
Abstract
Flowering is an essential stage of fruit production. To understand the molecular mechanisms controlling flowering in maloid fruit tree species, we isolated and analyzed genes homologous to Arabidopsis LEAFY (LFY; flower meristem identity gene) and TERMINAL FLOWER 1 (TFL1; inflorescence meristem identity gene) from six fruit tree species in the subfamily Maloideae of the Rosaceae; apple (Malus × domestica), Japanese pear (Pyrus pyrifolia), European pear (Pyrus communis), quince (Cydonia oblonga), Chinese quince (Chaenomeles sinensis), and loquat (Eriobotrya japonica). Two LFY homologues and two TFL1 homologues were cloned from all six maloid species by rapid amplification of 3′ and 5′ cDNA ends, reverse transcription-PCR, and PCR with genomic DNA. Phylogenetic analysis by the neighbor-joining method showed that the two LFY homologues and two TFL1 homologues were classified into two distinct clades. The presence of multiple copies of LFY and TFL1 homologues is discussed with reference to the polyploid origin of the subfamily Maloideae.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Flowering is an essential stage for fruit production, and thus an understanding of the genetic mechanisms underlying the flowering event is important for efficient fruit production. During the last decade, molecular mechanisms of flowering have been studied extensively in herbaceous “model” plants such as Arabidopsis and snapdragon (Antirrhinum majus). Genetic models for flowering time, floral meristem identity, and flower differentiation have been proposed upon the identification of several transcriptional regulator genes (Araki 2001; Battey and Tooke 2002; Parcy et al. 1998; Ratcliffe et al. 1999; Liljegren et al. 1999). In contrast, studies on the molecular mechanisms of flowering in fruit trees have just begun. Several flowering-related genes have been cloned from grapevine (Vitis vinifera) (Carmona et al. 2002; Boss et al. 2001), kiwifruit (Actinidia deliciosa) (Walton et al. 2001), and apple (Malus × domestica) (Kotoda et al. 2000, 2002; Wada et al. 2002; Sung et al. 1999, 2000; Jeong et al. 1999). In this paper, we isolated genes homologous to Arabidopsis LEAFY (LFY) and TERMINAL FLOWER 1 (TFL1) from six fruit tree species of the subfamily Maloideae of the Rosaceae.
LFY is a transcriptional regulation gene thought to play a primary role in determining flower meristem identity. FLORICAULA (FLO), a LFY homologue in snapdragon, has been shown to have almost the same role. Loss of function mutants of these genes result in the conversion of flowers into indeterminate secondary shoots (Coen et al. 1990; Weigel et al. 1992). Conversely, constitutive expression of LFY and its homologues has been shown to be sufficient to promote flower initiation and development in Arabidopsis, poplar (Weigel and Nilsson 1995), and citrus (Pena et al. 2001). Furthermore, alternation of inflorescence architecture, from indeterminate to determinate, was observed in Arabidopsis transformed with a chimeric LFY gene expressed constitutively under the control of the cauliflower mosaic virus (CaMV) 35S promoter.
TFL1 encodes a protein that likely plays a role in signaling, perhaps as an inhibitor of mitogen-activated protein kinase pathways (Corbit et al. 2003), and is a putative regulator gene involved in the control of flowering time and floral architecture. The Arabidopsis tfl1 mutant forms a terminal flower immediately after bolting and inflorescence architecture is altered from indeterminate to determinate. Furthermore, the mutants bolt and flower earlier than the wild type (Shannon et al. 1991; Bradley et al. 1997). CENTRORADIALIS (CEN), a TFL1 homologue in snapdragon, has almost the same function as Arabidopsis TFL1, and the cen mutant shows altered inflorescence architecture (from indeterminate to determinate; Bradley and Meeks-Wagner 1996). A recent study in pea (Pisum sativum) revealed the presence of three homologues of TFL1. Interestingly, one of these is involved in inflorescence development and another in the transition from vegetative to reproductive phase (Foucher et al. 2003).
The Maloideae is one of the four subfamilies of the Rosaceae, which includes several important fruit tree species such as apple (M. × domestica), Japanese pear (Pyrus pyrifolia), European pear (Pyrus communis), quince (Cydonia oblonga), Chinese quince (Chaenomeles sinensis), and loquat (Eriobotrya japonica). Their floral morphological and phenological traits, such as inflorescence architecture and bloom season, vary. Pear forms a raceme inflorescence with eight to ten flowers. The pear inflorescence is considered to be indeterminate because side or lateral flowers bloom first. Apple also forms a raceme inflorescence with five flowers. In contrast to the pear inflorescence, that of apple is considered to be determinate because a terminal flower blooms first (Westwood 1978). Loquat forms a big inflorescence with 50–60 flowers, which is comprised of several panicles. In contrast to these species, quince and Chinese quince bear solitary flowers on shoot apexes without forming peduncles. The bloom season of maloid fruit tree species also varies, with apple, pear, and quince blooming in April and loquat from November to February in Japan.
