
Abstract
Key message
Density was more important in shaping crown structure than neighbor species identity. Both species showed high crown plasticity at alternative levels, which may explain species coexistence in mixed broadleaved forests with functionally similar species.
Abstract
Understanding crown response to local competition is essential to predicting stand development in mixed stands. We analyzed data from an 8-year-old field experimental plantation mixing two species according to a crossed gradient of density and species proportion to quantify the effect of a broad range of local neighborhood conditions on the development of young trees at multiple crown levels. We used Fagus sylvatica and Acer pseudoplatanus, as two model deciduous species. They are considered functionally equivalent at the young stages, but with contrasting architectural patterns. For both species: (1) changes in density explained more of the variation on crown development than species proportion (2) much of the effect of competition was accounted for by variables at the stem level, while branch and leaf development within crowns were not directly altered by competition. Both species were able to modify their crowns at the stem level to compete with intra- and inter-specific neighbors: Acer and Fagus were taller with a highest proportion of Fagus as neighbors; Fagus displayed a lower crown base when the proportion of Fagus decreased, while Acer had a lower crown base when the proportion of Fagus around it increased. Both species showed common shapes in allometric relationships but contrasting responses at alternative crown levels. Acer exhibited broader intra-specific variation in its height–diameter relationship and in its crown length, while Fagus displayed higher individual variation of branch development and leaf area than Acer. This study demonstrates that differences in crown development strategy of each species in response to changes in local neighborhood conditions are an important factor in maintaining species coexistence in broadleaved forests and designing mixtures that persist over time.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Individual tree crown architecture, namely the position, size and orientation of the branches and foliar arrangement along the branches, is a primary determinant of forest function and productivity. At the tree scale, it plays a critical role in the competition for space. Tree crown architecture strongly influences the ability to compete with neighboring trees, as it determines the amount of light intercepted by the tree. At the same time, the competitive environment of a tree strongly affects its available space to grow in and its light environment and consequently its crown dimensions and architecture (Osada 2006; Schröter et al. 2012). Most studies to date have typically used simple response variables to study tree response to varying neighborhood conditions, such as variations in stem or crown dimensions (Purves et al. 2007; Davies and Pommerening 2008). Few studies have aimed at revealing the detailed responses of tree crown architecture (Lintunen and Kaitaniemi 2010), but such measurements may be essential to better understand stand development in spatially diverse stands such as mixed stands.
Overall, several studies show that crown size and architecture respond to a variety of exogenous factors such as local competition (Thorpe et al. 2010), site fertility (Dieler and Pretzsch 2013), topography (Lang et al. 2012; Antin et al. 2013) and climate (Lines et al. 2012) and to endogenous factors such as tree size (Thorpe et al. 2010, Antin et al. 2013) and species identity (Purves et al. 2007, Weiskittel et al. 2010). In all studies, endogenous factors appeared to be far more important than exogenous factors, with tree species and tree size being the primary drivers of most crown variables. Competition with neighboring trees is also a major factor, which has been shown to affect crown size and architecture through its various components: proximity, size and species identity of neighbors (Davies and Pommerening, 2008; Dieler and Pretzsch 2013). However, analyses regarding the combined effects of density and neighboring species identity on crown variables are presently scarce or dependent on the response variable examined. For example, Kaitaniemi and Lintunen (2010) found that the effect of density on height and diameter growth was more prominent than the effect of neighbor identity in 4- to 35-year-old mixtures of conifers and broadleaved species. However, in another study performed on similar mixtures, Lintunen and Kaitaniemi (2010) identified that many branch-level variables were mainly affected by neighbor species identity.
Studies analyzing the effects of competition on crown size and architecture have focused on particular sets of architectural parameters: crown length or width (Thorpe et al. 2010), crown rise (Ilomäki et al. 2003), crown asymmetry (Rouvinen and Kuuluvainen 1997), branch angle and branch diameter (Mäkinen and Colin 1999; Lintunen and Kaitaniemi 2010), or branch characteristics and leaf area (Medhurst and Beadle 2001; Osada 2006); with very few attempts to include a response at multiple crown levels (Weiskittel 2003). Quantifying the effects of local competition on the various components of crown architecture may be essential to understanding individual tree responses and to predict species ability to compete with neighboring trees. Parameters describing the various crown components are strongly correlated within and among trees, and an integrated approach that takes into account these correlations is required to analyze the crown components, their interactions and their combined effects in response to competition (Pretzsch and Dieler 2012; Weiskittel et al. 2007).
In this study, we focus on Fagus sylvatica L. and Acer pseudoplatanus L., two species with similar ecological niches that are commonly found in intimate mixture in Western and Central Europe (Hein et al. 2009). Despite their similar growth in the young stages (Hein et al. 2007; Wagner et al. 2010), the two species show enhanced growth in mixture with the other species (Collet et al. 2014). The highest production observed in even mixtures suggests a complementary effect for resource acquisition. The species have contrasting crown architectural patterns (Bell 1991), with Fagus conforming to Troll’s architectural model while Acer follows Rauh’s model. We hypothesize that the complementary effect observed between the two species may originate from differences in crown development and crown responses to competition.
