
Abstract
How tree morphology develops in mixed-species stands is essential for understanding and modelling mixed-stand dynamics. However, research so far focused on the morphological variation between tree species and neglected the variation within a species depending on intra- and interspecific competition. Our study, in contrast, addresses crown properties of nine mature Norway spruces (Picea abies [L.] Karst.) of a pure stand and compares them with ten spruces growing in mixture with European beech (Fagus sylvatica [L.]). The same was done with 11 pure stand beeches and 12 beeches growing in mixture with spruce. Through application of a terrestrial laser scanner and a new skeletonization approach, we deal with both species’-specific morphological traits such as branch angle, branch length, branch bending, crown volume and space occupation of branches within the crown, some of which were hardly accessible so far. Special attention is paid to distinct differences between trees growing in mixed and pure stands: for spruce, our study reveals significantly longer branches and greater crown volumes in the mixed stand when compared to the pure stand. In case of European beech, individuals growing in mixture show flatter branch angles, more distinct ramification, greater crown volumes and a lower share of a single branch’s space occupation in the total crown volume. The results show that the presented methods yield detailed information on the morphological traits analyzed in this study and that interspecific competition on its own may have a significant impact on crown structures. Implications for production ecology and stand dynamics of mixed-species forests are discussed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
According to the principle of function convergence the structure, physiology and phenology of plants indicate the prevailing environmental conditions under which they grow (see e.g., Assmann 1970; Walter 1931). This explains why Oldemann (1990) or Roloff (2001) use tree morphology and foliation for assessing tree vitality. For tree crowns that mean that their size and form result from the local environmental factors and resource supply which in forest stands are mainly determined by the tree’s inter- or intraspecific neighbourhood within the stand.
The structure of a tree’s crown is crucial in the feedback loop between tree functioning, structure and environment in pure stands (Hari 1985), but even more important in mixed stands where different species demonstrate their abilities to acclimate their structures to capture contested resources more efficiently or deny competitors access to the same resources (Pretzsch 2009).
The importance of crown structure and crown plasticity for the dynamic of mixed species stands is recognized and addressed (Pretzsch 2003; Price et al. 2010; Richards et al. 2010) but hardly elucidated so far, especially in mature stands where crown structure is difficult to measure.
Measurement of tree crowns by e.g., Badoux (1946) and Burger (1939) constitutes the paradigm shift from stand to individual tree level thinking and modelling in forest science. Terrestrial methods for measuring (Röhle and Huber 1985), quantifying (Assmann 1970) and modelling (Pretzsch 1992, 2009) tree crowns were continuously refined. The main objective was to understand, predict and maximize tree growth based on empirical relationships between growing space of the tree, crown size and stem volume growth (Assmann 1970; Pretzsch 2006). A further aspect was the effect of silviculture on crown structure and wood quality (Zobel and van Buijtenen 1989 Wilhelmsson et al. 2002; Seifert 2003). New motivation for crown analysis came from allometry (Niklas 1994) and especially from the metabolic scaling theory (MST) which provides promising synthesis, scaling approaches and models for the functioning and structure of plants from organ to ecosystem level (Enquist et al. 1998; West et al. 2009).
Understanding and modelling of tree growth in mixture require tracing of the causes of over- or underyielding from stand to the tree or even organ level. Space-filling principles within the crown such as branch angle, branch length, branch number and ramification in mixture may differ from pure stands and indicate changed resource supply, resource capture, or resource use efficiency (Binkley et al. 2004). However, these space filling principles and the according morphological patterns triggered by interspecific competition can hardly be revealed by classical crown measurement by recording the crown length and cross section area in order to estimate the extension of the convex hull (Röhle and Huber 1985).
Terrestrial laser scanning (TLS) is widely utilized for high precision measurements in architecture, engineering and archaeology (Vosselman and Maas 2010) and in the last decade also used to measure forest parameters for inventory and even individual tree parameters (Huang and Pretzsch 2010; Pretzsch et al. 2011). The extraction of a structure description from TLS point clouds such as a collection of connected lines is called skeletonization and allows detailed analysis of morphological traits. Automated skeletonization approaches already exist and show great potential (e.g., van Leeuwen et al. 2011; Bucksch et al. 2010; Bremer et al. 2012). These methods however, are often highly species specific, require several scan positions per tree as well as generally free-standing test trees and suffer from difficulties dealing with inhomogeneous point densities, data gaps and noise within the dataset (Bucksch 2011; Côté et al. 2011). Thus, there is still a lot of need for further research until those methods can be used to reliably measure crown parameters at the branch level, especially within forest stands where occlusion is considerably stronger.
In order to contribute to the understanding of inter- and intraspecific competition and its impact on structural parameters, we developed and applied a new, software based manual skeletonization method for TLS point clouds to Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) dealing with the following questions:
-
1.
Is the presented method suitable to reveal structural crown properties, what problems arise and how may they be overcome?
-
2.
What are the characteristic branch angles of both species and does growing within a mixed stand cause variations of these angles?
-
3.
What are the characteristic branch length and branch bending of both species and does growing within mixture influence them?
-
4.
What are the specific length-sum of the branches, number of branches and their distribution regarding branch order of both species and does interspecific competition affect these parameters?
-
5.
How large is the crown volume and the space that individual branches occupy within the crown on a fine scale and do these parameters in mixed stands differ from those in pure stands?
