
Abstract
Key message
Intra-annual tree-ring parameters of Quercus brantii contain high-resolution intra-annual climate signals which enable us to trace seasonal aspects of climate change and to reconstruct high-resolution climate data for the semi-arid region.
Abstract
Environmental conditions affect growth potential and wood-anatomical features of tree species. Hence, valuable climate signals can be extracted from intra-annual tree-ring features. In this study, we evaluated the potential of intra-annual wood parameters of Quercus brantii Lindel growing in the semi-arid southern Zagros Mountains, Iran. We analyzed earlywood width (EWW), latewood width (LWW), total ring-width (TRW), and several vessel features of the oak species. Standard chronologies have been developed for ring-width and vessel parameters using dendrochronological and quantitative wood-anatomical approaches. Correlations with local climate data showed that precipitation during the pre-growing and growing seasons had positive effects on EWW, LWW, TRW, and latewood vessel size. In contrast, earlywood vessel size showed positive correlations with precipitation in the active growing period (January–April). EWW, LWW, and TRW showed negative correlations with temperature during the pre-growing and growing seasons. Earlywood and latewood vessel features showed stronger negative correlations with mean monthly temperatures during the vessel formation period. Our results revealed that EWW, LWW, and earlywood and latewood anatomical variables of the trees contain valuable climatic signals complementing each other over different seasons. Despite the often low common signal strength of the anatomical variables shared between trees, they showed strong climate–growth relationships which can be useful for the reconstruction of seasonally resolved climate parameters in a multi-parameter tree-ring approach.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Within the Zagros Mountains in West Iran, oak woodlands cover about 5 million hectares and form around 40% of Iran’s total forest cover (Sabeti 2002; Sagheb -Talebi et al. 2014). Hence, they play an important role from an ecological and economic perspective. However, the oak woodlands are under strong pressure due to climate change, the occurrence of dust storms (Moradi et al. 2017), and anthropogenic effects including wide-spread fires and overgrazing (Jazirehi and Ebrahimi-Rastaghi 2003; El-Moslimany 1986). Recorded instrumental climate data show an increase of regional temperatures accompanied by a decline in annual precipitation, probably aggravating drought stress. Based on Iran’s Meteorological Organization, 96% of the total area of the country has experienced mild-to-extreme droughts during the last 7 years (2011–2017), and the 2017 autumn was reported as the driest autumn on record throughout the country. The seasonal climate changes and climate extremes have negatively affected most of the ecosystems and water sources in Iran (Gohari et al. 2013; Delju et al. 2013; Tabari and Hosseinzadeh-Talaee 2011). However, it is difficult to bring the climatic fluctuations and the current oak forest decline into a long-term context, since information about regional past climatic fluctuations and existing knowledge about the history of the oak woodlands and their bioclimatic requirements are still scarce.
To evaluate the impact of climatic stress on the growth behavior of tree species and to obtain information on past climate variability, tree-ring width (TRW) has been widely used (e.g., D’Arrigo et al. 1997; Jacoby et al. 2004; Akkemik et al. 2005; Gervais 2006; Griggs et al. 2007; Cufar et al. 2008; Friedrichs et al. 2008; Arsalani et al. 2014; Gildehaus et al. 2015; Dawson et al. 2015; Leal et al. 2015), but recent progress in wood-anatomical techniques (Gärtner et al. 2015) allow for the inclusion of additional parameters that might add valuable information about climate parameters to which ring width does not respond.
In ring-porous oak species, the boundaries of earlywood (EW) and latewood (LW) can easily be delineated from the distribution of vessels of strongly varying size. Since the intra-annual tree-ring features are affected by environmental conditions during wood formation (Eckstein 2004), the width of earlywood (EWW), latewood (LWW), and intra-annual vessel parameters in various oak species have been used to extract intra-annual climate signals (e.g., Garcia-Gonzalez and Eckstein 2003; Kern et al. 2013; González-González et al. 2014, 2013; Kniesel et al. 2015; Fonti and González-González 2004; Tardif and Conciatori 2006; Leal et al. 2007; Fonti and González-González 2008; Matisons and Dauskane 2009; Campelo et al. 2010; Fonti et al. 2013; Oladi et al. 2014; Matisons et al. 2015; Bräuning et al. 2016). Using a multiple tree-ring parameter approach may enable us to obtain intra-annual climate signals to trace seasonal aspects of climate change.
The most dominant oak species of the Zagros–Anti-Taurus deciduous oak woodlands is Quercus brantii Lindel which is the most drought tolerant oak species in the region. In the central Zagros Mountains, only a few studies have yet been conducted using TRW of Q. infectoria for studying climate–growth relationships and reconstructing climate variables (Arsalani et al. 2014; Azizi et al. 2013; Nadi et al. 2016). Also in the southern Zagros, where Q. brantii reaches its southern bioclimatic limit, dendroclimatic studies on the species have not yet been accomplished (Arsalani et al. 2018). Since additional climate signals may be obtained from intra-annual tree-ring features of ring-porous oak species (Fonti and Garcia Gonzalez 2008), such analyses may enable us to resolve intra-annual climate signals and improve our knowledge about the effects of the seasonal climate fluctuations on the species’ performance in the southern Zagros Mountains.
In the present study, we examined, for the first time, the suitability of TRW, EWW, LWW, and wood-anatomical parameters in EW and LW of Q. brantii for climate and environmental reconstructions in the southern Zagros Mountains. The main objectives of this study are (1) to evaluate the potential of EWW, LWW, and vessel features of Q. brantii as climate proxies; (2) to identify the strongest climate signals from the intra-annual and annual wood parameters for climate reconstruction purposes; (3) to understand how growth–climate relationships of the species vary throughout a vegetation period. Since regional climate data are only available for the period 1987–2015, we restricted our analysis to this common period, although the collected trees show higher ages and the TRW chronology covered 329 years (1796–2015).
