066
Bradford M (1976) A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Branquinho C, Serrano HC, Pinto MJ, Martins-Loução MA (2007) Revisiting the plant hyperaccumulation criteria to rare plants and earth abundant elements. Environ Pollut 146:437–443
Buurman P, Jongmans AG (2005) Podzolisation and soil organic matter dynamics. Geoderma 125:71–83
Ellis RH, Roberts EH (1981) The quantification of ageing and survival in orthodox seeds. Seed Sci Technol 9:373–409
Ezaki B, Kiyohara H, Matsumoto H, Nakashima S (2007) Overexpression of an auxilin-like gene (F9E10.5) can suppress Al uptake in roots of Arabidopsis. J Exp Bot 58:497–506
Ezaki B, Nagao E, Yamamoto Y, Nakashima S, Enomoto T (2008) Wild plants, Andropogon virginicus L. and Miscanthus sinensis Anders, are tolerant to multiple stresses including aluminum, heavy metals and oxidative stresses. Plant Cell Rep 27:951–961
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Gonçalves S, Martins N, Romano A (2009) Micropropagation and conservation of endangered species Plantago algarbiensis and P. almogravensis. Biol Plant 53:774–778
Gui R, Leng H, Zhuang S, Zheng K, Fang W (2011) Aluminum tolerance in Moso Bamboo (Phyllostachys pubescens). Bot Rev 77:214–222
Halliwell B, Gutteridge JMC (1993) Free radicals in biology and medicine. Clarendon, Oxford
Kranner I, Colville L (2011) Metals and seeds: biochemical and molecular implications and their significance for seed germination. Environ Exp Bot 72:93–105
Labra M, Gianazza E, Waitt R, Eberini I, Sozzi A, Regondi S, Grassi F, Agradi E (2006) Zea mays L. protein changes in response to potassium dichromate treatments. Chemosphere 60:1234–1244
Lamhamdi M, Bakrim A, Aarab A, Lafont R, Sayah F (2011) Lead phytotoxicity on wheat (Triticum aestivum L.) seed germination and seedlings growth. R C Biologies 334:118–126
Larson RA (1988) The antioxidants of higher plants. Phytochem 27:969–978
Lefèvre I, Marchal G, Corréal E, Zanuzzi A, Lutts S (2009) Variation in response to heavy metals during vegetative growth in Dorycnium pentaphyllum Scop. Plant Growth Regul 59:1–11
Lichtenthaler HK (1987) Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Meth Enzymol 148:350–382
Loreto F, Velikova V (2001) Isoprene produced by leaves protects the photosynthetic apparatus against ozone damage, quences ozone products, and reduces lipid peroxidation of cellular membranes. Plant Physiol 127:781–787
Marciano DPRO, Ramos FT, Alvim MN, Magalhaes JR, França MGC (2010) Nitric oxide reduces the stress effects of aluminum on the process of germination and early root growth of rice. J Plant Nutr Soil Sci 173:885–891
Martins N, Gonçalves S, Palma T, Romano A (2011) The influence of low pH on in vitro growth and biochemical parameters of Plantago almogravensis and P. algarbiensis. Plant Cell Tissue Organ Cult 107:113–121
Martins N, Gonçalves S, Palma T, Romano A (2012) Seed germination of two critically endangered plantain species, Plantago algarbiensis and P. almogravensis (Plantaginaceae). Seed Sci Technol 40:144–149
Martins N, Gonçalves S, Andrade P, Valentão P, Romano A (2013a) Changes on organic acid secretion and accumulation in Plantago almogravensis Franco and P. algarbiensis Samp under aluminum stress. Plant Sci 198:1–6
Martins N, Gonçalves S, Romano A (2013b) Metabolism and aluminum accumulation in Plantago almogravensis and P. algarbiensis in response to low pH and aluminum stress. Biol Plant 57:325–331
Martins N, Osório ML, Gonçalves S, Osório J, Palma T, Romano A (2013c) Physiological responses of Plantago algarbiensis and P. almogravensis shoots and plantlets to low pH and aluminum stress. Acta Physiol Plant 35:615–625
