
Abstract
Key message
Earlywood vessel features indicate different adaptations of Quercus petraea and Q. pyrenaica , which are probably related with their corresponding Atlantic and sub-Mediterranean ecological requirements.
Abstract
We studied the climatic signal of the earlywood anatomy of a temperate [Quercus petraea (Mattuschka) Liebl.] and a sub-Mediterranean (Quercus pyrenaica Willd.) oak species growing under similar climatic conditions in a transitional area between the Atlantic and Mediterranean regions of the Iberian Peninsula. We hypothesized that both species react differently in their wood anatomy due to their contrasting ecological requirements, and we test the usefulness of earlywood anatomical features to study the behaviour of these ring-porous oaks upon climate. For this, we measured the earlywood vessels, and obtained annual series of several anatomical variables for the period 1937–2006 using dendrochronological techniques, considering whether the vessels belonged to the first row or not. After optimizing the data set by principal component analysis and progressive filtering of large vessels, we selected maximum vessel area and total number of vessels as they resulted to be the optimal variables to describe vessel size and number, respectively. Vessel size of Q. pyrenaica was dependent on precipitation along the previous growing season, whereas it did not show any clear climatic response for Q. petraea. On the contrary, vessel number was related to winter temperature for both species. These relationships observed between climate and anatomy appeared to be stable through time. The results obtained reinforce the utility of earlywood vessel features as potential climate proxies.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Climate change has been recently affecting both ecosystems and the organisms that inhabit them (Parmesan 2006). Current environmental changes occur at a fast rate, which is imposing several physiological limitations to the capacity of trees to cope with new climatic conditions (Alcamo et al. 2007), and only plants able to take advantage of a warmer and longer growing season will probably dominate in the future (Pop et al. 2000). Phenotypical plasticity can be an immediate mechanism of adaptation if the stressful conditions lie within a species tolerance range (Valladares et al. 2007). On the contrary, long-term changes in the environment through many generations can force evolutionary changes within the populations. But when climatic changes occur at a higher intensity or a faster rate than the capacity of tree populations to adapt or evolve, such species need to migrate to survive (Gian-Reto et al. 2002). Tree species are expected to move up to a higher altitude at a rate of 8–10 m per decade (Grabherr et al. 1994), and many species limited to mountain tops could even become extinct (Bradley et al. 1999). Such species displacements have been estimated at different scales and regions, mostly through climate models (Hansen et al. 2001; Thuiller et al. 2005), which clearly predict a high turnover in many tree species and ecosystem composition. In addition, disturbance by human activities lead to fragmentation of forest ecosystems, which should also increase the risk of species extinctions, especially in marginal areas.
One of the most important adaptive strategies of trees to cope with this future scenario would be to react through their xylem architecture, gaining a compromise between conductive efficiency (optimal hydraulic flow), safety (embolism resistance), an optimal water economy (photosynthesis/transpiration balance), and reserve support for dormancy and growth resumption (assimilation/respiration rates) (Martínez-Vilalta et al. 2002; Mencuccini et al. 2010). Maximum hydraulic conductivity and vulnerability to cavitation determine the overall transport efficiency of the xylem to a great extent (Zanne et al. 2010). Conductivity directly depends on the size of the conducting elements, while embolisms are believed to spread along conduits as a function of the maximum size of the pores in the inter-conduit pit membrane (Hacke and Sperry 2001). Although pore size has often been hypothesized to be associated with the diameter of the conduit, other evidences point out that the relationship between conduit diameter and vulnerability to cavitation by water stress greatly varies (Hacke et al. 2006). The ‘rare pit hypothesis’ has been recently proposed to explain air-seeding cavitation in ring-porous trees, but Christman et al. (2012) state that even under this hypothesis vessel size-vulnerability trade-off is complex for large vessels, and probably highly variable across species. However, plants can use different strategies to react to drought stress, because an efficient conducting system may lead to hydraulic failure, whereas drought avoidance could result in carbon starvation (McDowell et al. 2008).
Water conductivity is strongly related to the distribution of the conducting cells within the annual tree rings, mainly vessels or tracheids. Thus, tree-ring anatomy, i.e. the measurement of anatomical features throughout continuous sequences of dated annual rings, seems to be a useful methodological approach for the evaluation of the impact of the predicted climate change on future vegetation dynamics and distributions, based on both current climate and the ecophysiological behaviour of trees (Fonti et al. 2010). Though sometimes focused on tracheid lumen size of conifers (e.g. Eilmann et al. 2006; Bryukhanova and Fonti 2013), or vessels of diffuse-porous trees (Sass and Eckstein 1995; Schume et al. 2004), this tool has been especially successful for the earlywood vessels of ring-porous species such as oak (García-González and Eckstein 2003; Fonti and García-González 2008; Gea-Izquierdo et al. 2012) or chestnut (Fonti et al. 2007), which are clearly related to tree performance.
Ring-porous deciduous oaks such as Q. robur L. and Q. petraea Mattuschka (Liebl.) are dominant in many forest ecosystems throughout Europe, and reach their distribution boundary towards the Mediterranean region. Within the Iberian Peninsula, they prevail at their southwestern limit in ecosystems of the Atlantic region, but are first coexisting and then progressively replaced by stands of marcescent oaks (mainly Q. pyrenaica Willd. but also Q. faginea Lam. or Q. pubescens Willd.) as Mediterranean conditions, i.e. a stronger water deficit in summer, increase (Sánchez-DeDios et al. 2009). But ongoing warming and reduced rainfall are particularly expected to affect the vegetation of the Iberian Peninsula (Hernández-Santana et al. 2009), where numerous species reach their limit of distribution, so that their populations become very susceptible to changes. Thus, the Iberian Peninsula is an adequate territory for studying the potential changes in the physiological and functional responses of existing forests that might result from climate change (Sánchez-DeDios et al. 2009). Specifically, northern regions of Spain occupy a key biogeographical position to understand the behaviour of species to cope with environmental changes, due to their transitional position between the Atlantic and the Mediterranean regions. Nemoral oaks such as Q. petraea could consequently be replaced by sub-Mediterranean oaks as Q. pyrenaica, and these temperate broad-leaved species could become relegated to some enclaves within the northern mountains of the Iberian Peninsula if climate becomes warmer (Benito-Garzón et al. 2008).
