
Abstract
The iron storage proteins, ferritin and hemosiderin, enable electron microscopic visualization thanks to their electron-dense iron content, which is not present in other compounds involved in transport or metabolism of iron such as transferrin, lactoferrin, or hemoglobin. It is this electron density which contributed to the unraveling of stages in absorption, transport, deposition, storage, and release of iron. In recent years, additional methods of investigation have further supported the information achieved by the ultrastructural studies. Even while using new analytical methods, the seminal morphological observations remain valid for understanding the role of iron in health and disease. In this review, we will illustrate a few basic findings of electron microscopy in humans, experimental animals, and cell cultures. The importance of H chain ferritin as a transporter across the blood–brain barrier is just an example of a new role revealed for an “old” storage protein, explaining some controversial observations on the presence of iron in the brain.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
In December 1976, Ciba Foundation, London convened a group of scientists interested in “Iron Metabolism” as an initiative of the late Alan Jacobs, Editor of the British Journal of Haematology. Intentionally, the group was small, but the participants were each specialized in different aspects of iron, either in basic research or clinical aspects. As it happened, the small group had ample time to exchange views resulting in productive information for all. My presence there was the result of an encounter, a few months before, with Alan Jacobs during his visit to the Children’s Hospital of Los Angeles; I was involved in a thorough electron microscopic study of iron overload in thalassemic children. Alan was enthusiastic of my findings on the ultrastructural features of ferritin and hemosiderin and deposition in various organs, from an early stage to advanced siderosis. He considered these findings important enough to be presented at the forthcoming Ciba London meeting (Ciba Foundation Symposium 51: Symposium on Iron Metabolism). Indeed, the use of unstained ultrathin sections enabled identification, location, and relative quantification of ferritin and hemosiderin during various stages of iron-loading conditions. Subsequently, stimulated by the interest shown by the colleagues in the group, I began expanding the study to morphologic features of iron overload in (1) spontaneous iron accumulation in humans and animals, (2) iron-loading diseases, (3) experimental models of iron overload, (4) iron in tumors, (5) iron in cultured cells, and (6) the effect of iron chelators on various models. Most of these studies were supplemented by a variety of complementary data in support of the electron microscopic findings. Even today, after 35 years in iron ultrastructure research, I still find the intra- and extracellular electron-dense iron compounds as fascinating markers of metal presence.
M. B. H. Youdim was among the distinguished group of iron researchers present at the Ciba Foundation Symposium in 1976. He presented a paper entitled “Biogenic monoamine metabolism and functional activity in iron deficient rats: behavioral correlates” (together with A. R. Green). We thus learned the other aspect of iron metabolism, the effects of deficiency, worldwide more important than the iron-loading conditions. As a pediatrician, I was naturally attracted by this aspect and followed Youdim’s studies on iron, behavior, and development with great interest. It came as a surprise when Moussa informed me of his appointment at the Technion Faculty of Medicine as Professor of Pharmacology. Neighbors in the same building, we continued our common interest in various aspects of iron metabolism ever since, recently focused on iron chelators.
The present review is intended to show some ultrastructural aspects of iron-containing compounds, mainly ferritin and hemosiderin, in an attempt to correlate these morphological findings with clinical, biochemical, and other analytical findings. They may help us in visualizing the distribution of iron compounds in different cellular and sub-cellular compartments. The various chemical, biological, histopathological, and electron microscopic methods have been described in detail in recent reviews.
Ferritin
The definition of this archaeoprotein varies according to those interested in its study. It is different for biochemists, biologists, clinicians, physicists, and electron microscopists to name only a few groups of researchers. Notwithstanding the variable focus, it can be stated that ferritin “is a ubiquitous intracellular protein that stores iron and releases it in a controlled fashion”. The protein is produced by almost all living organisms, including bacteria, algae and higher plants, and animals. In humans, it acts as a buffer against iron deficiency and iron overload. It is a globular protein complex consisting of 24 protein subunits and is the primary intracellular iron-storage protein in both prokaryotes and eukaryotes, keeping iron in a soluble and non-toxic form. Ferritin that is not combined with iron is called apoferritin. Apoferritin is a polymer composed of variable proportions of two units (H heavy, or Heart m.w. 21 kDa and L light or liver m.w. 19 kDa).