The general objectives of the study are: to analyze crown response of Fagus and Acer to competition from neighboring trees, to estimate whether differences in crown development may explain the complementary effect observed between the two species in the young stages. More specifically, we aim to answer the following questions: (1) To what extent are species-specific relationships among crown components influenced by density, size and species identity of neighboring trees? (2) To what extent are the effects of competition on crown components accounted for indirectly by competition effects on stem parameters (diameter and height)? (3) Do the effects of competition on crown development differ between Acer and Fagus?
We used an experimental Fagus–Acer plantation where soil and climate were not limiting factors and where large gradients of density and species proportion allowed observing changes in crown architecture directly related to inter- and intra-specific competition for light or physical contact by neighboring trees. We developed a model describing tree crown at multiple levels (stem, growth units, branches and leaves) and analyzed relationships among crown components following a hierarchical approach where parameters reflecting higher level components were systematically introduced in models describing lower level components.
Materials and methods
Study site
The study was conducted in the forest of Haye, near Nancy, North-Eastern France (48°38′17.18″N, 6°8′43.03″E), located on a limestone plateau at approximately 400 m a.s.l.. The climate is oceanic with a continental trend (January temperature: 1.2 °C, July temperature: 18.3 °C, annual precipitation: 740 mm). The soil is a brunisol consisting of a silt eutrophic brown soil layered over limestone.
Experimental design
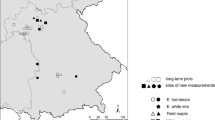
One-year Fagus and Acer saplings grown in nursery were planted on the site in November 1998 following a density-mixing double-clinal design (i.e., an experiment crossing two gradients that vary systematically; Deleuze et al. 1996; Vanclay 2006). The central zone of the study site (65 × 40 m) included 2,014 trees planted according to two perpendicular clines. The first cline was a density gradient, where local tree density varied exponentially with the location on the x-axis, ranging from 1,500 to 25,000 stem ha−1 and the second cline was a mixing gradient where the local proportion of the two species varied linearly with the location on the y-axis, from pure Fagus to pure Acer (Fig. 1). This central zone was surrounded by a 10-m-wide buffer zone with the same tree density and species mixing conditions. In the two zones, all trees were mapped. All other plants (mainly other tree species, Carex sp., Rubus sp. and Cytisus sp.) were removed periodically to avoid any competition other than from planted Fagus and Acer. The site was fenced to prevent browsing damage.

Map of the experimental plot representing trees of the two species studied (Acer in grey and Fagus in black) over the density and the mixture gradients. Thick dotted lines represent the external borders of a 3 m buffer zone (i.e., radius of the neighborhoods used to compute competition indices), thick plain lines represent the external borders of the central zone, and fine plain lines represent the borders of the 25 plots used to select the 120 trees of sample 3. Average density and average species proportion of the 25 plots are indicated in the top and right margins of the figure
Sampling and tree measurements
The modular structure of crowns allows high structural variability at multiple hierarchical levels. Therefore, we designed a hierarchical sampling scheme for tree measurements at three levels (stem, annual growth unit and branch) that were assessed within five nested samples (Table 1).
Sample 1 contained all trees from the central zone (2,014 trees) and trees located in the first buffer zone at less than 3 m from the central zone (612 trees). To calculate competition indices for the trees of the central zone, stem height (H st; Table 2) and diameter at 15 cm (D st; Table 2) were measured on these trees in 2006. Height of the crown base (i.e., height of the first living branch longer than 10 cm; Hcrownst; Table 2) was measured for every tree in the central zone (sample 2).
We considered trees as branched systems where the main stem is referred to as the first-order axis, the branches inserted on the stem as secondary branches, and so on (Barthélémy and Caraglio 2007). A smaller subset including both species was defined to analyze the branching pattern (sample 3, Table 1). To summarize the range of stand conditions being analyzed, the central zone was divided into 25 unequally sized plots with the purpose of having a similar number of trees in each density class (Fig. 1). The x-axis (corresponding to the density gradient) was divided into five classes of decreasing size when the density increased (plot size along the x-axis: 6, 7, 12, 15 and 20 m). The average density for each class was: 2,500, 5,500, 9,000, 16,500 and 21,000 tree.ha−1. The y-axis (corresponding to the mixing gradient) was divided into two small outward classes with an almost “pure” composition (95 % of Acer or 95 % of Fagus) and three equally sized inward plots with various mixing conditions (25, 50 or 75 % of Fagus and 75, 50 and 25 % of Acer). Each of the 15 inward plots included ca. 130 trees and each of the 10 outward “pure” plots ca. 30 trees. In each of the 25 plots, three trees per species were selected (in the “pure” plots, only one species was available), with a total of 120 trees (sample 3). In each plot and for each species, one tree was randomly selected from the 10 % smallest diameter trees of the plot, one tree from the 10 % largest trees and one tree from the 80 % intermediate trees. Tree dimension variability in the sample was maximized to account properly for possible size-dependent effects.
