
Abstract
Aegilops geniculata Roth is an annual grass relative to cultivated wheats and is widely distributed in North Africa. In order to understand the diversity of this species, 14 populations collected in different bioclimatic areas in northern Algeria were analyzed using morphological and biochemical characters. Principal component analyses (PCA) based on the inflorescence characters and ecological parameters allowed the separation of populations in two mainly bioclimatic clusters characterized by different morphological patterns. Populations originated from humid and sub-humid coastal areas were characterized by vigorous spikes. Samples collected from intermediate and high mountains with humid and semi-arid conditions had long and lanceolate spikes. Individuals with small and narrow spikes were characteristics of steppic highlands in semi-arid conditions and high mountains with humid bioclimate. Individuals were distinguished successively by spike width, spike length, rachis length and awns number. Electrophoretic analyses of high molecular weight glutenin subunits (HMW-GS) based on the phenotypic variability and genetic distances revealed a significant variation within and between populations associated with bioclimatic conditions, in particular winter temperature. Genetic diversity was higher in populations growing under warm bioclimates than in those from cold bioclimates. These results suggest that a part of the variation for HMW glutenin variability is adaptive.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Aegilops geniculata Roth is an annual, self-fertile, allotetraploid species (2n = 4x = 28, MU-genomes), belonging to tribe Triticeae Dumort., subtribe Triticinae Griseb. (van Slageren 1994). This species has a wide distribution around the Mediterranean Sea, showing adaptations to a large range of environmental constraints. Among the 22 species of the genus Aegilops, some species, including Ae. geniculata, are particularly interesting as sources of resistance to various diseases and pests, drought and salinity (Gill et al. 1985; Farooq et al. 1996; Rekika et al. 1998; Zaharieva et al. 2001).
Perrino et al. (1993) and Zaharieva et al. (1999), using morphological and phenological characters found a high variation between Ae. geniculata populations in Italy and Bulgaria, respectively. This variation was mainly explained by the adaptation of this species to different pedoclimatic conditions. Intraspecific variation within Ae. geniculata could also be due to the occurrence of natural hybridization with other tetraploid Aegilops species sharing the U-genome (Pazy and Zohary 1965; Zohary 1965; Kimber and Feldman 1987). The tendency of these tetraploid species to form mixed populations facilitates further hybridization. Feldman (1965) described many intermediate and introgressed types occurring in mixed populations of Ae. peregrina (Hack.) Maire and Weiller, Ae. biuncialis Vis. and Ae. geniculata. This considerable variability led to some taxonomists to propose different classifications. Maire (1955) included Ae. geniculata, together with Ae. neglecta Req. ex Bertol, in the same taxon and considered Ae. neglecta as a subspecies of Ae. geniculata. Quezel and Santa (1962) clustered together Ae. geniculata and Ae. neglecta into the same species, Ae. triuncialis L.
In Algeria, Ae. geniculata is found in a large range of climatic conditions. In some areas, it forms mixed populations with Ae. neglecta and many intermediate types resulting from hybridizations can be observed (Bandou, unpublished).
Some genetic diversity studies based on protein or DNA polymorphism have been carried out on Aegilops species, but more frequently on the diploid species that are the genome donors to polyploid Aegilops and Triticum species (Fernandez-Calvin and Orellana 1990; Rodriguez-Quijano et al. 2000; de Bustos and Jouve 2006; Sun et al. 2006). Fernandez-Calvin and Orellana (1990) analyzed the variation of HMW-GS in diploid Aegilops species of section Sitopsis and concluded that Ae. speltoides Tausch was the probable donor of the B-genome of wheat. Rodriguez-Quijano et al. (2000) analyzed the polymorphism of HMW glutenin subunits in Ae. umbellulata Zhuk., Aegilops comosa Sibth. and Sm. and Ae. markgrafii Greuter L. (donors of the U-, M- and C- genomes, respectively) identified new alleles that could be used in wheat quality improvement. Sun et al. (2006) identified two HMW-GS subunits (x and y) from several accessions of Ae. searsii Feld. and Kis. that were similar to those found on the Glu-1 locus of wheat species.
De Bustos and Jouve (2006), analyzing the HMW-GS in three diploids species, Ae. comosa, Ae. uniaristata Vis. and Ae. speltoides, highlighted new glutenins coded by Glu-Mx, Glu-Unx and Glu-Sy, respectively at these species. They showed that x and y subunits were heterogeneous between species and allowed to differentiate rye from wheat and Aegilops.
There are few genetic diversity studies in Ae. geniculata, except some works using either RFLP (Zaharieva et al. 2001) or RAPD polymorphism (Zhang et al. 1996; Monte et al. 1999). Zaharieva et al. (2001) found a large variability between populations originating from different Mediterranean regions and distinguished two groups, from the North and the South of the Mediterranean Sea. Monte et al. (1999), by studying the genetic variation for RAPD markers in Spanish populations of Ae. geniculata, found a significant correlation between some RAPD bands and ecogeographical factors. They suggested that part of the observed variation could be adaptive.