Materials and methods
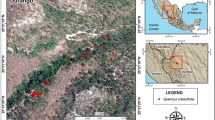
The Deh-Braftab study site (30°46′N, 51°30′E; 1810 m a.s.l.) is located close to the species’ natural southern bioclimatic limit (Fig. 1). In the study forest, the oak trees are scattered (crown coverage below 20%) and the forest structure has been partly degraded by human activities. One characteristic of old Q. brantii trees is heart rot in the inner heartwood part of the stems. The competition among trees for light is low, and the soil of the sampling site is calcareous and very shallow. The Zagros oak woodlands cover a vast area of the Zagros Mountain ranges from Piranshahr (36°00′N, 45°00′E) in the northwest of Iran to the vicinity of Firoozabad (29°00′N, 52°30′E) south of Shiraz, with an average length and width of 1300 and 200 km, respectively (Ghazanfari et al. 2004). In general, Q. brantii is continuously distributed from 1000 to 2400 m a.s.l. in the Zagros Mountains, but, in some parts of the southern Zagros, it is scattered in a wider range from 700 to 2700 m a.s.l. (Jazirehi and Ebrahimi-Rastaghi 2003). In the northern Zagros Mountains, Q. brantii is intermixed with Q. infectoria and Q. libani which are less xerophilous oak species (Jazirehi and Ebrahimi-Rastaghi 2003), whereas, in the central and southern regions of Zagros, Q. brantii is the only oak species. Q. brantii can tolerate low precipitation (< 350 mm/year) and very low winter temperatures (Djamali et al. 2010) and is the exclusive oak species in Iran growing in latitudes south of 34°N (El-Moslimany 1986; Zohary 1973). In the Zagros region, very low (below freezing) temperatures in autumn are the causes for cambial dormancy (Arsalani et al. 2018) and the deciduousness of the oak trees. The ring-porous structure of the oak species is probably also associated with a short growing season. The large vessels formed in the beginning of the growing season have a high hydraulic conductivity, allowing a high stomatal conductance and a high carbon gain. Deciduousness is the strategy enabling this oak species to cope with the seasonal patterns of precipitation and temperature.

Location of the study site and nearest meteorological station
The climate of the study site is described using the instrumental data of Yasuj meteorological station (30°41′N, 51°33′E; 1816 m a.s.l.), which is the closest climate station to the sampling site (in a distance of 25 km) and provides 29 years of instrumental data (1987–2015). The climate at the study site is characterized by a long-dry season from June to September and a wet season from November to April (Fig. 2a), when precipitation is brought by westerly disturbances. Mean annual precipitation is 801 mm, 95% of which falls during the wet season and mean annual temperature is 15 °C. Mean annual maximum temperature has increased over the period 1987–2015 (Fig. 2b) and frost days occur in December, January, and February (Fig. 2c).

Climate diagram of Yasuj meteorological station. a Mean monthly temperatures and monthly precipitation for the period 1987–2015; b Inter-annual variability of total annual precipitation and mean annual maximum temperature; c Mean monthly minimum temperatures of the climate station. The dotted black lines indicate the linear trends of the climate variables
In April 2016, 30 samples from 18 trees including 18 increment cores from 12 standing trees and 6 disks (12 measured radii) from dead trees were collected in the field. The samples were air-dried and mounted on wooden holders, and the surfaces of the samples were sanded using sandpapers with grain sizes from 120 to 1200. We used both air pressure and high-pressure water blast to remove tyloses and wood dust inside the vessels (Fig. 3a). To enhance the contrast of the sample surface for both ring-width measurements and quantitative wood-anatomical image analyses, we rubbed white chalk into all large and small vessels (Fig. 3b). Latewood and, in particular, the heartwood part of the species are dark. Hence, when the vessels were filled by white chalk, a strong visual contrast was obtained, so that the ring boundaries and the vessels were clearly distinguishable from the darker background. This procedure helped taking high-quality images from the surface of the samples for image analyses. It should be noted that the first row of earlywood cells of the year 2016 was already present on the collected samples, indicating that wood formation has already started in March.

Surface pictures from two smoothed cores of Q. brantii, illustrating the tree-ring boundaries and vessel distribution of the species after removing tyloses and wood dust inside the vessels (a) and filling the vessels with chalk (b). Growth direction from right to left. The upper part of the image (a) was modified for the labelling
EWW, LWW, and TRW were measured from bark to pith with a LINTAB 6 measuring system (Rinntech, Heidelberg, Germany) at a precision of 0.01 mm using the software TSAP-Win (Rinn 2003). First, two growth curves of each tree were cross-dated by both visual and statistical tests (Fritts 1976), and a mean curve was computed for each sampled tree by averaging the two radii. Then, site chronologies for the width parameters were established by averaging all mean curves.