We assumed that the variation in inflorescence architecture in maloid fruit tree species could be at least partially regulated by LFY and TFL1 homologues because LFY and TFL1 homologues determine flower and inflorescence meristem identities and flowering time in many plant species as described above. In this paper, therefore, we have isolated and characterized LFY and TFL1 homologues from six maloid fruit species as the first step towards our ultimate goal of understanding the molecular mechanism controlling flowering and inflorescence development in maloid fruit species.
Materials and methods
Plant materials
Apple (M. × domestica) cv. Fuji, Japanese pear (P. pyrifolia) cv. Housui, European pear (P. communis) cv. Bartlett, quince (C. oblonga) cv. Smyruna, Chinese quince (C. sinensis) cv. unknown, and loquat (E. japonica) cv. unknown were used. All plant materials were collected from adult trees grown in the field. Buds before floral differentiation in June, floral buds in September and December, mature leaves, hypanthiums with calyx lobes and ovaries, styles with stigmas, stamens, petals, and peduncles were collected for RNA isolation. Young leaves for DNA isolation were collected in spring. They were stored at −80°C until use.
RNA and DNA isolation
Total RNA was isolated from 0.3–0.5 g plant material by the modified cetyltrimethylammmonium bromide (CTAB) method, as described by Kotoda et al. (2000). Single strand cDNA was synthesized from 1 μg total RNA with oligo-dT adaptor primer by reverse transcriptase ReverTra Ace (Toyobo, Japan) after DNase I treatment (Takara Bio, Japan). Genomic DNA was isolated from 2 g leaves by the CTAB method (Doyle and Doyle 1987) and purified by PEG precipitation (Rowland and Nguyen 1993).
Cloning of homologues by PCR
We used a combination of polymerase chain reaction (PCR) of genomic DNA, reverse transcription (RT)-PCR, and rapid amplification of cDNA ends (RACE) techniques to isolate LFY and TFL1 homologues (Table 1). First, partial sequences of LFY and TFL1 homologues were isolated by RT-PCR with degenerate primers LFY-F-1/LFY-R-1 and TFL-F-1/TFL-R-1, which were designed from conserved regions of LFY and TFL1, respectively. PCR was performed with TaKaRa Ex Taq DNA polymerase (Takara Bio) and PCR products were cloned in pGEM-T easy vector (Promega, Madison, Wis.). DNA sequences of the clones were determined with BigDye (Applied Biosystems, Foster City, Calif.) or Dynamic (Amersham Biosciences, Piscataway, N.J.) terminator cycle sequencing using ABI PRISM 310 Genetic Analyzer (Applied Biosystems). Several LFY and TFL1 homologue gene-specific primers for further experiments were designed based on the partial DNA sequences (Table 1).
3′- and 5′-RACE to obtain full length clones were performed using cDNA of floral buds with a GeneRacer kit (Invitrogen, Carlsbad, Calif.). Touchdown PCR was performed with five cycles of 94°C for 30 s, 72°C for 60 s, followed by five cycles of 94°C for 30 s, 70°C for 60 s and finally 30 cycles of 94°C for 30 s, 56°C for 30 s, 72°C for 60 s using gene-specific primers (LFY-F-1, LFY-R-2, TFL-F-2, and TFL-R-2) paired with GeneRacer primers. Nested PCR was performed with 30 cycles of 94°C for 30 s, 56°C for 30 s, 70°C for 60 s using gene-specific primers (LFY-F-2, LFY-R-3, TFL-F-3, and TFL-R-3) paired with GeneRacer nested primers.
Since two kinds each of LFY and TFL1 homologues were isolated from Japanese pear, quince, and loquat by RACE, two different gene-specific primer sets that could be used to amplify almost full length LFY and TFL1 homologues were designed from conserved regions present at the 5′- and 3′- untranslational regions of the cDNAs (Table 1). RT-PCR was performed with 35 cycles of 94°C for 30 s, 30 s of designated annealing temperatures for the genes to be amplified, 72°C for 60 s. The annealing temperatures used were 54°C for LFY-1, 55°C for LFY-2, 55°C for TFL1-1, and 50°C for TFL1-2. TFL1-2 homologues of apple, quince, and Chinese quince were cloned by PCR with the genomic DNA using the same primers with 35 cycles of 94°C for 30 s, 50°C for 30 s, 72°C for 120 s.