The primary objective of this paper is to assess and understand the genetic diversity of Ae. geniculata in Algeria in order to improve the management of genetic resources. For this purpose, we analyzed the phenotypic and genetic diversity of natural populations in relation to bioclimatic factors from a wide sampling through a large range of ecological conditions. Algeria is characterized by a North–South bioclimatic gradient expressed by showing contrasted ecological conditions. Genetic diversity in natural populations was poorly investigated except some works in Poaceae species (Aïnouche et al. 1995, 1999; Amirouche and Misset 2003, 2007). In the present study, populations of Ae. geniculata were collected under various bioclimatic and ecological conditions from the coastal plains in the North to the steppic highlands regions in the South, including the mountains of Tellian Atlas. The analysis of genetic diversity was based on the variation of morphological characters and HMW-GS. Principal component analyses (PCA) based on inflorescence characters and frequencies of glutenin subunits in relation to ecological parameters, were performed. The morphological variations within and between populations and population groups were also estimated using variance analyses (ANOVA) and the diversity analyses of glutenins was completed by an analysis of genetic distances.
Materials and methods
Collection and sampling
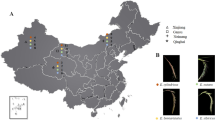
Fourteen populations of Ae. geniculata were collected in various bioclimatic and ecological conditions of northern Algeria (Fig. 1, Table 1). The collecting sites extended from the coastal region to the Atlas Tellian mountains, including the coastal plains and the steppic highlands, along a gradient of increasing aridity from North to South. Each sampling site was characterized by the main ecological factors of Mediterranean climate using the bioclimatic coefficient (Q2). The average annual rainfall (P) and the average temperatures of the hottest month in summer (M) and in particular those of the coldest month in winter (m) correlated with altitude were indicating continentality and bioclimatic variants. In each sampling site, individual plants were randomly collected (separated by at least one meter) for the morphological analyses, and one spike per plant was sampled to analyse the polymorphism of subunits of HMW glutenins.

Location of sampled populations of Aegilops geniculata Roth in Algeria. Bioclimatic limits according to Stewart (1974)
Morphological analyses
Morphological analyses were based on 11 morphological characters of spikes, spikelets and caryopses (Table 2). These characters were measured in ten individuals per population. The morphological characters were selected according to criteria used in taxonomic studies (Zhukovsky 1928; Eig 1929; Maire 1955; Kimber and Feldman 1987) and diversity studies (Zaharieva et al. 2001, 2003a, b). The choice was also based on numerical taxonomy studies (Sneath and Sokal 1973). To evaluate the morphological variability of a population, a first PCA and an analysis of variance (ANOVA) were applied to the 140 individuals. ANOVA was based on the F test which represented a ratio of two variances; one is for example the variance between groups and the other the general variance. The values of this test were compared with limit values given by Snédécor’s tables. In order to characterize the populations at the ecological level, a second PCA was performed on the average of the 14 populations and five ecological parameters. (P = annual rainfall, M = hottest temperature in summer, m = coldest temperature in winter, Q2 = bioclimatic coefficient, and A = altitude).
Glutenin analyses
Glutenin extraction was performed according to the sequential method described by Sing et al. (1991) and modified by Nieto-Taladriz et al. (1994). This method separates the glutenin in two parts, i.e. HMW and low molecular weight (LMW) subunits. At least five grains per plant were used for protein extraction. Proteins were extracted from one-half grain collected from the first spikelet on the base of a spike. Glutenin extraction was made into a solution of Tris–HCl buffer (pH 8.0) and propanol (50% v/v). In this solution, the proteins were reduced at 60°C with dithiothreitol at 1% and 4-vinylpyridin at 1.4%, respectively. After centrifugation, the supernatant was dissolved in 0.08 M Tris–HCl buffer (pH = 8) including 2% of sodium dodecyl sulfate (SDS), 40% glycerol and 0.02% bromophenol. SDS-PAGE experiment was carried out in 12% polyacrylamide. The samples of Aegilops populations separated by those from wheat cultivars, Cajeme and Ablaca, were loaded on the acrylamide gel. After running at constant current of 40 mA, the gels were fixed using trichloroacetic acid (TCA) and stained in Coomassie blue R 250. Analyses were carried out on the populations treated in the morphological analysis (Table 1). The different HMW-GS were first numbered according to their mobility without taking into account their genetic determination. These data, collected for each plant, enabled to compute and analyze the relationship between populations and ecological parameters, using PCA. Comparisons based on the genetic constitution of HMW-GS reported on hexaploid and tetraploid wheats and related species (for review see Shewry et al. 2006) with the different subunits inherited in group of two bands enabled to attribute each group to one of the two loci which were provisionally named here locus A and locus B. An alphabetic nomenclature was attributed to the allelic variants attributed at each locus. Genetic variations were estimated from allelic frequencies for each population. For these genetic variations, a dendrogram was built using UPGMA method and BIOSYS-1 program (Swofford and Selander 1989). The genetic distance was the Rogers distance (Wright 1978).
Chromosome counting
The glutenins when separated by one-dimensional SDS-PAGE generally give two subunits (two bands) in diploid species, four subunits in tetraploid species and six subunits in hexaploid species. In order to verify and to confirm the ploidy level of some studied samples, chromosome countings were carried out on meristematic cells collected from root tips of germinating caryopses. Feulgen staining was performed after pre-treatment in α-bromonaphtalene at room temperature and fixation in alcohol-acetic acid 3:1 v/v at 4°C. Some collected samples (having six subunits) of Ae. neglecta were proved to belong to the hexaploid form (2n = 6x = 42) and other (having four subunits) to the tetraploid form (2n = 4x = 28) by chromosomes counting. These samples of Ae. neglecta were analyzed at level population variability of glutenin patterns and they were not taken into account in morphological and glutenin analyses of phenotypic and genetic diversity. These samples were not taken into account in morphological and glutenin analyses.