In this paper, we hypothesize that Q. petraea and Q. pyrenaica growing under similar climatic conditions in a transitional area between the Atlantic and Mediterranean regions of the Iberian Peninsula react differently in their wood anatomy, as a result of their contrasting ecological requirements. Our specific aims are (1) to analyse whether these differences are recorded in their earlywood vessel features; and (2) to evaluate the usefulness of these features to study the behaviour of both ring-porous oaks upon climate at their distribution boundary.
Materials and methods
Study sites and study species
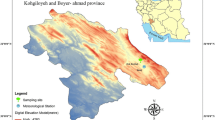
The study site is located within a semi-natural woodland area of about 11,250 ha of deciduous forest in Monte Hijedo Natural Park, in the southern part of the Cantabrian Mountains, Northern Iberian Peninsula (Fig. 1). Mean monthly temperature reaches 18 °C in the hottest month (August), 2.5 °C in the coldest (January), and mean annual precipitation is 789 mm. Rainfall suffers variation along the year, with a maximum during winter, and soil water reserves can be seasonally scarce, although a period of a remarkable summer drought, which is typical of Mediterranean climate, is absent. The area lies at the boundary between the Atlantic and Mediterranean bioclimatic regions (Sánchez-DeDios et al. 2009).

Location of the study area and climatic diagram for the period 1937–2006
Both study species grew at different locations, but only 400–500 m away from each other. Soils developed on sandstone, and were similar at both sites, although water-holding capacity was probably slightly higher for Q. petraea, due to most humid conditions and therefore a higher content of organic matter in the upper horizon. Q. petraea was sampled on a moderate slope with western exposure (42º54′9.4″N, 03º57′7.9″W; 900 m a.s.l.), forming a mixed stand with Fagus sylvatica L., and accompanied by Corylus avellana L., Ilex aquifolium L., Crataegus monogyna Jacq., and Taxus baccata L. Other species present at his site were indicative of a shady woodland, such as Vaccinium myrtillus L., Daphne laureola L., Deschampsia flexuosa (L.) Trin., Anemone nemorosa L., Viola riviniana Rchb., or abundant ferns (e.g. Dryopteris dilatata Hoffm., Dryopteris affinis (Lowe), Polystichum setiferum Forssf. or Blechnum spicant L.). By contrast, Q. pyrenaica formed a monospecific woodland in weak eastern exposure and on a similar slope (42º53′39.1″N, 03º56′1.6″W; 822 m a.s.l.), with some scattered, young individuals of Fagus sylvatica, Acer campestre L. or Ilex aquifolium. Shrubs were scarce, with occasional presence of Erica vagans L., Ruscus aculeatus L. and Rubus sp. Herbaceous species typical of moister environments hardly occurred, but still some were found (Anemone nemorosa, Primula veris L.), whereas ferns were nearly absent.
Tree sampling and wood preparation
For each species, two increment cores were extracted at breast height from 15 dominant trees. The cores were air-dried, and mounted on wooden supports to cut and polish a cross-sectional surface. The cut surface was sanded with progressively finer sandpaper (grain sizes from 280 to 1,200). Vessel lumina were cleaned with high-pressure water blast to remove both tyloses and wood dust. The wooden surface was coloured black with printer ink, and the vessels were filled with white chalk to make them clearly visible due to the contrast between the lumina and the wood matrix.
Variable survey and automatic image analysis
A tree-ring measuring device linear stage (Velmex TA UniSlide, Velmex Inc. Bloomfield NY, USA) combined with a binocular microscope was used to measure earlywood (EW) and latewood width (LW). The boundary between earlywood and latewood was distinguished by vessel size (much larger in earlywood than in latewood) and distribution (latewood dominated by fibres, with abundant small vessels forming flame-like radial groups). Total tree-ring width (RW) was calculated as the sum of EW and LW. The measurements were visually cross-dated to assign each ring to the corresponding calendar year, and we finally used the COFECHA software (Grissino-Mayer 2001) to statistically verify the tree-ring dating accuracy.
We selected 10 out of the 15 sampled trees to carry out vessel measurements. Among the trees with series covering the common period 1937–2006, we considered those with an optimal vessel appearance, as well as trees without conspicuous growth reductions, which would facilitate image analysis. Images were captured for each ring using a digital camera (NIKON DMX 1200 F) attached to a binocular microscope. The image resolution was 3,200 × 2,560 pixels, with ring boundaries aligned throughout the largest dimension, and each ring was cropped from the original image and saved into a new TIFF file.
Earlywood vessels were detected and measured using ImageJ software (Rasband 2009), which facilitates automatic object recognition in digital images with grey scales ranging from 0 to 255. Vessel lumen area (white) was distinguished from the background tissue (black) on a threshold grey level, which was occasionally adjusted during measurements. However, this process was not enough to correctly recognize all vessels, because of the presence of rays, dust spots, and other features also detected by the system. Therefore, two filters, a size filter and a shape filter, were applied to detect an object as a vessel. The minimum value of the size filter was set at 7,500 μm2 (diameter 120 μm), while the shape filter excluded objects whose length was more than twice their width. Manual corrections of the image to properly identify all vessels were performed when some objects could not be excluded by the filters.
Once all vessels had been correctly recognized, we visually selected a subset of earlywood vessels belonging to the first row, as they often lead to more successful results when comparing to climate (García-González and Fonti 2008). This first row was considered to include all vessels that were located immediately after the ring boundary, and also those whose beginning did not surpass the middle line of these vessels. All other vessels were referred to ‘not belonging to the first row’, as further rows were absent in some rings, or they were not always clearly arranged tangentially, especially in the transition to latewood and in wide rings.