Material and methods
Study area
For scrutiny of structural traits of spruce and beech and their differences in pure versus mixed stands, we selected the age series SON 814 (west–east spread: 10°28′45″E–0°31′39″E, north–south spread: 47°31′05″N–47°32′31″N) which includes both, pure and mixed stands of Norway spruce and European beech and is located 785–800 m above sea level in the ecological region 14.4 “Schwäbisch-Bayerische Jungmoräne und Molassevorberge. Westliche kalkalpine Jungmoräne” in southern Bavaria, about 60 km southwest of Munich, Germany. The mean annual temperature is 6.8 °C with a precipitation of 1,114 mm. During the vegetation period of 140 days (days ≥10 °C) the temperature averages 13.9 °C accompanied by 648 mm of precipitation. The natural vegetation would be a Luzulo-Fagetum association.
We took our sample trees from plot SON814/3 which is composed of parts where both species occur in pure as well as in mixed stands with otherwise equal underlying conditions. An overview of the stand characteristics for SON814/3 is given in Table 1. We considered a tree that had no other tree’s crown in between its own crown and the crown of the sample tree a neighbour. The selection of our sample trees was done in such a way as to ensure that trees classified as growing in pure stand were surrounded exclusively by neighbours belonging to the same species. Accordingly, a sample tree was only classified as growing in mixture if all of its neighbours are members of the opposite species. Furthermore, we closely paid attention to tree size and competitive situation to ensure that trees of both pure and mixed stands grow under comparable conditions and differences in structural crown properties are caused by mixture itself and not by differences in local stand density or tree size. The sample spruces in pure and mixed stand are 128-year old and of comparable average height (pure 38.9 m, mixed 39.2 m) and relative height (pure 0.93, mixed 0.94), i.e. their height in relation to the highest neighbour within a radius of 10 m around the sample tree. The local basal area (pure 64.82 m2/ha, mixed 63.28 m2/ha), i.e. the sum of basal areas of all trees also in a 10 m radius around the sample tree, up-scaled to ha is approximately the same. Spruce’s average dbh differs significantly between pure (45.7 cm) and mixed stand (59.1 cm). The selected beeches are at the age of 148 years in both pure and mixed stand. Mean height (pure 36.8 m, mixed 35.9 m), relative height (pure 0.94, mixed 0.91), local basal area (pure 52.83 m2/ha, mixed 53.39 m2/ha) and dbh (pure 43.7 cm, mixed 42.2 cm) are comparable. A summary of the characteristics of our sample trees is given in Table 2.
Data acquisition
We used the Riegl LMS-Z420i TLS system. This laser scanner system works on the so-called time-of-flight principle. A short laser impulse is fired in a specified direction to a target. A part of the light is reflected back to the scanner. The scanner measures the time between the firing and the return of the signal. Multiplying this time with the speed of light in air gives the double distance to the object. The accuracy for these distance measurements is about 1 cm. The angles azimuth and inclination to the target are recorded with a precision of 0.002°. For each laser impulse the angles are slightly varied (0.06°) resulting in a scanning action. The consecutive distance measurements result in a range image of the desired region. With one setup of the Riegl LMS-Z420i the maximum region is 360° (horizontally) by 80° (vertically) wide. Tilting the scanner by 90° and doing a second scan results in a nearly complete scan of the whole sphere. The originally spherical coordinates (azimuth, inclination and distance) are translated to Cartesian coordinates (x, y, z) for further processing.
Obviously, the measurement density drops with increasing distance to the scanner as the angles are widening with distance. Moreover, the infrared the laser beam is normally not able to penetrate the tree compartments and so it is not possible to make measurements behind an obstacle. These two effects result in rather sparse measurement densities in higher crown regions, especially if the crown parts near to the scanner are dense, like the conifer crowns are (Hilker et al. 2010). Keeping this in mind we set up the scanner between the trees of interest, in a way that the tree crown was covered as much as possible by the laser scan. In addition, we used a distance measurement mode which is called last-pulse or last-target. In this mode the last echo of the laser impulse is used for distance recording, allowing the laser beam to penetrate the tree crown further than using the normal, first-pulse mode.
Skeletonization
In order to determine crown structures as entirely as necessary to gain insights on crown structures, we developed a software-based approach for the manual skeletonization of measured TLS point clouds where the skeletonization of a tree’s point cloud is done semi-manually, using software specifically developed for this purpose. The code is written in the programming language Python to facilitate fast and relatively unproblematic adjustments and good interoperability with other programming languages such as C/C++ for time-critical calculations or R for statistical analysis.
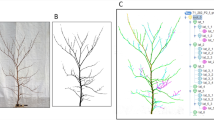
First, trees of sufficient scan quality were manually extracted from the point cloud of the whole scan position retrieved by the terrestrial laser scanner. These single tree point clouds were then skeletonized by visualizing the tree, utilizing the Mayavi program library for Python (Ramachandran and Varoquaux 2008) and letting the user measure the course of a branch by successively defining as many branch segments as possible through a dedicated user interface to get a sufficient image of the tree structure (Fig. 1). The definition of the branch segments is done by picking appropriate members of the point cloud, i.e. points that actually belong to the specific branch with the help of interactive call-back functions and internal plausibility checks. Additional queried information for every branch through user input includes branch type (main axis, secondary axis or branch), the branch order, the branch’s parent-branch and whether the branch is dead or alive. The entirety of the measured branches finally defines the tree’s skeletal structure.