Within the central cavity of the hollow apoferritin molecules variable amounts of iron can be accommodated. While natural apoferritin contains only traces of iron, a fully loaded molecule of ferritin contains up to 4,500 atoms of iron (Fe III). The core is variably described as “iron-hydroxy-phosphate”, “iron-oxyhydroxide” or “hydrous ferric-oxide-phosphate” or abbreviated (FeOOH) x . It is not yet clear whether the core consists of a single crystallite, as seen directly by electron microscopy, or of a number of crystallites (Harrison and Arosio 1996; Clegg et al. 1980).
The ferritin molecule: electron microscopy
The findings of iron subunits within the core are now considered artifacts since in fact the iron core is homogenous (Bessis 1973). In unstained biological material, only the electron-opaque cores are seen, and they vary in size and shape according to their iron content, i.e. “iron-poor”, “iron-rich”, and “intermediate iron content” (Iancu 1983, 1992). In iron-overloaded tissues, unstained sections show the cores with a diameter of about 6.0 nm (5.3–6.5 nm). The size and shape of the cores can be influenced by their state of hydration, the focus of the electron micrograph, and the degree of tilting. In solution, fully iron loaded and hydrated molecules appear larger (7.0–7.5 nm; Fishbach et al. 1971). By negative staining (holo) ferritin molecules measure 10.5–11.0 nm (Harrison et al. 1980).
The size and shape of individual ferritin particles is now considered typical enough for their identification in high-resolution micrographs, but disagreement arises when iron-containing particles join forming clusters or other arrangements in which the particles are densely packed (Harrison et al. 1974). Within lysosomes, the ratio of iron/protein increases, implying degradation of the apoferritin shell. According to Fishbach et al. (1971), ferritin molecules must be separated by at least 5.0 nm, representing twice the 2.5 nm apoferritin coat, in order to be considered undegraded ferritin molecules (Fig. 1). The electron microscopic study of unstained sections from iron-overloaded tissues produced the following conclusions:

The putative pathway of iron segregation in lysosomes (siderosomes) shows ferritin (holoferritin) with an electron-dense core being taken up by a single membrane-bound body. Within the siderosomes, the particles may become associated with phospholipid membranes forming arrays, may form clusters or, after degradation of the protein coat, may form an extremely electron-opaque mass, considered to be hemosiderin (Richter 1958, 1980; Richter and Bessis 1965)
-
1.
Iron-poor and iron-rich ferritin cores can be identified in most tissues, after spontaneous or experimental overload (Figs. 2, 3, 4, 5, 6, 7, 8, 9).