Chronologies of growth variables
A huge set of earlywood variables was initially calculated from the data obtained, with the aim of optimizing it for subsequent analyses. The variables considered for every ring were total number of vessels (NV), mean vessel lumen area (MVA) for all measured vessels, mean vessel lumen area of vessels larger than the distribution median (M50) for each ring, maximum vessel lumen area (MAX), total vessel lumen area (TVA), and water conductivity (KS), which was estimated as the fourth power of the radius of every vessel of the ring (Zimmermann 1983). NV, TVA, and KS were standardized to a frame of 10 mm of tangential width. For each variable, we considered three values: for all earlywood vessels, only for those belonging to the first row (r1), and for those not belonging to that row (nr1). However, NV in the first row was discarded, as it was nearly constant among rings. Three further variables were considered relating the average of vessels not belonging to the first row to those in the whole earlywood: number of vessels (NV%nr1), conductive area (TVA%nr1) and water conductivity (KS%nr1). In addition to anatomical variables, total ring width (RW), earlywood width (EW), and latewood width (LW) were also included in the analyses. This yielded a total set of 23 variables per species.
Growth-related trends were removed from the series because they could lead to a misrepresentation of the climatic signal. For this, we filtered all series by a cubic smoothing spline function (Cook 1992) with 32-year stiffness and a 50 % cutoff, mainly retaining year-to-year variability. The same method was applied to all variables to avoid differences associated with detrending procedures. After detrending, the obtained time series of growth indices were averaged by a biweight robust mean into a chronology for each variable.
Individual series and chronologies were characterized by standard descriptive statistics, also including the mean sensitivity (MS) and the first-order autocorrelation (AR1). The statistical quality of each single chronology was evaluated using classical dendrochronological parameters (Briffa and Jones 1992), such as the mean correlation between trees (Rbt), the percentage of variance in the first eigenvector (PC1), the expressed population signal (EPS), and the signal-to-noise ratio (SNR). The period for most analyses was 1937–2006, because it was covered by data from almost all study trees and climatic time series, and also to avoid the presence of juvenile wood in some individuals.
Comparison among chronologies
As we initially considered a high number of anatomical and growth variables (23), many of them were expected to be related to each other. Therefore, we identified groups of variables yielding similar information to reduce their number for further analyses. This was performed by principal component analysis (PCA), which proved to be successful at identifying groups in time series of anatomical variables (Fonti and García-González 2004; Bryukhanova and Fonti 2013). For each species, we applied PCA on the correlation matrix of all chronologies, and identified groups using the loadings on the two first principal components, after following a varimax rotation. These analyses were carried out in R Core Development Team (2012).
In addition to the vessel size variables considered (MVA, M50, and MAX), we checked the variations in the common and climatic signal of vessel size by selecting a subset of vessels, and therefore verified if the selection of the largest vessels, or vessels formed at different times, improved the climatic signal (García-González and Fonti 2006). For this, a progressive filter was applied to gradually exclude small vessels from the original dataset to track changes in the signal expressed at each step. This procedure was performed by retaining only the n-percent of the largest vessels, and progressively removing the lowest percentile of earlywood vessels. For each data subset, individual MVA time series were established, and therefore the statistical analyses had to be recalculated for them.
Relationships to climate
Relationships between climatic series and the indexed chronologies of ring widths and anatomical variables were established by means of correlation functions (Pearson’s correlation coefficient). Significance level of the correlation coefficients was determined using a bootstrap approach; we computed 10,000 iterations for each coefficient, and applied the correction proposed by Mason and Mimmack (1992) for the percentile bootstrap confidence intervals. For this, we used a routine written in Embarcadero Delphi® XE2.
We preliminarily computed these relationships for all anatomical variables considered; in addition, the analyses had to be recalculated for each MVA subset following the selection filter. However, the results focus only on those variables yielding the most reliable responses to climate, due to the huge amount of data generated. The period for these analyses was also 1937–2006 (70 years).
Climatic data came from regional records obtained by averaging data from three nearby stations (Reinosa, 42º59′N, 4º09′W, 870 m a.s.l.; Polientes, 42º48 N, 3º56 W, 720 m a.s.l; and Cervera de Pisuerga, 42º52 N, 4º31 W, 1,140 m a.s.l.). The series were obtained by means of the routine MET in the Dendrochronology Program Library software (Grissino-Mayer 2008).
We took monthly values of mean temperature and total precipitation, from previous June to current May for the earlywood variables, and from previous August to current October for latewood and total ring width. We also averaged consecutive months with similar climate–growth relationships, as the dormancy-quiescence period or the previous summer, to verify if the climatic responses were improved as compared to those obtained when single months were considered. Finally, the temporal variation of climate–growth relationships was analysed with moving correlation functions (Biondi 1997), considering periods of 40 years shifted by 1 year to each other. For this procedure, we used the longest possible period of the series that could be reliable for the analyses, i.e. 1927–2006 for Q. petraea (8 trees) and 1935–2006 for Q. pyrenaica (7 trees).
Results
An average of 2,386 vessels for Q. petraea, and 2,546 for Q. pyrenaica were measured in each tree (Table 1); about a half of the vessels (48.5 %) belonged to the first row in Q. petraea, and less (39.0 %) in Q. pyrenaica. Mean vessel area was very similar for both species (40,620 μm2 for Q. petraea; 40,095 μm2 for Q. pyrenaica). The distribution frequency of all measured lumina showed that small vessels were much more abundant for both species (skewness was 0.37 for Q. petraea; 0.51 for Q. pyrenaica), and even more for those not located in the first row (0.79 for Q. petraea; 0.64 for Q. pyrenaica). Skewness was almost zero for the first row of earlywood vessels, highlighting that vessels are more homogeneous at this position within the tree ring.
PCA identified three groups of variables, the results being similar for both species (Fig. 2). One group comprised variables related to vessel size for all measured vessels (MAX, MVA, and M50); and all variables derived from the first row (r1), regardless of being associated with vessel size or not. A second group included NV, TVA, EW, variables related to the average of vessels not belonging to the first row (%nr1), those not located in the first row of earlywood vessels (nr1), and intermediate variables as KS, which is determined by both size and number of vessels. A third group comprised only two growth variables including latewood width (RW and LW), and was closer to the second group for Q. pyrenaica than for Q. robur. In summary, PCA analysis indicated that there were two distinct groups of anatomical variables determined either by number or size, which recorded a different kind of environmental information. NV appeared to be the most appropriate variable among those defining the numbers, while a subsequent analysis was necessary for size variables.

