
Abstract
The poultry industry has emerged as one of the largest and fastest growing public sectors in the developed and developing countries. Unfortunately, this industry is under a major threat from diseases that are viral (Newcastle disease, infectious bursal disease, influenza, hydropericardium syndrome), bacterial (colibacillosis, pasteurellosis, salmonellosis, mycoplasmosis), parasitic (coccidiosis, histoplasmosis) or nutritional (dyschondroplasia, osteoporosis). Among these diseases, hydropericardium syndrome (HPS) is one of the important emerging diseases occurring in the specific areas of the world where broilers (chickens) are reared under intensive conditions. HPS was first observed in 1987 at Angara Goth, an area near Karachi, Pakistan, where broilers are raised. Since then, HPS has been reported in many countries of the world. From these reported cases, an adenovirus that was either isolated from or visualized electron microscopically in the liver of affected broilers has been implicated in the syndrome. The syndrome has been reproduced by inoculation of isolated fowl adenovirus (FAdV) strains, and hence, the syndrome is also called infectious hydropericardium syndrome. To our knowledge, HPS has not been observed in humans, so it is not considered a zoonotic disease, but it is of economic importance and causes huge losses to the poultry industry. Efforts have been made to develop conventional vaccines against this disease, which were formulated from infected liver homogenate. Formalin-inactivated liver organ vaccines have failed to protect the poultry industry. Hence, there is a dire need to develop a suitable vaccine to combat this disease. Currently, recombinant vaccine candidates are being developed by using molecular biology and biotechnological approaches for the prevention and control of infectious diseases, including HPS. Therefore, it is suggested that the immunogenicity of these recombinant proteins should be evaluated for their use as subunit vaccines.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Hydropericardium syndrome (HPS)
History
HPS was first reported in 1987 in 3- to 6-week-old broiler chickens in Karachi, Pakistan [57, 66]. HPS was described as an accumulation of a straw-colored “jelly”-like fluid in the pericardial sac, and discolored and enlarged liver with basophilic intranuclear inclusion bodies. Kidneys were congested and high mortality rates (up to 70%) were observed [5, 13, 48]. The disease was observed in rapidly growing broilers [11, 24, 57]. Occasionally, the disease was also reported in 20-week-old layers and breeders and wild pigeons [14, 86].
Inclusion body hepatitis (IBH), which had previously been reported in the United States in 1963, has some clinical similarities with this emerging syndrome, but the accumulation of fluid in the pericardial sac was not found in inclusion body hepatitis. Therefore, researchers differentiated this syndrome from inclusion body hepatitis and a new name, “infectious hydropericardium” was given [2, 80]. Later, HPS was observed in various parts of the world, and a few other names were coined for this syndrome, such as Angara disease [57] and Litchi heart disease [42].
Clinical signs/pathology
Diseased birds exhibit different signs, including lethargy, huddling with ruffled feathers, loss of appetite, and mucoid yellow droppings. Reduction in weight gain has been recorded in infected flocks, which results in a poor feed conversion ratio [13]. Asrani et al. [14] indicated that the birds remained active until death, and in the terminal stages of infection, the chicks were dull, depressed with ruffled feathers, and reluctant to move, and they tended to gather in corners.
Parameters of clinical pathology indicate severe anemia and significant reduction of all hematological values in birds with HPS [14, 89]. Serum protein profiles were also found altered, with a decrease in albumin and an increase in β-globulin [73]. Reduction in the osmotic pressure of colloidal plasma results in the leakage of fluid into the pericardial sac. Blood glucose and plasma protein levels are decreased, while serum uric acid, potassium, calcium and triglycerides levels are significantly increased, which appears to be due to accumulation of fluid in the pericardial sac and abdomen [19]. Serum enzyme activities of AST, ALT and CPK are low in normal birds and high in HPS-infected birds [56, 128]. All of these changes indicate involvement of the liver, kidney and heart in HPS.
Necropsy findings
Gross lesions found during necropsy include enlarged, pale, friable liver, occasionally with necrotic foci. Accumulation of fluid in the pericardium is the most prominent macroscopic lesion [13, 24, 96]. Lesions are observed in vital organs, including the liver, heart, kidneys and lungs. In addition, an enlarged spleen and atrophy of the thymus are observed in most dead birds with HPS. Up to 20 mL of clear, straw-colored fluid is observed in the pericardial sac, giving it a balloon-like shape [14, 24, 70]. The liver is swollen, congested, friable and yellow colored with areas of necrosis and patechial hemorrhages. Kidneys are pale, swollen and friable with urates in tubules [95]. Researchers have observed the same lesions during experimental induction of HPS [12, 31].
Etiology
Initially, a nutritional disorder was considered to be the cause of HPS. The causative factors were thought to be fishmeal, rancid fat, and vitamin and mineral imbalances [57, 96]. Anjum [12] attempted to reproduce the disease using various strategies, including using feed samples from farms where natural outbreaks of the disease had occurred and feed with high levels of mycotoxins (100 ppm), but subcutaneous inoculation with infected liver homogenate was able to transmit the disease successfully, which supported the possibility of an infectious nature of this disease. A viral etiology of the disease was indicated by the presence of basophilic intranuclear inclusion bodies in hepatic cells [13, 41, 42]. The detection of icosahedral-shaped adenovirus particles in a liver extract by electron microscopy verified the viral etiology [24]. Later, Mazaheri et al. [80] reported biotypes of fowl adenovirus 4 to be the cause of the disease. It was found that fowl adenovirus 12 alone or in association with fowl adenovirus 4 was responsible for inclusion body hepatitis/hydropericardium syndrome in poultry flocks in India [99]. In Pakistan, adenovirus serotype 4 was isolated from field cases of HPS [125]. Mazaheri et al. [80] characterized field isolates of fowl adenovirus from several outbreaks of HPS and found that a serum neutralization test showed a positive reaction against antibodies of serotype 4 and 10. It was revealed using immunological and molecular techniques that fowl adenoviruses 4 (FAdV-4) and 10 are closely related [33]. Zhang et al. [129] have reported that HPS is prevalent in many countries, including China. They found that FAdV-4 has been the main causative agent of HPS in China for many years. However, they observed that the virulence of this virus has been increasing since 2015.
Natural transmission
Hydropericardium syndrome is a highly pathogenic disease [66], that can spread rapidly from flock to flock and farm to farm [27]. It is a contagious disease that can be transmitted horizontally either mechanically or by the oral-fecal route [2, 22, 107]. Cowen et al. [28] have demonstrated a role of wild birds in the spread of the disease. Hafez [45] showed that vertical transmission can occur through eggs and that the virus can spread from parent to progeny.
Experimental transmission
The infectious nature of the disease was demonstrated by experimental transmission of the disease by subcutaneous inoculation with an infected liver homogenate [12, 14, 23]. Anjum [12] and Naeem et al. [87] attempted to reproduce the disease experimentally using an infected liver homogenate via the subcutaneous route, the oral route and by direct contact with infected birds. Successful experimental reproduction of disease was observed within 2-5 days after subcutaneous inoculation, with typical symptoms of the disease. However, the oral route was suggested to be the natural route of infection.