Fig. 2 
Synthesis of apoferritin by bound polyribosomes and its progression along the pathway for secretory protein. It has been suggested that iron may permeate apoferritin at a later stage in or around the Golgi complex. However, the direct formation of an iron-containing secondary lysosome, possibly by autophagy, from iron present in the cell sap, is more widely accepted (Hernandez-Yago et al. 1980)
Fig. 3 
Uptake of ferritin clusters into lysosomes has been suggested as an additional mechanism: After cytosolic ferritin particles coalesce and form clusters, they may join a “digestive vacuole” (primary or secondary lysosome) resulting in a membrane-bound body. The process of segregation in vacuoles is considered now as autophagocytosis, and the resultant organelles are described as autophagosomes. These may contain all forms of storage iron: individual ferritin particles, hexagonal arrangements, and hemosiderin “clumps” of variable electron density
Fig. 4 
Electron microscopic illustration of the above stages: The enterocyte of a carbonyl-iron-fed rat shows (after 4 months) a membrane-bound body which contains iron-rich cores of ferritin (unstained)
Fig. 5 
After 6 months of iron-supplemented feeds, iron-rich ferritin particles are seen in relationship with a cytosolic droplet (thin arrow) while hexagonal arrangements of ferritin are also present adjacent to plasma membrane (broad arrow). There is no preferential location of ferritin on or along the plasma membrane. Occasionally, siderosomes appear “trapped” in between plasma membrane of two adjoining cells. Hexagonal arrangements of ferritin cores are always in membrane-bound bodies, i.e. siderosomes (unstained)
Fig. 6 
15 months of carbonyl-iron-containing diet produces a further increase in the cytosiderosis: hexagonal paracrystalline arrangements of ferritin are frequent in liver cells (unstained)
Fig. 7 
At this stage of overload, massive presence of individual ferritin particles is noticed in cytosol in addition to numerous siderosomes with hexagonal ferritin arrangements. Macrophage (Kupffer cell) (unstained)
Fig. 8 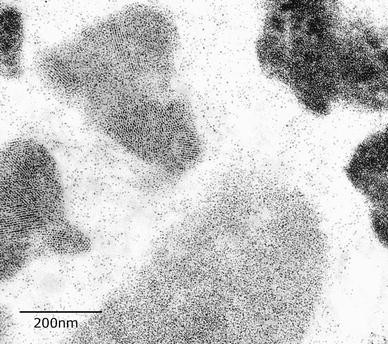
This macrophage illustrates variable densities of ferritin particles. In some bodies, individual particles can still be resolved, but in the hemosiderin-containing siderosomes the apoferritin coat has disappeared and the siderosome density is increased (unstained)
Fig. 9 
This micrograph shows various forms of iron overload present in the same field: cell sap individual particles, paracrystalline ferritin arrangement (arrow), and a hemosiderin electron-dense mass. Liver, 15 months of carbonyl-iron feeding (unstained)
-
2.
Most iron-poor cores are randomly dispersed within the cell sap.
-
3.
Most iron-rich cores are seen in clusters or, more frequently, in membrane-bound bodies (lysosomes—siderosomes) (Figs. 1, 4, 5).
-
4.
Most ferritin cores seen in paracrystalline arrangements or in relationship with phospholipidic membranes and forming arrays are of the iron-rich variety (Figs. 3, 7, 8).
-
5.
Electron-dense particles which are smaller and/or more densely packed than ferritin cores are seen only in membrane-bound bodies (i.e. hemosiderin within siderosomes) (Figs. 1, 4, 5).
-
6.
Hemosiderin particles do not form symmetrical arrangements but tend to coalesce and for clumps (Figs. 4, 7, 8).








Hemosiderin
In contrast to ferritin, this iron storage compound is heterogeneous to such an extent as to make biochemical and ultrastructural characterization extremely difficult. It was first recognized in morphological studies as a brown pigment-forming intra- and extracellular granules which gave a positive Prussian Blue (PB) Perls’ stain reaction. Attempts to isolate hemosiderin were based on some other properties: insolubility in water, heat coagulability, and isolation of heavy hemosiderin granules by centrifugation (Vidnes and Helgeland 1973). It is generally agreed that the iron is hydrous ferric oxide and is responsible for the brown color of ‘rusted’ iron-overloaded tissues, as well as for the positivity of Perls’ PB reaction. Degraded ferritin molecules may provide most of the non-heme iron present in hemosiderin of endogenous nature, with lipid, heme, and carbohydrate as additional components of this heterogeneous conglomerate, which is mostly located in secondary lysosomes.