Stem and branch measurements were made on each of the 120 trees: the stem was marked every 20 cm from the ground and, at each mark, the average of two diameters measured in orthogonal directions was recorded. The scars left each year by the winter buds on the stem were located and their height along the stem (HGU) was measured. They were used to determine the length (LGU) and age (AgeGU) of the successive annual growth units on the stem. The insertion height (Hbr) and basal diameter (Dbr) of each living branch inserted on the stem were measured. On Acer and Fagus, branch diameter does not vary linearly with the insertion height on the stem. Both species show an acrotonic growth pattern within each GU, where branches inserted near the top of the GU are larger (Nicolini et al. 2001). To model this pattern and to take into account branch distribution within the GUs, for each GU, the insertion height of each branch within the GU (Hinsbr, Table 1) was computed as a percentage of GU length. Cross-sectional area (SbaseGU, Table 1) at the base of each GU was obtained from stem taper measurements made every 20 cm along the stem, one immediately below the GU base and one immediately above the GU base and from the basal diameter of the branches inserted between these two stem measurements. For each GU, the cross-sectional area at the end of the GU (SendGU, Table 1) was then considered identical to the cross-sectional area at the base of the following GU.
Branch length (L br) and angle (A br) were measured on nine second-order branches on each of the 120 trees of sample 3 (sample 4, 60 trees per species, Table 1). The definition of sample 4 was based on the branching data previously described. Each tree of sample 3 was divided into three equally sized height sections (lower, middle and upper) positioned between the lowest and the highest second-order branch of the tree. In each of the three height sections, three branches were randomly selected by basal diameter criteria, one from the 10 % smallest branches of the height section, one from the 10 % largest branches and one from the 80 % intermediate-sized branches.
Additional measurements on higher order branches were performed for the largest trees, per species and per plot of sample 4 (sample 5, 20 trees per species, Table 1). On the nine second-order branches per tree, all branches of higher order were identified (a total of 360 second-order branches and 7,648 higher order branches (up to seventh order) for both species). For each branch higher than second order, we measured insertion height of the branch (Hinsbr) on its mother branch; branch length (L br); branch basal diameter (D br) and branch taper (for every 20 cm, the average of two diameters measured in orthogonal directions was recorded). The total volume carried by a branch (V br) was calculated as the sum of the volume of the branch and the volumes of all higher order branches carried by that branch.
In October 2007, we collected leaves on branches of sample 5. Leaves of the second or higher order branches were pooled. All leaves were counted, flattened and scanned and total leaf area was estimated using Image-J (Schneider et al. 2012). On large samples, a representative leaf subsample was scanned and both sample and subsample were weighted. The whole sample leaf area was estimated using its mass ratio to the subsample leaf area. The total leaf area per branch (LfAbr) was calculated as sum of the leaf area of the axis and the leaf area of all higher order branches carried by the axis.
Data analysis and modeling
To analyze the effects of local inter-tree competition on the various stem and branch parameters, we computed for each individual tree three simple competition indices from measurements performed on all neighboring trees located in a 3 m circular neighborhood around the target tree:
-
Local density (Dens), calculated from the total number of trees in the neighborhood;
-
Local species proportion (Mix), calculated as the number of Fagus over the total number of trees in the neighborhood;
-
Local dominant height (Hdom), calculated as the average height of the five tallest neighboring trees in the neighborhood (if less than five neighboring trees were available, Hdom was calculated over the total number of neighboring trees). At the GU level, the Hdom index was replaced by HrelGU (HGU/Hdom) to account for the effect of GU location in relation to the upper canopy level.
The three indices had been selected after testing a wide range of competition indices that combined species, height, diameter and distance to target tree. The radius of the circular neighborhood (i.e., 3 m) was selected after testing a range of distances from 50 cm to 10 m (best correlation, results not shown).
For all data sets, we analyzed Fagus and Acer data separately, including the three competition indices within the explanatory variables. Our dataset presented a hierarchical structure with branches nested within GUs, and within trees. Therefore, random effects were explored for those levels (Table 2). For all models, we tested a spatial varying model to take into account local spatial autocorrelation that may arise between neighboring trees (i.e., incorporating spatial linkages of proximity of the observations with a spatial weight matrix), and may result in a wrong estimation of model parameters and in model inaccuracy if not taken into account (Zuur et al. 2009). Additionally, for several of the models, we included an exponential variance submodel to account for residual heteroscedasticity. All analyses were conducted in the statistical program R version 2.14.1 (R Development Core Team 2011).
Stem-level variables
We analyzed stem-level variables collected on sample 1 trees using non-linear mixed models. To analyze the relation between height and base diameter (Eq. 1; Table 2), we followed Näslund (1937). Hcrownst (height of the crown base) was analyzed using a two-step process (Eqs. 2, 3; Table 2). First, we estimated an upper boundary for the relation Hcrownst to Hst: we divided tree height into twenty equal-sized classes and we selected the two trees with the highest crown basis in each class. These trees were used to estimate maximum crown basis height (Hcrownmaxst), following Eq. 2 (Table 2). In a second step, we calculated a reductor of Hcrownmaxst for each tree as the ratio of Hcrownst to the corresponding point on the boundary relation. A value of 1 for the reductor indicates that Hcrownst for this tree reached the upper boundary, while a value close to 0 indicates that Hcrownst was close to the ground level (i.e., no self-pruning). We analyzed the relationship between the reductor and the explanatory variables using a non-linear model (Eq. 3, Table 2).
