
Abstract
This study tests whether crown and stem development in Norway spruce could be described using a modified profile theory. 29 trees from three age-groups (25, 67, 86) with different treatments (unthinned, normally and intensively thinned) were destructively sampled. Crown ratio and crown length varied between age groups and treatments. Crown width was positively correlated with crown length, but branch length along the crown depended on tree age and growing space. Foliage mass density peaked at a relative crown height of 50–70% in middle-aged and mature stands, while young crowns were densest and widest at the base. Foliage mass was predictable from branch and stem cross-sectional area, provided the distance from the top was included. The ratio of foliage mass to branch cross-sectional area increased for 2–4 m down from the tip of the crown, then started to decrease. The relationship between cumulative foliage mass and stem cross-sectional area was non-linear along the stem in the upper crown, but the ratio of cumulative branch to stem cross-sectional area was linear. Trees in the mature and unthinned stands had more cross-sectional area in branches relative to stems than in the young and thinned stands. We conclude that the profile theory needs modification regarding (1) crown shape which varies with age and growing space, and (2) the ratio of foliage mass to branch area which varies along the stem. Both aspects emphasise the need to include impacts of disuse of sapwood pipes in models of crown and stem development.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The regular structure of trees, based on the idea that the shape of the organism is stable during the growing process, is empirically predicted by several allometric equations (e.g. Braekke 1986; Marklund 1987, 1988; Kershaw and Maguire 1995; Ingerslev and Hallbäcken 1999; Johansson 1999; West et al. 1999; Turner et al. 2000; Grote 2002; Ilomäki et al. 2004). A theory of the balanced interaction between the tree components is, as well, widely proposed as a basis for predicting the growth allocation and structure of trees (Mäkelä et al. 2002). The crown vertical profile has been related to a balanced relationship between foliage and its supporting structure in several studies: Shinozaki et al. (1964a, 1964b) created the widely applied (e.g. Kaufmann and Troendle 1981; Waring et al. 1982; Oren et al. 1986a; Dvorak et al. 1996; Mäkelä and Vanninen 2001) pipe model theory, and Chiba et al. developed (1988) the theory of stem growth related to crown profile increment. Osawa et al. (1991) developed further this profile theory of tree growth, which was afterwards tested by Kershaw and Maguire (2000) for Western hemlock [ Tsuga heterophylla Raf. (Sarg.)], and Balsam fir [ Abies balsamea (L.) Mill.], and tested and applied by Mäkelä and Vanninen (2001) for Scots pine ( Pinus sylvestris L.).
The pipe model theory (Shinozaki et al. 1964a, 1964b) explains that there is a proportional relationship between foliage and sapwood area. When the tree grows taller and/or develops more foliage, a corresponding amount of new sapwood is formed in stems and branches. On the other hand, when crown rise occurs, a corresponding amount of sapwood pipes become disused, eventually accumulating as heartwood in the stems and branches. The profile theory (Chiba et al. 1988; Osawa et al. 1991) assumes that active sapwood pipes become disused at the same specific rate as foliage turns over, that the ratio of sapwood cross-sectional area (or area increment) to the foliage above is constant over the entire stem, and that the vertical foliage mass distribution is constant over time, only moving upward with height growth and crown rise.
When modified appropriately on the basis of empirical results (Mäkelä and Vanninen 2001) and combined with a carbon balance model, the pipe model and the profile theory have lead to a realistic description of structural growth in Scots pine (Mäkelä 2002). The carbon balance model predicts the development of foliage mass, tree height and crown base, while the vertical distribution of foliage mass and branch and stem cross-sectional area are derived using the profile theory. Empirical analysis showed that as hypothesized by Osawa et al. (1991), the overall shape of the vertical foliage density distribution could reasonably be approximated with a function that is constant over time and over a range of social positions, although its size varied a lot in both dimensions (Mäkelä and Vanninen 2001). However, the ratio of foliage mass to branch cross-sectional area is not constant over the stem, but increases from the top downwards before decreasing again from the middle crown (Oren et al. 1986a; Dvorak et al. 1996; Kershaw and Maguire 2000; Mäkelä and Vanninen 2001). This is consistent with the pipe model (Shinozaki et al. 1964a) and the hypothesis that active sapwood pipes have a longer life span than foliage and inactivate gradually, leading to “reuse” of old active pipes by new foliage (Nikinmaa 1992), but implies that the profile theory has to be modified (Mäkelä 2002).
The objective of this study is to investigate whether, and under what conditions, a similar modified profile theory could also be applicable to the moderately shade tolerant Norway spruce [ Picea abies (L.) Karst.]. Defining such conditions would allow us to describe the crown dynamics and predict the development of stem structure in Norway spruce using a model similar to that for Scots pine (Mäkelä 2002). To investigate this, we shall analyse the vertical foliage density distribution, and the vertical distribution of the foliage mass : sapwood area relationships, as well as the branch and stem area relationships along the stem, in Norway spruce trees of different ages, sizes, and competitive status.