DNA sequence and phylogenetic analysis
cDNA and genomic DNA sequences were aligned using GENETYX (GENETYX, Tokyo, Japan) with manual adjustment. Putative introns in the genomic DNA sequences were determined and excluded from the analyses. Amino acid sequences were aligned and identity scores were calculated using a CLUSTAL X program (Thompson et al. 1997). Phylogenetic trees were constructed using the neighbor-joining method (Saitou and Nei 1987).
Expression analysis
Transcription of LFY and TFL1 homologues of Japanese pear, European pear, apple, and quince were determined by RT-PCR. Single strand cDNAs synthesized as described above were used as template. PCR was performed using gene specific primers with an amount of cDNA equivalent to the amount synthesized from 25 ng total RNA (Table 1) and a program consisting of 40 cycles of 94°C for 30 s, 30 s of the appropriate annealing temperatures for the genes to be amplified, 72°C for 45 s. The annealing temperatures used were 54°C for LFY-1, 55°C for LFY-2, 55°C for TFL1-1, and 50°C for TFL1-2. As a reference, actin gene-specific primers, ACT-F-1 and ACT-R-1, were used with a PCR program consisting of 30 cycles of 94°C for 30 s, 50°C for 30 s, 72°C for 45 s. The PCR products were electrophoresed on a 1% (w/v) agarose gel, stained with ethidium bromide, and detected using a UV illuminator. The experiment was repeated six times to confirm reproducibility.
Results and discussion
LFY homologues in six maloid fruit tree species
Two different types of cDNAs for LFY homologues were isolated from each maloid species used in this study; PpLFY-1 and PpLFY-2 for Japanese pear, PcLFY-1 and PcLFY-2 for European pear, CoLFY-1 and CoLFY-2 for quince, CsLFY-1 and CsLFY-2 for Chinese quince, and EjLFY-1 and EjLFY-2 for loquat. Since the two apple LFY homologues isolated in this study showed more than 99.5% amino acid sequence identity to those already reported (Wada et al. 2002) and seemed to be their alleles, we used the nomenclature of Wada et al. (2002) with the name of the cultivar used, i.e., AFL1-Fuji and AFL2-Fuji.
Deduced amino acid sequence alignment of LFY homologues revealed several species-specific insertions/deletions (indels) (Fig. 1). At around position 35, there are species-specific differences in the number of Ala residues. The number of Ala residues in CoLFY-1, CoLFY-2, CsLFY-1, and CsLFY-2 was less than in the others. Deletion of a few residues specific to CoLFY-1, CoLFY-2, EjLFY-1, PpLFY-1, PpLFY-2, and PcLFY-2 was observed in the central region. At position 180, a single residue insertion was observed in EjLFY-2.

Amino acid sequence alignment of maloid LFY homologues, Arabidopsis LFY, and snapdragon FLO. Plant species from which sequence data are derived are denoted by initials of their scientific name followed by the name of the group of LFY homologues, e.g., PpLFY-1 means LFY-1 of Pyrus pyrifolia. AFL1-Fuji and AFL2-Fuji are LFY homologues of Malus × domestica cv. Fuji (see text for details). Lines below alignment Highly conserved region and four motifs (see details in text), arrows above alignment positions of amino acid sequences used to design primers. Residues conserved in more than 11 sequences are shaded
LFY and its homologues in many plant species contain several characteristic motifs such as a single highly conserved region and four characteristic motifs as described below (Weigel et al. 1992; Frohlich and Meyerowitz 1997). The highly conserved region present in the C-terminal half of the protein shows very low variation among LFY homologues of different plant species. This conserved region is also found in maloid LFY homologues cloned in this study (Fig. 1, conserved region). The first characteristic motif found in LFY homologues of many plant species is the Pro-rich region located at the N-terminal end and known to be well conserved in LFY homologues of angiosperm species, while it is not well conserved in those of gymnosperm (see alignment data of Carmona et al. 2002; Frohlich and Meyerowitz 1997). In maloid LFY homologues, however, this region contains only one or two Pro residues followed by a continuous stretch of Ala residues (Fig. 1, motif I). The second characteristic motif is a short putative Leu zipper structure with regular Leu residues. This motif appeared to be completely conserved in the maloid species tested (Fig. 1, motif II). The third characteristic motif is a basic region consisting mainly of Arg and Lys residues, and the fourth motif is an acidic region consisting mainly of Asp and Glu residues. These two motifs are positioned centrally within the protein and are known to be conserved in LFY homologues of angiosperms (Frohlich and Meyerowitz 1997). They were also found in the maloid LFY homologues and consisted of nine basic residues and about ten acidic residues, respectively (Fig. 1, motifs III and IV).