Results
Morphological variability
The PCA analysis performed on 140 individuals and based on eleven inflorescence characters, explains 66.82% of the total variation with 38.61%, 14.49% and 13.72% for axis 1, 2 and 3, respectively. The characters were distributed in decreasing order of loading and correlation along the PC 1 (Fig. 2). Diameter of spike (K), width of glume and lemma (M, Q), caryopsis (U) and rachis (S) were more correlated between them and showed the highest loadings in relation to PC 1. However, lengths of these characters (C, P, L and T) were less correlated and had the lower loadings on the same PC. The length of rachis (R) contributed weakly to PC 1 but had a significant loading in relation to PC 2. This character expressed the length structure of inflorescence. The number of awns (N) contributed essentially to the construction of PC 3 (data not shown).

Principal component analysis of inflorescence characters of 14 populations belonging to Ae. geniculata Roth. K, M, S, Q and U character group of diameter and widths; C, P, L and T character group of lengths; R and N independent characters
The individuals were separated into two mainly clusters along the PC 2 (Fig. 3) according to bioclimatic conditions. Each cluster comprised three groups distributed along the PC 1. The first cluster (I) included the populations collected in Oranie and Aurès regions, under sub-humid and semi-arid conditions. The individuals of this cluster were located in the positive part of PC 2 and distributed along the PC 1 into three groups (1, 2 and 3) according to an increasing gradient of aridity, from sub-humid in positive part to semi-arid in negative part. The group 1 was composed by populations collected from the coastal area (Oued-Mellah) and the plain (Tessala 1 and 2), under temperate sub-humid bioclimate. The plants of this group were characterized by short and wide spikes (4.46–5.40 mm). Spikelets were inserted on short and wide rachis. Awn numbers varied from three to five. The group 2 comprised populations from Aurès mountains (Chélia 1 and 2), collected under cold conditions, intermediate between the sub-humid and the semi-arid climate. The individuals of this group were distinguished by long spikes (2.13–2.32 cm) with spaced spikelets on a long rachis (6.26–6.80 mm) and carried few awns (3 awns). The group 3 was made up of populations collected under fresh semi-arid climate from steppic highlands (Hassasna 1 and 2). Individuals of these populations were characterized by short and narrow spikes (1.45–1.73 cm in length and 3.47–3.57 mm in width) and their spikelets were weakly spaced on a short rachis (5.01–5.1 mm) and had three or four awns. The second cluster (II) comprised populations collected in Kabylie under humid conditions. Individuals of this cluster were situated in the negative part of PC 2, and were distributed along PC 1 along an increasing gradient of altitude into three groups: 4, 5 and 6. The group 4 was composed of populations of the coastal area (Tigzirt 1 and 2) and the plain (Mekla). These populations grew under temperate and warm and humid climates. The group 5 included populations from mountains situated at a mean altitude comprised between 700 and 880 m (Beni-Yenni, Aït-Méraou), in a fresh and humid climate. The group 6 comprised populations from mountains (Tikjda, Yakouren) located at high altitude from 900 to 1478 m and having a temperate humid climate. Plants collected from the plain and the coastal area (Mekla, Tigzirt 1 and 2) were characterized by short and wide spikes. Those from mountains located at middle altitude (Beni-Yenni and Aït-Méraou) had long and lanceolate spikes. Finally, the populations from high-altitude mountains (Tikjda and Yakouren) were differentiated by small and narrow spikes.

Principal component analysis of 140 individuals from 14 populations of Ae. geniculata Roth separated into two bioclimatic clusters. Cluster I: sub-humid and semiarid bioclimatic conditions (with group 1: from coast and plain open star; group 2: from mountains of high altitude shaded star and group 3: from high steppic lands filled star). Cluster II (with group 4: from coast and plain open triangle; group 5: from mountains of mean altitude shaded triangle and group 6: from mountains of high altitude filled triangle)
Variability within and between populations and groups
Table 3 shows the values of F test and the probabilities obtained from variance analysis. The F test revealed no significant difference (P < 0.05) within populations for all characters, indicating morphological similarities between individuals within the same populations. Whereas, it exhibited a significant difference (P < 0.05) between populations for all characters. Significant differences were expressed mainly for the length of spike, glume, lemma and caryopsis at group levels. However, rachis and caryopsis lengths and number of awns expressed a significant variation at cluster levels. These results confirmed the hierarchy of inflorescence characters obtained by the PCA. The characters of length, particularly those of rachis and caryopsis, as well as awns number, differentiated the groups and clusters and correlated with environmental conditions (Fig. 4b). Consequently, they expressed more ecological diversity than the thickness characters. These latter characters explained primarily the variation between the populations.