Materials and methods
Materials
The data were collected at the end of the 2001 growing season. The material consists of trees from three stands (young, middle-aged and mature) in southern Finland (Table 1). The stands, located on mineral soil, were classified as the Oxalis type (Cajander 1949), representing fertile stands with H 100 ca. 27–33 m (Vuokila and Väliaho 1980). In the young stand, 5 trees were sampled from one unthinned plot, while in each of the older stands, a total of 12 trees were sampled from three plots with different thinning treatments, including unthinned control, normal commercial thinning, and intensive thinning (Table 2). A larger number of trees were sampled from the unthinned and normal thinned plots, because of greater size variation among these trees.
In the middle-aged and mature stands, the sample trees were approximately of the same size (diameter at breast height: n =24, F =3.10, P =0.07, tree height: n =24, F =3.13, P =0.07) (Table 2). The smallest trees were located in the unthinned plots, and largest in the thinned plots, of which the maximum tree size appeared to be in the normally thinned plot. The specific growing space (Hegyi 1974) of individual sample trees was most limited in the unthinned plots, and widest in the intensively thinned ones ( n =24, F =15.49, P <0.01) in the two older stands (no thinnings were available for the young stand).
Crown width was measured in two directions—the maximum width and the width perpendicular to it. Projected crown area was estimated as a circle, of which the diameter was the mean of these two measurements. The trees were felled and each branch in whorls (no internodal branches were considered) within the living crown were tallied and measured in the field for their distance from the treetop, and for diameter over bark at the stem junction immediately above the basal swelling, after which the tally branch quality (live, dead) was registered, and stem diameter over bark below each whorl was measured. Ten living sample branches were taken systematically throughout the living crown, such that the crown was divided into ten sections of equal length, and one living branch was taken from each section at random (no branches were available in the top section in the young stand).
The fresh sample branches were weighed in the field, put in plastic bags, and taken to the laboratory where their length and basal diameter in the horizontal and vertical directions were measured, and they were oven dried at 70°C for 48 h. The dry mass of needles and branch wood was determined. The cross-sectional area of all branches, which was calculated using the field data, was corrected for systematic measurement error using the laboratory data of the sample branches. The diameter of seven sample disks, from different heights of the tree (including the crown base), was measured for heartwood and sapwood separately.
Methods
The vertical distributions of branch length, branch wood mass and foliage mass were estimated using regression analysis on the sample branches, and applying the resulting models of the biomass components to all branches. Branch diameter (or cross-sectional area) and distance from the top were used as independent variables as they were measured from all branches. The models were fitted for trees in different stand age groups or thinning treatments separately; branch length was fitted for the thinning treatments and stand age groups, and branch wood mass and branch foliage mass were fitted for the age groups. The number of sample trees (Table 2) and sample branch observations was therefore different in each subgroup.
The following branch length model was determined by investigating the relationship between the measured branch distance from the top and the branch length in the sample branches:
where y is branch length, and x is the distance of the branch from the top of the tree, and a and b are parameters. To fit the model using linear regression, a new variable z was defined for a given value of b as
The parameter a of Eq. 1 was found using linear regression with zero intercept for a range of b -values. The combination of a and b was chosen that gave the smallest mean square error. The intercept of the linear regression could be assumed to be zero because it was not statistically significant ( P >0.05) in any of the regressions.
The branch or foliage dry mass was estimated separately for all individual tally branches, of which the location in the crown and diameter were measured. The total mass in a crown was calculated by summing up the predicted (wood or foliage) masses of all living branches. The vertical distribution of foliage and branch wood mass density was evaluated for each 20% length of the crown and the estimate placed at the middle point of the interval (Mäkelä and Vanninen 2001).
The following branch biomass model was selected by investigating the relationship between the measured independent variables (branch cross-sectional area, distance from tree top) and the dry mass of foliage and branches in the sample branches:
where y is either foliage mass ( W f) or branch mass ( W b), x is relative distance down the living crown (top=0, crown base=1), A b is branch cross-sectional area over bark, and a, b , c and n are parameters. The model was fitted for the three stands separately, using linear regression on the log-transform of the model when the exponent n was fixed. Several values of n were tested, and the one minimising the mean square error was chosen. In the model, the factor n accounts for the observations that the ratio of foliage (branch) mass to branch cross-sectional area increased rapidly when moving down from the top of the crown in older trees and reduced (for foliage) at base of the crown in young trees (Fig. 4 e, g). Parameters not significantly different from zero ( P >0.05) were not fitted. The stand-specific parameters were used when estimating the crown mass of individual trees of the stands. The parameter d was calculated separately for those individual trees for which it was found to be significantly different from the parameter estimated for the whole stand ( P <0.01). The comparison was done using dummy variables.
The linearity of dependence is analysed by log-transformation equation of the form:
where y is dependent variable, x is independent variable, and a and b are parameters. We assume that the relationship is linear if 0.95≤ b ≤1.05.