From point cloud to a tree’s skeletal structure: (l) isolated individual tree from scan position image (m) skeletal structure surrounded by original point cloud data (r) isolated tree skeleton
After the skeletonization process, the skeletal structure serves as a basis for various calculations such as branch angles, branch length and branch bending. Furthermore, the point cloud of a tree or the combination of point cloud and skeletal structure facilitates the calculation of crown volume and the space requirement of a branch, i.e. the volume of a branch including smaller branches, twigs and foliage.
Branch angles
We defined a branch’s angle φ as the angle between the vectors \( \overrightarrow {bs} \) and \( \overrightarrow {z} \) where \( \overrightarrow {bs} \) is the vector from the branch’s base to a point s lying on the branch in an arbitrary distance r. \( \overrightarrow {z} \) is a vector of arbitrary length along the z-axis of the coordinate system which is aligned to the centre of gravity in negative direction. Since the skeletal approach allows for angle investigation in different distances r and therefore angle distributions along the course of a branch, the coordinates of s are dependent on r. The point s
with
can be determined by calculating the intersection point of the surface of a sphere with radius r and centre b and the line segment with \( \overrightarrow {{\left| {bp_{1} } \right|}} < r < \overrightarrow {{\left| {bp_{2} } \right|}} \) where p 1 is the first point defining the branch segment and p 2 is the second respectively (Fig. 2). Finally, the branch angle φ is determined by the scalar product of \( \overrightarrow {bs} \) and \( \overrightarrow {z} \) (Eq. 5).
For each branch of each skeletonized tree, we calculated branch angles in absolute distances ranging from 0.5 to 10 m in steps of 0.5 m. In addition, we determined angles ranging from 0.5 to 5.0 m measurement distance in reverse direction, meaning from the branch’s top to its base in 0.5-m steps as well and lastly, we calculated angles in distances relative to the branch in steps of 10 % of the branch’s length.

Angle between branch base b and point on the branch in distance r
Branch length
We defined two different lengths of a branch. One described simply by the straight distance l s between the branch base b and its end e (Eq. 6).
The second length l c we defined achieves higher accuracy by following the curvature of the branch and summing up the lengths of all n segments belonging to the branch’s skeletal structure as shown in Eq. (7).
Volume calculations
In order to estimate the spatial occupation of branches within the crown, additional points in a distance of 10 cm or less from each other were created within every branch segment to achieve higher density and homogeneity of the skeleton’s defining points. Then, every member of the TLS point cloud was assigned to its nearest neighbour of the enriched skeleton point crowd by application of the kd-tree-based nearest neighbour algorithm described by Maneewongvatana and Mount (1999). In computational geometry, an α-shape is a generalization of the convex hull of a point set based on Delaunay triangulation. The so-called α-value is a parameter that controls the level of detail in which the calculated shape represents the point cloud. It can range from \( \alpha \to 0 \) which yields the point cloud itself to \( \alpha \to \infty \) yielding the convex hull of the point cloud. For detailed explanation see Edelsbrunner and Mücke (1994).
The specific point cloud for every single branch resulting from the nearest neighbour assignment was used for three-dimensional α-shape calculations. The volume V α of the α-shape of a branch was calculated by summing up the volumes of all n tetrahedrons being part of the α-shape as shown in Fig. 3 and Eq. (8).

Tetrahedron edges as in Eq. (8)
Since we calculated the α-shape for several alpha values, ranging from coarse (convex hull: α = ∞) to fine (α = 0.25 m), it is possible to estimate the three-dimensional hull and spatial requirement of a tree’s branches including smaller branches, twigs and foliage originating from it in different resolutions (Fig. 4).

Original point cloud (l) and three-dimensional model of the same tree assembled by its single branch α-shapes (r)
To estimate the crown volume of a given tree’s point cloud, a three-dimensional α-shape of all members of the cloud with a z-value higher than that of the point where the crown commences was calculated. Its volume was determined analogue to the calculation of a single branch volume. It represents the growing space occupied by the tree, respectively its branches.
Results
The results summarized in Table 3 refer to all branches that were alive and exceeded 5 % of their tree’s height in length at the time the scans were made. For spruce, our study reveals significantly longer branches and significantly higher crown volumes in the mixed stand when compared to the pure stand. In case of European beech, individuals growing in mixture show flatter branch angles, more distinct ramification, higher crown volumes and a lower proportion of single branch volumes within the crown. In order to take the grouping of the branches on tree level into account, we applied a linear mixed effects model (lme) whenever it was appropriate.
Branch angles
The branch angles, measured from branch base to branch end, over the entirety of all branches show significant differences between pure and mixed stands for European beech (Fig. 5). The mean angles of Norway spruce both in the pure stand and the mixed stand are about 89° on average (Table 3). European beech’s branch angles average at 138.6° in pure stands and 128.5° in mixed stands (p < 0.001). Moreover, the results for European beech present significant differences between pure and mixed stands for branch angles over the whole course of a branch in all measurement distances relative to the total branch length. While the angles derived from a lme stay on a relatively constant level at about 140° in pure stands, in mixed stand they decline from about 133.8° to 128.5° with increasing measuring distance (Fig. 6). Both pure and mixed stand branch angles flatten towards the branch end. To illustrate this, the branch angles of beech’s crown periphery measured in reversed direction, i.e. from branch top in direction to the branch base in absolute measurement distance intervals of 0.5 m, are presented in Fig. 7.