A phylogenetic tree based on deduced amino acid sequences of maloid LFY homologues revealed two distinct clades: groups LFY-1 and LFY-2. About 95 and 97% amino acid sequence identity was observed among homologues in groups LFY-1 and LFY-2, respectively, while there was about 90% identity between homologues in different groups (Table 2, Fig. 2). These different groups were also discriminated by specific differences in amino acid residues in the sequence, e.g., Asp (LFY-1) and Glu (LFY-2) at position 14, Tyr and Cys at position 44, Arg and Glu at position 50, and Asn and Lys at position 82, etc. (Fig. 1).

Phylogenetic tree of LFY homologues constructed by the neighbor-joining method (Saitou and Nei 1987). Maloid LFY homologues are all located in the portion of the tree indicated by the circle, an enlarged version of which is shown above the tree. Abbreviations for maloid LFY homologues as in Fig. 1. Abbreviations and accession numbers of other LFY homologues in the tree: Antirrhinum majus (FLORICAULA, M55525), Arabidopsis thaliana (LEAFY, M91208), Brassica oleracea (BOFH, Z18362), Escholzia californica (EcFLO, AY188789), Eucalyptus globules (ELF1, AF034806), Ginkgo biloba (GinLFY, AF108228), Ionopsidium acaule (AY219226), Idahoa scapigera (AY219228), Juncus effuses (JunefLFY, AF160481), Leavenworthia crassa (AY219227), Lycopersicon esculentum (FALSIFLORA, AF197934), Nicotiana tabacum (NFL1 and NFL2, U15798 and U15799), Nymphaea odorata (NymodLFY, AF105110), Oryza sativa (RFL, AB005620), Peperomia (PepspLFY, AF106843), Petunia × hybrida (ALF, AF030171), Physcomitrella patens (PPLFY1 and PPLFY2, AB052251 and AB052252), Pisum sativum (UNIFLOLIATA, AF03516), Platanus racemosa (PlaraLFY, AF106842), Populus balsamifera (PTLF, U93196), Titanotrichum oldhamii (GFLO, AY526319), Trochodendron aralioides (TroLFY, AF230078), Vitis vinifera (VFL, AF450278), Welwitschia mirabilis (WelLFY, AF109130), Zamia furfuracea (ZamfulLFY, AF105107), Zea mays (ZFL1 and ZFL2, AY179882 and AY179881)
Since LFY homologues have generally been identified as single copy genes in diploid plant species such as Arabidopsis (Weigel et al. 1992), snapdragon (Coen et al. 1990), tomato (Morinero-Rosales et al. 1999), rice (Oryza sativa) (Kyozuka et al. 1998), and grapevine (Carmona et al. 2002), the presence of two different LFY homologues in maloid plants may reflect the polyploid origin of Maloideae. The origin of Maloieae has been discussed extensively (Sax 1933; Challice 1974, 1981; Phipps et al. 1991). Evidence from isozyme analyses of apples supports an allopolyploid origin (Chevreau et al. 1985; Weeden and Lamb 1987). Recently, phylogenetic analysis of Maloideae species based on DNA sequences for the granule-bound starch synthase gene apparently demonstrated their polyploid origin (Evans and Campbell 2002). As in Maloideae, multiple copies of LFY homologues were found in tobacco (Nicotiana tabacum) (Kelly et al. 1995) and maize (Zea mays) (Bomblies et al. 2003), both of which are tetraploids.
There appeared to be differences in transcriptional pattern between the two LFY homologues; however, both homologues were transcribed extensively in buds where floral differentiation takes place (Fig. 3). Although LFY-1 homologues in pear, PpLFY-1 and PcLFY-1, were transcribed only in the buds, those of apple and quince, AFL1-Fuji and CoLFY-1, were transcribed not only in the buds but also in the leaves, hypanthiums, and styles/stigmas. LFY-2 homologues in pears, PpLFY-2 and PcLFY-2, were transcribed in all tissues examined, while AFL2-Fuji and CoLFY-2 were transcribed only in buds, hypanthiums, styles/stigmas, and stamens. Although the expression pattern of AFL2-Fuji in this study was the same as that previously reported for AFL2, a putative allele of AFL2-Fuji (Wada et al. 2002), some differences between the expression patterns of AFL1-Fuji and AFL1, a previously reported putative allele of AFL1-Fuji (Wada et al. 2002), were found. In contrast to the results of Wada et al (2002), slight transcription of AFL1-Fuji was found in leaves, hypanthiums, and styles/stigmas in our experiment. Differences in PCR sensitivities could have led to the discrepancy between our results and the previous report (Wada et al. 2002). The different transcriptional patterns observed between the two LFY homologues, as well as differences between species, may be due to differences in regulatory sequences. Nonetheless, regardless of the types and species, the two LFY homologues were expressed in buds where flower primordia are formed, suggesting that both homologues could play an important role in floral bud formation in Maloideae.