The pipe model theory postulates that the cumulative branch cross-sectional area is linearly proportional to stem cross-sectional area at any height along the stem, at least for the part of the stem where no considerable branch shedding has occurred (Shinozaki et al. 1964a, Fig. 5). In the case of analysing this relationship between cumulative branch area and stem cross-sectional area down the crown with linear regression, the number of data points depended on the number of whorls on the tree of a subgroup. In order to compare these subgroups, weights were introduced that gave observations in each of the subgroups (three stands, and two of them thinned with different thinning treatments) equal relative weights. The weights were defined as follows:
where N is the total number of observations and n i is the number of observations in the subgroup.
The number of sample trees varied between different stands and thinning treatments. Therefore, in the regression analysis of the whole data set, weights were introduced that gave observations in each of these subgroups (three stands, and two of them thinned with different thinning treatments) equal relative weights. Weighted regression was used when analysing maximum density of foliage and branch wood as a function of crown projected area, and branch wood mass, foliage mass, and branch cross-sectional area as a function of stem cross-sectional area at the tree level.
Tree height, slenderness, crown dimensions (length, height to crown base), biomass components, and branch length were compared between age, and thinning categories using one-way ANOVA, and further the means were compared using Tukey’s test.
Results
Tree height and height to crown base
The trunk height to the base of the living crown (m), and the length of the living crown (m) varied distinctly between the stands and the thinning treatments. Crown ratio (the ratio of crown length to tree height) decreased with tree age ( n =29, F =44.86, P <0.01) being greatest (91%) in the young stand. In the absolute scale, the crowns were shorter in the mature stand relative to the middle-aged stand ( n =29, F =33.47, P <0.01). In the middle-aged and mature stands, height to the crown base was significantly highest in the unthinned plots and lowest in the intensively thinned ones ( n =24, F =3.45, P <0.05). The trees in the unthinned plots were the most slender ( n =24, F =3.48, P <0.05) and their crowns were clearly shorter ( n =24, F =12.13, P <0.01) than in the normally and intensively thinned plots.
Projected crown area and branch length
Spruce crown is conical and therefore the crowns were widest at the very base of the living crown. The long crowned trees had the widest crowns ( n =29, R 2=0.82, P <0.01), but branch length along the crown depended considerably on the age and in the lower crown on the growth space of the tree. Eq. 1 explained 69–87% of the variation in branch length (cm), depending on the subset considered (Table 3).
In the mature stand, the sample branches were longest, and in the young stand clearly shortest on average in the crown ( n =29, F =60.78, P <0.01) (Fig. 1a, b). The differences were apparent in the modelled branches in the lower crown (Fig. 1b).

Branch length for all sample branches: a measured and b modelled (Eq. 1). Lines represent the young stand, pluses the middle-aged stand, and circles the mature stand
Between the different thinning treatments, there was significant homogeneity in the branch length particularly along the upper crown in the middle-aged and mature stands (no thinnings were available in the young stand) (Fig. 2a, b). For 5 m down the crown, the branch mean length was nearly equal in the sample branches, showing the maximum values in the unthinned and minimum in the normally thinned forest stands ( n =24, F =0.62, P >0.05). In the lower crown, the branches of unthinned forest stands were slightly shorter. The crowns tend to be longest and still vigorous at the bottom crown and therefore widest in the thinned plots.

Branch length for all sample branches: a measured and b modelled (Eq. 1). Thinning intensity in the plots of middle-aged and mature stands: circles represent unthinned plots pluses normally thinned and lines intensively thinned
Foliage and branch mass of sample branches
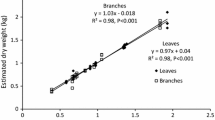
There was an almost linear relationship between foliage mass (g) and branch cross-sectional area (cm2) in the sample branches (Eq. 4), but the scatter around this relationship was very large (Fig. 3). The relationship between branch woody mass (g) and branch cross-sectional area (cm2) was clearly non-linear but with much less scatter than the foliage relationship (Fig. 3).

The relationship between biomass components, and branch cross-sectional area in the sample branches. Solid circles are branch woody mass, and circles are foliage mass
Foliage mass (g) on the sample branches increased from the tip of the crown downwards (Fig. 4a, b), starting to decrease in both older stands in the top half of the crown (Fig. 4b). The branch mass of sample branches also increased initially, but unlike in foliage, the increase continued down the crown or levelled off. Largest living branches, with dwindling but assimilative foliage, were located at the middle part of the living crown, particularly in the middle-aged and mature stands (Fig. 4c, d). In the young stand the dry mass of foliage and branch wood both peaked at the lower third of the crown (Fig. 4b, d). The cross-sectional area (mm2) of sample branches along the crown was similar to branch wood mass (Fig. 4c). Big dead branches were located at the bottom of the living crown, or below it. The size variation of branches of all qualities in individual trees was large.