All branch angles measured from branch base to branch top for Norway spruce and European beech in pure and mixed stands. Differences between pure and mixed stands are highly significant for European beech (linear model, p < 0.001). Sp Norway spruce in pure stand, Sm Norway spruce in mixed stand, Bp European beech in pure stand, Bm European beech in mixed stand

Mean relative angles and single standard error bars for European beech in pure and mixed stands derived from a linear mixed effects model. Trees growing in pure stands show significantly higher values than trees growing in mixture over the whole course of a branch. While the angles stay on a relatively constant level in pure stands, they decline with increasing measurement distance. p values range from 0.047 (worst) to 0.002 (best)

Absolute branch angle means and double standard error bars of European beech measured from branch end towards branch base. Means and errors are derived from a linear mixed effects model. p values range from 0.002 (worst) to <0.001 (best)
Branch length and branch bending
Norway spruce features significantly distinct branch lengths l s . In pure stands the mean measured branch length is 2.16 m compared to 2.78 m in mixed stands (p = 0.047; see Table 3). For European beech, the results show no significant differences over the entirety of all branches (p = 0.070) as well as within the various branch orders.
Regarding the branch bending (BB), here defined as the quotient l s /l c of branch length measured from branch base to top and the branch length revealed by the sum of all measured branch segments belonging to a branch, Norway spruce features mean values of about 96 × 10−2 in pure stands and in mixed stands without significant differences (p = 0.556). European beech however shows significantly distinct values of 96.21 × 10−2 in pure stand and 94.39 × 10−2 in mixed stand, meaning that branches in mixed stands are less straight (p = 0.002).
Length sum, branch count and branch orders
For Norway spruce the number of measured branches fulfilling the above-mentioned criteria averages 20.1 branches in pure and 26.0 branches in mixed stands resulting in no significant difference (p = 0.261). European beech, however, with 18.6 branches in the pure stand compared to 35.8 branches in the mixed stand, showed significantly more measureable branches over all in mixed stands (p < 0.001; Table 3). Especially branches of the second, third and fourth orders are significantly more numerous in mixed stands as shown in Fig. 8. While there is a tendency but no significant difference in the average branch length of European beech in pure and mixed stand (p = 0.070; see Table 3), due to the higher quantity of branches the sum branch lengths (SBL) differs significantly between pure (89.3 m) and mixed (153.8 m) stands (p < 0.001).

Mean number of branches and double standard error for European beech in pure and mixed stands by branch order. In mixture, European beech shows significantly more branches of the second, third and fourth order (p 2 < 0.001, p 3 = 0.011, p 4 = 0.004)
Volume of the α-shape of individual branches and the crown as a whole
Crown volumes (CV), based on volume calculations of α-shapes (α = 0.25 m) of the individual tree point clouds, were significantly higher in mixed stands for both Norway spruce and European beech as presented in Table 3. In the case of Norway spruce, the differences in crown volumes between pure and mixed stands are mainly caused by the greater average branch lengths in mixed stands compared to pure stands (R 2 = 0.34, p = 0.005) (see Fig. 9). Branch angles of spruce appear to have no significant influence on crown volume. The differences in crown volumes of European beech on the other hand are related to the differences of the branch angles (R 2 = 0.36, p = 0.002) (see Fig. 10), while branch length shows no significant influence.

Relation between average branch length and crown volume derived from α-shapes with α = 0.25 m for Norway spruce. \( \overline{{V_{\alpha } }} = - 4 5. 0 { + 28} . 1 *l_{s} \), R 2 = 0.34, p = 0.005, shaded area marks the 95 % confidence region of the regression line

Relation between mean branch angle measured from branch base to branch end per tree and crown volume derived from α-shape with α = 0.25 m for Europen beech. For better readability, angles were subtracted from 180°. Here, increasing angles mean flatter branches resulting in a larger crown volume. \( \overline{{V_{\alpha } }} = { - 42} . 7 { + 1} . 9 * ( 1 8 0 { - }\varphi ) \), R 2 = 0.36, p = 0.002, shaded area marks the 95 % confidence region of the regression line
The relative branch volume (RBV), i.e. the branch volume in relation to the tree’s crown volume, shows no significant differences between pure and mixed stands when a linear mixed effects model is applied. However, the logarithmic relative branch volume (RLBV) is significantly higher in pure stands for European beech for both standard linear models as well as linear mixed effects models (p lme = 0.005; Table 3). Since due to occlusion caused by foliage, the skeletonization of Norway spruce proved to be difficult in the higher crown regions, branch volume calculations did not yield reliable results.

Schematic drawing of crown structures of European beech in pure (left) and mixed (right) stands to illustrate the different branching structures. While individuals growing in pure stands develop steeper branches, those growing in mixture with Norway spruce show flatter branches with a higher number of smaller branches originating from them
Discussion
How different tree crowns may develop under intra- and interspecific competition is illustrated in Fig. 11 by the example of the results we found for European beech. Tree crown development plays an essential role during stand development of pure and even more of mixed-species forests. In order to explain the relevance of crown development and intraspecific plasticity in the interaction between two tree species during stand development, Fig. 12 illustrates the feedback between their functioning, structure, and local environment within the stand. The distinction between functioning (e.g., growth), structure formation (e.g., crown extension, crown space filling) and environment (e.g., light supply in the canopy, water supply below ground) is made in accordance to the ecosystem approach by Hari (1985, p 28) and Pretzsch (2009, p 226). The trees can modify their local environment within the stand via their structure and their functioning.