Transcription of LFY and TFL1 homologues in various organs of Japanese pear, European pear, apple, and quince. RT-PCR amplification of LFY-1 homologues (LFY-F-4 and LFY-R-4), LFY-2 homologues (LFY-F-4 and LFY-R-5), TFL1-1 homologues (TFL-F-6 and TFL-R-2), TFL1-2 (TFL-F-5 and TFL-R-4), and an actin gene (ACT-F-1 and ACT-R-1) was performed. Total RNA was isolated from peduncles, petals, stamen (anthers and filaments), styles/stigmas, hypanthiums with ovaries and calyx lobes, mature leaves, buds in June (before floral differentiation), floral buds in September, and floral buds in December. In quince, there is no peduncle because a solitary flower attaches directly to the end of the short shoot
TFL1 homologues in six maloid fruit tree species
As with LFY, two different types of cDNA for TFL1 homologues were isolated from each maloid species used in this study, namely PpTFL1-1 and PpTFL1-2 for Japanese pear, PcTFL1-1 and PcTFL1-2 for European pear, MdTFL1-1 and MdTFL1-2 for apple, CoTFL1-1 and CoTFL1-2 for quince, CsTFL1-1 and CsTFL1-2 for Chinese quince, and EjTFL1-1 and EjTFL1-2 for loquat. All homologues consisted of 172 deduced amino acid residues and no species-specific indels were found (Fig. 4). However, a total of 25 substituted residues in the sequence alignment of maloid homologues were found. Among them, 17 substitutions (positions 2, 3, 4, 5, 8, 33, 52, 69, 90, 96, 104, 105, 111, 128, 134, 159, 167) were species-specific, while 8 were specific to the type of TFL1 homologue (positions 23, 26, 59, 60, 73, 99, 127, 140), TFL1-1 or TFL1-2.

Amino acid sequence alignment of maloid TFL1 homologues, Arabidopsis TFL1, and snapdragon CEN. Plant species from which sequence data are derived are denoted by the initials of their scientific names followed by the name of the group of TFL1 homologues, e.g., PpTFL1-1 means TFL1-1 of Pyrus pyrifolia. Lines below alignment Amino acid residues important for ligand binding: D-P-D-X-P motif (70–74), His residue at position 86, and G-X-H-R motif (116–119) (Banfield and Brady 2000); arrows above alignment positions of amino acid sequences used to design primers. Residues conserved in more than 11 sequences are shaded
The maloid TFL1 homologues were classified into two distinct clades in a phylogenetic tree based on the deduced amino acid sequences. The TFL1 homologues belonging to the two different clades, TFL1-1 and TFL1-2, shared around 90% identity, while homologues belonging to the same clade had identities of 98% (TFL1-1) and 97% (TFL1-2) (Fig. 5, Table 3). As with multiple copies of LFY homologues, the presence of two distinct TFL1 homologues in the maloid species tested in this study could be attributed to the polyploid origin of Maloideae.