Epidemiology
The first epidemic of HPS was reported in 1987 on poultry farms in Angara Goth, which is an extensive broiler-producing area near Karachi, Pakistan [57, 65]. Later, the disease was reported in different countries, including Iraq [1], India [42, 85], Mexico, Ecuador, Peru, Chile [28], South and Central America [117], Canada [43], Hungary [59], Poland [91], Korea [26], Russia [20], China [129] and Japan [3, 78]. Apart from the broilers, rare outbreaks were also reported in other avian species, including pigeons and quail [60, 86]. Asthana et al. [15] reported that FAdVs are commonly present in chickens worldwide and that HPS is the most severe disease associated with FAdV serotype 4.
Zhang et al. [129] reported that during the past few years, the incidence of HPS has increased in many countries, including China. It was found that FAdV-4 was present in China before 2014 but no severe HPS outbreaks were reported. However, recent outbreaks of HPS have been reported with high mortality rates on broilers in small and medium-sized chicken farms in China. Moreover, duck farms have also experienced heavy economic losses due to HPS. It has been shown that strains circulating in China before 2014 and after 2015 had different ancestors and that new strains circulating in China were derived from earlier Indian strains, with some genetic differences.
Adenoviruses
Nicklin et al. [90] and Robinson et al. [104] have shown that adenoviruses (Ads) have non-enveloped virions with a diameter of 70-90 nm. Adenoviruses were discovered in the 1950s in outgrowths of human adenoids and tonsils. It was found that humans and other vertebrates are infected by adenoviruses. Different isolates differ in their ability to agglutinate rat and rhesus erythrocytes. Based on these properties, adenoviruses have been classified into five subgroups [30].
Aviadenoviruses
Aviadenoviruses are a diverse group of pathogens that cause a variety of infections in birds [34, 105]. The aviadenoviruses are further divided into three groups. Group I comprises 12 serotypes of avian adenoviruses from chickens, geese, turkeys and other species that have a common group of antigens [64, 82, 121]. The aviadenovirus that causes HPS is classified as group I, serotype 4 [29, 52, 88, 122]. The virus responsible for acute inclusion body hepatitis is classified as group I, serotype 8 [33, 44, 101].
The second group of avian adenoviruses is known as group II, and its members which cause infections such as hemorrhagic enteritis in turkeys, marble spleen disease in pheasants, and splenomegaly in chickens. The group II viruses also share a common group of antigens that distinguishes them from other groups [32].
The third group of avian adenoviruses is known as group III, and its members are responsible for egg drop syndrome in laying chickens [83].
Morphological characteristics
Aviadenoviruses are non-enveloped virus with icosahedral morphology and a diameter of approximately 90 nm. The viral capsid is composed of 720 hexon subunits arranged as 240 trimers and 12 vertex penton capsomers with one or two fibers protruding from the surface. Aviadenoviruses are easily identified by their typical icosahedral shape and triangular facets of capsomers. This symmetry is typical for all members of the family Adenoviridae [124].
Shah et al. [114], Nicklin et al. [90], and Laver et al. [71] observed aviadenoviruses by electron microscopy and found hexagonal-shaped viral particles with a diameter of 70 to 90 nm.
Molecular characteristics
The aviadenovirus genome is a linear, non-segmented, dsDNA that is 35–36 in kbp length. The DNA has guanine/cytosine content of 53–59%. Viral genome encodes about 40 proteins, and it has terminal redundant sequences with inverted terminal repeats (ITR). Both ends of the viral genome are covalently attached to terminal proteins (TP) [124].
Russel [108, 109] reported that adenovirus genome encodes about 13 structural proteins. He found that the capsid of the viral particle consists of three major proteins, i.e., hexon, penton base and knobbed fiber. The remaining proteins are minor structural proteins including cement proteins (VI, VIII, IX, IIIa) and core proteins (V, VII, Mu, terminal protein, IVa2, protease).
Immunological characteristics
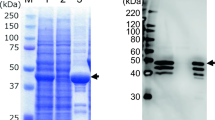
Balamurugan et al. [17] and Kumar and Chandra [68] studied the protein profiles of FAdV-4 and reported eight to twelve protein fractions of isolates with molecular weights between 13.8 and 110 kDa. Among these protein fractions, seven immunogenic polypeptides were detected (with molecular weights from 15.8 to 110 kDa) using western blot analysis with FAdV-4 serum. Hexon, penton base and fiber were found among the highly immunoreactive fractions. Fingerut et al. [36] and Jucker et al. [58] found that structural capsid proteins of adenovirus, i.e. penton and fiber, interact with cell receptors during penetration of the virus into the cell. Among the nonstructural proteins, 100K has importance, because Hong et al. [53] described that this protein may have a role in intracellular transport and folding of viral proteins during viral replication.
Phylogeny and genetic organization
According to Davison et al. [30], adenoviruses are non-enveloped, icosahedral viruses that belong to the family Adenoviridae which is divided into five genera, i.e., Atadenovirus, Siadenovirus, Mastadenovirus, Aviadenovirus and Ichtadenovirus. They replicate inside the nucleus. They have double-stranded, 35- to 36-kbp linear DNA. They are classified as medium-sized DNA viruses. The genomes have inverted terminal repeats (ITR) ranging from 36 to 200 bp, and their 5’ ends have a terminal protein (TP).
Mase et al. [77] compared the nucleotide sequences of the short fiber genes of various fowl adenovirus 4 isolates from Japan, India and Pakistan. Phylogenetic analysis showed that the fowl adenovirus 4 isolates from chickens with HPS grouped into a different cluster from fowl adenovirus 4 strains isolated from non-HPS cases. It was therefore suggested that fowl adenovirus 4 strains can be distinguished on the basis of the nucleotide sequence of the short fiber gene.
Harrach et al. [46] suggested a new classification for adenoviruses in which turkey adenovirus was assigned to the genus Aviadenovirus and the species in the family were renamed by adding a letter designation, i.e., “Possum adenovirus A, Goose adenovirus A, Canine adenovirus A, Tree shrew adenovirus A and Frog adenovirus A”.
Proteins encoded by aviadenoviruses
During viral replication, aviadenoviruses penetrate into the nucleus of host cells. Transcription of early and late genes encoding viral proteins and assembly of new virions occurs in the nucleus. The capsid consists of three major structural proteins, the hexon, fiber, and penton base. The hexon forms the majority of the structural components of the capsid, which has 240 trimeric hexon capsomeres and 12 pentameric penton bases. The trimeric fiber protein protrudes from the penton base at each of the 12 vertices of capsid and has a knobbed rod-like structure. The major difference in the capsids of fowl adenovirus C is the presence of two thin fibers, as most other aviadenoviruses have one fiber [76]. The structure and function of some important viral proteins are described below.