Principal component analysis of 14 Algerian populations belonging to Ae. geniculata Roth based on five ecological factors ( P, M, m, Q2 and Alt). Cluster I: sub-humid and semiarid bioclimatic conditions (with group 1: from coast and plain open star; group 2: from mountains of high altitude shaded star and group 3: from steppic highlands filled star). Cluster II (with group 4: from coast and plain open triangle; group 5: from mountains of mean altitude shaded triangle and group 6: from mountains of high altitude filled triangle). Distribution in scattergraph of PC 1 and PC 2 a of populations, b ecological parameters (Q2, P, M, m and Alt) and morphological characters (C, K, L, M, N, P, Q, R, S, T and U)
Ecological characterization of morphological groups
A second PCA was performed on the averages of the 14 populations by taking into account both the morphological characters and climatic parameters (P, M, m and Q2) with altitude (Alt) (Fig. 4). The first three axes accounted for 71.99% with 39.43%, 21.47% and 11.09% for axis 1, 2, and 3, respectively. The populations showed the same grouping in the plane 1–2 (Fig. 4a) as in the first morphological PCA. The cluster I of populations collected under sub-humid and semi-arid climatic conditions from Oranie and Aurès (Tessala 1 and 2, Oued-Mellah, Chélia 1 and 2, and Hassasna 1 and 2) occupied mainly the positive part of PC2 and the negative part of PC1, whereas, the cluster II of populations collected under humid climate from Kabylie (Tigzirt 1 and 2, Mekla, Yakouren, Tikjda, Beni-Yenni and Aït-Méraou) was located essentially in the negative part of PC 2 and positive part of PC 1. The populations collected from Aurès mountains (Chélia 1 and 2) and steppic highlands (Hassasna 1 and 2) as well as the populations sampled in humid coastal area (Tigzirt 1 and 2) were clearly distinguished from the others and corresponded to morphological patterns described in the first PCA.
This distribution of populations could be explained by a close relationship of three morphological characters (R, T and N) with the ecological parameters (Fig. 4b). The length of the rachis between the first and the second spikelet (R) was strongly linked with altitude (r = 0.59) and m (mean of coldest temperature in winter) (r = −0.67) and the number of awns upper glume (N) exhibited moderate correlations with P (annual rainfall) and Q2 (bioclimatic coefficient) r = 0.41 and r = 0.40, respectively. These characters opposed Chélia 1 and 2 populations, growing under cold semi-arid climate, characterized by long spikes with the spaced spikelets on a long rachis and having few awns, to Tigzirt 1 and 2 populations sampled under warm humid climate and having short spikes with spikelets inserted on short rachis and carrying several awns. The populations Tessala 1 and 2 characterized by small spikes with long caryopsis, growing under cold semi-arid climate with hot summer, were differentiated by the length of caryopsis (T) which exhibited a strong correlation (0.52) with M (mean of hottest temperature in summer).
Glutenin variability
Glutenin patterns and population variability
The electrophoretic spectra (Fig. 5) showed two areas in decreasing order of mobility. The first one included the subunits of HMW-GS while the second one included those of LMW-GS. For a large majority of samples analyzed, the spectra of HMW-GS subunits showed four bands. In some rare samples three bands were observed (Fig. 5 lanes 2, 3) and more rarely six bands (Fig. 5, lanes 1, 4). Three and four bands were encoded at the two loci of the tetraploid species (2n = 4x = 28) Ae. geniculata. Six bands reflected three loci corresponding to hexaploid samples belonging to Ae. neglecta collected from Aït-Méraou and Beni-Yenni. These two homologous loci that were provisionally named locus A and locus B, correspond to the Glu-1 locus of the wheat genome.

SDS-PAGE patterns of high molecular weight and low molecular weight glutenin subunits of endosperm proteins from populations of Ae. geniculata Roth. Beni-Yenni (lanes 2, 3), Yakouren (lanes 5, 6, 7), Tikjda (lanes 10, 11, 12), Aït-Méraou (lanes 13, 14), Mekla (lanes 15, 16, 17), Tigzirt 2 (18, 19); from Ae. neglecta Req. ex Bertol. Aït-Méraou (lane 1), Beni-Yenni (lane 4) and from wheat cultivars Cajeme (lane 8) and Ablaca (lane 9)
Sixteen subunits (bands) were found in all HMW-GS of the tetraploid populations (Fig. 6). They resulted from the activity of the two loci A and B. We named these bands from b1 to b16, according to the decreasing mobility. Among them, five (b2, b3, b6, b10 and b13) had the highest frequencies (0.077–0.121). They could be characteristic of the tetraploid samples, in particular b13 and then b6 and b3 which were occurring in many populations. Several phenotypes were generated resulting from the combination of 16 bands related to the two loci, whose 28 were more frequent, i.e., the phenotypes observed at least in three individuals by population. The most frequent phenotypes were P13 and P17 observed in populations from humid bioclimate, Beni-Yenni (0.55) and Tikjda (0.45) and P21 and P27 in populations from semi-arid, Hassasna 1 and Chélia 2 (0.50).