Phylogenetic tree of TFL1 homologues constructed by the neighbor-joining method (Saitou and Nei 1987). Maloid TFL1 homologues are all located in the portion of the tree indicated by the circle, an enlarged version of which is shown above the tree. Abbreviations for maloid TFL1 homologues as in Fig. 4. Abbreviations and accession numbers for other TFL1 homologues included in the tree: Antirrhinum majus (CEN, S81193), Arabidopsis thaliana (TFL1, NM_120465; ATC, NM_128315; FT, NM_105222; TSF, NM_118156; MFT, NM_101672; BFT, NM_125597), Brassica napus (BNTFL1-1, AB017525), Nicotiana tabacum (CET1, AF145259; CET2, AF145260; CET4, AF145261), Lolium perenne (LpTFL1, AF316419), Pisum sativum (PsTFL1a, AY340579; PsTFL1c, AY343326), Lycopersicon esculentum (SP, U84140; SP2G, AY186734; SP5G, AY186736; SP6A, AY186737; SP3D, AY186735; SP9D, AY186738)
TFL1-like proteins are similar to mammalian phosphatidylethanolamine binding proteins (PEBPs), also known as Raf-1 kinase inhibitor protein (RKIP). A crystal structure of CEN, a TFL1 homologue in snapdragon, revealed several important motifs that could be involved in interaction with kinase by ligand binding (Banfield and Brady 2000). SP, a TFL1 homologue in tomato (Lycopersicon esculentum), has also been shown to encode a modular protein with the potential to interact with a variety of signaling pathways (Pnueli et al. 2001). Since the maloid TFL1 sequences contain key amino acid residues that are conserved in TFL1 homologues of other genera, TFL1 homologues of maloid species may function in the same way as TFL1 and CEN.
The function of TFL1 in Arabidopsis involves determination of inflorescence meristem identity as well as floral transition (Bradley et al. 1997). CEN is also involved in inflorescence development in snapdragon (Bradley et al. 1996). SP in tomato, however, is expressed in vegetative apical meristem and controls regulation of the vegetative-reproductive switch (Pnueli et al. 1998). Furthermore, in pea, PsTFL1a and PsTFL1c were identified as TFL1 homologues and have distinct functions. The former plays a role in inflorescence development and the latter is involved in flower initiation (Foucher et al. 2003). Although, as with PsTFL1a and PsTFL1c, the two different types of maloid TFL1 homologues could have distinct functions, we assume that the two maloid types can be attributed to the polyploid nature of maloid species, and that they have the same function. This assumption could be supported by the fact that maloid TFL1-1 and TFL1-2 groups are placed very close in the phylogenetic tree as compared with PsTFL1a and PsTFL1c.
Maloid TFL1 homologues were transcribed mainly in buds (Fig. 3). In all species examined, both TFL1 homologues, TFL1-1 and TFL1-2, were expressed at high levels in buds before floral differentiation, and their expression seemed to decrease after floral differentiation. These observations could support the idea that TFL1 homologues play an important role in floral differentiation. Species-specific patterns of organ-specific expression were also observed, e.g. the presence of MdTFL1-1 transcripts in hypanthiums of apple, MdTFL1-2 transcripts in stamens of apple, and those of PpTFL1-2 and PcTFL1-2 in peduncles of pears. It is unclear whether TFL1 homologues are involved in other pathways in floral organs. Further analysis will be needed to reveal the functions of TFL1 homologues in tissues other than buds.
In conclusion, in this study we have cloned two clones each for LFY and TFL1 homologues from six maloid fruit tree species with different flower architecture and flowering habit. There appeared to be several indels and substitutions of amino acid residues in maloid LFY homologues. As compared to LFY homologues, maloid TFL1 homologues appeared to be highly conserved, although they contain several substituted amino acid residues. The difference in maloid LFY homologues may reflect functional differences, which could result in the differences in flowering habit and inflorescence architectures observed among maloid species. These differences will be analyzed in future studies once transformation systems have been established for all maloid species tested in this study. In the short term, it may be useful to see if any difference can be observed when the homologous genes are expressed heterologously in model plant species such as Arabidopsis, as has been done with AFL1 and AFL2 (Wada et al. 2002).