Principal component analysis (PCA) of the correlation matrix of all chronologies, after extracting the two first PCs following varimax rotation. Series were analysed along the period 1937–2006 (70 years). RW total ring width, LW latewood width, EW earlywood width, MVA mean vessel area, M50 mean vessel area using 50 % of the vessels larger than the median, MAX maximum vessel area, TVA total vessel area, KS water conductivity, NV number of vessels. Anatomical variables refer to all vessels when no indication is provided, otherwise vessels belong to the first row (r1), those not located in the first row (nr1), or the ratio between vessels not in the first row and all vessels (%nr1)
Within the groups identified by PCA, correlations with climate were similar to each other (Table 2), although some variables appear to optimize the signal. We found very weak climatic responses for RW and LW both in Q. petraea and Q. pyrenaica, with only significant correlations (P < 0.05 for Q. petraea and up to P < 0.01 for Q. pyrenaica) for previous winter and April precipitation, and also to winter temperature for Q. pyrenaica. However, RW had the best chronology statistics among all studied variables, with higher values of Rbt (0.46 for Q. petraea; 0.52 for Q. pyrenaica) and EPS (0.89 for both species), as well as the highest values of year-to-year variability, expressed by MS (0.16–0.18). AR1 was low for RW and LW (0.08–0.29) as compared to those reached in some anatomical variables (0.08–0.52).
Vessel size variables did not strongly respond to precipitation for Q. petraea, with only significant correlations (P < 0.05) for previous August or October and current March; such result was even observed after filtering out the smallest vessels. On the contrary, vessel size of Q. pyrenaica was clearly positively correlated with summer rainfall of the previous growing season (Table 2), bearing significant correlations for all size variables, so that a subsequent analysis was performed to select the optimal anatomical variables related to vessel size for their relationships to climate; this method was not applied for Q. petraea due to the lack of a clear response in its vessel size. Pearson’s correlation coefficient between meteorological time series and mean vessel size after each step of the n-percent filter (i.e. removing an increasing percentage of the smallest vessels but retaining the largest ones) increased steadily (Fig. 3a), and the maximum correlation (r = 0.52; P < 0.0001) was reached when only the largest vessel was kept, which corresponds to the variable MAX. This clearly indicates that MAX maximized the climatic signal in comparison to the mean area of the majority of vessels. However, common signal (Rbt) was optimal when ca. 75 % of the smallest vessels had been removed, and it drastically decreased afterwards, even when MAX was the variable considered (Fig. 3b). Furthermore, MAX was highly related to climate in Q. pyrenaica despite having a low common signal (Fig. 4b, c).

Evolution of the climatic and common signal of mean vessel area (MVA) of Quercus pyrenaica when progressively removing small vessels in each ring, respectively, expressed as, a bootstrapped Pearson’s correlation coefficient to previous summer precipitation (June–September), and b mean correlation between trees (Rbt). Dot size in (a) is proportional to the significance level (P < 0.05, P < 0.01, P < 0.001, or P < 0.0001) of the bootstrapped correlation coefficient

Climate–growth relationships for vessel size, expressed as maximum vessel area (MAX). Bootstrapped correlation functions for, a Q. petraea, and b Q. pyrenaica; c adjustment between MAX chronology of Q. pyrenaica and precipitation during the previous summer (June–September). Statistics of chronology quality for MAX are indicated in (a) and (b): mean correlation (Rbt), percentage of variance (PC1), expressed population signal (EPS), signal-to-noise ratio (SNR), mean sensitivity (MS), and first-order autocorrelation (AR1). All correlations and statistics are for the common period 1937–2006
Variables dependent on vessel number showed a strong climatic signal both for Q. petraea and Q. pyrenaica, with slight differences among them. Therefore, this response was observed for most of the variables included in the third group of the PCA analysis, but mainly for TVA, EW, and variables expressing the average number of vessels not belonging to the first row (%nr1); but also for variables not located in the first row of earlywood vessels (nr1), and intermediate variables as KS, although with lower significant levels for the latter. This result can be easily summarized in the response of NV.
Thus, NV was mainly negatively related to temperature during the dormancy-quiescence period, i.e. December through March (r = –0.49 for Q. petraea; r = –0.46 for Q. pyrenaica, P < 0.0001 in both cases; Table 2; Fig. 5). The highest correlation for these single months was found with February, but its value was still lower than considering a longer time span (December–March). But despite the strong correlations found for both species with climate, chronology quality was rather low for NV. Values of Rbt and EPS were clearly lower as compared to those reached by width variables (Fig. 5a, b); MS was also low, but contrary to MAX, values for AR1 were high.

Climate–growth relationships for the total number of vessels (NV). Bootstrapped correlation functions for, a Q. petraea, and b Q. pyrenaica; adjustment between the average chronology of NV and mean temperature during the dormant period (December–March) for, c Q. petraea, and d Q. pyrenaica. Statistics of chronology quality for NV are indicated in (a) and (b): mean correlation (Rbt), percentage of variance (PC1), expressed population signal (EPS), signal-to-noise ratio (SNR), mean sensitivity (MS), and first-order autocorrelation (AR1). All correlations and statistics are for the common period 1937–2006
Finally, the most important relationships showed a great temporal stability along the whole study period (Fig. 6). The moving correlations for 40-year segments always remained at a significance level of at least P < 0.0001 for MAX for Q. pyrenaica. NV was quite stable for both species, with probabilities between P < 0.01 and P < 0.001; however, Q. petraea exhibited a horizontal trend along most of the period, with only a slight increment from the 60 s, whereas Q. pyrenaica was more irregular, reaching the highest values in recent years.