Electron microscopy of hemosiderin
The earliest electron microscopic studies of hemosiderin granules revealed its heterogeneous nature, as well as the presence within these membrane-bound bodies of typical ferritin particles, along with other electron-dense particles and pleiomorphic osmophilic material. Richter and Bessis (1965) described hemosiderin granules in five different forms: (1) aggregates of ferritin molecules, often surrounded by a membrane; (2) characteristic ferritin crystals, which may or may not be situated on the interior of a membrane; (3) aggregates of dense particles, measuring 0.7–3.0 nm, which probably correspond to micelles contained in ferritin molecules; (4) a mixture of 1, 2, and 3; (5) complex bodies which contain a mixture of osmiophilic material (e.g. lipofuscin), myelin figures and other membranes, as well as glycoproteins. Being enclosed by a membrane, the bodies (granules) were considered to be secondary lysosomes and in view of their iron content, named siderosomes. According to Zuyderhoudt et al. (1978), the non-ferritin depot iron (hemosiderin) results from the difference between total liver depot iron and total ferritin iron.
Electron microscopically, we consider hemosiderin to be formed of extremely variable electron-dense particles, from the smallest visible particles to long specular crystals (Richter 1978) all being different presentations of hydrous ferric oxide. The extent to which these particles coexist with other components, such as carbohydrates, lipids, and proteins within secondary lysosomes, depends, in our opinion, on the cell type and the degree of iron overload. Of note, hemosiderin granules have an increased iron/protein ration, in parallel with their increased frequency of these granules along the progressive process of overload (Zuyderhoudt et al. 1978).
Basic ultrastructural features of iron storing proteins
The iron-loaded liver of humans or experimental animals provided the easiest information on the features, amount, and location of ferritin and hemosiderin. It is beyond the scope of this review to describe these classical findings (Arstila et al. 1970). Suffice to recall (1) the presence of randomly dispersed individual ferritin molecules in the cell sap, (2) their occasional assembly in clusters, and (3) coalescence in membrane-bound bodies in lysosomes (siderosomes), probably through autophagy. The explanation of the putative pathway of their formation is graphically depicted in Figs. 1, 2, and 3. Figures 4, 5, 6, 7, 8, and 9 show, in previously unpublished electron micrographs, the stages of cytosiderosis in an experimental model: the carbonyl-iron-fed mouse (Iancu et al. 1987a, b).
In parallel to the studies on spontaneous iron overload in humans with iron-loading diseases (Iancu et al. 1977), the investigations of spontaneous and experimental iron overload in experimental animals have provided additional information on these conditions. More recently, we used cell cultures to study the effect of iron supplements and that of chelating agents (Iancu et al. 1987a, b; Cooper et al. 1988; Hirsh et al. 2002). Because of the link between iron and neoplasia, tumors (e.g. neuroblastoma) were also examined to understand their iron segregation (Iancu 1989a, b). The association of ferritin and hemosiderin with various lipidic compounds (e.g. lipofuscin, melanin) is under ongoing study.
In recent years, the studies on brain iron have multiplied, including humans and experimental animals. These investigations were partly stimulated by the new information on the importance of iron in such conditions as Parkinson’s disease (Takanashi et al. 2001), Alzheimer’s (Mandel et al. 2007), as well as rarer conditions, e.g. NBIA1—former Hallervorden-Spatz disease; the term Neurodegeneration with Brain Ion Accumulation type1 (NBIA-1) has been used in more recent publications, and the most recently used term for this condition is pantothenate kinase-associated neurodegeneration (Gregory et al. 2009).
In addition to the role of iron deposition in basal ganglia, the importance of iron deficiency in behavioral disorders came into focus (Youdim et al. 1982). In a different area of research, the finding of numerous iron-containing neurones throughout the caudate putamen was consistent with the supposition that iron is directly involved with dopamine neurotransmission at the neuronal level (Ben-Shachar et al. 1985).
In my early years of study of iron-loading conditions, I was impressed by the brown color of the meninges of children undergoing post-mortem examination after lethal ß-thalassemia major. However, surprisingly, only vascular endothelium and some macrophages would show a positive Perls’ stain. The brain sections, at all levels, were essentially negative for iron. Years later, I had the opportunity (thanks to Dr. Joanna Hill 1985, 1988) to study ultrathin sections (glutaraldehyde-fixed and epoxy-embedded) of globus pallidus, substantia nigra (pars reticulata) nucleus accumbens, and caudate putamen. The sections were from adult patients and were unstained. Here again, we were unable to identify electron-opaque iron particles, with the exception of a few hemosiderin granules in cells of the capillary endothelium.