SDS bands and frequency of 28 phenotypes (P1–P28) and 16 HMW-GS (b1− b16) of Ae. geniculata Roth
Within and between-population phenotypic variability
PCA performed on the 14 populations by taking into account both the frequencies of sixteen bands and the five ecological parameters as shown in Fig. 7. The first two axes accounted 42.66% with 24.12%, and 18.54% for axis 1 and 2, respectively. The distribution of populations in the scattergraph of PC 1 and PC 2 (Fig. 7a) revealed a similar grouping as in the morphological analyses. The cluster I of populations collected under semi-arid and sub-humid climatic conditions (Hassasna 1 and 2, Chélia 1 and 2, Tessala 1 and 2) was distinguished in negative values of PC1. Whereas, the cluster II of populations collected under humid climate (Tigzirt 1 and 2, Tikjda, Mekla, Yakouren, Aït-Méraou, Beni-Yenni) was characterized by positive values of PC1. The population of Oued-Mellah, of sub-humid climate, belonging to cluster I was associated with the populations of cluster II. The correlations between the bands and ecological parameters were shown in scattergraph of PC 1 and PC 2 (Fig. 7b). The bands b4, b8, b10, b14 and b16 of low frequencies (0.025–0.077) and the band b2 of high frequency (0.092) showed close relationships with ecological parameters. The bands b2, b4 and b16 were associated with the populations of semi-arid and sub-humid climate (cluster I). The bands b8, b10 and b14 were specifics to the populations of humid climate (cluster II). In cluster I, the band b2 was characteristic of populations collected under semi-arid climate from steppic highlands (Hassasna 1 and 2) and Aurès mountains (Chélia 1 and 2). It revealed strong negative correlations (r = −0.67 and r = −0.52) with P (annual rainfall) and Q2 (bioclimatic coefficient), respectively. Whereas, the bands b4 and b16 linked with m (mean coldest of temperature in winter) (r = −0.79) and M (mean hottest of temperature in summer) (r = 0.69) and were specifics to populations characterized by cold winter (Chélia 1 and 2) and hot summer (Hassasna 1 and 2), respectively. In the cluster II, the band b8 exhibited strong correlations with climatic factors, (r = 0.53) with P (annual rainfall) and (r = 0.62) with Q2 (bioclimatic coefficient) and moderate correlation (0.49) with m. This cluster grouped the populations growing under humid climate with warm winter and originated from coastal area (Tigzirt 1 and 2), plain (Mekla) and mountains (Tikjda). These populations formed the first, warmer group (H1) of humid climate. Whereas, the bands b10 and b14 correlated between each other (r = 0.51) and with M (r = −0.55), were specific to populations growing under humid climate with moderate summer and cold winter (Aït-Méraou, Beni-Yenni, Yakouren). These populations formed the second colder, group (H2) of humid climate. However, populations growing under sub-humid climate had no specific bands.

Principal component analysis of 14 Algerian populations belonging to Ae. geniculata Roth based on five ecological parameters (P, M, m, Q2 and Alt) and frequencies of 16 glutenin subunits. Distribution in scattergraph of PC 1 and PC 2 1 a of populations, b ecological parameters (Q2, P, M, m and Alt) and glutenins subunits (from b1 to b16)
Structuration of genetic diversity
The UPGMA dendrogram (Fig. 8) based on the allelic frequencies separated the populations into one major group at a level distance of d = 0.25 and three minor groups at level distances of d = 0.30, d = 0.33 and d = 0.37. These groups were separated on the basis of higher frequent alleles located on the two loci which were labeled A and B (Table 4). The major group included the populations characterized by warm winter from sub-humid (Oued-Mellah, Tessala 1 and 2) and humid (Tigzirt 2, Mekla, Tikjda, Yakouren) regions and was differentiated by the first alleles a (b2–b13 and b6–b10) located on A and B locus, respectively, except the population of Mekla which was characterized by the alleles i (b1–b13) on locus A and e (b8–b10) on locus B. The populations of this group were distinguished by mean frequency in both loci ranged from 0.085 to 0.117 and had 8–11 alleles per locus. The populations from semi-arid (Chélia 1, Hassasna 1 and 2) regions as well as two populations (Beni-Yenni, Aït-Méraou) from humid regions formed minor groups characterized by low temperatures. The alleles b (b3–b13), c (6b–b14) and d (b2–b16) located on locus A, differentiated these minor groups composed by populations Chélia 1, Beni-Yenni and Aït-Méraou, and Hassasna 1 and 2, respectively. In comparison with the major group, the mean allelic frequency of these minor groups was lower (0.053–0.077) and they had two to five alleles per locus.

UPGMA dendrogram using genetic distances of Rogers (Wright 1978) showing the relationships among 14 populations of Ae. geniculata Roth sampled in Algeria. Cophenetic correlation = 0.788. Alleles of A locus: a = b2–b13; b = b3–b13, c = b6–b14, d = b2–b16, f = b3–b15. Bioclimate: Humid open circle, Sub-humid shaded circle, Semiarid filled circle. The 14 populations form one major group characterized by warm bioclimatic tendency (with following populations: Oued-Mellah, Tessala 1 and 2, Mekla, Tikjda and Yakouren) and three minor groups characterized by cold bioclimatic tendency (first group with Chélia 1, the second group with Hassasna 1 and 2 and the third group with Beni-Yenni and Aït-Méraou)
Discussion
Morphological diversity and glutenin polymorphism discriminated the populations of Ae. geniculata collected in northern Algeria, in various ecological environments. Characters (Fig. 2) expressing spike thickness, particularly the diameter of spike, were those differentiating better the populations. These characters highly varied between populations (Table 3) but poorly differed between groups and clusters that reflected environmental conditions. Characters related to the length of organs, as the length of spikes varied more significantly between groups. Finally, rachis and caryopsis lengths and number of awns were the traits that differed more between climatic clusters. They were strongly correlated with ecological factors, particularly winter and summer temperatures and altitude. Taxonomists (Zhukovsky 1928; Eig 1929; Hammer 1980; Witcombe 1983; Kimber and Feldman 1987) and in particular the regional florists (Battandier and Trabut 1902; Maire 1955; Quezel and Santa 1962) often used the length of glumes, lemmas and awns to differentiate species and subspecies. In our study, these characters discriminated less the studied populations than thickness characters. The analysis of variance revealed a strong homogeneity within populations and a significant variation between populations for all characters. This variation pattern is specific in most annual colonizing species and particularly in autogamous species (Hamrick and Godt 1997). The reduction of genetic variation was considered as main factor in colonizing success in most annuals and facilitated their rapid colonization specifically of disturbed sites (Hegde et al. 2002).