The five samples that showed the highest correlations of their EW, LW, and TRW growth curves with the mean chronology were selected for quantitative analysis of wood-anatomical features. High-resolution digital images (5936 pixels × 3392 pixels) from cross sections of the last 29 growth rings (1987–2015) were obtained using a digital microscope ZEISS Smartzoom 5. The contrast of vessel lumen area from the background tissue was increased using the image processing options of the microscope. This procedure enables image analysis software to correctly recognize and measure all vessels. WinCELL PRO 2012a (Regent Instruments, Canada) was applied to analyze earlywood and latewood vessels from the digital images. Earlywood and latewood vessels were measured ring by ring separately. EW was defined by one or two rows of large vessels with an area of more than 20,000 µm2, whereas LW contained a large number of small vessels with higher density. The boundaries between EW and LW were distinguished based on the vessel size differences and were traced manually. We excluded rays from the measurements and also defined a size (> 1000 µm2) and shape filter to exclude non-vessel objects that appeared bright on the images. The accuracy of measurements was visually checked and in some cases when the filter did not exclude all non-vessel objects, the image was corrected manually and vessels were re-measured. From the digital images, the following anatomical variables were computed for each tree ring: average vessel lumen area (AVLA, µm2), number of vessels (NV), vessel density (VD, n/mm2), and total vessel lumen area (TVLA, %). In addition, maximum vessel lumen area (MAX µm2) and mean vessel lumen area of vessels larger than the distribution median (M50, µm2) were computed for the earlywood part of the tree rings. The individual time series of the variables were cross-dated visually and statistically using the software TSAP-Win (Rinn 2003). To remove possible age effects and non-climatic trends in the time series of wood parameters, all series were detrended by fitting a 20-year cubic smoothing spline function (Cook and Peters 1981) in the ARSTAN software. After detrending, all series were averaged into chronologies using a biweight robust mean. ARSTAN produces several types of chronologies. Since standard chronologies (STD) of ring width and earlywood and latewood anatomical parameters yielded stronger correlations with climate variables in comparison with residual (RES) and ARSTAN (ARS) chronologies, we selected the STD chronologies of all parameters for further analyses.
The quality of the chronologies was evaluated using the parameters: mean sensitivity (MS), mean correlation between trees (Rbt), expressed population signal (EPS; Wigley et al. 1984), signal-to-noise ratio (SNR), standard deviation (SD), and first-order autocorrelation (AC1) (Fritts 1976). Similarities between tree-ring variables and relationships between the chronologies and climate data were evaluated by calculating Pearson’s correlation coefficients. In addition to Pearson’s correlation coefficients, we used principal component analysis (PCA) to characterize the relation between the 12 ring-width and anatomical variables.
Monthly precipitation from previous October to current May (there is no effective precipitation during June to September at the study site) and mean monthly temperatures (hereafter, MMT) from October of the previous year to September of the growing season were used to calculate climate-chronology relationships of TRW, EWW, LWW, and the wood-anatomical parameters (AVLA, NV, VD, TVLA, MAX, and M50) over the common period 1987–2015. In addition to monthly temperature and precipitation, we used monthly Palmer Drought Severity Index (PDSI) (CRU scPDSI 3.25) data for grid points nearest to the study site from KNMI Climate Explorer (The Royal Netherlands Meteorological Institute; http://climexp.knmi.nl/) to evaluate the relationship between ring widths and earlywood and latewood anatomical standard chronologies with the drought index. Statistical analyses of the present study were performed using SPSS version 22 (IBM, SPSS Statistics 2011).
Results
For the study period 1987–2015, the highest values of TRW, EWW, and LWW occurred in 2004 and the lowest values of TRW and LWW occurred in 2009, while the lowest value for EWW occurred in 2010 (Fig. 4a). Most of earlywood anatomical variables showed low values in 1999, 2002, 2005, 2009–2010, and 2013 years (Fig. 4b). In addition, low values for latewood anatomical variables were observed in 1997 and 2008 (Fig. 4c).

Standard chronologies of total ring-width (TRW), earlywood width (EWW), and latewood width (LWW) (a) and earlywood (b) and latewood (c) anatomical variables for Q. brantii in the southern Zagros Mountains over the period 1987–2015
A total number of 1461 vessels for EW and 2990 vessels for LW were measured from the 29 studied rings. The maximum number of EW and LW vessels of the five analyzed trees were 14 (in 1989) and 30 (in 2002), respectively (Fig. 5a). AVLA of earlywood ranged from 34,528 to 60,392 µm2 and AVLA of latewood ranged from 6209 to 12,368 µm2 (Fig. 5b).

Earlywood and latewood number of vessels (NV) (a) and average vessel lumen area (AVLA) (b) of Q. brantii over the period 1987–2015. The data for the two variables are raw data before detrending
Both LWW and TRW showed higher values of Rbt, EPS, SNR, and MS statistics in comparison with EWW (Table 1), indicating a higher synchronicity and common forcing. For most of the studied tree-ring parameters, the values of autocorrelation (ACBD) before detrending of the series are high (> 0.5), indicating a strong influence of a value to the value of the following year. For example, EWW showed a high value of AC1, indicating that it is strongly influenced by previous year’s growth. This emphasizes the necessity of detrending to remove possible age-related trends from the series before correlating them with climate variables. Among anatomical variables, the highest value of chronology quality statistics was obtained for latewood NV. In general, anatomical variables of both earlywood and latewood showed rather weak common signals (low-to-moderate Rbt and EPS values). It should also be noted that only TRW, LWW, and latewood NV passed the recommended EPS threshold of 0.85 (Wigley et al. 1984).
LWW (r = 0.91) and EWW (r = 0.63) were significantly correlated with TRW (Table 2). None of the earlywood anatomical variables were correlated to TRW, while all latewood anatomical variables except VD were significantly correlated with TRW.
Inter-correlations among the earlywood anatomical variables showed significant positive correlations between TVLA, M50, and AVLA and EWW, while MAX, NV, and VD were not correlated with EWW. Besides, high and significant (p < 0.01) correlations were found between AVLA with M50 (r = 0.93), MAX (r = 0.72), VD (r = − 0.72), and TVLA (r = 0.50). M50 showed significant positive correlations with MAX and TVLA and significant negative correlations with VD. Furthermore, MAX was negatively correlated with VD (r = − 0.53) and positively correlated with both AVLA (r = 0.72) and M50 (r = 0.84). NV showed positive correlations with EWW and negative correlations with AVLA, M50, and MAX.