Temporal variation of the main climate–growth relationships obtained for, a Q. pyrenaica and b Q. petraea in the anatomical variables maximum vessel area (MAX) and total number of vessels (NV), expressed as moving bootstrapped Pearson’s correlations for 40-year periods. Dot size is proportional to the significance level (P < 0.05, P < 0.01, P < 0.001, or P < 0.0001) of the bootstrapped correlation coefficient
Discussion
Dendroclimatological meaning of growth and anatomical variables
Width series appear to have a better quality for dendrochronology than anatomical variables. According to our results, RW showed a stronger common signal, coupled to a higher year-to-year variability than vessel features for both species. In fact, common signals of anatomical chronologies were very low, especially for vessel areas. This is a common situation, also observed in previous studies for different ring-porous species, such as Quercus spp. (García-González and Eckstein 2003; Fonti and García-González 2008; Gea-Izquierdo et al. 2012), Castanea sativa (Fonti and García-González 2004), or even for diffuse-porous trees such as Q. ilex (Campelo et al. 2010). Although this finding apparently suggests that RW would be more appropriate for dendrochronological studies (Fonti and García-González 2004; Campelo et al. 2010; Matisons and Brūmelis 2012), common signal only indicates the statistical quality of each variable, without providing the recorded climatic information (Campelo et al. 2010), which needs comparing chronologies between each other and also to climatic data. García-González and Fonti (2006) showed that the common signal of vessel size of oak and chestnut tended to remain low even if sample size was increased, while climate–chronology relationships were very robust to a considerably small sample size. This seems to be the case for our study, since a higher common signal in RW or LW did not allow identifying clear climate–growth relationships; on the contrary, they were strong and reliable for most vessel variables, despite even having a negligible common signal in some cases.
Tree-ring variables were not independent of each other, which was evidenced not only by comparisons among chronologies, but also in the climate–chronology relationships, which showed groups of chronologies yielding a similar result. Previous works had already analysed the differences or analogies in the climatic signal of several anatomical variables (García-González and Eckstein 2003; Olano et al. 2012; Gea-Izquierdo et al. 2012), but to our knowledge, no previous work dealt with a large data set as the one considered initially in this work (23 variables) for hardwoods, while Bryukhanova and Fonti (2013) obtained multiple variables from tracheid measurements in two conifer species. We combined different methodological approaches to optimize the climatic signal, and consequently reduce the number of study variables, namely PCA for chronology comparison (Fonti and García-González 2004; Bryukhanova and Fonti 2013), vessel selection according to the position within the ring (García-González and Fonti 2008; Campelo et al. 2010), and vessel selection upon size (García-González and Fonti 2006; Campelo et al. 2010). The first two methods allowed us to identify three groups of variables with a similar response, whereas the third facilitated the optimization of a climatic response already identified. Thus, apart from the two variables including latewood width (RW and LW), which clearly differed from the others, anatomical variables were split into two groups. These groups were determined by vessel size or number, and by vessel position, i.e. vessels lying in the first row or in additional rows.
The appearance of the earlywood produced in ring-porous oaks is mainly determined by the balance between vessel number (usually distributed into only 1–3 rows), and vessel size (normally very large). As the largest vessels tend to remain in the first row, vessel number in this first row is rather constant, and this explains why the total vessel number (NV) was grouped with all variables not belonging to the first row; but even “intermediate” variables, i.e. those influenced not only by number but also by size (TVA, KS, and EW), were closer to NV, indicating that the total earlywood formed and its associated conductivity depend more on the number of vessel rows than on the number of vessels.
A second group comprised vessel size variables, but together with the rest of variables belonging to the first row of earlywood. In addition, vessel size and number appear to be nearly independent from each other, so that there does not seem to be an efficiency vs. safety trade-off (Sperry et al. 2008) within the earlywood; this means that the existence of smaller earlywood vessels is not associated with an increase in their number and vice versa; however, we cannot rule out that size and distribution of other conducting elements within the ring (latewood vessels or tracheids), which are probably mostly involved in safety, could be somehow related to the characteristics of the earlywood produced. Although all variables within this group had a similar response to climate, we chose MAX instead of MVA for Q. pyrenaica, because the n-percentage filter clearly showed that the correlation was maximized when most small vessels had been eliminated. Consequently, the largest vessels in the tree ring, nearly always located in the first row, recorded the strongest climatic signal, as has been pointed out before in other studies (García-González and Fonti 2006; Campelo et al. 2010). This result proves the need for an optimization of climate–growth relationships when using vessel size, which is also of great methodological relevance. If some of the main climatic signals can be recorded when measuring only the largest vessels of the tree rings, the detection limit could be raised during the process of image analysis, and thus accelerated the process by using images with a lower resolution (Fonti et al. 2009). As regards Q. petraea, vessel size appears to be nearly independent of climate, since we found very weak responses that could not be improved by using the size filter.
Ecophysiological interpretation of climate–chronology relationships
Although several authors often found that vessel features were controlled by climatic conditions during vessel formation (e.g. García-González and Eckstein 2003; Fonti and García-González 2008; Matisons and Brūmelis 2012; Gea-Izquierdo et al. 2012), which probably affect vessel ontogeny directly, all our findings indicate that both vessel size and number of vessels are related to processes that occurred previously. This situation is not uncommon for anatomical variables, and has also been evidenced in other oak species (Matisons and Brūmelis 2012). Specifically, our results point out that factors affecting non-structural carbohydrate dynamics are the most plausible explanation. In fact, deciduous oaks are ring-porous trees, which usually form the large earlywood vessels before bud break (Hinckley and Lassoie 1981; Bréda and Granier 1996), whose formation is therefore supported by previous reserves. But the driving mechanisms are different for the number of vessels and for vessel size.
The number of vessels showed a clear negative relation to temperature during December–March, both in Q. petraea and Q. pyrenaica, with no remarkable differences between species. Due to the mountain conditions of the study area that should delay cambial reactivation and phenology, this timing is likely to totally extend within the dormant period. As a result, the observed response is probably a consequence of consumption of stored reserves for winter respiration. A higher temperature increases respiration rate in the tree, so that a lower amount of carbohydrates would be available for growth in early spring (Gea-Izquierdo et al. 2012), resulting in the productions of fewer cells (and therefore vessels) in the wood. In fact, Hoch et al. (2003) found a very pronounced decrease in non-structural carbohydrates during the early growing season for oak, which was attributed to its ring-porous condition. But we cannot rule out that other more complex mechanisms are involved in the observed response, because the strongest response was observed during February, and numerous hormonal (e.g. Lachaud et al. 1999) and cell structural changes (Farrar and Evert 1997) occur during the quiescent period, thus affecting vessel features. In fact, Fonti et al. (2007) associated a similar relationship to processes occurring during quiescence for vessel size in chestnut, which were probably mediating auxin sensitivity in the cambium.