The PCP from same individuals and populations (Figs. 3, 4) showed that the large variability observed between populations was associated to ecological parameters, namely, the mean of annual rainfall, the summer and winter temperatures and the altitude. In particular, altitude and winter temperature allowed distinguishing three phenotypes. The first one, characterized by large and wide spikes, carrying spikelets on a short rachis and having three to five awns, was mainly present in the coastal areas and plains, under warm, humid and sub-humid climates. The second phenotype, with long and lanceolate spikes, spaced spikelets on a long rachis and few awns, was represented at mean and high altitudes, in cold semi-arid and humid regions. The third phenotype was characterized by small and narrow spikes, with spikelets weakly spaced on a short rachis. This phenotype distinguished the groups located at steppic highlands of fresh semi-arid and at high mountain of cold humid climate.
Perrino et al. (1993) analyzing the diversity of Italian populations of Ae. geniculata on the basis of morphological characters, showed that the morphological variability was associated with ecological conditions. Zaharieva et al. (2003a, b), analyzing numerous populations of Ae. geniculata and other Aegilops species sampled in Bulgaria noted significant differences between populations for most of the morphological characters and observed a significant correlation between the morphology and the ecogeographical region of the studied populations.
The populations of Ae. geniculata collected from Algeria were adapted to a large range of rainfalls, from weak rainfalls (400 mm) in semi-arid areas, to abundant rainfalls (1,350 mm) in humid regions. A large variation in the adaptation to drought conditions was observed in a collection of Ae.geniculata from different geographic origins by Zaharieva et al. (2003a, b) and in a collection from the semi-arid region of Lebanon by Baalbaki et al. (2006).
The electrophoretic analysis of HMW-GS revealed as good way to determinate the polyploidy levels. Some hexaploid samples of Ae. neglecta (2n = 6x = 42) were detected as expressing six subunits and were separated from the tetraploid species (2n = 4x = 28) which had only three to four subunits. The subunits HMW-GS were widely analyzed in diploid species, particularly in Ae. umbellulata which is the one diploid at the origin of the tetraploid species Ae. geniculata. These subunits are controlled by a gene located on the long arm of 1U chromosome (Lawrence and Shepherd 1981; Law and Payne 1983). The proteins encoded at Glu-1 locus generally give two subunits (two bands) when separated by one-dimensional SDS-PAGE (Rodriguez-Quijano et al. 2000).
In Ae. geniculata, the frequency analysis of glutenin subunits revealed a large variability within and between populations. Sixteen bands of HMW-GS generated 28 main phenotypes. These resulted from the allelic combination between the two independent homologous loci (Fig. 6). In Ae. umbellulata, diploid relative of Ae. geniculata, Rodriguez-Quijano et al. (2000) found also a large variability expressed by 11 bands.
The phenotypic variability based on PCA analyses was associated to bioclimatic conditions and altitude. The bands b2, b4 and b16 were specific to populations growing under semi-arid climate. Band b2 characterized the weak rainfall of semi-arid region, b4 distinguished the populations from Aurès mountains (Chélia 1 and 2) characterized by cold winter and band b16 was specific to populations originated from steppic highlands (Hassasna 1 and 2) with a warm summer, whereas, the bands b8, b10 and b14 characterized the populations of humid climate and separated them into warmer (Tigzirt 1 and 2, Tikjda and Mekla) and colder (Aït-Méraou, Beni-Yenni) sub-regions. Therefore, this result suggests that a part of glutenin variability would be adaptive. Monte et al. (1999) analyzing different tetraploid species of Aegilops from Spain and using RFLP markers came to similar conclusions. In populations of wild barley from Algeria, Amirouche and Misset (2003) found also a significant correlation between some subunits of hordein and environmental factors. The variability between populations also reflected the variation due to environmental conditions. Some alleles were associated to winter temperatures. The first allele of locus A (a: b2–b13) differentiated, in a major group, most of the populations growing under warm climates (sub-humid and humid climates). The alleles b: b3–b13, c: b6–b14 and d: b2–b16 clustered into many minor groups the populations growing under cold bioclimates (semi-arid and humid). Nevo et al. (1982), using isozyme markers in wild tetraploid wheat, highlighted a similar correlation between specific isozyme loci and some environmental factors.
The populations growing under warm climates showed more genetic variability, compared with those of cold climates. Their allelic frequency in both loci ranged from 0.085 to 0.117 (8–11 alleles per locus), while the allelic frequency in both loci in populations from cold climates exhibiting a low genetic variability varied from 0.053 to 0.077 (five to seven alleles per locus). The differences of genetic variability between these populations could be explained by fluctuations of gene flow. The warm temperature in humid and sub-humid environments would constitute a favorable factor which increases the diversity within the populations. Hegde et al. (2000) observed in diploid and tetraploid wild wheats large-scale morphological similarities and low-level genetic variation, and suggested that these taxa possess similar types of genetic potential in similar ecological interactions which lead them to specialized adaptation. The spread of genetic diversity between populations would be attributed to a fine-scale adaptation to microhabitat such as soil type and climatic regime. They suggested that genetic variation could also be influenced by other micro- and/or macro-evolutionary forces such as meiotic process. Indeed, Badaeva et al. (2004), using C-banding and FISH (Fluorescence In Situ Hybridization), observed in Ae. geniculata significant intraspecific variability which was due to introgressive hybridization associated with chromosomal rearrangements. They concluded that this species is still undergoing an intensive speciation process. The concentration of diploid species to the Eastern of Mediterranean basin (Zhukovsky 1928; Eig 1929; van Slageren 1994) and their absence from Algeria confirmed the allopatric origin of Ae. geniculata. This species could be considered as young species (Stebbins 1971) which as not yet finished its speciation. This is in agreement with the conclusions of Badaeva et al. (2004).