Feedback between stand structure, environmental conditions and tree functioning in a two-specific mixed forest stand. Further explanation is given in the text
The functioning (e.g., growth, partitioning) can change the tree structure (e.g., crown width, branch angles) and the resulting tree and stand structure affects the local environmental conditions for tree and stand growth (slow feedback loop functioning → structure → environment → functioning, represented by bold arrows). The extinction of light in the stand canopy results from the tree and stand structure which are analyzed in this study. The tree functioning can also directly affect the environment (rapid feedback loop functioning → environment → functioning, represented by bold arrow and opposite thin arrow). Supply or efficiency of soil water and nutrient exploitation may be reduced or enhanced e.g., if shallow rooting species and deeper rooting neighbours complement each other (Grams et al. 2002; Bolte and Villanueva 2006).
Our results on the variability of crown structure concern the slow feedback loop functioning → structure → environment → functioning. The crown development revealed for mixed stands is beyond the range known from pure stands and indicates crown properties emerging in interspecific environment.
The principle differences in the crown properties between spruces and beeches in mono-specific stands were already subject of detailed analysis (Assmann 1961; von Droste zu Hülshoff 1969; Pretzsch and Schütze 2005) and we just discuss additional details like species-specific branch angles, numbers and volumes. In contrast, the intra-specific crown plasticity of both species in mixed versus pure stands which we focused on was hardly analyzed so far (Purves et al. 2007; Richards et al. 2010).
The shallower branch angle, further crown extension, and denser crown space filling of beech in interspecific versus intraspecific neighbourhood may favour the competition of beech versus spruce in two different ways, via access to contested resources and hindering of a neighbour’s approach by effective shading and space occupation.
A higher structural crown plasticity and more complex canopy structure creates a higher variability of environmental conditions and in return triggers the plasticity at organ, crown or tree structure level for an optimal capture of resource or acclimation to the environment altered by competing neighbours. As a result of the feedback between growth, structure and local environmental conditions, structural variability can have an effect on interspecific competition, production ecology, structural diversity and biodiversity as shown in the following.
Methodological considerations on TLidar application
Our semi-manual skeletonization approach facilitates the retrieval of crown structures from imperfect TLS point cloud data and does so regardless of tree species. Even, due to occlusion and little scan positions, imperfect data as retrieved within actual forest stands yield usable tree architectures. However, occlusion, inhomogeneous point densities and noise within the dataset can still be an issue. Especially crown regions of Norway spruce which are higher than about 70 % of the tree’s height were hard to capture due to high occlusion caused by its foliage. Because of that, our method did not deliver reliable branch α-shapes and their volumes for Norway spruce sample trees. In fact the number of measurable branches in the higher crown regions is much lower than the actual number of branches of the tree, which in combination with the nearest neighbour assignment of the TLS point cloud members leads to a large overestimation of the actual point cloud of a branch. The establishment of more scan positions and the utilization of a more advanced TLS-device such as a full waveform scanner may help to overcome this problem. European beech on the other hand, if scanned outside the vegetation period under foliage free condition, proves to be a lot less problematic in that regard and yields viable tree architectures even in the higher crown regions.
Another issue that needs further investigation is the optimal α-value for the hull and volume calculations. The higher the chosen value the coarser the hull, which means that noisy point cloud members lying outside the actual crown or branch gain larger influence on the resulting volume calculation. On the other hand, α-values that are too small can lead to an underestimation of the actual crown or branch volume since holes in the point cloud data that are caused by occlusion are omitted in the hull calculation (Edelsbrunner and Mücke 1994). Therefore, depending on the quality of the dataset and the applied scan resolution, the range of reasonable α-values can vary. In our case, an α-value of 0.25 m provided a practicable trade-off between over- and underestimation of the actual growing space while ensuring that the potential point density in an assumed maximum measurement distance of 50 m is still sufficient for every α-shape facet.
Intraspecific variation of morphological traits in mixed-species versus pure stands
The longevity of forests enables a particular continuous formation of complex crown and stand structure. Due to uniform crown structure the canopy as a whole is rather homogeneous in pure stands but can become much more heterogeneous in mixed stands, e.g., when species such as spruce and beech with different compensation points of photosynthesis establish in multiple layers.
In order to avoid a mixing of merely size- and density-dependent changes in tree allometry and architecture with reactions triggered by an interspecific environment (Poorter et al. 2012), we selected trees with rather similar tree size in terms of stem diameter at breast height and local competition (Table 2). The structural traits our method reveals were presented in isolation in the “Results” section but are discussed in connexion with each other because of their close mechanical and physiological relationship.
The characteristic branch angles we report do not significantly differ between the pure and the mixed stand for Norway spruce. European beech, however, turned out to be much more variable in branch angles and lateral branch extension in mixed versus pure stands. This contributes to an increase in growing space (see Fig. 10) and may explain why under interspecific competition with spruce, beech is able to penetrate and occupy crown space very efficiently with low biomass investments (Pretzsch and Schütze 2005).
Regarding branch length and branch bending we found that compared to the pure stand, Norway spruce in mixture with European beech developed longer branches, thus resulting in more voluminous crowns. European beech shows no difference in branch lengths. However, it demonstrates its plasticity by growing branches that are less straight in mixture.
The number of branches, their distribution among branch orders and the length sum of branches within a crown for Norway spruce in a pure stand does not differ significantly from spruces in a mixed stand. European beech again shows its high crown plasticity by developing more branches, especially in the second, third and fourth branch order resulting in considerably more ramified crowns. While the branch length does not change, beech’s increased number of branches in mixed stand leads to significantly higher sums of branch lengths within the crown, contributing to a considerably denser filling of the growing space and increased fractal scaling of crown surface structure.