Hexon
The icosahedral capsid of adenovirus is mainly composed of hexon protein. There are four types of hexon, designated as H1, H2, H3 and H4 [21]. H1-type hexons are sixty in number, and they are associated with pentons at the 12 apices, so they are also called peripentonal hexons. The remaining hexons are designed as ‘group of nine’ (GON) on the 20 faces of the icosahedra. The H2-type hexon are on the twofold axes, H3 are on the threefold axes, and the remaining ones are H4. The size of hexon is different in the different serotypes, the largest hexon comprises of 967 amino acids. Each hexon has up to nine hypervariable regions, and they are situated at the top of the molecule [111]. These regions represent the type-specific antigens of the hexon, and at least one of them has a major role in virus-neutralizing activity [79, 103, 110].
Penton base
The penton base monomer comprises approximately 471 amino acids. The covalent complex of two proteins (homopentameric penton base and homotrimeric fiber protein) forms the penton capsomere. The penton base is situated at strategic positions at the apices of the icosahedral capsid, and it plays a key role in stabilization of the capsid as it interacts with the neighbouring capsomeres, peripentonal hexons, and other associated proteins. Moreover, the penton base is sensitive to heat, trypsin, pH and changes in ionic strength [126]. It has been found that the penton base, in association with some other proteins plays a major role during penetration and entry of the virion into the cell [35]. At the post-entry stage, the role of the penton base is not clear, but it has been found that the penton base protein interacts with cellular components, and neutralizing antibodies against the penton base have been detected in the sera of patients [54].
Fiber
The fiber polypeptide comprises approximately 582 amino acids, and it binds noncovalently to the penton base from its N-terminal side [130]. The fiber has three regions, including the tail, shaft and knob. The fiber is the first component of the virus that interacts with host cells/tissues. The ability of the virus to bind to the host cell is a key feature of the infection process and affects the pathogenicity of the virus. Variable sequences on the fiber knob determine the haemagglutinating properties of adenoviruses, and these properties are used to classify the species from A to F [92]. The fiber knob also has an important role in fiber protein synthesis and encapsidation [49].
Studies on the morphology of the penton of CELO virus revealed that aviadenoviruses have a different pentameric base structure, which is associated with two fibers, one long and one short. It was found that the two fibers have different receptors, so it was suggested that both fibers are required, one for virus attachment and the other for internalization [51, 119]. Mase et al. [77] determined the nucleotide sequence of the short fiber of fowl adenovirus serotype 4, and its amino acid sequence was deduced. They found that the short fiber comprises 479 amino acids, and its nucleotide sequence can be used to differentiate various fowl adenovirus serotype 4 strains. It was also found that epitopes affecting virulence, cell-surface-binding receptors, and receptors for tissue tropism exist within the fiber. Thus, the main role of the fiber protein is tethering (attachment) of the viral capsid to the host cell surface by its interaction with cellular receptors [90, 109].
100K
100K is an important nonstructural protein (NSP) of the virus. Xie et al. [127] found that antibodies specific for the 100K and 33K nonstructural proteins were detected in chickens experimentally infected with FAdVs, but not in chickens vaccinated with inactivated FAdVs. They also found that antibodies against NSPs can be used to distinguish FAdV-infected from vaccinated chickens [116]. This protein has been shown to play an important role in transport of viral proteins during viral replication in host cells. The 100K protein of adenoviruses of subgroups B and C has been identified as a chaperone protein that assists in trimerization and nuclear localization of hexons. Experiments have been conducted in which the hexon protein was expressed alone in insect cells or co-expressed in the presence of the 100K protein. When expressed alone, the hexon protein was detected in the cytoplasm in inclusion bodies, but when the hexon and 100K proteins were co-expressed, surprisingly, the hexon was found in a soluble trimeric form. It has been suggested that the 100K protein acts as a scaffold protein for the hexon, because evidence suggests that it plays a role in folding, self-assembly and nuclear import of the hexon in insect cells [53].
Diagnosis
Different techniques have been used by various researchers for accurate and precise diagnosis of HPS, which include conventional microbiological and molecular techniques.
Conventional microbiological techniques
Clinical diagnosis of HPS is a difficult task because of the sudden onset and acute nature of the disease [37, 38, 72]. Usually, initial diagnosis has been done by observing gross lesions and by histopathology by detection of intra-nuclear inclusion bodies of hepatocytes [13]. The diagnostic significance of electron microscopy has been discussed by Cheema et al. [24]. The presence of icosahedral viral particles detected by negative staining electron microscopy in an infected liver homogenate supported the view that the disease was caused by an aviadenovirus. Isometric, roughly spherical particles with morphology resembling that of adenoviruses were also found using transmission electron microscopy [23, 39].
Microbiological and serological methods that are used for diagnosis of subclinical HPS include agar gel precipitation test [47], indirect haemagglutination assay [75, 102], dot immunobinding assay (Rabbani et al., [98], enzyme-linked immunosorbent assay [47, 112], immunoperoxidase test [106], and tissue culture and embryonated hen egg culture along with virus neutralization test [97].
The ability of the virus to be propagated in embryonated hen eggs confirmed the viral etiology of the disease [24, 61, 62]. Mahmood and Hassan [73] propagated the fowl adenovirus in embryonated duck eggs. They injected the virus through the yolk sac and chorioallantoic sac. Death of embryos along with stunted growth and hemorrhages were reported by both routes. They also reported intranuclear inclusion bodies in hepatocytes of inoculated embryos.
The virus has also been propagated and isolated from cell culture [8, 17, 18, 22]. Khawaja et al. [66] propagated and isolated the virus in cell culture of chicken embryo kidney cells. They reported cytopathic effects in cell culture, e.g., degeneration, detachment of cells from the surface, and basophilic intra-nuclear inclusion bodies. Afzal and Ahmad [4] made some efforts to propagate the HPS-associated virus in QT 35 and Vero cell lines, but the virus could not be successfully cultured in these cell lines and the virus was unable to produce any cytopathic effects. Roy et al. [106] adapted the HPS virus to Vero cells by subjecting it to four blind passages. Later, they observed characteristic cytopathogenic effects (CPE) 96 hours after inoculation of Vero cells.
Kumar et al. [69] and Mahmood et al. [72] cultivated the HPS-associated virus in chicken embryo hepatocyte suspension and found a biological titer of 103.6 LD50 units per mL, while the same virus showed a biological titer of 105.6 LD50 units per gram of infected liver homogenate. Thus, it was found that 100 times more virus can be harvested from infected liver homogenate.