In conclusion, our genetic and morphological study of Ae. geniculata populations from Algeria showed that all individuals were morphologically homogeneous and that there was little variability within populations. The studied populations clustered into morphological groups adapted to various ecological environments. These groups were differentiated into three main morphological patterns associated to climate, altitude and winter temperature. The wide distribution of Ae. geniculata from coast to Saharian Atlas including mountains of Tellian Atlas, was due to adaptive variation revealed by significant correlations between, on the one hand, some subunits of glutenins and bioclimatic conditions and on the other hand, between some alleles and the same ecological factors. The populations from cold climates were genetically less diverse than those of warm climates.
References
Aïnouche ML, Misset M-T, Huon A (1995) Genetic diversity in Mediterranean diploid and tetraploid Bromus L. (Section Bromus Sm) populations. Genome 38:879–888
Aïnouche ML, Bayer RJ, Gouret JP, Defontaine A, Misset MT (1999) The allotetraploid invasive weed Bromus hordeaceus L. (Poaceae): genetic diversity, origin and molecular evolution. Folia Geobot 34:405–419
Amirouche R, Misset M-T (2003) Hordein polymorphism in diploid and tetraploid Mediterranean populations of the Hordeum murinum L. complex. Pl Syst Evol 242:83–99
Amirouche N, Misset M-T (2007) Morphological variation and distribution of cytotypes in the diploid-tetraploid complex of the genus Dactylis L. (Poaceae) from Algeria. Pl Syst Evol 264:157–174
Baalbaki R, Hajj-Hassan N, Zurayk R (2006) Aegilops species from semiarid areas of Lebanon: variation in quantitative attributes under water stress. Crop Sci 46:799–806
Badaeva ED, Amosova AV, Samatadze TE, Zoshchuk SA, Chikida N, Zelenin AV, Raupp WJ, Friebe B, Gill BS (2004) Genome differentiation in Aegilops. 4. Evolution of U-genome cluster. Pl Syst Evol 246:45–76
Battandier JA, Trabut L (1902) Flore analytique et synoptique de l’Algérie et de la Tunisie. edn. Vve Girault, Alger. pp 410
De Bustos A, Jouve N (2006) Characterization and phylogenetic analysis of the genes coding for high molecular wei glutenin subunits in three diploid species of Aegilops. Int J Pl Sci 167:359–366
Eig A (1929) Monographisch-kritische Übersicht der Gattung Aegilops. Feddes Repert 55:1–228 in German
Farooq S, Shah TM, Asghar M (1996) Intergeneric hybridization for wheat improvement: V. Production of and metaphase 1 chromosome analysis in F1 hybrids of wheat (Triticum aestivum) with Aegilops ovata L. Cereal Res Commun 24:155–161
Feldman M (1965) Further evidence for natural hybridization between tetraploid species of Aegilops. sect. Pleionathera. Evolution 19:162–174
Fernandez-Calvin B, Orellana J (1990) High molecular weight glutenin subunit variation in the Sitopsis section of Aegilops. Implications for the origin of the B genome of wheat. Heredity 65:455–463
Gill BS, Sharma HC, Raupp WJ, Browder IE, Hatchett JH, Harvey TL, Moseman JG, Waines JG (1985) Evaluation of Aegilops species for resistance to wheat powdery mildew, wheat leaf rust, Hussian fly, and greenbug. Plant Dis 69:314–316
Hamrick JL, Godt MJW (1997) Effects of life history on genetic diversity in plants species. In: Silvertown J, Franco M, Harper JL (eds) Plant life histories-ecology phylogeny and evolution. Cambridge University Press, London, p 313
Hammer K (1980) Vorarbeiten zur Monographischen Darstellung von Wildpflanzen-Sortimenten: Aegilops L. Kulturpflanze 28:33–180
Hegde SG, Valkoun J, Waines JG (2000) Genetic diversity in wild wheats and goatgrass. Theor Appl Genet 101:309–316
Hegde SG, Valkoun J, Waines JG (2002) Genetic diversity in wild and weedy Aegilops, Amblyopyrum, and Secale species—A preliminary survey. Crop Sci 42:608–614
Kimber G, Feldman M (1987) Wild wheat. An introduction. Special Report 353, College of Agriculture, University of Missouri, Columbia, p 142, Kimber and Sears 1984
Law CN, Payne PI (1983) Genetical aspects of breeding for improved grain protein content and type in wheat. J Cereal Sci 1:79–93
Lawrence GJ, Shepherd KW (1981) Chromosomal location of genes controlling seed protein in species related to wheat. Theor Appl Genet 59:25–31
Maire R (1955) Flore de l’Afrique du Nord. Eds Le Chevalier, vol III, Paris, pp 65–69
Monte JV, Casanova C, Soler C (1999) Genetic variation in Spanish populations of the genus Aegilops revealed by RAPDs. Agronomie 19:419–427