Annual growth unit-level variables
At this level, we tested a random tree effect in the models to take into account possible correlations among GUs within trees. Growth unit basal diameter (DbaseGU) and the sum of branch cross-sectional areas per GU (SGU) were described as linear models of their parameters and were modeled only for second-order branches (Eq. 4, 5; Table 2). The number of carried branches (NbrGU) on an annual growth unit (Eq. 6; Table 2) was modeled using a Poisson distribution following Mäkinen and Colin (1999).
Branch-level variables
At this level, we tested random tree and branch effects in the models to take into account possible correlations between GUs within trees, and among branches within the same second-order branch. As correlation amongst branches from a same cluster was likely, all the models included a single multilevel random effect combining tree identifiers with branch identifiers to take into account correlations among branches from the same tree as well as among n + 1th order branches carried by the same nth order branch.
Branch length (Lbr) was modeled as a linear regression of Dbr for Fagus, and as a combination of a linear model and a negative exponential model (to account for small branches) for Acer (Eq. 8; Table 2). Branch insertion angle (A br) was modeled as a linear regression of branch insertion height (HinsGU) and Dbr (Eq. 9, Table 2). Branch volume (V br) was a non-linear model adapted from Muukkonen (2007), which describes branch volume as a power function of branch basal diameter (Eq. 10, Table 2). Branch leaf area (LfAbr) was described as a non-linear model of branch cross-sectional area (Sbr) and branch order (Ordbr; Eq. 11; Table 2).
Results
Stem-level variables
The shape of the H st − D st (stem height − stem diameter) relationship did not differ between Acer and Fagus (Fig. 2). The H st − D st relationship reached an asymptote at approximately 4 m at the lowest densities, and at 5.5 and 5 m at the highest densities for Acer and Fagus, respectively (representing an increase of 27 and 20 %, respectively; Fig. 3; Table 2; Eq. 1). Density (Dens) and the proportion of Fagus in the local neighborhood (Mix) affected the slope and the asymptote of the H st − D st allometry. For a given D st value, high density caused considerably larger H st values and the magnitude of this effect increased for larger trees. Both Acer and Fagus saplings tended to be taller with a highest proportion of Fagus as neighbors. For both species, the species proportion effect was of a smaller magnitude than the density effect (asymptote varied between 4.5 at the lowest percentage of Fagus proportion for both species, and 4.75 and 5 m for Fagus and Acer at the highest percentage of Fagus proportion, respectively; representing an increase of 5 and 10 %). The effect of Hdom (local dominant height) on H st was significant and positive for both species.

Fitted allometries for H st (stem height) against D st (stem diameter) in Acer and Fagus (Eq. 1; Table 2; Sample number 1; Table 1). The density indicated for observed individual tree values is the average density for the five classes in which the density gradient was divided. We also show the simulated density-dependent effects on the regression lines fixed at three levels of density (500, 12,000, 25,000), while keeping species proportions at its mean (0.5)

Changes in young trees crown height with Dens (local density) in Acer and Fagus. The density indicated for individual tree values is the average density for the five classes in which the density gradient was divided. a Upper boundary line between Hcrownmaxst (maximum crown basis height) and H st (stem height); the black dots represent the two trees with the highest crown basis in each of the twenty equally sized classes in which tree heights were divided; the grey dots represent the rest of the trees (Eq. 2; Table 2; Sample 2; Table 1) b Relationship between the reductor and D st (stem diameter) (Eq. 3; Table 2; Sample 2; Table 1). A value of 1 for the reductor indicates that the crown height for this tree reached the upper boundary, while a value close to 0 indicates that the height crown was close to the ground level
The upper boundary line of Hcrownmaxst (maximum crown basis height) against H st was well described by a positive linear function for both species, with a lower slope for Fagus (0.39) than for Acer (0.46) (Fig. 3a; Table 2; Eq. 2). For both species, the reductor could be modeled as a negative exponential function of D st, meaning that for a given height, trees with a larger diameter had their crown base lower on the stem (Fig. 3b; Table 2; Eq. 3). For both species, the relationship was a function of density. For a given Hst and Dst, trees growing at higher density showed a Hcrownst closer to the maximum value (i.e., smaller crown length and higher reductor). In addition, Fagus showed a lower crown base (i.e., larger crown length and smaller reductor) when Fagus proportion decreased, while Acer had a lower crown base height when the proportion of Fagus around it increased. Crown base height was on average almost half for Fagus (2.86 cm) than for Acer (5.57 cm) and the difference between species increased when the proportion of Fagus decreased in the local neighborhood.
Annual growth unit-level variables
Most relationships at the annual growth unit-level (GU) were not directly affected by competition indices. For both species, DbaseGU (GU basal diameter) could be fit as a positive linear relationship of D st combined with a negative interaction between H GU/H st and D st (Fig. 7, Table 2; Eq. 4; H GU/H st was not statistically significant). For both species, none of the three competition indices affected DbaseGU.
S GU (sum of branch cross-sectional areas per GU) was a linear function of the difference between SbaseGU and SendGU (cross-sectional area at the base and end of the GU, respectively; Fig. 8; Table 2; Eq. 5). The relationship was unaffected by AgeGU (GU age) or by competition indices in Fagus, but was affected by AgeGU and by HrelGU (GU height relative to the canopy level) in Acer. For a given difference between SbaseGU and SendGU, Fagus exhibited a wider individual variability of SGU than Acer, as shown by the adjusted R 2 of their respective model, resulting in lower correlation quality.