Molecular diagnostic techniques
In the case of adenoviruses, the use of serological techniques is associated with problems in interpretation of results, because antibodies are commonly present in both healthy and infected birds [45, 120]. Molecular techniques target and detect the pathogen itself rather than the antibodies induced by it, and in this way, early detection of the disease is ensured. The invention of the polymerase chain reaction (PCR) technique has provided high sensitivity and specificity for the diagnosis of HPS virus by targeting the hexon gene [29, 40, 120, 123]. Ganesh et al. [40] extracted viral DNA from infected liver tissue or purified virus and used it to amply a variable part of the hexon gene by PCR using specific primers. The results revealed the successful amplification of a 700-bp fragment of the hexon gene. This fragment of the hexon gene was subsequently also used as a probe for detection of the virus by dot blot hybridization of viral DNA. Rau and Hess [100] and Hess [50] amplified a 1319-bp portion of the hexon gene using specific primers. They also studied the restriction fragment length polymorphism (RFLP) after digestion of the PCR product with the enzyme HpaII. Mase et al. [77] found that PCR-RFLP analysis of the short fiber gene using the enzyme AluI was useful to distinguish various fowl adenovirus 4 strains. Thakor et al. [120] amplified an 890-bp DNA fragment of the hexon gene using gene-specific primers for detection of HPS-IBH virus in field samples.
In developing countries, in spite of keeping in view the economic importance of the disease, none of the techniques described above have been applied in the field for diagnosis and control of disease [65].
Prevention and control
Various strategies have been used by researchers for prevention and control of HPS since the initial outbreaks in 1987. It has been suggested that maintaining good husbandry practices, such as disinfection, maintaining strict biosecurity, and proper ventilation, may significantly reduce the chances of infection [10]. The addition of iodophor solution to drinking water at a concentration of 0.07-0.1% has been found to be efficacious in reducing the course and mortality of HPS in broilers [1, 2]. In cases of outbreaks, there is no specific treatment available against this viral disease; however, antibiotics are used for prevention of secondary bacterial infections [84, 118].
Vaccines
Standard vaccines are available against many other poultry diseases, but there are few against HPS. Two types of vaccines are currently reported in the literature against HPS. These include parenteral inactivated (killed) whole-cell vaccine and oral live attenuated vaccine. A list of the various types of available licensed vaccines against HPS are given in Table 1.
Parenteral inactivated (killed) whole-cell vaccines
Initially, the disease was controlled by an inactivated liver homogenate vaccine that was formulated after the first outbreak, and many researchers reported that the vaccine had successfully prevented the disease [7, 9, 25]. It was found that vaccination at 15-18 days or two vaccinations at 10 and 21 days of age gave the best protection [6, 47].
Kataria et al. [63], and Chandra et al. [22] reported that HPS can be brought under control by inactivated liver homogenate vaccines (0.25 mL/bird) or by inactivated cell culture vaccines (103.5 LD50/bird) injected subcutaneously at 10-15 days of age. Moreover, they found that the vaccine is effective in cases of outbreaks and that it significantly reduces mortality.
Khan et al. [65] and Mahmood et al. [72] suggested that outbreaks of HPS mostly occur post-vaccination in Pakistan. For this reason, the local vaccine production has been minimized. The poultry industry is still at the same risk as it was in 1987. It was suggested that the virus should be propagated on specific-pathogen-free embryonated hen eggs and cell cultures to produce killed and live attenuated vaccines. They also suggested that the unhealthy liver homogenate vaccine should not be used.
Kim et al. [67] showed that cell-culture-propagated inactivated oil-emulsion FAdV-4 vaccine conferred broad cross-protection against various serotypes of FAdV field isolates to vaccinated birds as well as the progeny of vaccinated breeders. It was concluded that cell-culture-propagated inactivated oil-emulsion FAdV-4 vaccine could be effective in preventing not only FAdV-4 infection but also other serotypes of FAdV. Previously, Icochea et al. [55] evaluated the efficacy of inactivated vaccines in Peru. They also reported that oil-adjuvanted cell culture IBH vaccine provided better protection when compared to the autogenous vaccine.
Oral live attenuated vaccine
Mansoor et al. [74] developed a live attenuated vaccine against HPS. They adapted a field isolate of HPS virus to chicken embryonated eggs by subjecting it to four blind passages. The chicken-embryo-adapted virus was further passaged up to 12 times for its complete attenuation. They tested this attenuated virus by giving it through the oral and parenteral routes to broilers at 14 days of age. They compared this attenuated vaccine with the old liver homogenate vaccines. It was found that the antibody response measured by ELISA was significantly higher in the group of broilers that were immunized with 16th-passage attenuated HPS virus at 7, 14 and 21 days post-immunization. In a challenge protection test at 24 days post-immunization, it was found that broilers immunized with 16th-passage attenuated virus were conferred with 95% protection, while the broilers immunized with the liver homogenate vaccine showed significantly lower protection (55%). Thus, the attenuated vaccine was more immunogenic and effective against HPS virus in broilers.
Recombinant (subunit) vaccine
Currently, inactivated vaccines [12] and attenuated vaccines [74] are available against HPS. The lack of full attenuation or inactivation and the oncogenic potential of adenoviruses have prevented their use in routine vaccines. A subunit vaccine has the advantage that it eliminates the danger of disease outbreaks caused by incomplete inactivation or attenuation [36].
The effectiveness of subunit vaccines against infectious bursal disease (IBD) virus in chickens [93], egg drop syndrome (EDS) virus in chickens [36], hemorrhagic enteritis virus (HEV) in turkeys [94], and hepatitis B virus in humans [81] have been demonstrated previously. Balamurugan and Kataria [16] and Khan et al. [65] suggested that modern practices of recombinant DNA technology should be applied to meet the needs of the present era for diagnosis and prevention of HPS. Pitcovski et al. [94] also suggested that subunit vaccines might be useful against adenoviruses. Shah et al. [115] developed a subunit vaccine against HPS. For this purpose, the penton base protein of HPS virus was cloned, expressed in Escherichia coli and used as subunit vaccine in broilers. The immunogenicity of the recombinant penton base protein and a challenge protection test against pathogenic virus demonstrated the ability of the recombinant penton base protein to confer 90% protection. The results suggest that the recombinant penton base protein is a candidate for a subunit vaccine against HPS. Schachner et al. [113] compared the immunogenicity of two viral recombinant fiber proteins (fiber-1 and fiber-2). The recombinant fiber-2 was identified as a protective immunogen, so it was proposed as a candidate for a subunit vaccine. A challenge protection test revealed that the fiber-2-vaccinated group was protected against challenge. However, in spite of this protection, fecal excretion of viral DNA was detected by real-time PCR, demonstrating that the vaccine had prevented clinical symptoms, but not infection.
Conclusions
HPS has been affecting poultry throughout the world since 1987. In most cases, FAdV-4 has been reported with some genetic differences. However, some researchers have found FAdV-4 in association with FAdV-10 or FAdV-12 to be the cause of HPS. Recently, researchers have also found some genetic changes in the viral genome, and it is suspected that the virulence of FAdV-4 has been increased over the years. Moreover, the worldwide scarcity of vaccines has made the situation alarming. However, it has been reported that cell-culture-propagated inactivated oil-emulsion FAdV-4 vaccine gives better protection than other available vaccines. Therefore there is a need to develop effective vaccines against this disease using modern molecular techniques. Recently, the effectiveness of subunit vaccines has been demonstrated, and subunit vaccine candidates against HPS have been identified, including recombinant viral surface proteins, e.g., penton base and fiber-2 (short fiber). Biotechnological approaches may be used for further understanding of FAdV evolution, which will provide essential information for the development of vaccines against HPS.