Among the latewood anatomical variables, only VD showed negative and nonsignificant correlations with LWW. Correlations among latewood anatomical variables were significantly negative between AVLA and VD (r = − 0.82). A significant positive correlation was found between NV and TVLA (r = 0.47). In addition, latewood NV showed positive correlations with LWW and TRW and inverse correlation with AVLA.
Principal component analysis (PCA) showed that the 12 ring-width and anatomical variables of the species can be divided into two groups (Fig. 6). The two first principal components accounted for 60% of the total variance. PCA#1 was associated with LWW, TRW, LTVLA, LAVLA, and LNW, whereas PCA#2 was related with EWW, ETVLA, EAVLA, M50, and MAX. It should be noted that EVD, ENV, and LVD are affected by other factors.

Principal component analysis (PCA) of the correlation matrix of the 12 ring-width and anatomical variables for the period 1987–2015 (see Table 1 for the abbreviations of the variables)
All ring-width variables (EWW, LWW, and TRW) of Q. brantii showed positive relationships with precipitation from the previous October to the current May (Fig. 7a). In general, monthly precipitation in January and December showed the highest positive effect on tree growth. In comparison with EWW and LWW, TRW showed higher positive correlations with monthly precipitation from the previous October to the current April. Significant positive correlations between TRW and monthly precipitation were found in December (r = 0.47) and January (r = 0.53). In addition, significant correlations between LWW with precipitation were found in December (r = 0.37) and January (r = 0.41), while the relationship for EWW was only significant in January (r = 0.48). It should be noted that LWW showed higher correlations with precipitation in autumn (October–December) and also from February to May than EW.

Pearson’s correlation coefficients for standard tree-ring width chronologies (EWW, LWW, and TRW) with monthly precipitation from the previous October to the current May (a) and mean monthly temperatures from the previous October to the current September (b) over the period 1987–2015. Horizontal dashed gray and black lines indicate significance at 0.05 and 0.01 levels, respectively
Inverse correlations between EWW, LWW, and TRW with MMT were found from the previous October to the current September (Fig. 7b). Significant negative correlations between EWW and MMT were found in February (r = − 0.48), March (r = − 0.39), and May (r = − 0.38) of the growing season. In addition, LWW showed significant negative correlations with MMT in May (r = − 0.37), July (r = − 0.48), and August (r = − 0.52). Furthermore, negative correlations between TRW and MMT were found in March (r = − 0.39), May (r = − 0.41), July (r = − 0.55), August (r = − 0.59), and September (r = − 0.39) of the growing season. EWW showed significant negative correlations with winter temperature (January–March), while both LWW and TRW showed higher negative correlations with summer temperature (July–September).
Among the earlywood anatomical variables, TVLA (r = 0.39) and M50 (r = 0.37) were significantly correlated with precipitation in the previous November and January, respectively (Table 3). AVLA, MAX, and M50 showed positive correlations with precipitation during the growing season (January–April), while the relationships during the pre-growing season (October–December) were negative. Relationships between precipitation and NV were mostly negative from the previous October to May.
AVLA, M50, MAX, and TVLA showed negative correlations with MMT from the previous October to September. Significant negative correlations for AVLA and M50 with MMT were found in January and February. In addition, MAX showed the highest significant negative correlations with MMT in the previous October (r = − 0.39), January (r = − 0.54), and February (r = − 0.37). Significant negative correlations between NV (r = − 0.38) and TVLA (r = − 0.42) with MMT were found in November and June, respectively. Relationships between VD and climate variables were almost absent. Clear relationships between NV and precipitation were not found, while the variable negatively correlated with temperature in the previous October to May. Furthermore, AVLA, M50, MAX, and TVLA showed significant negative correlations with winter temperature (January–March).
AVLA, VD, and TVLA of latewood showed positive correlations with monthly precipitation from the previous October to the current May (Fig. 8a). TVLA (r = 0.38) and VD (r = 0.37) showed significant positive correlations with monthly precipitation in December and February, respectively. It should be noted that the highest correlations with monthly precipitation for both AVLA and TVLA were found during the pre-growing season, whereas negative correlations with MMT occurred from the previous October to the current September, with highest negative correlations in August and July (Fig. 8b). NV showed unclear relationships with precipitation and only moderate negative correlations with temperature in July and August. VD showed mostly weak positive relations with temperature during the pre-growing and growing seasons. AVLA and TVLA showed significant positive correlations with autumn precipitation (October–December) and significant negative correlations with autumn and summer (July–September) temperatures.

Pearson’s correlation coefficients between the latewood anatomical standard chronologies (AVLA, NV, VD, and TVLA) and monthly precipitation from the previous October to the current May (a) and mean monthly temperatures from the previous October to the current September (b) for the period 1987–2015. Horizontal dashed gray and black lines indicate significance at 0.05 and 0.01 levels, respectively
Correlations between EWW, LWW, and TRW standard chronologies with monthly PDSI were positive for EWW during the growing season and pre-growing season, while both TRW and LWW showed positive relationships with PDSI from April to September and negative correlations from October to March (Fig. 9a). Significant negative correlations between LWW and PDSI were found for the previous October (r = − 0.47) and November (r = − 0.42), respectively. Among earlywood anatomical variables, only vessel density (VD) showed negative correlations with monthly PDSI during the pre-growing and growing seasons (Fig. 9b). NV of earlywood showed significant positive correlations with PDSI in November, December, and January. All latewood anatomical variables showed positive correlations with PDSI from April to September (Fig. 9c). VD of latewood showed positive correlations with PDSI during the pre-growing and growing seasons, while the relationships for AVLA and TVLA were negative from the previous October to the current March. In addition, NV negatively correlated with PDSI from the previous October to the current January, while the correlation from February to September was positive.