Vertical distribution of foliage mass ( a, b) and branch cross-sectional area ( c, d) of the sample branches as a function of absolute and relative distance from treetop. Vertical distribution of the ratios of foliage mass ( e, f) and branch wood mass ( g, h) to branch cross-sectional area in the sample branches as a function of absolute and relative distance from treetop. Lines young stand, pluses middle-aged stand, and circles mature stand
The pipe model relation, i.e. ratio of foliage mass to branch cross-sectional area, clearly increased for 1 m down the crown, peaking at 2–4 m (Fig. 4e). After this peaking point, corresponding to ca. 10-year-old branches, the ratio started to decrease. In the lower crown (>10m) of the older trees the foliage-branch relationship stabilised and was nearly constant. Regardless of tree age, size, or stand thinnings, the pipe relation was fairly similar in all trees at the very top of the crown where the branch cross-sectional area can be assumed to consist of active pipes only. The similarity of the relationship across trees was clearer when taken with respect to absolute rather than relative crown length (Fig. 4e, f). The ratio of branch wood mass to branch cross-sectional area increased down the crown, and stabilised at 5 m (Fig. 4g, h). Young trees had not reached this tree height, so the ratio increased all the way down the crown.
The foliage and branch wood mass models (Eq. 3) gave good fits for foliage and branch biomass in trees. The coefficient of determination varied between 0.58 and 0.86 for foliage mass, and between 0.87 and 0.95 for branch wood mass (Table 4). The worst prediction was for foliage mass in the mature stand, probably because foliage mass varied considerably in the lower part of the crown between trees and branches, receding in shaded branches, while a few branches in good light had a vivid foliage mass even down in the crown.
Vertical density distribution of foliage and branch wood mass
The shape of the vertical foliage density distribution (g m−1) varied between stands of different ages (Fig. 5). In the young stand, the maximum density was reached at 30% relative height from the crown base upwards, whereas in the older stands, the peak of the foliage mass was at about 50–70% up the living crown. The living branch mass (g m−1) was densest at the middle of the crown or lower.

Vertical density distributions of foliage and living branch wood in selected sample trees as a function of relative and absolute height. Stands are: 1 young, 2 middle-aged, and 3 mature stand. Thinning intensity is: a unthinned, b normal thinning, and c intensive thinning. Thick line is most dominated and thin line is most suppressed tree in a plot
The foliage and branch wood mass were clearly greater in dominant than in suppressed trees (Fig. 5). In the intensively thinned plots, there were no really suppressed trees left. Suppressed trees of all ages tended to have shorter crowns with maximum foliage and branch wood density higher (in relative scale) along the stem than dominant trees.
The maximum density of foliage and branch wood (g m−1) correlated with the tree and crown dimensions—stem diameter at breast height and crown base, stem sapwood area at the crown base, tree height, crown length, and crown projected area (the linear relationship between crown projected area and the maximum foliage density in the whole data set: n =29, R 2=0.59, P <0.001, and the maximum branch wood density: n =29, R 2=0.74, P <0.001) (Fig. 6).

Relationship between maximum branch density in vertical direction (g m−1) and crown projected area (m2). Solid circles are branch woody mass, and circles are foliage mass
The ratio of branch cross-sectional area and foliage mass to stem cross-sectional area
The presumption of linearity in the pipe model was tested (Eq. 4) between the cumulative branch cross-sectional area (cm2) and stem cross-sectional area over bark (cm2) and they were found to correlate linearly throughout the crown in the whole data set ( n =987, R 2=0.95, P <0.001) (Fig. 7a). However, the ratio was decreasing in the lower crown, probably due to shedding of branches. In addition, the linear regression lines differed between the (age × thinning) categories, the slope of the line varying in the range 1.15–1.59, and the intercept in the range 0–29. No systematic trends related to age or thinning category could be detected, however.

Cumulative branch cross-sectional area as a function of stem cross-sectional area over bark a in the whole data set, and b defined as the upper 5 m of the living crown by tree sample trees. Lines represent a young tree (sample tree 33), pluses a middle-aged tree (tree 2), and circles a mature tree (tree 16)
In order to test further the pipe model ratio of branches to stem, the top 5 m of the living crown was analysed separately (for the young trees, this covered the entire crown), which generally in all tree size classes appears to be above the maximum foliage density in the crown. This would suggest that these branches are still growing and there is neither significant branch shedding nor heartwood development (Fig. 7b).
The ratio of cumulative branch cross-sectional area to stem cross-sectional area was larger in the top part than in the total crown, and the corresponding linear regression had zero intercept (Table 5). However, the ratio increased significantly with stand age, being greatest for the mature stand, as was demonstrated for the unthinned plots (no thinnings were available for the young stand) (Table 5) (Fig. 7b). The different thinning regimes also showed differences; the cumulative branch area relative to stem cross-sectional area was smallest when normally thinned and greatest when unthinned (Table 5). The average number of branches per whorl was four when unthinned or thinned.