Vessel size responded strongly to precipitation of the previous year (June–September), but only in Q. pyrenaica. No significant responses were found for Q. petraea, which reflects that rainfall is not a limiting factor for vessel growth of this species in the study area. A strong relationship between vessel size and precipitation is common for ring-porous species, but affecting in different moments during the growing season, mainly during spring (García-González and Eckstein 2003; Fonti and García-González 2008), but also in the previous year (Matisons and Brūmelis 2012). Our climatic responses were clearly related to conditions in the previous growing season, namely summer time, suggesting carbohydrate storage as the most reliable explanation. Barbaroux and Bréda (2002) observed that the accumulation of the reserves occurred mainly during summer months for oaks, and this process is greatly influenced by climatic conditions (Gallé et al. 2007). Considering that rainfall minimum occurs during summer at the study area, and water shortage can consequently be limiting for photosynthesis, rainfall during the previous summer should improve the filling of carbon storage pools (Michelot et al. 2012), facilitating the formation of larger vessels at the beginning of the following season for Q. pyrenaica; or the opposite situation if water availability strongly decreases during summer. Carbohydrate storage is very important for this species, because ring-porous trees as Q. pyrenaica develop their first earlywood vessels just before or at the time of bud break (Suzuki et al. 1996; Schmitt et al. 2000), and the beginning of earlywood formation should be supported by the mobilization of reserves stored during the previous season. Furthermore, Yang and Midmore (2005) reported that the main amounts of stored reserves are used for development of leaves in the current growing season. As expanding leaves are the main sources of auxin, which regulates cambial activity (Fukuda 2004), this could also indirectly influence the variation in vessel size observed for Q. pyrenaica.
Relevance for climatic and ecological studies
The results of this work confirm the usefulness of dendrochronological techniques applied to tree-ring anatomy to study global change (Fonti et al. 2010). We established climatic signals on wood anatomy that resulted to be robust, compatible with reliable physiological hypotheses, and, in addition, stable through time.
Many dendroclimatological studies based on tree-ring widths pointed out clear instabilities in the climatic responses (e.g. Briffa et al. 2004; Büntgen et al. 2006) which are usually called the ‘divergence problem’ (D’Arrigo et al. 2008). This process is defined as a change in climate sensitivity especially around the middle of the twentieth century, and attributed to climate change. Unstable climatic responses have also been observed for oaks, both in high latitudes and temperate conditions (Friedrichs et al. 2009; Visser et al. 2010). Mérian et al. (2011) evaluated the spatio-temporal variation for 31 tree-ring chronologies of Q. petraea along a wide range of climate conditions in France, highlighting a general instability of climate–growth relationships. Moreover, some lack of temporal stability was also found in vessel features (Fonti et al. 2009), suggesting that vessel size in Q. petraea was not an optimal proxy in ecological reconstructions. However, we showed that climate–growth relationships obtained for earlywood vessel features of Q. petraea and Q. pyrenaica, especially vessel size, revealed a strong temporal stability. Though limited to a period slightly shorter than one century because of series length and the availability of climatic data, dendrochronology applied to anatomical features is probably an adequate proxy for long-term climate reconstructions in the study area. But this should be further confirmed by other studies involving longer tree-ring series.
The climatic responses found in our study are of great importance to better understand the possible consequences of climatic variation on both Q. petraea and Q. pyrenaica, as they point out some ideas on their higher or lower capacity to cope with rising temperatures and decreasing rainfall. In a rapidly changing environment, narrowly adapted species with a low plasticity would be highly prone to be displaced by other species (Valladares et al. 2007). Two properties of the xylem, namely size and number of vessels, could trigger a wider plasticity in both oaks. Wide vessels have the advantage of a greater water transport capacity, but they are also more vulnerable to drought-induced cavitation than narrow vessels, emerging the importance of the concept efficiency versus safety, widely cited in many studies (e.g. Hacke and Sperry 2001; Hacke et al. 2006; Gea-Izquierdo et al. 2012). On the other hand, vessel numbers could compensate for an optimal conductivity when vessel size is reduced.
We demonstrated that climate modulates vessel number in both oak species, but also vessel size in Q. pyrenaica. Therefore, higher winter temperatures would be expected to lead to formation of a xylem with fewer vessels, with consequent loss of water conductivity both in Q. petraea and Q. pyrenaica. We hypothesize that the strong adaptation of vessel size in Q. pyrenaica to previous year conditions could indicate more plasticity of this species, i.e. a better ability of individuals to modify their behaviour, anatomy, or physiology in response to altered environmental conditions (Gallé et al. 2010; Gea-Izquierdo et al. 2012). In contrast, Q. petraea does not seem to have this capacity for adjusting its vessel size according to water conditions, which would confer it a lower probability of adaptation. The apparently different plasticity between both species was identified through the statistical analysis of the anatomical time series, but a slightly higher variability among rings and individuals of Q. pyrenaica could be occasionally observed on the core surface (Fig. 7). However, we must be aware of the limitations of this work, which was carried out only in one geographical area, and the analysis performed did not allow us to establish unequivocally if the obtained responses to climate are more likely to indicate adaptive plasticity or vulnerability to harsh climatic conditions. But it clearly demonstrates the exceptional value of vessel chronologies as a tool to study the behaviour of these oaks through time, and should be considered for further studies involving tree-ring networks along the transition area between the Atlantic and Mediterranean regions.