Tree crowns as described by Oldemann (1990), Purves et al. (2007) and Roloff (2001) lie somewhere in the continuum between the borderline cases of an umbrella-like crown with the whole leave surface area allocated close to the convex hull (surface dimension n = 2) and a broom-like crown with leave surface area distributed all over the crown space (surface dimension n = 3) (Zeide 1998). Species mixing can obviously modify the fractal dimension of the crown surface area and leaf area from a lower space filling (Euclidian scaling) towards higher space filling of the crown volume (fractal scaling) (Pretzsch and Dieler 2012). Concerning crown volume and the space that individual branches occupy within the crown, Norway spruce features larger crowns in mixture. The crown volumes of European beech too are significantly larger in the mixed stand. The average share of a single branch on beech’s crown volume however is smaller in the mixed stand. This may be explained through the finding that the larger overall crown volume in mixture is outweighed by the higher number of smaller branches when compared to the pure stand.
The considered higher crown plasticity in mixed versus pure stands is a emergent property which is crucial for better analyzing, understanding and modelling of stand dynamics (Purves et al. 2007; Richards et al. 2010). The latter applies in particular to mixed species forests where the structural as well as functional heterogeneity are much higher compared to pure stands and trigger manifold space occupation strategies, plant forms and shapes. Spruce and beech represent rather different species in terms of functional and structural traits: spruce is a quick, vertically oriented but rather unplastic species, while beech is a slow, rather laterally and very plastic one. Combination of such complementary species means a significant change of the canopy structure compared with pure stands and might trigger the particular broad intraspecific plasticity not known from pure stands. However, other common species combinations, e.g., oak/beech, pine/beech, eucalypt/acacia, populus/white spruce, represent similar structural and functional differences and might trigger comparable crown plasticity in inter- versus intraspecific environments (e.g., Bauhus et al. 2004).
Relevance of the structural crown variability for the growth performance of mixed versus pure stands
Crown structures at the individual tree level constitute a key aspect in the mechanisms of inter- and intraspecific interaction and its impact on a successful exploitation of potentially available environmental resources. Above ground, differences in growth and yield between pure and mixed stands can be caused by the occupation of more canopy space, by variations in the utilization efficiency of the given space or a combination of both. Since organs such as branches, twigs and leaves constitute a considerable share of the productivity; structural differences may presumably not only influence the filling of canopy space but crown efficiency (biomass production per unit of crown projection area and year) and competitiveness in resource exploitation as well (Matyssek et al. 2005).
The plasticity in lateral crown expansion prevents spruce from being outcompeted by beech, drives it into higher canopy layers where light can be captured more efficiently due to less shading. Latter reaction may explain the overyielding of spruce in mixture with beech (Pretzsch et al. 2010). Combination of both species can finally result in an average overyielding of about 20 % compared with pure stands (Pretzsch and Schütze 2009).
While our results for Norway spruce, apart from longer branches and higher crown volume in the mixed stand, show no statistically significant differences in structural crown parameters between pure and mixed stands, an increase in crown efficiency is likely at least partially caused by structural differences within the crown on branch or smaller level. European beech on the other hand is known to feature no considerable increase of crown efficiency when grown in mixture on our experimental plot. In fact, the crown efficiency slightly decreases (Pretzsch and Schütze 2009). As European beech does not profit from mixture-induced improvements of soil properties as much, the overyielding is presumably directly related to the differing structural crown properties we found. Since the actual distribution and availability of resources within the canopy space is hard to measure, space serves as an abstraction for above-ground resources. According to Grams and Lüttge (2011) space may even be considered a resource itself. In pure stands under intraspecific competition, potential niches and gaps in canopy space are heavily contested. Larger crowns, higher branch ramification, higher branch bending as well as flatter branch angles and more branches overall lead to a denser filling of the available canopy space, thus enhancing space occupation efficiency, adaption and multi-layering which enhances light interception and diameter growth (Binkley et al. 2011) and contributes to the frequently reported overyielding of beech in mixed versus pure stands (Pretzsch 2009; Pretzsch et al. 2010). The space occupation and suppression of neighbouring species reflect the competitive strength of beech and explain its domination in most Central European mixed stands (Fischer 1995).
Structural heterogeneity as component of biodiversity
Apart from productivity, structural crown properties also play an important role in habitat formation and its impact on biodiversity. The habitat heterogeneity hypothesis assumes that more complex habitats likely induce increased species diversity as they provide more niches and means to utilize potential environmental resources (e.g. MacArthur and MacArthur 1961; Bazzaz 1975). The distribution and interaction of animal species are often notably influenced by the plant community of a habitat which defines the majority of the physical environmental structure (Lawton 1983; McCoy and Bell 1991). In our case, more complex habitats is equivalent to mixed stands due to their higher complexity achieved by two instead of one main tree species and by more complex crown structures of European beech when grown in mixture with Norway spruce. According to Tews et al. (2004), the majority of studies about habitat heterogeneity and its relation to animal species diversity found a positive correlation with plant species diversity or structural complexity defined by parameters such as branch angles and ramification. Therefore, not only the mixture of Norway spruce and European beech in itself but also its impact on crown structures on branch or even lower level may significantly enhance animal species diversity.
Conclusions
A new method using terrestrial LiDAR in combination with point cloud skeletonization and other methods such as alpha shape calculations to determine structural crown properties of trees independent of tree species was developed. This method was applied to sample trees of an experimental plot providing comparable trees of both Norway spruce and European beech in pure stand as well as in mixture with each other. The results of this study have shown that our method may constitute a valuable non-destructive tool to facilitate a deeper understanding of the intra- and interspecific interaction of trees and its impact on productivity and ecological traits, e.g., biodiversity in pure and mixed-species forest stands.