Nevo E, Goldberg E, Beiles A, Brown AHD, Zohary D (1982) Genetic diversity and environmental associations of wild wheat. Triticum dicoccoides in Israel. Theor Appl Genet 78:260–264
Nieto-Taladriz MT, Branlard G, Dardevet M (1994) Polymorphism of omega-gliadin in durum wheat as revealed by the two-step APAGE/SDS-PAGE technique. Theor Appl Genet 87:1001–1008
Pazy B, Zohary D (1965) The process of introgression between Aegilops polyploids. Natural hybridization between A. variabilis, A. ovata and A. biuncialis. Evolution 19:385–394
Perrino P, Laghetti G, Cifarelli S, Volpe N, Spagnoletti-Chichester PL, Zeuli UK (1993) Wild wheats in southern Italy. In: Damania AB (ed) Biodiversity and wheat improvement. Wiley, Chichester, pp 361–368
Quezel P, Santa S (1962) Nouvelle flore de l’Algérie et des régions désertiques méridionales. Tome I. Edition du CNRS, Paris, p 558
Rekika D, Zaharieva M, Stankova P, Xu X, Souyris I, Monneveux P (1998) Abiotic stress tolerance in Aegilops species. In: Nachit MM et al. Durum Research Network, Proceeding of the SEWANA, South Europe, West Asia and North Africa, ICARDA, Aleppo, Syria, pp 113–118
Rodriguez-Quijano M, Nieto-Taladriz MT, Carrillo JM (2000) Polymorphism of high molecular weight glutenin subunits in three species of Aegilops. Genet Resour Crop Evol 48:599–607
Shewry PR, Halford NG, Lafiandra D (2006) The high-molecular-weight subunits of glutenin. In: Wrigley C, Békés F, Bushuk W (eds) Gliadin and glutenin. The unique balance of wheat quality. AACC, St Paul, pp 143–169
Sing NK, Shepherd KW, Cornich GB (1991) A simplified SDS-PAGE procedure for separating LMW subunits of glutenin. J Cereal Sci 14:203–208
Sneath PHA, Sokal RR (1973) Numerical taxonomy. The principles and practise of numerical classification. Freeman & Co, San Francisco, p 571
Stebbins GL (1971) Chromosomal evolution in higher plants. Arnold, London, p 21
Stewart P (1974) Un nouveau climagramme pour l’Algérie et son application au barrage vert. Bull Soc Hist Nat Afrique du Nord 65:239–248
Sun X, Hu S, Qian W, Hao S, Zhang A, Wang D (2006) Characterization of HMW glutenin subunits from Aegilops searsii and identification of a novel variant HBM glutenin subunit. Theor Appl Genet 113(4):631–641
Swofford DL, Selander RB (1989) Biosys-1. A computer program for the analysis of allelic variation in population genetics and biochemical systematics. Release 1.7. Illinois Natural History Survey, Illinois, pp 43
van Slageren MW (1994) Wild Wheats: a monograph of Aegilops L. and Amblyopyrum (Jaub. & Spach) Eig (Poaceae). Wageningen Agricultural University Papers 94–7, Wageningen, The Netherlands
Witcombe JR (1983) A guide to the species of Aegilops L. International Board for plant Genetic Resources. Wheat Program IBPGR, Rome, p 74
Wright S (1978) Evolution and the genetics of populations, vol 4. Variability within and among natural populations. University of Chicago Press, Chicago
Zaharieva M, David J, This D, Monneveux P (1999) Analyse de la diversité génétique d’Aegilops geniculata Roth en Bulgarie. (In French, with English abstract). Cahiers Agric 8:181–188
Zaharieva M, Gaulin E, Havaux M, Acevedo E, Monneveux P (2001) Drought and heat responses in the wild wheat relative Aegilops geniculata Roth: potential interest for wheat improvement. Crop Sci 41:1321–1329
Zaharieva M, Dimov A, Stankova P, David J, Monneveux P (2003a) Morphological diversity and potential interest for wheat improvement of three Aegilops L. species from Bulgaria. Genet Resour Crop Evol 50:507–517
Zaharieva M, Suenaga K, H.M. William HM, Mujeeb-Kazi A (2003) Microsatellite markers for detection Aegilops geniculata Roth and U genome chromosomes in wheat background. Annual Wheat Newslett 49:75–78
Zhang XY, R-C Wang R, Dong YS (1996) RAPD polymorphisms in Aegilops geniculata Roth. (Ae. ovata auct. non L.). Genet Resour Crop Evol 43:429–433
Zhukovsky PM (1928) Kritiko-systematischeskii obzor vydov roda Aegilops L. (Specierum generis Aegilops L. revisio critica). Trudy Prikl Bot Genet Selekc 18:417–609 in Russian
Zohary D (1965) Colonizer species in the wheat group. In: Baker HG, Stebbins GL (eds) The genetics of colonizing species. Academic Press, New York, pp 403–423
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bandou, H., Rodriguez-Quijano, M., Carrillo, J.M. et al. Morphological and genetic variation in Aegilops geniculata from Algeria. Plant Syst Evol 277, 85–97 (2009). https://doi.org/10.1007/s00606-008-0106-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00606-008-0106-z