The relationship between cumulative foliage mass (g) and stem cross-sectional area (cm2) was clearly non-linear (Eq. 4) in the whole data set, as well as in individual sample trees of different ages (Fig. 8a). The foliage mass in the uppermost crown was first accumulating slowly for 0.8 m, after which escalating with simultaneously multiplying twig surface (Fig. 8b). The upper crown foliage mass was accumulating most effectively in the mature trees. In the canopy-shaded branches lower down, the foliage mass accumulation levelled off (Fig. 8a).

Cumulative foliage mass as a function of stem cross-sectional area over bark a from the top to the bottom of the living crown, and b defined as the upper 2m of the living crown by tree sample trees. Lines represent a young tree (sample tree 33), pluses a middle-aged tree (tree 2), and circles a mature tree (tree 16)
Branch wood and foliage mass at tree level
The crowns had twice as much branch mass (kg) as foliage mass (kg) per cross-sectional area (cm2) of stem or sapwood at the crown base. Stem sapwood cross-sectional area (cm2) was a slightly better predictor of foliage mass (kg) of a tree than stem cross-sectional area (cm2), whereas the opposite was true of branch wood mass (kg) and branch cross-sectional area (cm2) (Table 6). Almost as good predictors for the branch dimensions were projected crown area or crown length and stem diameter at crown base or at breast height.
On the basis of the exponent in Eq. 4, foliage mass correlated linearly with stem cross-sectional area, and branch cross-sectional area correlated linearly with sapwood area. Foliage mass : stem cross-sectional area ratio seemed to be independent of stand age (in the whole data set n =29, F =0.12, P >0.01). The relationships between branch characteristics (wood mass and cross-sectional area) and the stem cross-sectional area at the crown base were non-linear (Fig. 9). In the young stand, branch wood weighed clearly less per total branch cross-sectional area or stem cross-sectional area at the crown base than the branches in the two older stands.

Relationships between crown characteristics (total foliage mass, branch wood mass, and branch cross-sectional area over bark), and stem cross-sectional area at the crown base. Pluses are observations under bark of the stem, and circles represent stem sapwood
The crowns were lighter (in the two older stands: n =24, F =9.18, P <0.01) and thin-branched ( n =24, F =12.96, P <0.01) in the unthinned plots. However, the ratios of foliage mass, branch wood mass, and total branch cross-sectional area to stem cross-sectional area under bark at the crown base, did not differ between the unthinned, normally thinned or intensively thinned sites (in the whole data set: n =29, P >0.01). The correlations were slightly weaker when analysed with respect to stem sapwood, probably because sapwood area is difficult to define accurately.
Discussion
The present results on the crown dimensions are largely similar to those of previous studies. The crown ratio is reducing and the crown radius is enlarging in ageing Norway spruces (Colin and Houllier 1992; Deleuze et al. 1996; Richardson et al. 2000). In addition, narrow growth space accelerates crown rise (Colin and Houllier 1992; Johansson 1992), but in the upper crown does not affect crown width, as also indicated by the results of Nilsson and Gemmel (1993), whereas lower down the availability of growth space seems to control crown horizontal growth and vertical shape. According to Deleuze et al. (1996) the crown in open-grown Norway spruce is expanded, while it remains narrower in trees of denser growth space. On the other hand, Gilmore and Seymour (1997) have reported of Abies balsamea that the crown radius down the crown is widest in short crowned suppressed trees and narrower in more dominating trees.
Unlike the fairly constant shaped Scots pine crown (Mäkelä and Vanninen 2001), the vertical profile of the foliage was found to be strongly age-dependent in Norway spruce (Hakkila 1989; Pulkkinen 1991). In young spruces (ca. 20 years.) the foliage is densest at the lower crown, at 30% of relative height, and after canopy closure in older trees focusing higher up (ca. 60–90 years at 50–70%). This can be understood by the ecological differences of the two species. Being as moderately shade-tolerant as Douglas fir (Maguire and Bennett 1996) or Balsam fir (Gilmore and Seymour 1997), Norway spruce maintains its lower branches much longer than Scots pine, when the shaded part of the crown is extending with tree age and size.
The maximal foliage density in Norway spruce crown does not increase with the increase of the projected crown area as clearly as in Scots pine (Mäkelä and Vanninen 2001). The dependence between the two is slightly weaker for Norway spruce, and the regression passes more clearly through the origin in Scots pine (Mäkelä and Vanninen 2001). In different aged Scots pine stands the crown is widest and densest at the middle of the crown. In the middle-aged and mature forest stands the Norway spruce crown is widest in the lower part of the crown, but the densest foliage mass is located in the upper parts of the crown; the canopy shading in the lower crown directly reduces foliage mass but has a delayed effect on the horizontal growth of the crown. Therefore, it is only in the young Norway spruce stands before canopy closure that the crown is widest and densest at approximately the same height at the bottom of the crown. In addition, the foliage mass in Scots pine is uniformly covering the crown surface (Mäkelä and Vanninen 2001), whereas in Norway spruce the foliage mass is covering the individual branches generating gaps in the crown surface. According to Kuuluvainen (1991), the crown range of Norway spruce does not indicate directly the foliage density of a tree; the mean needle density per certain crown-projected area is smaller in suppressed trees compared to more dominant ones.