References
Abdul-Aziz TA, Al-Attar MA (1991) New syndrome in Iraqi chicks. Vet Record 129(12):272
Abdul-Aziz TA, Hassan SY (1995) Hydropericardium syndrome in broiler chickens: its contagious nature and pathology. Res Vet Sci 59:219–221
Abe T, Nakamura K, Tojo H, Mase M, Shibahara T, Yamaguchi S, Yuasa N (1998) Histology, immunohistochemistry and ultra structure of hydropericardium syndrome in adult broiler breeders and broiler chicks. Avian Dis 42:606–612
Afzal M, Ahmad I (1990) Efficacy of an inactivated vaccine against hydropericardium syndrome in broilers. Vet Record 126:59–60
Ahmad I, Afzal M, Malik MI, Hussain Z, Hanif W (1989) Studies on the disease pattern and etiology of hydropericardium syndrome (Angara disease) in broiler chickens in Pakistan. Pak J Agric Res 10:195–199
Ahmad I, Malik MI, Iqbal K, Ahmad K, Naz S (1990) Efficacy of formalized liver organ vaccine against Angara disease in broilers. Veterinarski Arkhiv 60:131–138
Ahmad K, Hasan S (2004) The efficacy of experimental Angara disease vaccines. Pak Vet J 24:101–103
Ahmad MD, Zaman S, Mushtaq MH, Anjum AA, Akram M (2011) Comparative pathogenicity of liver homogenate and cell culture propagated Hydropericardium syndrome virus in broiler birds. Pak Vet J 31(4):321–326
Akhtar M, Ahmad R, Hayat CS, Hussain I, Ashfaque M (2000) Comparative immune response of formalin inactivated and binary ethyleneimine inactivated Angara disease vaccines. Pak J Biol Sci 3:1313–1314
Akhtar, S., (1992). Studies on the rate of lateral spread of hydropericardium syndrome agent(s). In: Etiology, pathogenesis and control of hydropericardium syndrome in poultry. Board on Science and Technology for International Development (BOSTID), Washington, DC
Akhtar S (1994) Hydropericardium syndrome in broiler chickens in Pakistan. World’s Poultry Sci J 50:177–182
Anjum AD (1990) Experimental transmission of hydropericardium syndrome and protection against it in commercial broiler chickens. Avian Pathol 19:655–660
Anjum AD, Sabri MA, Iqbal Z (1989) Hydropericardium syndrome in broiler chickens in Pakistan. Vet Record 124:247–248
Asrani RK, Gupta BK, Sharma SK, Singh SP, Katoch RC (1997) Hydropericardium hepatopathy syndrome in Asian poultry. Vet Record 141:271–273
Asthana M, Chandra R, Kumar R (2013) Hydropericardium syndrome: current state and future developments. Arch Virol 158:921–931
Balamurugan V, Kataria JM (2004) The hydropericardium syndrome in poultry: a current scenario. Vet Res Commun 28:127–148
Balamurugan V, Kataria JM, Kataria RS, Verma KC, Nanthakumar T (2002) Characterization of fowl adenovirus 4 associated with hydropericardium syndrome in chicken. Comp Immunol Microbiol Infect Dis 25:139–147
Balamurugan V, Kataria JM, Tiwari AK, Verma KC, Toroghi R, Jadhao SJ (2001) Development of sandwich ELISA for the detection of fowl adenovirus 4 associated with hydropericardium syndrome in experimentally infected chickens. Acta Virol 45:95–100
Bhatti BM, Qureshi MS, Bajwa TM (1989) Hematology and clinical chemistry values in broiler chicken affected with hydropericardium syndrome prevalent in Pakistan. Veterinarski Arkhive 50:107–111
Borisov VV, Borisov AV, Gusev AA (1997) Hydropericardium syndrome in chicken in Russia. In: Proceedings of the tenth International Congress of World Veterinary Poultry Association, Budapest, p 258
Burnett RM (1985) The structure of the adenovirus capsid. II. The packing symmetry of hexon and its implications for viral architecture. J Mol Biol 185:125–143
Chandra R, Shukla SK, Kumar R (2000) The hydropericardium syndrome and inclusion body hepatitis in domestic fowl. Trop Anim Health Prod 32:99–111
Chandra R, Shukla SK, Kumar M, Garg SK (1997) Electron microscopic demonstration of an adenovirus in the hepatocytes of birds experimentally infected with hydropericardium syndrome. Vet Record 140:70–71
Cheema AH, Ahmad J, Afzal M (1989) An adenovirus infection of poultry in Pakistan. Revue Scientifique et Technique de l’ Office International des Epizooties 8:789–795
Chishti MA, Afzal M, Cheema AH (1989) Preliminary studies of the development of vaccine against the hydropericardium syndrome. Revue Scientifique et Technique de l’Office International des Epizooties 8:797–801
Choi KS, Kye SJ, Kim JY, Jeon WJ, Lee EK, Park KY, Sung HW (2012) Epidemiological investigation of outbreaks of fowl adenovirus infection in commercial chickens in Korea. Poult Sci 91:2502–2506
Cowen BS (1992) Inclusion body hepatitis anemia and hydropericardium syndromes: aetiology and control. World’s Poultry Sci J 48:247–254
Cowen BS, Lu H, Weinstock D, Castro AE (1996) Pathogenicity studies of fowl adenoviruses isolated in several regions of world. In: Proceedings of international symposium on adenovirus and reovirus infections in poultry, Rauischholzhauzen, pp 79–88
Dahiya S, Srivastava RN, Hess M, Gulati BR (2002) Fowl adenovirus serotype 4 associated with outbreaks of infectious hydropericardium in Haryana, India. Avian Dis 46:230–233
Davison AJ, Benko M, Harrach B (2003) Genetic content and evolution of adenoviruses. J Gen Virol 84(11):2895–2908
Deepak JN (1998) Studies on pathogenicity and immunosuppressive effects of fowl adenovirus serotype 4 isolated from hydropericardium syndrome in chicken, MVSc Thesis, Deemed University, IVRI, Izatnagar
Domermuth CH, Weston CR, Cowen BS, Colwell WM, Gross WB, DuBose RT (1980) Incidence and distribution of avian adenovirus group II splenomegaly of chickens. Avian Dis 24:591–594
Erny KM, Barr DA, Fahey KJ (1991) Molecular characterization of highly virulent fowl adenoviruses associated with outbreaks of inclusion body hepatitis. Avian Pathol 20:597–606
Fadly AM, Winterfield RW (1973) Isolation and some characteristics of an agent associated with inclusion body hepatitis, hemorrhages and aplastic anemia in chickens. Avian Dis 17:182–193
Fender P, Boussaid A, Mezin P, Chroboczek J (2005) Synthesis, cellular localization, and quantification of pentondodecahedron in serotype 3 adenovirus-infected cells. Virology 340:167–173