These findings were in striking discrepancy with the results of others attempting identification of brain iron, such as histocytochemistry, atomic absorption spectrophotometric analysis and above all, Perls’ reaction for ferric iron intensified by diaminobenzidine (Perls’ + DAB). The application of the latter methodology by Hill (1985) showed amounts of stainable iron in areas previously unknown, especially in the rat’s brain. It thus came to mind that the routine processing of brain tissue is not adequate to reveal its iron presence, especially as ferritin. Indeed, in contrast to the methodology used by Hill, it was (and still is) common procedure to immerse the whole brain for 10–12 days in 10% buffered formaldehyde, thus enabling its further sectioning without distortion. However, it is understandable that during such a procedure, part of the core-iron of the ferritin molecule is “washed out”. Furthermore, the ensuing processing for electron microscopy will further diminish the size or electron density of the iron-oxyhydroxide core. The remaining ferritin protein can be demonstrated, among other methods, by immunochemistry, assuming that the protein coat has not been degraded and has not lost its epitopes.
Other methods for identification of brain iron were reviewed by Perl and Good (1992) with special emphasis on laser microprobe mass analysis (LAMMA). This methodology produces a mass spectrum with simultaneous identification of multiple elements through the entire periodic table. In their review of this instrument, Perl and Good showed mass spectra from neuromelanin granules with high peaks of aluminum and iron in granules of a patient with Parkinson’s disease. Using energy-dispersive X-ray spectroscopy (EDS or EDX), Jellinger et al. (2006) demonstrated a neuromelanin-iron complex in dopaminergic substantia nigra pars compacta neurons in Parkinson’s disease. Another analytical method, electron energy loss spectroscopy (EELS) indentifies spectra of elements enabling quantification and is more sensitive than EDS. Among complementary methods permitting identification and quantification of iron, the PRIME magnetic resonance sequence (Graham et al. 2000) should be mentioned as it identified higher levels of iron in substantia nigra of patients with Parkinson’s disease.
Iron transport into the brain
The fact that the brain contains large amounts of iron, about a third probably as ferritin, is presently supported by numerous observations. Less clear is the pathway by which the iron reaches the brain tissue (Rouault 2001). The fact that ferritin was found in all organs in hypotransferrinemic mice indicated that transferrin is not the major transporter of iron into the brain. Fisher et al. (2007) were able to demonstrate through cell culture model of the blood–brain barrier that H-ferritin iron is transported across endothelial cells in the brain. These authors conclude that the relative amount of iron that could be delivered via H-ferritin could make this protein a predominant player in cellular iron delivery.
As for the transport of inorganic iron into the brain, a unitary concept is only beginning to emerge. The more recent reports by Moos et al. (2007) on iron trafficking inside the brain and by Rouault et al. (2009) on the putative role of the choroid plexus may unravel the mechanism of iron route into the brain. The latest comprehensive description of putative mechanisms for iron transport into the brain has been provided by Crichton (2009).
Conclusions
As this review is mainly concerned with ultrastructural aspects of iron, suffice to mention other methods in use for demonstration and quantification of iron in various locations, including neurological tissues. Immunocytohistiological staining, as well as the use of gold particles as markers, have been frequently used (Cooper et al. 1988; Hirsh et al. 2002). LAMMA (Iancu et al. 1995, 1996) has the advantage of the localization of the iron-containing organelles and determination of the metal amount.