NbrGU (number of branches per GU) varied as a linear function of GU length for both Acer and Fagus (Fig. 4; Table 2; Eq. 6). The slope of the relationship was affected by AgeGU and by Dens for both species: at a given GU length, older GUs or GUs from trees growing in high density plots carried fewer branches. The effect of Dens was less visible in younger GUs. The equations were insensitive to Mix and Hdom. The relationship showed an overall similar shape for both species but Fagus exhibited a higher individual variation in NbrGU at a given GU length with Dens at a given age.

Change in NbrGU (number of branches on the GU) with LGU (GU length). The density indicated for individual tree values is the average density for the five classes in which the density gradient was divided. We also show (a) the simulated density effects on the regression lines for Acer; and b the simulated AgeGU (the age of GU on the stem) effects on the regression lines for Fagus (Eq. 6; Table 2; Sample 3; Table 1)
For second-order branches, D br showed a clear acrotonic pattern (i.e., a pattern at the GU level) for Acer and for Fagus (Table 2; Eq. 7). For both species, D br increased as a cubic function of Hinsbr (branch insertion height). The slope of the relationship decreased for young branches, their acrotony being weaker. Additionally, for both species, D br also depended linearly on DbaseGU as larger GUs carried larger branches. Relationships for both species were unaffected by competition indices. In Fagus, there was a great dispersion of values for large branches resulting in an inferior model quality (Eq. 7; Table 2).
Branch-level variables
All relationships at the branch level were not directly affected by competition indices for both species. Several branch traits could be described at any branch order using D br. Fagus showed a higher variation in L br (branch length) and V br (branch volume) for a given D br than Acer (Figs. 9, 10; Eqs. 8, 10; Table 2). For a given D br, Acer showed on average longer branches than Fagus (Table 2; Eq. 8). This relation was independent of Ordbr (branch order). Branch angle (A br) decreased with D br and increased with Hinsbr (Table 2; Eq. 9). For both species, V br was expressed as a cubic function of D br (Fig. 10; Table 2; Eq. 10). Acer exhibited a slightly steeper slope than Fagus with greater V br for larger branches. Data variability increased for larger branches (Fig. 10) and could not be linked to any measured parameters.
For both species, LfAbr (branch leaf area) was expressed as a power function of S br (branch cross-sectional area) with power coefficients slightly above 1 (Fig. 5; Table 2; Eq. 11). For a given cross-sectional area, Fagus showed a higher leaf area than Acer and a higher variability reflected by the larger data dispersion around the modeled line and the worse goodness of fit. LfAbr was significantly correlated to Ordbr, with higher branches (where the amount of light is smaller) having a smaller LfAbr.

Discussion
In our study of an 8-year-old experimental plantation of Fagus and Acer, we found strong evidence for variation in crown structure in response to density and species identity of the local neighborhood. The variation occurred at alternative crown levels for the two species. This highlights the complexity of species interactions and the need for detailed information on crown development to better understanding species coexistence in mixed broadleaved stands.
Effects of size, density and species identity of neighbors
Even though our study was restricted to a juvenile stage, we observed that for most of the crown traits measured, larger trees exhibited greater variability than smaller trees, as shown by previous results (Mäkelä and Vanninen 1998; Ilomäki et al. 2003). It is likely that the response of crowns to the local neighborhood will vary with stand development, although crowns of older Fagus trees have also demonstrated high plasticity in response to changes in the local canopy conditions (Schröter et al. 2012; Dieler and Pretzsch 2013) suggesting that the response to competition could be sustained over tree life.
Tree density significantly affected crown architecture as reported previously (Lang et al. 2012), whereas neighbor species identity had a significant but less pronounced role despite the large range in species proportions examined. This lends support to the findings of Kaitaniemi and Lintunen (2010), who found that the effect of density (in a more limited range than in our study) and neighbor size (calculated as a competition index) alone were more important than the effect of neighbor identity when exploring the effect of Larix sibirica Ledeb. and Pinus sylvestris L. on Betula pendula Roth. Overall, our results are consistent with previous findings, where information on the amount, size and species of neighboring trees was necessary to adequately describe tree crown architecture and branching arrangement (Sumida et al. 2002; Kaitaniemi and Lintunen 2008; Lintunen and Kaitaniemi 2010).
Models at the stem level were generally improved directly by the competition indices accounting for density (Dens) and species proportion (Mix). However, at the GU and branch levels, most crown variables were not directly affected by competition, and could be mostly predicted through allometry, once the effect on some key covariates was accounted for. Most of the influence of the variations in density, neighbor identity and neighbor size were accounted for by the stem-level variables for both species and, for branch-level variables, secondarily by GU-level variables. One exception was branch number, which was affected by density and not by species identity as reported by Kaitaniemi and Lintunen (2010). In general, we found that allometry between crown attributes was less variable than the differences in allometry between stem diameter and crown attributes, which is in agreement with the findings on tree allometry in Scots pine by Mäkelä and Vanninen (1998). Similarly, several studies found that stem and crown properties were sufficient to predict reliable branch properties under contrasted competitive local environments (Hein et al. 2007; Weiskittel et al. 2007; Weiskittel et al. 2010; Antin et al. 2013).