Pearson’s correlation coefficients between ring widths (a), earlywood (b), and latewood (c) anatomical standard chronologies with monthly PDSI (CRU scPDSI 3.25) from the previous October to the current September for the period 1987–2015. Horizontal dashed gray lines indicate significance at 0.05 level
Discussion
In this study, TRW and wood-anatomical parameters of Q. brantii were analyzed to extract climate signals. A strong correlation between TRW and LWW (r = 0.91; Table 2) indicates that variations in total tree-ring width are mainly determined by variations of the width of latewood rather than that of the earlywood, which is also indicated by the higher inter-annual variability of the latewood indicated by MS (Table 1). LWW and TRW showed stronger common signals than EWW and the wood-anatomical variables (except latewood NV), which is in line with studies on other oak species (Kern et al. 2013; González-González et al. 2014; Fonti and García-González 2008). The common signals for both earlywood and latewood vessel areas were low, but, nevertheless, their climate–chronology relationships were strong (Table 3; Fig. 8). Fonti and García-González (2008) stated that, due to the effects of non-climatic disturbances on the common signal, a higher quality of a chronology does not guarantee a stronger climate signal. Low common signals and high climate–chronology relationships have also been reported for vessel features of several oak species (González-González et al. 2014; García-González and; Eckstein 2003; Fonti and García-González 2008; Fonti and García-González 2004; García-González and Fonti 2006).
EWW, LWW, and TRW showed positive correlations with precipitation during the pre-growing and growing seasons and inverse correlations with MMT from the previous October to the current September. Positive correlations between TRW of Q. brantii growing in the central Zagros Mountains with precipitation and negative correlations with mean and maximum temperatures have been reported by Arsalani et al. (2018) and Azizi et al. (2013). TRW of Q. infectoria in the Zagros oak woodlands shows positive correlations with precipitation and negative correlations with temperature (Nadi et al. 2016; Arsalani et al. 2014; Azizi et al. 2013). Furthermore, positive effects of precipitation on TRW of Q. ilex were reported in the Mediterranean region (Campelo et al. 2009; Corcuera et al. 2004; Cherubini et al. 2003).
Common responses of EWW and the earlywood anatomical parameters (AVLA, M50, and MAX) to precipitation were limited from January to April, which is before the active growing period. In contrast, LWW showed positive correlations with precipitation from February to May. The latewood anatomical parameters (AVLA and TVLA) showed positive, but not significant relationships with precipitation from the previous October–May. These findings underline the positive effects of water supply during the pre-growing and early growing seasons on the formation of the hydraulic system in EW and LW, respectively. Most of the high and low values of the ring-width and earlywood and latewood anatomical variables coincided with maxima and minima of the recorded climate data (Fig. 2). Earlywood and latewood NV were negatively affected by high temperatures in the previous autumn (October–December) and current summer (July and August), although no significant correlations between NV and precipitation were found. Based on the climate–growth relationships (Figs. 7, 8; Table 3), the climate factors are probably the main responsible for controlling width and anatomical parameters of the species. Low values of AC1 for vessel features (Table 1) indicate that these wood-anatomical parameters are not dependent on the previous ring, and based on the climate-growth relationships, they are mostly affected by environmental conditions during the current growth year. This is confirmed by the fact that highest negative correlations between temperature on earlywood and latewood vessel areas occurred during their active growing periods. The previous studies also found that vessel features are mostly affected by climate conditions during vessel formation (Fonti and Garcia Gonzalez 2008; Garcia-Gonzalez and Eckstein 2003). Beside poor climatic response, another reason for the unclear relationships between NV and VD of earlywood (Table 3) and latewood (Fig. 8) with precipitation and temperature might also be related to the decreasing trend observed for earlywood and latewood NV (Fig. 5a) before detrending. Hence, these vessel features might possibly be affected by the changing geometry with increasing tree size or age, as it has been documented for conifer tree species (Anfodillo et al. 2012). Currently, no information about variations of wood-anatomical parameters in our study species along the stem is available, but future studies should test if allometric relationships may also have an impact on the variation of anatomical parameters of Q. brantii.
The limiting effect of water supply is confirmed by negative correlations of the tree-ring width variables EWW, LWW, and TRW, and earlywood (AVLA, M50, MAX and TVLA) and latewood anatomical parameters (AVLA and TVLA) with MMT in the pre-growing and growing seasons. High temperatures increase evapotranspiration and hence aggravate water stress on the plants, leading to the formation of smaller vessels which provide higher safety and less risk of hydraulic failure (Sperry et al. 2008; Hacke et al. 2017; Fichtler and Worbes 2012; Pfautsch et al. 2016). However, vessel size and vessel number in EW as well as in LW vary independent from each other (Fig. 6), as also found for temperate and Mediterranean oak species (González-González et al. 2014). Hence, the occurrence of smaller earlywood vessels in EW and LW is not associated with their number; instead, the number of latewood vessels is correlated with the size of the latewood (Table 2). This leads to the conclusion that there is probably no trade-off between efficiency vs. safety in the hydraulic system of Q. brantii.
Correlations of wood-anatomical parameters in EW and LW to temperature were similar, but stronger than those of EWW and LWW. Based on the important role of temperature for the starting date of the growing season (El-Moslimany 1986), the active growing period of the species starts between March and April. The abundant precipitation during the pre-growing season provides sufficient soil moisture during earlywood formation. Correlation of growth and anatomical parameters with temperature are higher (but of opposite sign) than with precipitation, which is an effect frequently observed in semi-arid climates (Arsalani et al. 2014; Nadi et al. 2016). High temperatures occur in the dry summer season when the region is affected by the subtropical high-pressure belt. Therefore, the negative effects of temperature on latewood formation are stronger than on earlywood formation. Especially, latewood vessels are formed under the dry and hot summer conditions, when wood formation depends on precipitation that has fallen during prior months.