Martins N, Osório ML, Gonçalves S, Osório J, Romano A (2013d) Differences in Al tolerance between Plantago algarbiensis and P. almogravensis reflect their ability to respond to oxidative stress. Biometals. doi:10.1007/s10534-013-9625-3
Nataraj M, Parmar S (2008) Biochemical response during the germination of raya and fenugreek seeds under heavy metal stress. J Cell Tissue Res 8:1589–1594
Pereira LB, Mazzanti CM, Gonçalves JF, Cargnelutti D, Tabaldi LA, Becker AG, Calgaroto NS, Farias JG, Battisti V, Bohrer D, Nicoloso FT, Morsch VM, Schetinger MR (2010) Aluminum-induced oxidative stress in cucumber. Plant Physiol Biochem 48:683–689
Pimentel NL, Wright VP, Azevedo TM (1996) Distinguishing early groundwater alteration effects from pedogenesis in ancient alluvial basins: examples from the Palaeogene of southern Portugal. Sediment Geol 105:1–10
Polle E, Konzak CF, Kittrick JA (1978) Visual detection of aluminum tolerance levels in wheat by hematoxylin staining of seedling roots. Crop Sci 18:823–827
Serrano HC, Pinto MJ, Martins-Loução MA, Branquinho C (2011) How does an Al-hyperaccumulator plant respond to a natural field gradient of soil phytoavailable Al? Sci Total Environ 409:3749–3756
Shaff JE, Schultz BA, Craft EJ, Clark RT, Kochian LV (2010) GEOCHEM-EZ: a chemical speciation program with greater power and flexibility. Plant Soil 330:207–214
Singh PK, Tewari RK (2003) Cadmium toxicity induced changes in plant water relations and oxidative metabolism of Brassica juncea L. plants. J Environ Biol 24:107–112
Tahara K, Yamanoshita T, Norisada M, Hasegawa I, Kashima H, Sasaki S, Kojima K (2008) Aluminum distribution and reactive oxygen species accumulation in root tips of two Melaleuca trees differing in aluminum resistance. Plant Soil 307:167–178
Verma S, Dubey RS (2003) Lead toxicity induces lipid peroxidation and alters the activities of antioxidant enzymes in growing rice plants. Plant Sci 164:645–655
Xu FJ, Jin CW, Liu WJ, Zang YS, Lin XY (2011) Pretreatment with H2O2 alleviates aluminum-induced oxidative stress in wheat seedlings. J Integr Plant Biol 53:44–53
Yadav SK, Mohanpuria P (2009) Responses of Camellia sinensis cultivars to Cu and Al stress. Biol Plant 53:737–740
Yamamoto Y, Kobayashi Y, Devi SR, Rikiishi S, Matsumoto H (2003) Oxidative stress triggered by aluminum in plant roots. Plant Soil 255:239–243
Zelinová V, Haluškova L, Huttová J, Illéš P, Mistrík I, Valentovičová K, Tamás L (2011) Short-term aluminium-induced changes in barley root tips. Protoplasma 248:523–530
Zhang H, Tan Z-Q, Hu L-Y, Wang S-H, Luo J-P, Jones RL (2010) Hydrogen sulfide alleviates aluminum toxicity in germinating wheat seedlings. J Integr Plant Biol 52:556–567
Acknowledgments
N. Martins and S. Gonçalves acknowledge grants SFRH/BD/48379/2008 and SFRH/BPD/31534/2006 from the Portuguese Science and Technology Foundation (FCT). This work was supported by the FCT project PTDC/AGR-AAM/ 102664/2008.
Conflicts of interest
The authors declare that they have no conflicts of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling Editor: Néstor Carrillo
Rights and permissions
About this article
Cite this article
Martins, N., Gonçalves, S. & Romano, A. Aluminum inhibits root growth and induces hydrogen peroxide accumulation in Plantago algarbiensis and P. almogravensis seedlings. Protoplasma 250, 1295–1302 (2013). https://doi.org/10.1007/s00709-013-0511-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00709-013-0511-1