Acer and Fagus respond to neighborhood competition at alternative crown levels
Despite common shapes in the relationships among all the variables measured at the three levels considered (i.e., stem, annual growth unit, and branch) for Acer and Fagus, the two species reacted differently to variation in their local neighborhoods. Crown attributes were modified at different levels for the two species. Species-specific effects of neighbor competition on height–diameter relationships and crown allometries were expected from previous neighborhood analysis (Coates et al. 2009; Harja et al. 2012). Acer displayed higher intraspecific plasticity of crown length to changes in density, but both Acer and Fagus showed a similar plasticity of crown length to the neighboring species proportion. In our study, changes in the light environment are mostly a result of changes in density (results not shown). Given this, our results agree with the findings of (Petriţan et al. 2009) in an observational study of juvenile trees under varying light conditions beneath a shelterwood canopy in a mixed deciduous stand, where Acer showed a greater plasticity in crown shape than Fagus. In our experiment, the latter displayed a higher level of individual variability on branch development (i.e., branch diameter vs. volume, Fig. 10; difference of branch cross-sectional area at start and end vs. sum of branch cross-sectional area, Fig. 8) and on leaf area variation (Fig. 5) in response to changes in density and species proportion in its neighborhood. Although both species are defined as having similar resource use strategies (Collet et al. 2014), the low degree of modification of the crown length with increasing shade in Fagus, together with the high degree of leaf area variability, indicates a greater ability of Fagus to maintain the height they acquired in periods of high light availability (Collet et al. 2011), and to keep branches alive under shaded conditions, thus resisting to very low light environments.
Implications for predicting stand development in mixed-species forests
Our results suggest that responses of crown morphology should be inferred from a hierarchical model including variables at distinct scales (Fig. 6) rather than modular units that are autonomous with respect to their responses to local conditions (Kawamura 2010). This could provide a more mechanistic understanding of how different strategies of crown development contribute to species coexistence in mixed stands. Our models provide a steady-state calculation, and as such they are not able to characterize temporal dynamics, including the growth of the studied crown attributes. However, the static relationships among attributes provide a useful foundation for the dynamic analysis, by identifying the key attributes on which to focus, and by simplifying the relationships to study. Our static approach can also contribute to the understanding of the various development constraints (biomechanical, physiological, etc.) that eventually shape crown morphology (Fournier et al. 2013). Moreover, this approach provides the basis for a procedure to reconstruct crown geometry from a set of attributes that are relatively easy to measure in the stands, and that can afterwards be used as an input to growth models or light interception models (Lintunen et al. 2011). Using the relationships we established, we could simulate vertical leaf distribution within a mixed stand to test hypotheses, such as if potential complementarity in the leaf area display between the two species could explain the higher productivity found in the mixing where the proportion of both Fagus and Acer in the stand is 50 % (Collet et al. 2014).

Scheme of the scaling relationships among the variables measured for both species at the stem, GU, and branch levels, as affected by the competition indices: density (Dens), mixing (Mix) and dominant height (Hdom). An arrow connecting two variables indicates a significant relationship between both variables for both species (Table 2) If the effect is significant for only one species, the arrow connecting two variables is dotted
In mixed deciduous forests of temperate regions, shade tolerance is a trait that has been commonly reported to explain differences in crown plasticity (Ameztegui and Coll 2011). Our results indicate that in mixed-species stands formed by functionally equivalent species, with similar shade tolerance and similar total tree height trends, species coexistence may be partly explained by differences in species architecture, such as previously identified for co-occurring broadleaved species of the genera Fagus, Tilia, Acer, Fraxinus and Carpinus (Frech et al. 2003). Furthermore, our results are in line with other experiences of shade-intolerant species in mountain forests (Parish et al. 2008; Vieilledent et al. 2010), highlighting the importance of intraspecific differences in individual variability as a potential mechanism for the long-term persistence of mixed deciduous broadleaved forests.
Author contributions
F.N. and C.C. designed the sampling scheme and the measurement protocol. I.B and C.C. wrote code, ran the models, and analyzed output data. I.B, C.C. and F.N. wrote the manuscript.