Our results showed positive correlations between TRW, EWW, and LWW, and the vessel size variables of both earlywood (AVLA, M50, MAX, and TVLA) and latewood (ALVA and TVLA). In comparison with TRW, EWW, and LWW, the vessel size variables showed higher negative correlations with VD (Table 2). Pourtahmasi et al. (2011) and Stojnic et al. (2013) found similar relationships between TRW, AVLA, and VD of beech species. Positive correlations between ring-width and the vessel size variables can be explained by the positive effects of vessel area on water transport efficiency which allows a higher stomatal conductance and transpiration rates and a higher carbon gain, increasing productivity (Poorter et al. 2010; Stojnic et al. 2013). Earlywood NV showed positive correlations with EWW and negative correlations with AVLA, M50, and MAX of earlywood. In addition, latewood NV correlated positively with LWW and negatively with AVLA of latewood. González-González et al. (2014) found similar results for oaks growing in the Mediterranean region and stated that NV is also related to ring width and controlled by factors different from those controlling vessel size. The results of PCA for Q. brantii show that TRW is strongly related to LWW, which has also been observed for other oak species, e.g., for Q. robur and Q. pyrenaica (González-González et al. 2015) and for Q. alba and Q. rubra (Tardif and Conciatori 2006).
Intra-annual tree-ring features of Q. brantii showed statistically significant intra-annual climate signals, providing high-resolution climate and ecological information (Table 4). However, TRW showed strong relationships with both temperature and precipitation variables, earlywood anatomical parameters of oak species have been frequently found to record climate signals (González-González et al. 2014; Campelo et al. 2010; Fonti and García-González 2004). Our findings not only confirmed the high potential of EWW and earlywood anatomical parameters of Q. brantii, but also showed that some anatomical parameters of latewood, specifically AVLA and TVLA, contain valuable climate signals and are promising proxies for summer-temperature reconstructions, like they were successfully derived from other oak species (e.g., Kern et al. 2013; Pritzkow et al. 2016).
Since our study is the first to evaluate the potential of intra-annual wood parameters on any oak species growing in the Zagros Mountains, no other regional studies are available to compare our results. The present study represents a first step for using wood-anatomical features for climate reconstructions in this sensitive environment.
Conclusions
Climate signals of Q. brantii including ring-width and various anatomical parameters were studied in the southern Zagros Mountains, Iran. The intra-annual tree-ring features of the species showed valuable intra-annual climate signals. Although TRW is a useful climate indicator, EWW and LWW and also the anatomical parameters AVLA, NV, VD, TVLA, MAX, and M50 provide climate signals with monthly and seasonal resolutions. In addition, some of the anatomical parameters showed higher climate correlations than ring-width parameters.
Climate–chronology relationships revealed that precipitation has positive effects on EWW, LWW, TRW, and latewood vessel size during the pre-growing and growing seasons, while the relationship between earlywood vessel size variables and precipitation is positive in the active growing period. Ring-width, earlywood, and latewood vessel size variables showed negative correlations with MMT during the pre-growing and growing seasons, but earlywood and latewood anatomical parameters showed higher negative correlations with MMT during the vessel formation period.
Expanding the network of wood-anatomical chronologies would provide valuable information about climate limiting factors, and could improve our understanding of the growth rate and ecological performance of the species under varying climate conditions over different climatic provinces over Iran. The intra-annual tree-ring features provide high-resolution climate signals which are crucial for the reconstruction of high-resolution climate data for the region that is strongly affected by climate extremes. Intra-annual reconstruction of different climate parameters will enable the reconstruction of atmospheric circulation patterns in different seasons which can be applied in climate modeling and environmental planning.
Author contribution statement
MA designed and executed the study with assistance from AB and KB. Site selection and Sampling were conducted by MA, GA, KP, and HM. Sample preparation, ring-width measurements, building of chronologies, figure perpetration, image analyses, and statistical analyses were performed by MA with assistance from AB. MA wrote the draft of the manuscript. AB and KP provided revisions on the manuscript. The authors approved the final manuscript.
References
Akkemik ϋ, Dagdeviren N, Aras A (2005) A preliminary reconstruction (A.D. 1635–2000) of spring precipitation using oak tree rings in the western Black Sea region of Turkey. Int J Biometeorol 49(5):297–302
Anfodillo T, Deslauriers A, Menardi R, Tedoldi L, Petit G, Rossi S (2012) Widening of xylem conduits in a conifer tree depends on the longer time of cell expansion downwards along the stem. J Exp Bot 63(2):837–845
Arsalani M, Azizi G, Bräuning A (2014) Dendroclimatic reconstruction of May–June maximum temperatures in the central Zagros Mountains, western Iran. Int J Climatol 35:408–416