Densely grown Norway spruce is capable of compensating for the lost lower crown needle mass by accumulating the growth in the upper crown, as regards both height growth relative to diameter growth (Mård 1996), and the growth of branches and foliage (Maguire and Bennett 1996; Gilmore and Seymour 1997). In the unthinned forest stands, the trees are more slender than in the thinned stands, and the total branch area above a certain stem area is greater in the upper crown, where the foliage mass of suppressed trees tends to be densest as well. The lower branches, which are very much alive in the thinned stands, are typically dying or dead in the unthinned stands.
According to Oren et al. (1986a), in different sites foliage will be distributed differently along the crown. On the other hand, the ratio of foliage mass to branch cross-sectional area seems independent of stocking density or dominance position (Oren et al. 1986a), but dependent on crown depth. It increases from the top to the middle of the crown (Oren et al. 1986a; Dvorak et al. 1996; Kershaw and Maguire 2000; Mäkelä and Vanninen 2001), peaking in Scots pine in the middle crown in all age groups (Mäkelä and Vanninen 2001). Similarly, the present results show that in Norway spruce the increase and culmination (at 2–4 m from tip of the crown downwards) seem to be found as a function of distance in all trees regardless of age or social position. In the lower crown, where foliage shedding and heartwood formation occur, the ratio of foliage mass to branch cross-sectional area decreases, and in older trees levels off being eventually nearly constant. This relationship can be utilised for predictions of foliage mass from branch and stem cross-sectional area, provided that the distance from the top is also included (e.g. Mäkelä 2002).
The upper, assimilatively important part of the crown (Oren et al. 1986b; Niinemets 1997) with active pipes and no shedding, seems to remain similar in shape and even size, once entirely formed in a young tree. The size is probably related to the life span of needles, which is about 10 years in southern Finland. Kaufmann and Troendle (1981) observed that the upper part of a larger tree has the same leaf area to stem sapwood ratio as the entire crown of a smaller conifer tree. The crown profile of Norway spruce can therefore be understood as consisting of an upper, active part with a fairly constant profile, and a lower part, also with a constant profile, but the ratio of the two evolving with tree age, and probably also varying with the social position of the tree.
The clearly non-linear relationship between cumulative foliage mass and stem cross-sectional area along the stem in the upper crown contradicts the original form of the pipe model (Shinozaki et al. 1964a). The form of the foliage mass – branch cross-sectional area relationship from the top of the crown downwards stimulated the choice of the equation of foliage mass. Vertically the foliage mass tends to be densest at the upper half of the crown with increased shoot surface, which promotes the vigour accumulation of foliage mass after minor foliage gain at the very top. Similar results of non-linear relationship between cumulative foliage mass and stem cross-sectional area along the stem have been reported earlier in Scots pine (Mäkelä and Vanninen 2001), Norway spruce (Oren et al. 1986a), and Balsam fir (Gilmore and Seymour 1997).
The ratio of cumulative branch cross-sectional area to stem cross-sectional area along the stem can be taken to represent the pipe model ratio between branch and stem pipes for the part of the stem where no significant branch shedding has taken place. Assuming that heartwood formation proceeds at the same rate in branches and stems, their ratio should be the same for dead and live whorls. Again, the present results are similar to earlier studies in Scots pine (Kaipiainen and Hari 1985; Mäkelä and Vanninen 2001) showing that a linear relationship exists between cumulative branch cross-sectional area and stem cross-sectional area along the stem. However, notable differences could be found between age classes and thinning treatments. Trees in the mature and unthinned stands had more cross-sectional area in branches relative to stems than the young and thinned stands. Interestingly, an opposite observation was made in Scots pine: younger trees and more dominant trees had a bigger branch : stem cross-sectional area ratio than older and more suppressed trees (Mäkelä and Vanninen 2001). The difference between the two species may be related to the ecological adaptation to shade in Norway spruce.
It is interesting that although there is considerable variation in crown shape, and the pipe model ratios are not constant along the stems, the tree-level measures of the pipe model ratios were found to be very stable. The ratio of foliage mass to early wood area at the crown base seems independent of site quality, age, or crown class (Eckmüllner and Sterba 2000). Furthermore, the foliage-sapwood ratio and the branch to stem cross-sectional area ratio at the tree level are almost the same as in Scots pine (Kaipiainen and Hari 1985; Mäkelä and Vanninen 2001). The main differences between the species therefore appear to be found in the crown dynamics, manifested by the crown profile, and can probably be traced back to differences in the photosynthetic system (Marek et al. 1989; Leverenz and Hinckley 1990; Leverenz 1996).