References
Araki T (2001) Transition from vegetative to reproductive phase. Curr Opin Plant Biol 4:63–68
Banfield MJ, Brady RL (2000) The structure of Antirrhinum centroradialis protein (CEN) suggests a role as a kinase regulator. J Mol Biol 297:1159–1170
Battey NH, Tooke F (2002) Molecular control and variation in the floral transition. Curr Opin Plant Biol 5:62–68
Bomblies K, Wang RL, Ambrose BA, Schmidt RJ, Meeley RB, Doebley J (2003) Duplicate FLORICAULA/LEAFY homologs zfl1 and zfl2 control inflorescence architecture and flower patterning in maize. Development 130:2385–2395
Boss PK, Vivier M, Mastumoto S, Dry IB, Thomas MR (2001) A cDNA from grapevine (Vitis vinifera L.), which shows homology to AGAMOUS and SATTERPROOF, is not only expressed in flowers but also throughout berry development. Plant Mol Biol 45:541–553
Bradley D, Carpenter R, Copsey L, Vincent C, Rothstein S, Coen E (1996) Control of inflorescence architecture in Antirrhinum. Nature 379:791–797
Bradley D, Ratcliffe O, Vincent C, Carpenter R, Coen E (1997) Inflorescence commitment and architecture in Arabidopsis. Science 275:80–83
Carmona MJ, Cubas P, Martinez-Zapater JM (2002) VFL, the grapevine FLORICAULA/LEAFY ortholog, is expressed in meristematic regions independently of their fate. Plant Physiol 130:68–77
Challice JS (1974) Rosaceae chemotaxonomy and the origins of the Pomoideae. Bot J Linn Soc 69:239–259
Challice JS (1981) Chemotaxonomic studies in the Rosaceae and the evolutionary origins of the subfamily Maloideae. Preslia 53:289–304
Chevreau E, Lespinasse Y, Gallet M (1985) Inheritance of pollen enzymes and polyploidy origin of apple (Malus × domestica Borkh.). Theor Appl Genet 71:268–277
Coen ES, Romero JM, Doyle S, Elliott R, Murphy G, Carpenter R (1990) Floricaula: a homeotic gene required for flower development in Antirrhinum majus. Cell 63:1311–1322
Corbit KC, Trakul N, Eves EM, Diaz B, Marshall M, Rosner MR (2003) Activation of Raf-1 signaling by protein kinase C through a mechanism involving Raf kinase inhibitory protein. J Biol Chem 278:13061–13068
Doyle JJ, Doyle JL (1987) A rapid DNA isolation procedure from small quantities of fresh leaf tissues. Phytochem Bull 19:11–15
Evans RC, Campbell CS (2002) The origin of the apple subfamily (Maloideae; Rosaceae) is clarified by DNA sequence data from duplicated GBSSI genes. Am J Bot 89:1478–1484
Foucher F, Morin J, Courtiade J, Cadioux S, Ellis N, Banfield MJ, Rameau C (2003) DETERMINATE and LATE FLOWERING are two TERMINAL FLOWER 1/CENTRORADIALIS homologs that control two distinct phases of flowering initiation and development in pea. Plant Cell 15:2742–2754
Frohlich MW, Meyerowitz EM (1997) The search for flower homeotic gene homologs in basal angiosperms and gnetales: a potential new source of data on the evolutionary origin of flowers. Int J Plant Sci 158:S131–S142
Jeong DH, Sung SK, An G (1999) Molecular cloning and characterization of CONSTANS-like cDNA clones of the fuji apple. J Plant Biol 42:23–31
Kelly AJ, Bonnlander MB, Meeks-Wagner DR (1995) NFL, the tobacco homolog of FLORICAULA and LEAFY, is transcriptionally expressed in both vegetative and floral meristems. Plant Cell 7:225–234
Kotoda N, Wada M, Komori S, Kidou S, Abe K, Masuda T, Soejima J (2000) Expression pattern of homologues of floral meristem identity genes LFY and AP1 during flower development in apple. J Am Soc Hortic Sci 125:398–403
Kotoda N, Wada M, Kusaba S, Kano-Murakami Y, Masuda T, Soejima J (2002) Overexpression of MdMADS5, an APETALA1-like gene of apple, causes early flowering in transgenic Arabidopsis. Plant Sci 162:679–687
Kyozuka J, Konishi S, Nemoto K, Izawa T, Shimamoto K (1998) Down-regulation of RFL, the FLO/LFY homolog of rice, accompanied with panicle branch initiation. Proc Natl Acad Sci USA 95:1979–1982
Liljegren SJ, Gustafson-Brown C, Pinyopich A, Ditta GS, Yanofsky MF (1999) Interaction among APETALA1, LEAFY, and TERMINAL FLOWER 1 specify meristem fate. Plant Cell 11:1007–1018
Molinero-Rosales N, Jamilena M, Zurita S, Gomez P, Capel J, Lozano R (1999) FALSIFLORA, the tomato orthologue of FLORICAULA and LEAFY, controls flowering time and floral meristem identity. Plant J 20:685–693