References
Ameztegui A, Coll L (2011) Tree dynamics and co-existence in the montane–sub-alpine ecotone: the role of different light-induced strategies. J Veg Sci 22:1049–1061. doi:10.1111/j.1654-1103.2011.01316.x
Antin C, Pélissier R, Vincent G, Couteron P (2013) Crown allometries are less responsive than stem allometry to tree size and habitat variations in an Indian monsoon forest. Trees 27:1485–1495. doi:10.1007/s00468-013-0896-7
Barthélémy D, Caraglio Y (2007) Plant architecture: a dynamic, multilevel and comprehensive approach to plant form, structure and ontogeny. Ann Bot 99:375–407. doi:10.1093/aob/mcl260
Bell AD (1991) Plant form: an illustrated guide to flowering plant morphology. Oxford University Press, Oxford
Coates KD, Canham CD, LePage PT (2009) Above- versus below-ground competitive effects and responses of a guild of temperate tree species. J Ecol 97:118–130
Collet C, Fournier M, Ningre F et al (2011) Growth and posture control strategies in Fagus sylvatica and Acer pseudoplatanus saplings in response to canopy disturbance. Ann Bot 107:1345–1353. doi:10.1093/aob/mcr058
Collet C, Ningre F, Barbeito I et al (2014) Response of tree growth and species coexistence to density and species evenness in a young forest plantation with two competing species. Ann Bot 113:711–719. doi:10.1093/aob/mct285
Davies O, Pommerening A (2008) The contribution of structural indices to the modelling of Sitka spruce (Picea sitchensis) and birch (Betula spp.) crowns. For Ecol Manag 256:68–77. doi:10.1016/j.foreco.2008.03.052
Deleuze C, Herve J-C, Colin F, Ribeyrolles L (1996) Modelling crown shape of Picea abies : spacing effects. Can J For Res 26:1957–1966
Dieler J, Pretzsch H (2013) Morphological plasticity of European beech (Fagus sylvatica L.) in pure and mixed-species stands. For Ecol Manag 295:97–108. doi:10.1016/j.foreco.2012.12.049
Fournier M, Dlouhá J, Jaouen G, Almeras T (2013) Integrative biomechanics for tree ecology: beyond wood density and strength. J Exp Bot ert279. doi: 10.1093/jxb/ert279
Frech A, Leuschner C, Hagemeier M, Holscher D (2003) Neighbor-dependent canopy dimensions of ash, hornbeam, and lime in a species-rich mixed forest (Hainich National Park, Thuringia). Forstwissenchaftlisches Cent 122:22–35
Harja D, Vincent G, Mulia R, Noordwijk M (2012) Tree shape plasticity in relation to crown exposure. Trees 26:1275–1285. doi:10.1007/s00468-012-0703-x
Hein S, Mäkinen H, Yue C, Kohnle U (2007) Modelling branch characteristics of Norway spruce from wide spacings in Germany. For Ecol Manag 242:155–164. doi:10.1016/j.foreco.2007.01.014
Hein S, Collet C, Ammer C et al (2009) A review of growth and stand dynamics of Acer pseudoplatanus L. in Europe: implications for silviculture. Forestry 82:361–385. doi:10.1093/forestry/cpn043
Huang S, Meng SX, Yang Y (2009) Assessing the goodness of fit of forest models estimated by nonlinear mixed-model methods. Can J For Res 39:2418–2436
Ilomäki S, Nikinmaa E, Mäkelä A (2003) Crown rise due to competition drives biomass allocation in silver birch. Can J For Res 33:2395–2404. doi:10.1139/x03-164
Kaitaniemi P, Lintunen A (2008) Precision of allometric scaling equations for trees can be improved by including the effect of ecological interactions. Trees 22:579–584. doi:10.1007/s00468-008-0218-7
Kaitaniemi P, Lintunen A (2010) Neighbor identity and competition influence tree growth in Scots pine, Siberian larch, and silver birch. Ann For Sci 67:604. doi:10.1051/forest/2010017
Kawamura K (2010) A conceptual framework for the study of modular responses to local environmental heterogeneity within the plant crown and a review of related concepts. Ecol Res 25:733–744. doi:10.1007/s11284-009-0688-0
Lang AC, Härdtle W, Baruffol M et al (2012) Mechanisms promoting tree species co-existence: experimental evidence with saplings of subtropical forest ecosystems of China. J Veg Sci 23:837–846. doi:10.1111/j.1654-1103.2012.01403.x
Lines ER, Zavala MA, Purves DW, Coomes DA (2012) Predictable changes in aboveground allometry of trees along gradients of temperature, aridity and competition. Glob Ecol Biogeogr 21:1017–1028. doi:10.1111/j.1466-8238.2011.00746.x
Lintunen A, Kaitaniemi P (2010) Responses of crown architecture in Betula pendula to competition are dependent on the species of neighbouring trees. Trees 24:411–424. doi:10.1007/s00468-010-0409-x
Lintunen A, Sievänen R, Kaitaniemi P, Perttunen J (2011) Models of 3D crown structure for Scots pine (Pinus sylvestris) and silver birch (Betula pendula) grown in mixed forest. Can J For Res 41:1779–1794. doi:10.1139/x11-092
Mäkelä A, Vanninen P (1998) Impacts of size and competition on tree form and distribution of aboveground biomass in Scots pine. Can J For Res 28:216–227. doi:10.1139/x97-199
Mäkinen H, Colin F (1999) Predicting the number, death, and self-pruning of branches in Scots pine. Can J For Res 29:1225–1236. doi:10.1139/x99-065
Medhurst JL, Beadle CL (2001) Crown structure and leaf area index development in thinned and unthinned Eucalyptus nitens plantations. Tree Physiol 21:989–999. doi:10.1093/treephys/21.12-13.989