Fingerut E, Gutter B, Gallili G, Michael A, Pitcovski J (2003) A subunit vaccine against the adenovirus egg-drop syndrome using part of its fiber protein. Vaccine 21:2761–2766
Ganesh K, Suryanarayana V, Raghavan R, Gowda S (2001) Nucleotide sequence of L1 and part of P1 of hexon gene of fowl adenovirus associated with hydropericardium syndrome differs with the corresponding region of other fowl adenoviruses. Vet. Microbiol 78:1–11
Ganesh K (1998) Molecular studies on hexon gene fragment of fowl adenovirus associated with hydropericardium hepatitis syndrome. Ph.D. thesis, University of Agricultural sciences, Bangalore
Ganesh K, Raghvan R, Gowda RNS, Satyanarayana ML, Suryanarayana VVS (2002) Purification and characterization of the etiological agent of hydropericardium hepatitis syndrome from infected liver tissues of broiler chickens. Trop Anim Health Prod 34:7–17
Ganesh K, Suryanarayana VVS, Raghaven R (2002) Detection of fowl adenovirus associated with hydropericardium hepatitis syndrome by polymerase chain reaction. Vet Res Commun 26:73–80
Gowda RNS (1994) Leechi disease (hydropericardium syndrome), a mysterious and an emerging threat to poultry industry in India. Poultry Advisor 27:53–61
Gowda RNS, Satyanarayana ML (1994) Hydropericardium syndrome in poultry. Indian J Vet Pathol 18:159–161
Grgic H, Yang DH, Nagy E (2011) Pathogenicity and complete genome sequence of a fowl adenovirus serotype 8 isolate. Virus Res 156:91–97
Grimes TM (1992) Cause and control of a peracute form of inclusion body hepatitis. In: Proceedings of 40th western poultry disease conference, Sacramento, pp 42–43
Hafez HM (2011) Avian adenoviruses infection with special attention to Inclusion body hepatitis/hydropericardium syndrome and egg drop syndrome. Pak Vet J 31(2):85–92
Harrach B, Benkı M, Both GW, Brown M, Davison AJ, Echavarría M, Hess M, Jones MS, Kajon A, Lehmkuhl HD, Mautner V, Mittal SK, Wadell G (2011) In: King AMQ, Adams MJ, Carstens EB, Lefkowitz EJ (eds) Family Adenoviridae. virus taxonomy: classification and nomenclature of viruses. 9th report of the International Committee on Taxonomy of Viruses. Elsevier, San Diego, pp 95–111
Hassan NU, Afzal M, Hameed A, Khan AR (1994) Immune response to inactivated hydropericardium syndrome vaccine in broilers. Pak Vet J 14:5–10
Hassan SA (1989) Pakistan is mystified by hydropericardium syndrome. Poultry Misset 5:35–37
Henning P, Lundgren E, Carlsson M, Frykholm K, Johannisson J, Magnusson MK, Tang E, Franqueville L, Hong SS et al (2006) Adenovirus type 5 fiber knob domain has a critical role in fiber protein synthesis and encapsidation. J Gen Virol 87:3151–3160
Hess M (2000) Detection and differentiation of avian adenoviruses: a review. Avian Pathol 29:195–206
Hess M, Alain C, Rob WHR, Jadwiga C, Bernard J (1995) The Avian adenovirus Penton: two fibers and one base. J Mol Biol 252:379–385
Hess M, Raue R, Prusas C (1999) Epidemiological studies on fowl adenoviruses isolated from cases of infectious hydropericardium. Avian Pathol 28:433–439
Hong SS, Szolajska E, Schoehn G, Franqueville L, Myhre S, Lindholm L, Ruigrok RW, Boulanger P, Chroboczek J (2005) The 100Kchaperone protein from adenovirus serotype 2 (Subgroup C) assists in trimerization and nuclear localization of hexons from subgroups C and B adenoviruses. J Mol Biol 352:125–138
Hong SS, Habib NA, Franqueville L, Jensen S, Boulanger PA (2003) Identification of adenovirus (Ad) penton base neutralizing epitopes by use of sera from patients who had received conditionally replicative Ad (Addl1520) for treatment of liver tumors. J Virol 77:10366–10375
Icochea E, Alba M, Fiory L, Ramirez A (2001) Efficacia de tres vacunas inactivades contra la hepatitis a corpusculos de inclusion y syndrome de hidropericardio en el Peru. Revue Scientifique et Technique de Office International des Epizooties 115:1025–1030
Iqbal MJ, Bhatti BM, Khan KM (1994) Studies on serum enzymes in hydropericardium syndrome (Angara disease) of broilers. Pak Vet J 14:83–85
Jaffery MS (1988) A treatise on Angara disease (hydropericardium-pulmonary oedema-hepatonephritis syndrome). J Pak Vet Med Assoc 34:1–33
Jucker MT, McQuiston JR, van den Hurk JV, Boyle SM, Pierson FW (1996) Characterization of the haemorrhagic enteritis virus genome and the sequence of the putative penton base and core protein genes. J Gen Virol 77(3):469–479
Kajan GL, Kecskemeti S, Harrach B, Benko M (2013) Molecular typing of fowl adenoviruses, isolated in Hungary recently, reveals high diversity. Vet Microbiol 167:357–363
Karunamoorthy G, Manickam R (1998) Hydropericardium syndrome in quails. Poultry Times of India 2:1–3
Kataria JM, Verma KC, Jadhao SJ, Shah RL (1996) Studies on virus isolated from cases of inclusion body hepatitis in chickens. In: Proceedings of the 17th annual conference of the Indian Association of Veterinary Microbiologists, Immunologists and Specialists in Infectious Diseases, Bhubaneswar, p 24
Kataria JM, Verma KC, Jadhao SJ, Deepak JN, Shah RL (1997) Efficacy of an inactivated oil emulsified vaccine against inclusion body hepatitis hydropericardium syndrome (litchi disease) in chicken prepared from cell culture propagated fowl adenovirus. Indian J Comp Microbiol Immunol Infect Dis 18:38–42
Kataria JM, Verma KC, Shah RL, Jadhao SJ (1997a) Experimental studies with fowl adenovirus serotype 4 isolated from cases of IBH-HPS in chicks. In: Proceedings of the 18th conference of Indian Association of Veterinary Microbiologists, Immunologists and Specialists in Infectious Diseases, Ludhiana, p 69
Kawamura H, Shimizu F, Tsubahara H (1964) Avian adenovirus: its properties and serological classification. Natl Inst Anim Health Q (Tokyo) 4:183–193
Khan AA, Sabri AN, Mansoor MK, Hussain I (2005) Hydropericardium syndrome in Pakistan: a review. World’s Poultry Sci J 61:647–653
Khawaja DA, Ahmad S, Rauf MA, Zulfiqar MZ, Mahmood SMI, Hassan M (1988) Isolation of an adenovirus from hydropericardium syndrome in broiler chicks. Pak J Vet Res 1:2–17
Kim MS, Tae HL, Dong HL, Hana Y, Seong SY, Byoung YK, Soo WC, Cheong HJ, Jang HH, Chang SS (2014) An Inactivated oil-emulsion fowl adenovirus serotype 4 vaccine provides broad cross-protection against various serotypes of fowl adenovirus. Vaccine 32:3564–3568