Surface picture of two core fragments for both Q. petraea (a) and Q. pyrenaica (b), illustrating the most common anatomical appearance of both species. Although a wide range of vessel sizes and numbers are observed across individuals, Q. pyrenaica usually presents a higher variability within and among trees, and also more earlywood vessels outside the first row
References
Alcamo J, Moreno JM, Nováky B, Bindi M, Corobov R, Devoy R (2007) Europe: impacts, adaptation and vulnerability. Contribution of Working Group II to the fourth assessment report of the Intergovernmental Panel on Climate Change. Cambridge University Press, Cambridge, pp 541–580
Barbaroux C, Bréda N (2002) Contrasting distribution and seasonal dynamics of carbohydrate reserves in stem wood of adult ring-porous sessile oak and diffuse-porous beech trees. Tree Physiol 22:1201–1210
Benito-Garzón M, Sánchez-DeDios R, Sainz-Ollero H (2008) Effects of climate change on the distribution of Iberian tree species. Appl Veg Sci 10:877–885
Biondi F (1997) Evolutionary and moving response functions in dendroclimatology. Dendrochronologia 15:139–150
Bradley NL, Leopold AC, Ross J, Huffaker W (1999) Phenological changes reflect climate change in Wisconsin. Proc Natl Acad Sci USA 96:9701–9704
Bréda N, Granier A (1996) Intra and interannual variations of transpiration, leaf area index and radial growth of a sessile oak (Quercus petraea). Ann For Sci 53:521–536
Briffa K, Jones PD (1992) Basic chronology statistics and assessment. In: Cook ER, Kairiukštis LA (eds) Methods of dendrochronology. Applications in the environmental sciences. Kluwer, Boston, pp 137–153
Briffa KR, Osborn T, Schweingruber F (2004) Large-scale temperature inferences from tree rings. Global Planet Change 40:11–26
Bryukhanova M, Fonti P (2013) Xylem plasticity allows rapid hydraulic adjustment to annual climatic variability. Trees 27:485–496
Büntgen U, Frank D, Schmidhalter M, Neuwirth B, Seifert M, Esper J (2006) Growth/climate responses shift in a long subalpine spruce chronology. Trees 20:99–110
Campelo F, Nabais C, Gutiérrez E, Freitas H, García-González I (2010) Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 24:463–470
Christman MA, Sperry JS, Smith DD (2012) Rare pits, large vessels and extreme vulnerability to cavitation in a ring-porous tree species. New Phytol 193:713–720
Cook ER (1992) A conceptual linear aggregate model for tree rings. In: Cook ER, Kairiukštis LA (eds) Methods of dendrochronology. Applications in the environmental sciences. Kluwer, Boston, pp 98–104
D’Arrigo R, Wilson R, Liepert B, Cherubini P (2008) On the divergence problem in Nothern forest: a review of the tree-ring evidence and possible causes. Glob Planet Change 60:289–305
Eilmann B, Weber P, Rigling A, Eckstein D (2006) Growth reactions of Pinus sylvestris L. and Quercus pubescens Willd. to drought years at a xeric site in Valais, Switzerland. Dendrochronologia 23:121–132
Farrar JJ, Evert RF (1997) Seasonal changes in the ultrastructure of the vascular cambium of Robinia pseudoacacia. Trees 11:191–201
Fonti P, García-González I (2004) Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol 163:77–86
Fonti P, García-González I (2008) Earlywood vessel size of oak as potential proxy for spring precipitation in mesic sites. J Biogeogr 35:2249–2257
Fonti P, Solomonoff N, García-González I (2007) Earlywood vessels of Castanea sativa Mill. record temperature prior to their formation. New Phytol 173:562–570
Fonti P, Eilmann B, García-González I, von Arx G (2009) Expeditious building of ring-porous earlywood vessel chronologies without loosing signal information. Trees 23:665–671
Fonti P, von Arx G, García-González I, Eilmann B, Sass-Klaassen U, Gärtner H, Eckstein D (2010) Studying global change through investigation of the plastic responses of xylem anatomy in tree rings. New Phytol 185:42–53
Friedrichs DA, Büntgen U, Frank DC, Esper J, Neuwirth B, Loffler J (2009) Complex climate controls on 20th century oak growth in Central-West Germany. Tree Physiol 29:39–51
Fukuda H (2004) Signals that control plant vascular cell differentiation. Mol Cell Biol 5:379–391
Gallé A, Haldimann P, Feller U (2007) Photosynthetic performance and water relations in young pubescent oak (Quercus pubescens) trees during drought stress and recovery. New Phytol 174:799–810
Gallé A, Esper J, Feller U, Ribas-Carbo M, Fonti P (2010) Responses of wood anatomy and carbon isotope composition of Quercus pubescens saplings subjected to two consecutive years of summer drought. Ann For Sci 67:809
García-González I, Eckstein D (2003) Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiol 23:497–504
García-González I, Fonti P (2006) Selecting earlywood vessels to maximize their environmental signal. Tree Physiol 26:1289–1296
García-González I, Fonti P (2008) Ensuring a representative sample of earlywood vessels for dendroclimatological studies: an example from two ring-porous species. Trees 22:237–244
Gea-Izquierdo G, Fonti P, Cherubini P, Martín-Benito D, Chaar H, Canellas I (2012) Xylem hydraulic adjustment and growth response of Quercus canariensis Willd. to climatic variability. Tree Physiol 32:401–413
Gian-Reto W, Post E, Convey P, Menzel A, Parmesan C, Beebee TJC, Fromentin JM, Hoegh-Guldberg O, Bairlein F (2002) Ecological responses to recent climate change. Nature 416:339–395
Grabherr G, Gottfried M, Pauli H (1994) Climate effects on mountain plants. Nature 369:448
Grissino-Mayer HD (2001) Evaluating crossdating accuracy: a manual and tutorial for the computer program Cofecha. Tree-Ring Res 57:205–221
Grissino-Mayer HD (2008) A dendrochronology program library in R (dplR). Dendrochronologia 26:115–124
Hacke UG, Sperry JS (2001) Functional and ecological xylem anatomy. Perspect Plant Ecol Evol Syst 4:97–115
Hacke UG, Sperry JS, Wheeler JK, Castro L (2006) Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol 26:689–701