Arsalani M, Pourtahamsi K, Azizi G, Bräuning A, Mohammadi H (2018) Tree-ring based December–February precipitation reconstruction in the southern Zagros Mountains. Iran Dendrochronol 49:45–56
Azizi G, Arsalani M, Bräuning A, Moghimi E (2013) Precipitation variations in the central Zagros Mountains (Iran) since A.D. 1840 based on oak tree rings. Palaeogeogr Palaeoclimatol Palaeoecol 386:96–103
Bräuning A, De Ridder M, Zafirov N, García-González I, Petrov Dimitrov D, Gärtner H (2016) Tree-ring features: indicators of extreme event impacts. IAWA J 37(2):206–231
Campelo F, Nabais C, García-González I, Cherubini P, Gutiérrez E, Freitas H (2009) Dendrochronology of Quercus ilex L. and its potential use for climate reconstruction in the Mediterranean region. Can J Res 39:2486–2493
Campelo F, Nabais C, Gutiérrez E, Freitas H, García-González I (2010) Vessel features of Quercus ilex L. growing under Mediterranean climate have a better climatic signal than tree-ring width. Trees 24(3):463–470
Cherubini P, Gartner BL, Tognetti R, Bräker O, Schoch W, Innes JL (2003) Identification, measurement and interpretation of tree rings in woody species from Mediterranean climates. Biol Rev 78:119–148
Cook ER, Peters K (1981) The smoothing spline: a new approach to standardizing forest interior tree-ring width series for dendroclimatic studies. Tree-ring Bull 41:45–55
Corcuera L, Camarero JJ, Gil-Pelegrin E (2004) Effects of a severe drought on Quercus ilex radial growth and xylem anatomy. Trees 18:83–92
Cufar K, De Luis M, Eckstein D, Kaifez-Bogataj L (2008) Reconstructing dry and wet summers in SE Slovenia from oak tree-ring series. Int J Biometeorol 52(7):607–615
D’Arrigo R, Yamaguchi D, Wiles G, Jacoby G, Osawa A, Lawrence D (1997) A Kashiwa oak (Quercus dentata) tree-ring width chronology from northern coastal Hokkaido, Japan. Can J For Res 27(4):613–617
Dawson A, Austin D, Walker D, Appleton S, Gillanders BM, Griffin SM, Sakata C, Trouet V (2015) A tree-ring based reconstruction of early summer precipitation in southwestern Virginia (1750–1981). Clim Res 64(3):243–256
Delju AH, Ceylan A, Piguet E, Rebetez M (2013) Observed climate variability and change in Urmia Lake Basin, Iran. Theoret Appl Climatol 112:285–296
Djamali M, Akhani H, Andrieu-Ponel V, Braconnot P, Brewer S, de-Beaulieu JL, Fleitmann D, Fleury J, Gasse F, Guibal F, Jackson ST, Lézine AM Médail M, Ponel F, Roberts N, Stevens L (2010) Indian summer monsoon variations could have affected the early-Holocene woodland expansion in the Near East. The Holocene 20(5):813–820
Eckstein D (2004) Changes in past environments-secretes of tree hydrosystem. New Phytol 163:1–4
El-Moslimany A (1986) Ecology and late-Quaternary history of the Kurdo-Zagrosian oak forest near Lake Zeribar, western Iran. Vegetatio 68:55–63
Fichtler E, Worbes M (2012) Wood anatomical variables in tropical trees and their relation to site conditions and individual tree morphology. IAWA J 33:119–140
Fonti P, Garcia-Gonzalez I (2008) Earlywood vessel size of oak as a potential proxy for spring precipitation in mesic sites. J Biogeogr 35:2249–2257
Fonti P, García-González I (2004) Suitability of chestnut earlywood vessel chronologies for ecological studies. New Phytol 163(1):77–86
Fonti P, Heller O, Cherubini P, Rigling A, Arend M (2013) Wood anatomical responses of oak saplings exposed to air warming and soil drought. Plant Biol 1:210–219
Friedrichs DA, Bϋntgen U, Frank DC, Esper J, Neuwirth B, Löffler J (2008) Complex climate controls on 20th century oak growth in Central-West Germany. Tree Physiol 29:39–51
Fritts HC (1976) Tree rings and climate. Academic Press, London
Garcia-Gonzalez I, Eckstein D (2003) Climatic signal of earlywood vessels of oak on a maritime site. Tree Physiol 23:497–504
García-González I, Fonti P (2006) Selecting earlywood vessels to maximize their environmental signal. Tree Physiol 26:1289–1296
Gärtner H, Banzer L, Schneider L, Schweingruber FH, Bast A (2015) Preparing micro sections of entire (dry) conifer increment cores for wood anatomical time-series analyses. Dendrochronologia 34:19–23
Gervais BR (2006) A three-century record of precipitation and blue Oak recruitment from the Tehachapi Mountains, Southern California, USA. Dendrochronologia 24:29–37
Ghazanfari H, Namiranian M, Sobhani H, Marvi-Mohajer MR (2004) Traditional forest management and its application to encourage public participation for sustainable forest management in the northern Zagros Mountains of Kurdistan province, Iran. Scand J For Res 19(4):65–71
Gildehaus S, Arabas K, Larson E, Cipes-Gerbitz K (2015) The dendroclimatological potential of Willamette Valley Guercus Garryana. Tree-ring Research 71(1):13–23
Gohari A, Eslamian S, Abedi-Koupaei J, Massah-Bavani A, Wang D, Madani K (2013) Climate change impacts on crop production in Iran’s Zayandeh-Rud River Basin. Sci Total Environ 422:405–419
González-González BD, García-González I, Vázquez-Ruiz RA (2013) Comparative cambial dynamics and phenology of Quercus robur L. and Q. pyrenaica Willd. in an Atlantic forest of the northwestern Iberian Peninsula. Trees 6(27):1571–1585
González-González BD, Rozas V, García-González I (2014) Earlywood vessels of the sub-Mediterranean oak Quercus pyrenaica have greater plasticity and sensitivity than those of the temperate Q. petraea at the Atlantic–Mediterranean boundary. Trees 28(1):237–252
González-González BD, Vázquez-Ruiz RA, García-González I (2015) Effects of climate on earlywood vessel formation of Quercus robur and Q. pyrenaica at a site in the northwestern Iberian Peninsula. Can J For Res 45:698–709