Kumar R, Chandra R (2004) Studies on structural and immunogenic polypeptides of hydropericardium syndrome virus by SDS-PAGE and western blotting. Comp Immunol Microbiol Infect Dis 27(3):155–161
Kumar R, Chandra R, Shukla SK (2003) Isolation of etiological agent of hydropericardium syndrome in chicken embryo liver cell culture and its serological characterization. Indian J Exp Biol 41:821–826
Kumar R, Chandra R, Shukla SK, Agrawal DK, Kumar M (1997) Hydropericardium syndrome in India: a preliminary study on the causative agent and control of the disease by inactivated autogenous vaccine. Trop Anim Health Prod 29:158–164
Laver WG, Younghusband HB, Wrigley NG (1971) Purification and properties of chick embryo lethal orphan virus (an avian adenovirus). Virology 45:598–614
Mahmood MD, Khushi M, Masood R, Atif H, Irshad H (2011) In process quality control factors affecting efficacy of Hydropericardium syndrome virus vaccine. Pak J Zool 43(1):73–77
Mahmood T, Hassan Z (1995) Propagation of Angara disease virus in duck’s embryonated eggs. Pak J Livestock Poultry 1:104–105
Mansoor MK, Hussain I, Arshad M, Muhammad G (2011) Preparation and evaluation of chicken embryo-adapted fowl adenovirus serotype 4 vaccine in broiler chickens. Trop Anim Health Prod 43:331–338
Manzoor S, Hussain I (2003) Reverse passive haemagglutination (RPHA) test for the detection and quantification of hydropericardium syndrome virus. Pak J Life Soc Sci 1:141–143
Marek A, Nolte V, Schachner A, Berger E, Schlötterer C, Hess M (2012) Two fiber genes of nearly equal lengths are a common and distinctive feature of Fowl adenovirus C members. Vet Microbiol 156(3–4):411–417
Mase M, Kikuyasu N, Tadao I (2010) Characterization of fowl adenovirus serotype 4 isolated from chicken with hydropericardium syndrome based on analysis of the short fiber protein gene. J Vet Diagn Invest 22:218–223
Mase M, Nakamura K, Minami F (2012) Fowl adenoviruses isolated from chickens with inclusion body hepatitis in Japan, 2009–2010. J Vet Med Sci 74:1087–1089
Matsushima Y, Shimizu H, Phan TG, Ushijima H (2011) Genomic characterization of a novel human adenovirus type 31 recombinant in the hexon gene. J Gen Virol 92(12):2770–2775
Mazaheri A, Prusas C, Vob M, Hess M (1998) Some strains of serotype 4 fowl adenoviruses cause inclusion body hepatitis and hydropericardium syndrome in chickens. Avian Pathol 27:269–276
McAleer WJ, Eugene B, Buynak RZ, Maigetter D, Eugene W, William JM, Maurice RH (1984) Human hepatitis B vaccine from recombinant yeast. Nature 307:178–180
Mcferran JB, Adair B, Connor TJ (1975) Adenoviral antigens (CELO, QBV, GAL). Am J Vet Res 36:527–529
Mcferran JB, Connor TJ, Adair BM (1978) Studies on the antigenic relationship between an isolate (127) from the egg drop syndrome 1976 and a fowl adenovirus. Avian Pathol 7:629–639
Merck Sharp, Dohme Corp. (2011) Inclusion body hepatitis/hydropericardium syndrome: introduction. The Merck veterinary manual, 10th edn. A subsidiary of Merck & Co., Inc., Whitehouse Station. (http://www.merckvetmanual.com/mvm/index.jsp?cfile=htm/bc/200400.htm)
Mittal D, Jindal N, Tiwari AK, Khokhar RS (2014) Characterization of fowl adenoviruses associated with hydropericardium syndrome and inclusion body hepatitis in broiler chickens. Virus Dis 25:114–119
Naeem K, Akram HS (1995) Hydropericardium outbreaks in pigeon flock. Vet Record 138:296–297
Naeem K, Rahim K, Majeed IU (2001) Post infection dissemination pattern of avian adenovirus involved in hydropericardium syndrome. Pak Vet J 21:152–156
Nakamura K, Mase M, Yamaguchi S, Yuasa N (2000) Induction of hydropericardium in one day old specific pathogen free chicks by adenovirus from inclusion body hepatitis. Avian Dis 44:192–196
Niazi AK, Khan MZ, Siddique M (1989) Haematological studies on naturally occurring hydropericardium syndrome in broiler chicks. Vet Record 125:400
Nicklin SA, Wu E, Nemerow GR, Baker AH (2005) The influence of adenovirus fiber structure and function on vector development for gene therapy. Mol Ther 12:384–393
Niczyporuk JS (2016) Phylogenetic and geographic analysis of fowl adenovirus field strains isolated from poultry in Poland. Arch Virol 161:33–42
Pehler-Harrington K, Khanna M, Waters CR, Henrickson KJ (2004) Rapid detection and identification of human adenovirus species by adenoplex, a multiplex PCR–enzyme hybridization assay. J Clin Microbiol 42:4072–4076
Pitcovski J, Bezalel G, Gilad G, Martin G, Beny P, Gideon G, Simha K, Marisa B, Amnon M (2003) Development and large-scale use of recombinant VP2 vaccine for the prevention of infectious bursal disease of chickens. Vaccine 23:4736–4743
Pitcovski J, Fingerut E, Gallili G, Eliahu D, Finger A, Gutter B (2005) A subunit vaccine against hemorrhagic enteritis adenovirus. Vaccine 38:4697–4702
Qureshi AA (1988) Hydropericardium and kidney lesions. Poultry Int 27:48–50
Qureshi AA (1989) Hydropericardium and ascites. Poultry Int 28:44–48
Rabbani M, Naeem K (1996) In vitro and in vivo evaluation of avian adenovirus isolates from outbreaks of hydropericardium syndrome. In: Proceedings of international symposium on adenovirus and reovirus infections in poultry, Rauischholzhauzen, pp 26–31
Rabbani M, Muneer MA, Naeem K (1998b) A dot immunobinding assay on nitrocellulose for the detection of antigens and antibodies of avian PARC 1. In: Proceedings of the second pan commonwealth veterinary conference, pp 1325–1331
Rahul S, Kataria JM, Senthilkumar N, Dhama K, Sylvester SA, Uma R (2005) Association of fowl adenovirus serotype 12 with hydropericardium syndrome of poultry in India. Acta Virol 49(2):139–143
Rau R, Hess M (1998) Hexon based PCRs combined with restriction enzyme analysis for rapid detection and differentiation of fowl adenoviruses and egg drop syndrome virus. J Virol Methods 73:211–217
Reece RL, Barr DA, Grix DC (1987) Pathogenicity studies with a strain of fowl adenovirus serotype 8 (VRI-33) in chickens. Aust Vet J 64:365–367