Hansen AJ, Neilson RP, Dale VH, Flather CH, Iverson LR, Currie DJ, Shafer S, Coot R, Bartlein PJ (2001) Global change in forests: responses of species, communities, and biomes. Bioscience 51:765–779
Hernández-Santana V, Martínez-Vilalta J, Martínez-Fernández J, Williams M (2009) Evaluating the effect of drier and warmer conditions on water use by Quercus pyrenaica. For Ecol Manage 258:1719–1730
Hinckley TM, Lassoie JP (1981) Radial growth in conifers and deciduous trees: a comparison. Mitt Forstl Bundesvers Wien 142:17–56
Hoch G, Richter A, Körner C (2003) Non-structural carbon compounds in temperate forest trees. Plant Cell Environ 26:1067–1081
Lachaud S, Catesson AM, Bonnemain JL (1999) Structure and functions of the vascular cambium. C R Acad Sci III 322:633–650
Martínez-Vilalta J, Prat E, Oliveras I, Piñol J (2002) Hydraulic properties of roots and stems of nine woody species from a holm oak forest in NE Spain. Oecologia 133:19–29
Mason SJ, Mimmack GM (1992) The use of bootstrap correlation coefficients in climatology. Theor Appl Climatol 45:229–233
Matisons R, Brūmelis G (2012) Influence of climate on tree-ring and earlywood vessel formation in Quercus robur in Latvia. Trees 26:1251–1266
McDowell N, Pockman W, Allen CD, Breshears DD, Cobb N, Kolb T, Plaut J, Sperry J, West A, Williams DG, Yepez A (2008) Mechanisms of plant survival and mortality during drought: why do some plants survive while others succumb to drought? New Phytol 178:719–739
Mencuccini M, Martínez-Vilalta J, Piñol J, Loepfe L, Burnat M, Álvarez X, Camacho J, Gil D (2010) A quantitative and statistically robust method for the determination of xylem conduit spatial distribution. Am J Bot 97:1247–1259
Mérian P, Bontemps JD, Bergés L, Lebourgeois F (2011) Spatial variation and temporal instability in climate-growth relationships of sessile oak (Quercus petraea (Matt.) Liebl.) under temperate conditions. Plant Ecol 212:1855–1871
Michelot A, Bréda N, Damesin C, Dufrêne E (2012) Differing growth responses to climatic variations and soil water deficits of Fagus sylvatica, Quercus petraea and Pinus sylvestris in a temperate forest. For Ecol Manage 265:161–171
Olano JM, Eugenio M, García-Cervigón AI, Folch M, Rozas V (2012) Quantitative tracheid anatomy reveals a complex environmental control of wood structure in continental Mediterranean climate. Int J Plant Sci 173:137–149
Parmesan C (2006) Ecological and evolutionary responses to recent climate change. Ann Rev Ecol Evol Syst 37:637–669
Pop EW, Oberbauer SF, Starr G (2000) Predicting vegetative bud break in two arctic deciduous shrub species, Salix pulchra and Betula nana. Oecologia 124:176–184
Rasband W (2009) ImageJ. US National Institute of Health, Bethesda, Maryland, USA. http://rsb.info.nih.gov/ij. Accessed 7 March 2012
R Development Core Team (2012) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna
Sánchez-DeDios R, Benito-Garzón M, Sainz-Ollero H (2009) Present and future extension of the Iberian submediterranean territories as determined from the distribution of marcescent oaks. Plant Ecol 204:189–205
Sass U, Eckstein D (1995) The variability of vessel size in beech (Fagus sylvatica L.) and its ecophysiological interpretation. Trees 9:247–252
Schmitt U, Möller R, Eckstein D (2000) Seasonal wood formation dynamics of Beech (Fagus sylvatica L.) and Black Locust (Robinia pseudoacacia L.) as determined by the “pinning” technique. J Appl Bot 74:10–16
Schume H, Grabner M, Eckmullner O (2004) The influence of an altered groundwater regime on vessel properties of hybrid poplar. Trees 18:185–194
Sperry JS, Meinzer FC, McCulloh KA (2008) Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant Cell Environ 31:632–645
Suzuki M, Yoda K, Suzuki H (1996) Phenological comparison of the onset of vessel formation between ring-porous and diffuse-porous deciduous trees in a Japanese temperate forest. IAWA J 17:431–444
Thuiller W, Lavorel S, Araújo MB, Sykes MT, Prentice IC (2005) Climate change threats to plant diversity in Europe. Proc Natl Acad Sci USA 102:8245–8250
Valladares F, Gianoli E, Gomez JM (2007) Ecological limits to plant phenotypic plasticity. New Phytol 176:749–763
Visser H, Büntgen U, D’Arrigo R, Petersen AC (2010) Detecting instabilities in tree-ring proxy calibration. Clim Past 6:367–377
Yang Z, Midmore DJ (2005) Modelling plant resource allocation and growth partitioning in response to environmental heterogeneity. Ecol Model 181:59–77
Zanne AE, Westoby M, Falster DS, Ackerly DD, Loarie SR, Arnold SEJ, Coomes DA (2010) Angiosperm wood structure: global patterns in vessel anatomy and their relation to wood density and potential conductivity. Am J Bot 97:207–215
Zimmermann MH (1983) Xylem structure and ascent of sap. Springer, New York
Acknowledgments
We thank Sabrina Aldao and Adrián González for field and laboratory assistance, and the Servicio de Montes y Conservación de la Naturaleza de Cantabria for permission for coring trees in Monte Hijedo Natural Park. Rosa Ana Vázquez commented on an earlier version of his manuscript. B. D. González-González benefited from Maria Barbeito pre-doctoral fellowship by Galicia Government. V. Rozas benefited from research contracts by INIA-Xunta de Galicia and CSIC. This research was funded by Instituto Nacional de Investigación y Tecnología Agraria y Alimentaria (INIA), Spanish Ministry of Science and Innovation (RTA2006-00117). We are also grateful to two anonymous reviewers for their useful suggestions.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by A. Nardini.
Rights and permissions
About this article
Cite this article
González-González, B.D., Rozas, V. & García-González, I. Earlywood vessels of the sub-Mediterranean oak Quercus pyrenaica have greater plasticity and sensitivity than those of the temperate Q. petraea at the Atlantic–Mediterranean boundary. Trees 28, 237–252 (2014). https://doi.org/10.1007/s00468-013-0945-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-013-0945-2