Requiring less expensive instruments, immunostaining using specific anti-ferritin antibodies (polyclonal, Sigma, Israel) at the light microscopic level provide information on the presence of (holo) ferritin in the cells and their changes when exposed to chelating agents (Y. Bashenko et al., ongoing study). These investigations may establish a parallel between the ultrastructural appearance of the assembled iron-rich molecules and the presence of ferritin-protein alone.
References
Arstila AU, Bradford WD, Kinney TD, Trump BF (1970) Iron metabolism and cell membranes. Am J Path 53:419–449
Ben-Shachar D, Finberg JP, Youdim MBH (1985) Effect of iron chelators on dopamine D2 receptors. J Neurochem 45:995–1005
Bessis M (1973) Living blood cells and their ultrastructure. Springer, Berlin
Clegg GA, Fitton JE, Harrison PM, Trefry A (1980) Ferritin: molecular structure and iron-storage mechanisms. Prog Biophys Mol Biol 36:56–86
Cooper PJ, Iancu TC, Ward RJ, Guttridge KM, Peter TJ (1988) Quantitative analysis of immunogold labeling for ferritin in liver from control and iron-overloaded rats. Histochem J 20:499–509
Crichton R (2009) Iron metabolism—from molecular mechanisms to clinical consequences, 3rd edn. Wiley, New York
Fishbach FA, Gregory DW, Harrison PM, Hoy TG, Williams JM (1971) On the structure of hemosiderin and its relationship to ferritin. J Ultrastruct Res 37:495–503
Fisher J, Devraj K, Ingram J, Slagle-Webb B, Madhankumar AB, Liu X, Klinger M (2007) Ferritin: a novel mechanism for delivery of iron to the brain and other organs. Am J Physiol Cell Physiol 293:C641–C649
Graham JM, Paley MNJ, Grünewald RA1, Hoggard N, Griffiths PD (2000) Brain iron deposition in Parkinson’s disease imaged using the PRIME magnetic resonance sequence. Brain 123:2423–2431
Gregory A, Polster BJ, Hayflick SJ (2009) Clinical and genetic delineation of neurodegeneration with brain iron accumulation. J Med Genet 46:73–80
Harrison PM, Arosio P (1996) The ferritins: molecular proprieties, iron storage function and cellular regulation. Biochim Biophys Acta 1275:161–203
Harrison PM, Hoy TG, Macara IJ, Hoare RJ (1974) Ferritin iron uptake and release. Structure-function relationships. Biochem J 143:445–451
Harrison PM, Clegg GA, May K (1980) Ferritin structure and function. In: Jacobs A, Worwood M (eds) Iron in biochemistry and medicine. Academic Press, London, pp 131–171
Hernández-Yago J, Knecht A, Martinez-Ramon, Grisolia S (1980) Autophagy of ferritin incorporated into the cytosol of Hela cells by liposomes. Cell Tissue Res 205:303–309
Hill JM (1985) Iron concentration reduced in ventral pallidum, globus pallidus, substantia nigra by GABA-transaminase inhibitor, gamma-vinyl GABA. Brain Res 342:18–25
Hill JM (1988) The distribution of iron in the brain. In: Youdim MBH (ed) Brain iron: neurochemical and behavioral aspects. Taylor and Francis, London, pp 1–24
Hirsh M, Konijn AM, Iancu TC (2002) Acquisition, storage and release of iron by human cultured hepatoma cells. J Hepatol 36:30–38
Iancu TC (1983) Iron overload. Mol Aspect Med 6:1–100
Iancu TC (1989a) Iron and neoplasia: ferritin and hemosiderin in tumor cells. Ultrastruct Pathol 13:573–584
Iancu TC (1989b) Ultrastructural pathology of iron overload 2:475–495
Iancu TC (1992) Ferritin and hemosiderin in pathological tissues. Electron Microsc Rev 5:209–229
Iancu TC, Landing BH, Neustein HB (1977) Pathogenetic mechanisms in hepatic cirrhosis of thalassemia major: light and electron microscopic studies. Pathol Annu 12:171–200