Rehman SU, Ashfaque M, Anjum AD, Sandhu TA (1989) Indirect haemagglutination test for detecting Angara disease (hydropericardium) agent antibody. In: Proceedings of the international conference and trade show on poultry production, pp 73–74
Roberts DM, Nanda A, Havenga MJ, Abbink P, Lynch DM, Ewald BA, Liu J, Thorner AR, Swanson PE et al (2006) Hexon-chimaeric adenovirus serotype 5 vectors circumvent pre-existing anti-vector immunity. Nature 441:239–243
Robinson CM, Singh G, Henquell C, Walsh MP, Peigue-Lafeuille H, Seto D, Jones MS, Dyer DW, Chodosh J (2011) Computational analysis and identification of an emergent human adenovirus pathogen implicated in a respiratory fatality. Virology 409:141–147
Rosenberger JK, Eckroade RJ, Klopps S, Krauss WC (1974) Characterization of several viruses isolated from chickens with inclusion body hepatitis and aplastic anemia. Avian Dis 18:399–409
Roy P, Kotteeswaran A, Manickam R (2001) Serological, cytopathological and cytochemical studies on hydropericardium syndrome virus. Veterinarski Arhiv 71(2):97–103
Roy P, Muralimanohar B, Kotteeswaran A, Omparkash AV (2004) Experimental studies on hydropericardium syndrome in two different synthetic lines of broiler chickens. Veterinarski Arhiv 74:157–164
Russel WC (2000) Update on adenovirus and its vectors. J Gen Virol 81:2573–2604
Russel WC (2009) Adenoviruses: update on structure and function. J Gen Virol 90:1–20
Rux JJ, Kuser PR, Burnett RM (2003) Structural and phylogenetic analysis of adenovirus hexons by use of high-resolution X-ray crystallographic, molecular modeling, and sequence-based methods. J Virol 77:9553–9566
Saban SD, Silvestry M, Nemerow GR, Stewart PL (2006) Visualization of a-helices in a 6-A° ngstrom resolution cryoelectron microscopy structure of adenovirus allows refinement of capsid protein assignments. J Virol 80:12049–12059
Saifuddin M, Wilks CR (1990) Development of an enzyme linked immunosorbent assay to detect and quantify adenovirus in chicken tissue. Avian Dis 34:239–245
Schachner A, Marek A, Jaskulska B, Bilic I, Hess M (2014) Recombinant FADV-4 fiber-2 protein protects chickens against hepatitis-hydropericardium syndrome (HHS). Vaccine 32(9):1086–1092
Shah MS, Ashraf A, Khan MI, Rahman M, Habib M, Babapoor S (2011) Molecular characterization of fowl adenoviruses associated with hydropericardium syndrome in broilers. Afr J Microbiol Res 30:5407–5414
Shah MS, Ashraf A, Khan MI, Rahman M, Qureshi JA (2012) A subunit vaccine against hydropericardium syndrome using adenovirus penton capsid protein. Vaccine 30(50):7153–7156
Shah MS, Ashraf A, Khan MI, Rahman M, Habib M, Qureshi JA (2016) Molecular cloning, expression and characterization of 100K gene of fowl adenovirus-4 for prevention and control of hydropericardium syndrome. Biological 44:19–23
Shane SM (1996) Hydropericardium hepatitis syndrome, the current world situation. Zootecnica Int 18:20–27
Talha K, Moid S, Ahmad M, Hassaan H, Moazzam A (2011) Poultry industry; API presentation. www.slideshare.net/msaadafridi/poultry-industry-of-pakistan
Tan PK, Michou AI, Bergelson JM, Cotton M (2001) Defining CAR as a cellular receptor for avian adenovirus CELO using a genomic analysis of the two viral fiber proteins. J Gen Virol 82:1465–1472
Thakor KB, Dave CJ, Prajapati KS, Fefar DT, Jivani BM (2012) Molecular characterization of Avian adenovirus causing Inclusion body hepatitis—hydropericardium syndrome in broiler chicken of Anand, Gujarat, India. Vet World 5(3):178–182
Toro H, Gonzalez C, Cerda L, Hess M, Reyes F, Geisse C (2000) Chicken anemia virus and fowl adenoviruses: association to induce the inclusion body hepatitis/hydropericardium syndrome. Avian Dis 44:51–58
Toro H, Gonzalez O, Escobar C, Cerda L, Morales MA, Gonzalez C (2001) Vertical induction of inclusion body hepatitis/hydropericardium syndrome with fowl adenovirus and chicken anemia virus. Avian Dis 45:215–222
Toro H, Prusas C, Raue R, Cerda L, Geisse C, Gonzalez C, Hess M (1999) Characterization of fowl adenovirus from outbreaks of inclusion body hepatitis/hydropericardium syndrome in Chile. Avian Dis 43:262–270
Viralzone (2015). dsDNA viruses, adenoviridae, aviadenovirus. Swiss institute of bioinformatics. htp://www.viralzone.expasy.org
Voss M, Vieltz E, Hess M, Prusas CH, Mazaheri A (1996) Etiological aspects of hepatitis and hydropericardium syndrome caused by pathogenic adenoviruses in different countries. In: Proceedings of the international symposium on adenovirus and reovirus infections in poultry, Rauischholzhauzen, pp 75–78
Wiethoff CM, Wodrich H, Gerace L, Nemerow GR (2005) Adenovirus protein VI mediates membrane disruption following capsid disassembly. J Virol 79:1992–2000
Xie Z, Luo S, Fan Q, Xie L, Liu J, Xie Z, Pang Y, Deng X, Wang X (2013) Detection of antibodies specific to the non-structural proteins of fowl adenoviruses in infected chickens but not in vaccinated chickens. Avian Pathol 42(5):491–496
Zaman T, Khan MZ (1991) Serum enzyme profiles in hydropericardium affected broiler chicks. Pak Vet J 11:50–52
Zhang T, Qianyue J, Peiyang D, Yinbiao W, Yongxiao C, Yafei L, Xiao L, Jun L, Zhang G (2016) Molecular epidemiology of Hydropericardium syndrome outbreaks-associated serptype 4 fowl adenovirus isolates in central China. Virol J 13:188
Zubieta C, Schoehn G, Chroboczek J, Cusack S (2005) The structure of the human adenovirus 2 penton. Mol Cell 17:121–135
Acknowledgements
We are thankful to the Higher Education Commission (HEC), Government of Pakistan, for their support to accomplish this work.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
This article does not contain any studies with human participants or animals performed by any of the authors.
Conflict of interest
The authors of this manuscript have declared that no conflict of interest exists.
Rights and permissions
About this article

Cite this article
Shah, M.S., Ashraf, A., Khan, M.I. et al. Fowl adenovirus: history, emergence, biology and development of a vaccine against hydropericardium syndrome. Arch Virol 162, 1833–1843 (2017). https://doi.org/10.1007/s00705-017-3313-5
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00705-017-3313-5