The prediction of foliage mass have generally not been able to improve by studies combining sapwood area with sapwood conductivity when compared with sapwood area only (e.g. Whitehead et al. 1984). This is actually in line with the original pipe model theory (Shinozaki et al. 1964a), which perceives the importance of the sapwood as conductive tissue, but also the aspect of mechanical support. The pipe model is essentially based on empirical observation not on theories of tree hydraulics or mechanical strength, although both aspects are likely to be important if an explanation is sought for the observed structure. In addition, heartwood formation is an essential part in the pipe model theory giving an extra dimension to the model of tree structure.
In order to predict the vertical profile of foliage mass using branch or stem cross-sectional area information or modelling the tree growth using vertical foliage distribution as an input, it is important to realise the structural differences between the age-independent, regular formed upper crown (<5m) and the lower part established by the foliage shedding. In order to obtain a reliable prediction of the vertical distribution of foliage mass, it is therefore crucial to include branch location in the crown as an independent variable in the model for branch foliage mass. Otherwise, it is conceivable to predict the foliage mass for upper and lower crown separately. Similarly, the pipe model assumptions about the relationship between total foliage and sapwood are practical, when modelling the growth in a young stand (<10 years). After canopy closure, the crown rise and foliage shedding is accelerating, and the growth of stem and branches is regulated increasingly by the life span of active sapwood pipes.
References
Braekke FH (1986) Distribution and yield of biomass from young Pinus sylvestris and Picea abies stands on drained and fertilized peatland. Scand J For Res 1:49–66
Cajander AK (1949) Forest types and their significance. Acta For Fenn 56
Chiba Y, Fujimori T, Kiyono Y (1988) Another interpretation of the profile diagram and its availability with consideration of the growth process of forest trees. J Jpn For Soc 70:245–254
Colin F, Houllier F (1992) Branchiness of Norway spruce in northeastern France: predicting the main crown characteristics from usual tree measurements. Ann Sci For 49:511–538
Deleuze C, Herve J-C, Colin F, Ribeyrolles L (1996) Modelling crown shape of Picea abies. Spacing effects. Can J For Res 26:1957–1966
Dvorak V, Opustilova M, Janous D (1996) Relation between leaf biomass and annual ring sapwood of Norway spruce according to needle age-class. Can J For Res 26:1822–1827
Eckmüllner O, Sterba H (2000) Crown condition, needle mass, and sapwood area relationship of Norway spruce ( Picea abies). Can J For Res 30:1646–1654
Gilmore DW, Seymour RS (1997) Crown architecture of Abies balsamea from four canopy positions in central Maine, USA. Tree Physiol 17:71–80
Grote R (2002) Foliage and branch biomass estimation of coniferous and deciduous tree species. Silva Fenn 36:779–788
Hakkila P (1989) Utilization of residual forest biomass. Springer, Berlin Heidelberg New York, pp 11–99
Hegyi F (1974) A simulation model for managing jack-pine stands. In: Fries J (ed) Growth models for tree and stand simulation. Royal College of Forestry, Stockholm, Sweden, pp 74–90
Ilomäki S, Nikinmaa E, Mäkelä A (2004) Crown rise due to competition drives biomass allocation in silver birch (Betula pendula L.). Can J For Res 33:2395–2404
Ingerslev M, Hallbäcken L (1999) Above ground biomass and nutrient distribution n a limed and fertilized Norway spruce ( Picea abies) plantation. II. Accumulation of biomass and nutrients. For Ecol Manage 119:21–38
Johansson K (1992) Effects of initial spacing on the stem and branch properties and graded quality of Picea abies [L.] Karst. Scand J For Res 7:503–514
Johansson T (1999) Biomass production of Norway spruce ( Picea abies [L.] Karst.) growing on abandoned farmland. Silva Fenn 33:261–280
Kaipiainen L, Hari P (1985) Consistencies in the structure of Scots pine. In: Tigerstedt PMA, Puttonen P, Koski V (eds) Crop physiology of forest trees. Proceedings of an International Conference on Managing Forest Trees as Cultivated Plants. Helsinki University Press, Helsinki, pp 31–37
Kaufmann MR, Troendle CA (1981) The relationship of leaf area and foliage biomass to sapwood conducting area in four subalpine forest tree species. For Sci 27:477–482
Kershaw JA, Maguire DA (1995) Crown structure in western hemlock, Douglas-fir, and grand fir in western Washington: trends in branch-level mass and leaf area. Can J For Res 25:1897–1912