Muukkonen P (2007) Generalized allometric volume and biomass equations for some tree species in Europe. Eur J For Res 126:157–166. doi:10.1007/s10342-007-0168-4
Näslund M (1937) Skogsförsöksanstaltens gallringsförsök i tallskog. Medd Stat Skogsförsöksanst 29:1–169 (in Swedish)
Nicolini E, Chanson B, Bonne F (2001) Stem growth and epicormic branch formation in understorey beech trees (Fagus sylvatica L.). Ann Bot 87:737–750. doi:10.1006/anbo.2001.1398
Osada N (2006) Crown development in a pioneer tree, Rhus trichocarpa, in relation to the structure and growth of individual branches. New Phytol 172:667–678. doi:10.1111/j.1469-8137.2006.01857.x
Parish R, Nigh GD, Antos JA (2008) Allometry and size structure of trees in two ancient snow forests in coastal British Columbia. Can J For Res 38:278–288. doi:10.1139/X07-172
Petriţan AM, von Lüpke B, Petriţan IC (2009) Influence of light availability on growth, leaf morphology and plant architecture of beech (Fagus sylvatica L.), maple (Acer pseudoplatanus L.) and ash (Fraxinus excelsior L.) saplings. Eur J For Res 128:61–74. doi:10.1007/s10342-008-0239-1
Pretzsch H, Dieler J (2012) Evidence of variant intra- and interspecific scaling of tree crown structure and relevance for allometric theory. Oecologia 169:637–649. doi:10.1007/s00442-011-2240-5
Purves DW, Lichstein JW, Pacala SW (2007) Crown plasticity and competition for canopy space: a new spatially implicit model parameterized for 250 North American tree species. PLoS One 2:e870. doi:10.1371/journal.pone.0000870
Rouvinen S, Kuuluvainen T (1997) Structure and asymmetry of tree crowns in relation to local competition in a natural mature Scots pine forest. Can J For Res 27:890–902. doi:10.1139/x97-012
Schneider CA, Rasband WS, Eliceiri KW (2012) NIH image to image J: 25 years of image analysis. Nat Methods 9:671–675. doi:10.1038/nmeth.2089
Schröter M, Härdtle W, von Oheimb G (2012) Crown plasticity and neighborhood interactions of European beech (Fagus sylvatica L.) in an old-growth forest. Eur J For Res 131:787–798. doi:10.1007/s10342-011-0552-y
Sumida A, Terazawa I, Togashi A, Komiyama A (2002) Spatial arrangement of branches in relation to slope and neighbourhood competition. Ann Bot 89:301–310. doi:10.1093/aob/mcf042
Thorpe HC, Astrup R, Trowbridge A, Coates KD (2010) Competition and tree crowns: a neighborhood analysis of three boreal tree species. For Ecol Manag 259:1586–1596. doi:10.1016/j.foreco.2010.01.035
Vanclay JK (2006) Experiment designs to evaluate inter- and intra-specific interactions in mixed plantings of forest trees. For Ecol Manag 233:366–374. doi:10.1016/j.foreco.2006.05.034
Vieilledent G, Courbaud B, Kunstler G et al (2010) Individual variability in tree allometry determines light resource allocation in forest ecosystems: a hierarchical Bayesian approach. Oecologia 163:759–773. doi:10.1007/s00442-010-1581-9
Wagner S, Collet C, Madsen P et al (2010) Beech regeneration research: from ecological to silvicultural aspects. For Ecol Manag 259:2172–2182. doi:10.1016/j.foreco.2010.02.029
Weiskittel AR (2003) Alterations in Douglas-fir crown structure, morphology, and dynamics imposed by the Swiss needle cast disease in the Oregon Coast Range. Thesis
Weiskittel AR, Maguire D, Monserud RA (2007) Response of branch growth and mortality to silvicultural treatments in coastal Douglas-fir plantations: implications for predicting tree growth
Weiskittel AR, Seymour RS, Hofmeyer PV, Kershaw JA Jr (2010) Modelling primary branch frequency and size for five conifer species in Maine, USA. For Ecol Manag 259:1912–1921. doi:10.1016/j.foreco.2010.01.052
Zuur AF, Ieno EN, Walker N et al (2009) Mixed Effects Models and Extensions in Ecology with R, vol 1. Springer, Berlin
Acknowledgments
We thank C. Richter and A. Piboule (Office National des Forêts) for designing and installing the experimental site. Funding was provided by Region Lorraine (project “Jeune Equipe 2005”), Office National des Forêts (“ModelFor” project) and INRA (project “Forêts mélangées” from the ECOGer programme). The UMR 1092 LERFoB is supported by a grant overseen by the French National Research Agency (ANR) as part of the “Investissements d’Avenir” program (ANR-11-LABX-0002-01, Lab of Excellence ARBRE).
Conflict of interest
The authors declare that they have no conflict of interest.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Matyssek.
Appendices
Appendices


Relationship between S brGU (the sum of cross-sectional areas at the base of each GU) and the cross-sectional areas at GU base (SbaseGU) and GU distal end (SendGU) for both species (Eq. 5; Table 2; Sample 3; Table 1). Note that a single trendline is shown for both species because they are almost identical


Rights and permissions
About this article

Cite this article
Barbeito, I., Collet, C. & Ningre, F. Crown responses to neighbor density and species identity in a young mixed deciduous stand. Trees 28, 1751–1765 (2014). https://doi.org/10.1007/s00468-014-1082-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-014-1082-2