Parcy F, Nilsson O, Busch MA, Lee I, Weigel D (1998) A genetic framework for floral patterning. Nature 395:561–566
Pena L, Martin-Trillo M, Juarez J, Pina JA, Navarro L, Martinez-Zapater JM (2001) Constitutive expression of Arabidopsis LEAFY or APETALA1 genes in citrus reduces their generation time. Nat Biotechnol 19:263–267
Phipps JB, Robertson KR, Rohrer JR, Smith PG (1991) Origins and evolution of subfam. Maloideae (Rosaceae). Syst Bot 16:303–332
Pnueli L, Carmel-Goren L, Hareven D, Gutfinger T, Alvarez J, Ganal M, Zamir D, Lifschitz E (1998) The SELF-PRUNING gene of tomato regulates vegetative to reproductive switching of sympodial meristems and is the ortholog of CEN and TFL1. Development 125:1979–1989
Pnueli L, Gutfinger T, Hareven D, Ben-Naim O, Ron N, Adir N, Lifschitz E (2001) Tomato SP-interacting proteins define a conserved signaling system that regulates shoot architecture and flowering. Plant Cell 13:2687–2702
Ratcliffe OJ, Bradley DJ, Coen ES (1999) Separation of shoot and floral identity in Arabidopsis. Development 126:1109–1120
Rowland LJ, Nguyen B (1993) Use of polyethylene glycol for purification of DNA from leaf tissue of woody plants. Biotechniques 14:734–736
Saitou N, Nei M (1987) The neighbor-joining method—a new method for reconstructing phylogenetic trees. Mol Biol Evol 4:406–425
Sax K (1933) The origin of the Pomoideae. Proc Am Soc Hortic Sci 30:147–150
Shannon S, Meeks-Wagner DR (1991) A mutation in the Arabidopsis TFL1 gene affects inflorescence meristem development. Plant Cell 3:877–892
Sung SK, Yu GH, An G (1999) Characterization of MdMADS2, a member of the SQUAMOSA subfamily of genes, in apple. Plant Physiol 120:969–978
Sung SK, Yu GH, Nam J, Jeong DH, An G (2000) Developmentally regulated expression of two MADS-box genes, MdMADS3 and MdMADS4, in the morphogenesis of flower buds and fruits in apple. Planta 210:519–528
Thompson JD, Gibson TJ, Plewniak F, Jeanmougin F, Higgins DG (1997) The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Res 25:4876–4882
Wada M, Cao Q, Kotoda N, Soejima J, Masuda T (2002) Apple has two orthologues of FLORICAULA/LEAFY involved in flowering. Plant Mol Biol 49:567–577
Walton EF, Podivinsky E, Wu R (2001) Bimodal patterns of floral gene expression over the two seasons that kiwifruit flowers develop. Physiol Plant 111:396–404
Weeden N, Lamb R (1987) Genetics and linkage analysis of 19 isozyme loci in apple. J Am Soc Hortic Sci 112:865–872
Weigel D, Nilsson O (1995) A developmental switch sufficient for flower initiation in diverse plants. Nature 377:495–500
Weigel D, Alvarez J, Smyth DR, Yanofsky MF, Meyerowitz EM (1992) LEAFY controls floral meristem identity in Arabidopsis. Cell 69:843–859
Westwood MN (1978) Flowering. In: Westwood MN (ed) Temperate-zone pomology. Freeman, New York, pp 171–172
Acknowledgements
This work was supported by a Grant-in-Aid (no. 14656014) for Exploratory Research to R.T. from the Japan Society for the Promotion of Science. We gratefully acknowledge the gift of plant material from K. Banno and T. Minemura of Shinshu University (Nagano, Japan).
Author information
Authors and Affiliations
Corresponding author
Additional information
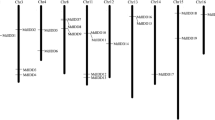
The nucleotide sequence data reported here are available in the DDBJ/EMBL/GenBank database under the accession numbers AB162028 (AFL1-fuji), AB162029 (PpLFY-1), AB162030 (PcLFY-1), AB162031 (CoLFY-1), AB162032 (CsLFY-1), AB162033 (EjLFY-1), AB162034 (AFL2-Fuji), AB162035 (PpLFY-2), AB162036 (PcLFY-2), AB162037 (CoLFY-2), AB162038 (CsLFY-2), AB162039 (EjLFY-2), AB162040 (MdTFL1-1), AB162041 (PpTFL1-1), AB162042 (PcTFL1-1), AB162043 (CoTFL1-1), AB162044 (CsTFL1-1), AB162045 (EjTFL1-1), AB162046 (MdTFL1-2), AB162047 (PpTFL1-2), AB162048 (PcTFL1-2), AB162049 (CoTFL1-2), AB162050 (CsTFL1-2), AB162051 (EjTFL1-2).
Rights and permissions
About this article
Cite this article
Esumi, T., Tao, R. & Yonemori, K. Isolation of LEAFY and TERMINAL FLOWER 1 homologues from six fruit tree species in the subfamily Maloideae of the Rosaceae. Sex Plant Reprod 17, 277–287 (2005). https://doi.org/10.1007/s00497-004-0239-3
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00497-004-0239-3