Griggs C, DeGaetano A, Kuniholm P, Newton M (2007) A regional high-frequency reconstruction of May–June precipitation in the north Aegean from oak tree rings. AD Int J Climatol 27:1089–1198 1075–1089
Hacke UG, Spicer R, Schreiber SG, Plavcová L (2017) An ecophysiological and developmental perspective on variation in vessel diameter. Plant Cell Environ 40:831–845
Jacoby G, Solomina O, Frank D, Eremenko N, D’Arrigo R (2004) Kunashir (Kuriles) oak 400-year reconstruction of temperature and relation to the Pacific Decadal Oscillation. Palaeogeogr Palaeoclimatol Palaeoecol 209:303–311
Jazirehi MH, Ebrahimi-Rastaghi M (2003) Silviculture in Zagros. Tehran University Press, Tehran
Kern Z, Patko M, Kazmer M, Fekete J, Kele S, Palyi Z (2013) Multiple tree-ring proxies (earlywood width, latewood width, and 13C) from pedunculate oak (Quercus robur L.), Hungary. Quatern Int 239:257–267
Kniesel B, Günther B, Von Arx G (2015) Defining ecologically relevant vessel parameters in Quercus robur L. for use in dendroecology: a pointer year and recovery time case study in Central Germany. Trees 29:1041–1051
Leal S, Nunes E, Pereira H (2007) Cork oak (Quercus suber L.) wood growth and vessel characteristics variations in relations to climate and cork harvesting. Eur J Forest Res 127(1):33–41
Leal S, Campelo F, Luisa AL, Carneiro MF, Santos JA (2015) Potential of oak tree-ring chronologies from Southern Portugal for climate reconstructions. Dendrochronologia 35:4–13
Matisons R, Dauskane I (2009) Influence of climate on earlywood vessel formation of Quercus robur at its northern distribution range in central regions of Latvia. Acta Universitatis Latviensis 753:49–58
MatisonsR,JansonsJ,KatrevičsJ,JansonsĀ(2015)Relation of tree-ring width and earlywood vessel size of alien Quercus rubra L. with climatic factors in Latvia.Silva Fennica49:1–14
Moradi A, Taheri Abkenar K, Afshar Mohammadian M, Shabanian N (2017) Effects of dust on forest tree health in Zagros oak forests. Environ Monit Assess 189(11):549
Nadi M, Bazrafshan J, Pourtahmasi K, Bräuning A (2016) Tree-ring based reconstruction of the joint deficit index in Javan-Roud Region, Kermanshah (Iran). Int J Climatol 37:420–429
Oladi R, Bräuning A, Pourtahmasi K (2014) Plastic and static behaviour of vessel-anatomical features on Oriental beech (Fagus orientalis Lipsky) in view of xylem hydraulic conductivity. Trees 28:493–502
Pfautsch S, Harbusch M, Wesolowski A, Smith R, Macfarlane C, Tjoelker MG, Reich PB, Adams MA (2016) Climate determines vascular traits in the ecologically diverse genus Eucalyptus. Ecol Lett 19:240–248
Poorter L, McDonald I, Alarcon A, Fichtler E, Licona JC, Pena-Claros M, Sterck F, Villegas Z, Sass-Klaassen U (2010) The importance of wood traits and hydraulic conductance for the performance and life history strategies of 42 rainforest tree species. New Phytol 185:481–492
Pourtahmasi K, Lotfiomran N, Bräuning A, Parsapajouh D (2011) Tree-ring width and vessel characteristics of oriental beech (Fagus orientalis) along an altitudinal gradient in the Caspian forests, Northern Iran. IAWA J 32(4):461–473
Pritzkow C, Wazny T, Heußner KU, Słowin´ski M, Bieber A, Dorado Liñán I, Helle G, Heinrich I (2016) Minimum winter temperature reconstruction from average earlywood vessel area of European oak (Quercus robur) in N-Poland. Palaeogeogr Palaeoclimatol Palaeoecol 449:520–530
Rinn F (2003) TSAP-Win: time series analysis and presentation for dendrochronology and related applications. In: Version 0.55 User reference. RINNTECH, Heidelberg. http://www.rimatech.com
Sabeti H (2002) Forests, trees and shrubs of Iran. Yazd University Press, Yazd
Sagheb-Talebi K, Sajedi T, Yazdian F (2014) Forests of Iran: a treasure from the past, a hope for the future plant and vegetation. Springer, Dordrecht
Sperry JS, Meinzer FC, McCulloh KA (2008) Safety and efficiency conflicts in hydraulic architecture: scaling from tissues to trees. Plant Cell Environ 31:632–645
Stojnic S, Sass-Klaassen U, Orlovic S, Matovic B, Eilmann B (2013) Plastic growth response of European beech provenances to dry site conditions. IAWA Journal 34(4):475–489
Tabari H, Hosseinzadeh-Talaee P (2011) Analysis of trends in temperature data in arid and semi-arid regions of Iran. Global Planet Change 79:1–10
Tardif JC, Conciatori F (2006) Influence of climate on tree rings and vessel features in red oak and white oak growing near their northern distribution limit, southwestern Quebec, Canada. Can J For Res 36:2317–2330
Wigley T, Briffa KR, Jones PD (1984) On the average value of correlated time series, with applications in dendroclimatology and hydrometeorology. J Appl Meteorol 23:201–213
Zohary M (1973) Geobotanical Foundations of the Middle East. 2 volumes. Gustav Fischer Verlag
Acknowledgements
The research stay of M. Arsalani at the Institute of Geography, Friedrich-Alexander University of Erlangen-Nuremberg was supported by the Iranian Ministry of Science, Research and Technology (MSRT) and by the German Academic Exchange Service (DAAD) in the framework of German–Iranian scholarship Program (GISP). We thank Iris Burchardt for technical support during the laboratory work in Germany. We also thank the two anonymous reviewers for their constructive comments and suggestions.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Liang.
Rights and permissions
About this article

Cite this article
Arsalani, M., Bräuning, A., Pourtahmasi, K. et al. Multiple tree-ring parameters of Quercus brantii Lindel in SW Iran show a strong potential for intra-annual climate reconstruction. Trees 32, 1531–1546 (2018). https://doi.org/10.1007/s00468-018-1731-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-018-1731-y