Iancu TC, Shiloh H, Link G, Bauminger ER, Pinson A, Hershko C (1987a) Ultrastructural pathology of iron-loaded rat myocardial cells in culture. Br J Exp Pathol 68:53–65
Iancu TC, Ward RJ, Peters TJ (1987b) Ultrastructural observations in the carbonyl iron-fed rat, an animal model for hemochromatosis. Virchows Arch B Cell Pathol Incl Mol Pathol 53:208–217
Iancu TC, Shiloh H, Raja KB, Simpson RJ, Peters TJ, Perl DP, Hsu A, Good PF (1995) The hypotransferrinaemic mouse: ultrastructural and laser microprobe analysis observations. J Pathol 177:83–94
Iancu TC, Perl DP, Sternlieb I, Lerner A, Leshinsky E, Kolodny EH, Hsu Amy, Good PF (1996) The application of laser microprobe mass analysis in the study of biological materials. BioMetals 9:57–65
Iancu TC, Deugnier Y, Halliday JW, Powell LW, Brissot P (1997) Ultrastructural sequences during iron overload in genetic hemochromatosis. J Hepatol 27:628–638
Jellinger K, Kienzel E, Rumpelmair G, Riederer P, Stachelberger H, Ben-Shachar D, Youdim MBH (2006) Iron melanin complex in substantia nigra of Parkinsonian brain: an X-ray analysis. J Neurochem 59:1168–1171
Mandel S, Amit Tamar, Bar-Am Orit, Youdim MBH (2007) Iron dysregulation in Alzheimer’s disease: multimodal brain permeable iron chelating drugs, possessing neuroprotective-neurorescue and amyloid precursor protein processing regulatory activities as therapeutic agents. Prog Neurobiol 82:348–360
Moos T, Rosengren Nielsen T, Skjorringe T, Morgan EH (2007) Iron trafficking inside the brain. J Neurochem 103:1730–1740
Perl DP, Good PF (1992) Comparative techniques for determining cellular iron distribution in brain tissues. Ann Neurol 32:S76–S81
Richter GW (1958) Electron microscopy of hemosiderin: presence of ferritin and occurrence of crystalline lattices in hemosiderin deposits. J Biophys Biochem Cytol 4:55–58
Richter GW (1978) The iron-loaded cell—the cytopathology of iron storage. Am J Pathol 91:362–404
Richter GW, Bessis MC (1965) Commentary on hemosiderin. Blood 25:370–374
Rouault TA (2001) Iron and the brain. Nat Genet 28:299–300
Rouault TA, Zhang DL, Jeong SY (2009) Brain iron homeostasis, the choroid plexus, and localization of iron transport proteins. Met Brain Dis 24:673–684
Takanashi M, Mochizuku H, Yokomizo K, Hattori N, Mori H, Yamamura Y, Mizuno Y (2001) Iron accumulation in the substantia nigra autosomal recessive juvenile parkinsonism (ARJP). Parkinsonism Relat Disord 7:311–314
Vidnes A, Helgeland L (1973) Sex and age differences in the hemosiderin content of rat liver BBA 328:365–372
Youdim MBH, Yehuda S, Ben-Shachar D, Ashkenazi R (1982) Behavioral and brain biochemical changes in iron-deficient rats: the involvement of iron in dopamine receptor function. In: Pollit E, Leibel RL (eds) Iron deficiency: brain biochemistry and behavior. Raven Press, New York, pp 39–56
Zuyderhoudt FMJ, Linthorst C, Hengeveld P (1978) On the iron content of human serum ferritin, especially in acute viral hepatitis and iron overload. Clin Chim Acta 90:93–99
Acknowledgments
This work was supported by grant 11-2009 by the Dan David Foundation and by a contribution from the Milman Fund for Pediatric Research.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Iancu, T.C. Ultrastructural aspects of iron storage, transport and metabolism. J Neural Transm 118, 329–335 (2011). https://doi.org/10.1007/s00702-011-0588-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00702-011-0588-7