Whether, and to what extent, our results may be applied to other stands is subject to further research. The experimental plot SON 814 is considered highly productive for both species. It can be assumed that nutrition and soil properties are provided in sufficient magnitude for each of the sample trees, largely independent of competition. It is imaginable that inter- and intraspecific competition for the most part manifest in above-ground structural differences induced by the competition for light. Hence, it is conceivable that structural differences in terms of structural reactions on differing neighbour species are less distinct on stands which offer less optimal growth conditions where below-ground competition mechanisms or the general resource supply and soil properties become more of an issue.
The potential applications of our method are manifold. For instance, not only may structural crown properties be analyzed, but also the interaction between crowns and the occupation of canopy space of stand neighbours on individual tree level. The method is currently applied on plots along an ecological gradient to analyze how the intraspecific crown plasticity is modified by site conditions.
References
Assmann E (1961) Waldertragskunde. Organische Produktion, Struktur, Zuwachs und Ertrag von Waldbeständen. BLV Verlagsgesellschaft, München
Assmann E (1970) The principles of forest yield study. Pergamon Press, Oxford
Badoux E (1946) Krone und Zuwachs. Mitt Schweiz Anst Forstl Versuchswesen 24:405–513
Bauhus J, van Winden AP, Nicotra AB (2004) Above-ground interactions and productivity in mixed-species plantations of Acacia mearnsii and Eucalyptus globulus. Can J For Res 34:686–694
Bazzaz FA (1975) Plant species diversity in old-field successional ecosystems in southern Illinois. Ecology 56:485–488
Binkley D, Stape JL, Ryan MG (2004) Thinking about efficiency of resource use in forests. For Ecol Manag 193:5–16
Binkley D, Campoe OC, Gspaltl M, Forrester DI (2011) Light absorption and use efficiency in forests: Why patterns differ for trees and stands. For Ecol Manage. doi:10.1016/j.foreco.2011.11.002
Bolte A, Villanueva I (2006) Interspecific competition impacts on the morphology and distribution of fine roots in European beech (Fagus sylvatica L.) and Norway spruce (Picea abies (L.) Karst.). Eur J For Res 125:15–26
Bremer M, Jochem A, Rutzinger M (2012) Comparison of branch extraction for deciduous single trees in leaf-on and leaf-off conditions—an eigenvector based approach for terrestrial laser scanning point clouds. EARSeL eProc 11(1):33–43
Bucksch AK (2011) Revealing the skeleton from imperfect point clouds. Dissertation, Delft University of Technology
Bucksch AK, Lindenbergh R, Menenti M (2010) SkelTre—Robust skeleton extraction from imperfect point clouds. Vis Comput 26:1283–1300
Burger H (1939) Holz, Blattmenge und Zuwachs. Mitt Schweiz Anst Forstl Versuchswesen 1939–1953, vol 15–29
Côté JF, Fournier RA, Egli R (2011) An architectural model of trees to estimate forest structural attributes using terrestrial LiDAR. Environ Model Softw 26:761–777
Edelsbrunner H, Mücke EP (1994) Three-Dimensional Alpha Shapes. ACM Transact Graph 13:43–72
Enquist BJ, Brown JH, West GB (1998) Allometric scaling of plant energetics and population density. Nature 395:163–165
Fischer A (1995) Forstliche Vegetationskunde. Pareys Studientexte 82. Blackwell Wissenschaft, Berlin, Wien
Grams TEE, Lüttge U (2011) Space as a resource. Prog Bot 72:349–370
Grams TEE, Kozovits AR, Winkler JB, Sommerkorn M, Blaschke H, Häberle K-H, Matyssek R (2002) Quantifying competitiveness in woody plants. Plant Biol 4:153–158
Hari P (1985) Theoretical aspects of eco-physiolocigal research. In: Tigerstedt PMA, Puttonen P, Koski V (eds) Crop physiology of forest trees. Helsinki Univ Press, Helsinki, pp 21–30 336p
Hilker T, van Leeuwen M, Coops NC, Wulder MA, Newnham GJ, Jupp DLB, Culvenor DS (2010) Comparing canopy metrics derived from terrestrial and airborne laser scanning in a Douglas-fir dominated forest stand. Trees 24:819–832
Huang P, Pretzsch H (2010) Using terrestrial laser scanner for estimating leaf areas of individual trees in a conifer forest. Trees 24:609–619
Lawton JH (1983) Plant architecture and the diversity of phytophagous insects. Ann Rev Entomol 28:23–39
MacArthur RH, MacArthur JW (1961) On bird species diversity. Ecology 42:594–598
Maneewongvatana S, Mount D (1999) It’s okay to be skinny, if your friends are fat. In: Proceedings of the 4th Annual CGC Workshop on Computational Geometry
Matyssek R, Agerer R, Ernst D, Munch JC, Oßwald W, Pretzsch H, Priesack E, Schnyder H, Treutter D (2005) The Plant’s Capacity in Regulating Resource Demand. Plant Physiol 7:560–580
McCoy ED, Bell SS (1991) Habitat structure: the evolution and diversification of a complex topic. In: Bell SS, McCoy ED, Mushinsky HR (eds) Habitat structure: the physical arrangement of objects in space. London, Chapman & Hall, pp 3–27
Niklas KJ (1994) Plant Allometry. Univ Chicago Press, Chicago
Oldemann RAA (1990) Forests: elements of Silvology. Springer, Berlin