Kershaw JA, Maguire DA (2000) Influence of vertical foliage structure on the distribution of stem cross-sectional area increment in western hemlock and balsam fir. For Sci 46:86–94
Kuuluvainen T (1991) Relationships between crown projected area and components of above-ground biomass in Norway spruce trees in even-aged stands: empirical results and their interpretation. For Ecol Manage 40:243–260
Leverenz JW (1996) Shade-shoot structure, photosynthetic performance in the field, and photosynthetic capacity of evergreen conifers. Tree Physiol 16:109–114
Leverenz JW, Hinckley TM (1990) Shoot structure, leaf area index and productivity of evergreen conifer stands. Tree Physiol 6:135–149
Maguire DA, Bennett WS (1996) Patterns in vertical distribution of foliage in young coastal Douglas-fir. Can J For Res 26:1991–2005
Mäkelä A (2002) Derivation of stem taper from the pipe theory in a carbon balance framework. Tree Physiol 22:891–905
Mäkelä A, Vanninen P (2001) Vertical structure of Scots pine crowns in different age and size classes. Trees 15:385–392
Mäkelä A, Givnish TJ, Berninger F, Buckley TN, Farquhar GD, Hari P (2002) Challenges and opportunities of the optimality approach in plant ecology. Silva Fenn 36:605–614
Mård H (1996) The influence of a birch shelter ( Betula spp.) on the growth of young stands of Picea abies. Scand J For Res 11:343–350
Marek M, Barták M, Pirochtová M (1989) Vertical topography of photosynthetic activity and crown structure in Norway spruce. Acta Sc Nat Brno 23:1–52
Marklund LG (1987) Biomass functions for Norway spruce ( Picea abies [L.] Karst.) in Sweden. Department for Forest Survey Report 43. Swedish University of Agricultural Sciences, Uppsala
Marklund LG (1988) Biomass functions for pine, spruce and birch in Sweden. Department for Forest Survey Report 45. Swedish University of Agricultural Sciences, Uppsala
Niinemets Ü (1997) Acclimation to low irradiance in Picea abies: influences of past and present light climate on foliage structure and function. Tree Physiol 17:723–732
Nikinmaa E (1992) Analyses of the growth of Scots pine; matching structure with function. Acta For Fenn 235:1–68
Nilsson U, Gemmel P (1993) Changes in growth and allocation of growth in young Pinus sylvestris and Picea abies due to competition. Scand J For Res 8:213–222
Oren R, Werk KS, Schulze E-D (1986a) Relationships between foliage and conducting xylem in Picea abies [L.] Karst. Trees 1:61–69
Oren R, Schulze E-D, Matyssek R, Zimmermann R (1986b) Estimating photosynthetic rate and annual carbon gain in conifers from specific leaf weight and leaf biomass. Oecologia 70:187–193
Osawa A, Ishizuka M, Kanazawa Y (1991) A profile theory of tree growth. For Ecol Manage 41:33–63
Pulkkinen P (1991) Crown form and harvest increment in pendulous Norway spruce. Silva Fenn 25:207–214
Richardson AD, Berlyn GP, Ashton PMS, Thadani R, Cameron IR (2000) Foliar plasticity of hybrid spruce in relation to crown position and stand age. Can J Bot 78:305–317
Shinozaki K, Yoda K, Hozumi K, Kira T (1964a) A quantitative analysis of plant form: the pipe model theory. I. Basic analyses. Jpn J Ecol 14:97–105
Shinozaki K, Yoda K, Hozumi K, Kira T (1964b) A quantitative analysis of plant form: the pipe model theory. II. Further evidence of the theory and its application in forest ecology. Jpn J Ecol 14:133–139
Turner DP, Acker SA, Means JE, Garman SL (2000) Assessing alternative allometric algorithms for estimating leaf area of Douglas-fir trees and stands. For Ecol Manage 126:61–76
Vuokila Y, Väliaho H (1980) Growth and yield models for conifer cultures in Finland. Comm Inst For Fenn 99.2:1–271
Waring RH, Schroeder PE, Oren R (1982) Application of the pipe model theory to predict canopy leaf area. Can J For Res 12:556–560
West GB, Brown JH, Enquist BJ (1999) A general model for the structure and allometry of plant vascular systems. Nature 400:664–667
Whitehead D, Edwards WRN, Jarvis PG (1984) Conducting sapwood area, foliage area, and permeability in mature trees of Picea sitchensis and Pinus contorta. Can J For Res 14:940–947
Acknowledgements
The work carried out in the Finnish Forest Research Institute (project no. 3283). The authors would like to acknowledge the project leader Risto Ojansuu and colleagues Mikko Kukkola and Jaakko Repola. The data were collected in co-operation with Harri Mäkinen, Tapio Nevalainen, Pasi Aatsinki and his team. A number of people participated in the laboratory work led by Pasi Aatsinki and Jouni Väisänen. The study was funded by the Foundation for Research of Natural Resources in Finland.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Kantola, A., Mäkelä, A. Crown development in Norway spruce [ Picea abies (L.) Karst.]. Trees 18, 408–421 (2004). https://doi.org/10.1007/s00468-004-0319-x
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00468-004-0319-x