Poorter H, Niklas KJ, Reich PB, Oleksyn J, Poot P, Mommer L (2012) Biomass allocation to leaves, stems and roots: meta-analysis of interspecific variation and environmental control. New Phytol 193:30–50
Pretzsch H (1992) Modellierung der Kronenkonkurrenz von Fichte und Buche in Rein- und Mischbeständen. AFJZ 163(11/12):203–213
Pretzsch H (2003) Diversität und Produktivität von Wäldern. AFJZ 174:88–98
Pretzsch H (2006) Species-specific allometric scaling under self-thinning. Evidence from long-term plots in forest stands. Oecologia 146:572–583
Pretzsch H (2009) Forest dynamics, growth and yield: From measurement to model. Springer, Berlin
Pretzsch H, Dieler J (2012) Evidence of variant intra- and interspecific scaling of tree crown structure and relevance for allometric theory. Oecologia. doi:10.1007/s00442-011-2240-5
Pretzsch H, Schütze G (2005) Crown allometry and growing space efficiency of Norway spruce (Picea abies (L.) Karst.) and European beech (Fagus sylvatica L.) in pure and mixed stands. Plant Biol 7:628–639
Pretzsch H, Schütze G (2009) Transgressive overyielding in mixed compared with pure stands of Norway spruce and European beech in Central Europe: evidence on stand level and explanation on individual tree level. Eur J For Res 128:183–204
Pretzsch H, Seifert S, Huang P (2011) Beitrag des terrestrischen Laserscannings zur Erfassung der Struktur von Baumkronen. Schweiz Z Forstwes 162:186–194
Pretzsch H, Block J, Dieler J, Dong PH, Kohnle U, Nagel J, Spellmann H, Zingg A (2010) Comparison between the productivity of pure and mixed stands of Norway spruce and European beech along an ecological gradient. Ann For Sci 67. doi:10.1051/forest/2010037
Price CA, Gilooly JF, Allen AP, Weitz JS, Niklas KJ (2010) The metabolic theory of ecology: prospects and challenges for plant biology. New Phytol 188:696–710
Purves DW, Lichstein JW, Pacala SW (2007) Crown plasticity and competition for canopy space: a new spatially implicit model parameterized for 250 North American tree species. PLoS ONE 2:e870. doi:10.1371/journal.pone.0000870
Ramachandran P, Varoquaux G (2008) Mayavi: making 3D data visualization reusable. In: Varoquaux G, Vaught T, Millman J (eds) Proceedings of the 7th Python in Science Conference. Pasadena, CA USA, pp 51–56
Richards AE, Forrester DI, Bauhus J, Scherer-Lorenzen M (2010) The influence of mixed tree plantations on the nutrition of individual species: a review. Tree Physiol 30:1192–1208
Röhle H, Huber W (1985) Untersuchungen zur Methode der Ablotung von Kronenradien und der Berechnung von Kronengrundflächen. Forstarchiv 56:238–243
Roloff A (2001) Baumkronen. Verständnis und praktische Bedeutung eines komplexen Natur-phänomens, Ulmer
Seifert T (2003) Integration von Holzqualität und Holzsortierung in behandlungssensitive Waldwachstumsmodelle. Dissertation, Technical University of Munich
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92
van Leeuwen M, Hilker T, Coops NC, Frazer G, Wulder MA, Newnham GJ, Culvenor DS (2011) Assessment of standing wood and fiber quality using ground and airborne laser scanning: a review. For Ecol Manag 261:1467–1478
von Droste zu Hülshoff B (1969) Struktur und Biomasse eines Fichtenbestandes auf Grund einer Dimensionsanalyse an oberirdischen Baumorganen. Ph.D thesis, LMU München, 209 p
Vosselman G, Maas HG (2010) Airborne and terrestrial laser scanning. Whittles Publishing, Dunbeath
Walter H (1931) Die Hydratur der Pflanzen und ihre physiologisch-ökologische Bedeutung. Gustav Fischer Verlag, Jena
West GB, Enquist BJ, Brown JH (2009) A general quantitative theory of forest structure and dynamics. PNAS 106:7040–7045
Wilhelmsson L, Arlinger J, Spångberg K, Lundqvist SO, Grahn T, Hedenberg Ö, Olsson L (2002) Models for predicting wood properties in Stems of picea abies and pinus sylvestris in Sweden. Scand J For Res 17:330–350
Zeide B (1998) Fractal analysis of foliage distribution in loblolly pine crowns. Can J For Res 28:106–114
Zobel B, van Buijtenen J (1989) Wood Variation—its causes and control. Springer, Berlin
Acknowledgments
We thank the Bavarian State Ministry for Nutrition, Agriculture and Forestry for permanent support of the project W 07 “Long-term experimental plots for forest growth and yield research” (# 781-20400-2012). Thanks are also due to Dr. Peter Biber for advice on the statistical analysis, Gerhard Schütze for participating in the field work and assistance in the data preparation, Ottilie Arz for assistance in field work, the skeletonization work as well as artwork creation in the course of her master thesis and reviewers for their constructive criticism.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by R. Matyssek.
Rights and permissions
About this article
Cite this article
Bayer, D., Seifert, S. & Pretzsch, H. Structural crown properties of Norway spruce (Picea abies [L.] Karst.) and European beech (Fagus sylvatica [L.]) in mixed versus pure stands revealed by terrestrial laser scanning. Trees 27, 1035–1047 (2013). https://doi.org/10.1007/s00468-013-0854-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-013-0854-4