
Abstract
Most of radial growth studies of Nothofagus pumilio (locally known as ‘lenga’) in Tierra del Fuego have focused on mature forests and climate relations, but little is known about growth dynamics after harvesting and even less after applying intermediate treatments. The objective of this chapter is to analyze the effect of thinning and pruning on radial growth of N. pumilio in Tierra del Fuego (Argentina) in the Aguas Blancas experimental long-term forest, which was harvested in 1967 and had different combinations of interventions in 1984–1985 and 1999. Three treatments were analyzed: (i) one thinning (1 T, thinned in 1984–85), (ii) two thinnings (2 T, thinned in 1984 and 1999), and (iii) two thinnings with pruning of 2/3 of the living crown (2 T + P, thinned in 1984 and 1999 with pruning). Using standard dendrochronological methods, ring widths were cross-dated, measured and the mean ring width (RW), basal area increment (BAI) and the corresponding percentages of change in growth (PGC) in 5-year windows were calculated. The three treatments showed significant increments in PGC in both RW (129%) and BAI (419%) after the 1984–1985 thinning. However, from 1999 there were significant differences in periodic growth between the three treatments with the highest RW and BAI in the thinned with pruning treatment. In these forests, dendrochronology emerges as a complementary method to quantify and to reconstruct the individual response of trees affected by silvicultural treatments.
Access provided by Autonomous University of Puebla. Download chapter PDF
Similar content being viewed by others


Keywords
1 Introduction
Forest management, whatever its objectives and modalities, substantially modifies the original forest structure. Silvicultural interventions produce changes in microclimatic conditions, such as increased solar radiation input, variations in the redistribution of the intercepted rainfall (Caldentey et al. 1998; Otero et al. 1998; Martínez-Pastur et al. 2011), changes in temperature, nutrient cycling (Thibodeau et al. 2000), moisture availability (Fayle 1983) and air humidity (Barg and Edmonds 1999). These changes, according to their intensity and the spatial distribution of the silvicultural interventions, influence and stimulate the growth of the remaining or residual trees (Thorpe et al. 2007; Vincent et al. 2009; Peri et al. 2013; Venturoli et al. 2015; Pamerleau-Couture et al. 2015). In this way, improved radial growth of residual trees generally leads to higher financial returns at the stand level despite the reduced tree density (Liu et al. 2007).
Dendroecology (Fritts and Swetnam 1989) is a useful tool to evaluate different processes of forest dynamics, e.g. regeneration, establishment, maturity or senescence of trees. In addition, it is used to date and quantify the effect of past disturbances in the tree canopy which can be translated into periods of suppression or releases of tree rings (Schweingruber 1996). Such information is required to build growth and yield models useful when predicting forest responses after the application of silvicultural treatments based on its past responses. In this way, radial growth releases due to thinning interventions and detected by dendrochronological methods were reported for several species in different parts of the world (Misson et al. 2003; Pérez-de-Lis et al. 2011; Ruano et al. 2013). In particular for Tierra del Fuego (Argentina), Franco (2015) analyzed the response to different combination of thinnings from below in an experimental Nothofagus betuloides secondary forest at Estancia Moat, located close to Beagle channel. She found that different thinning intensities produced differential effects on radial growth, where heavy thinning treatments resulted in higher individual radial growth rates than the control treatments.
Nothofagus pumilio (Poepp. et Endl.) Krasser (locally known as ‘lenga’) is the most important native tree forest species in economic terms of the sub-Antarctic forests of Argentina and Chile. On the Isla Grande de Tierra del Fuego (Argentina), the silvicultural systems have varied in the last decades, from high grading (Mutarelli and Orfila 1973), shelterwood cuts (Martínez-Pastur et al. 2000), and recently, variable retention harvesting (Martínez-Pastur et al. 2009). However, there are few examples of intermediate treatments that improve and lead the second growth forests (Martínez-Pastur et al. 2001; Peri et al. 2002, 2013), because forests are generally abandoned after the first cuts. Recently, the Dirección General de Bosques de Tierra del Fuego, within the framework of National Law No. 26,331, has promoted the large-scale implementation of these intermediate treatments in secondary forests of Tierra del Fuego (e.g. Martínez-Pastur et al. 2013). In the Argentinean forests of Tierra del Fuego, N. pumilio has been used in dendroclimatic studies because of its radial growth sensitivity to annual climate variability (Boninsegna et al. 1989; Gutiérrez 1992; Mundo et al. 2007; Massaccesi et al. 2008). However, dendrochronological methods have not been used until now for silvicultural management.
Intermediate treatments are performed to ensure the composition, stem quality, spacing and desirable characteristics within a stand (Daniel et al. 1982; Smith et al. 1998). The positive effects of thinning are to increase net productivity and the stand quality as well as to favor the growth of the remaining trees, i.e. those with the best characteristics for the management objective. However, so far, the effects of thinning, and even pruning, on radial growth to sustain their inclusion in silvicultural systems in the N. pumilio forests of Tierra del Fuego have not been quantified through tree-ring methods yet. In consequence, the objective of this study was to analyze the effect of thinning and pruning on N. pumilio radial growth through dendrochronological techniques in a long-term experimental stand.
2 Materials and Methods
2.1 Study Site
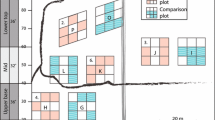
This study was based on samples from plots located in the Aguas Blancas long-term experimental forest (Fig. 14.1, 54°36′35” S; 67°15′43” W; 215 m a.s.l.) in the department of Ushuaia (Tierra del Fuego province, Argentina), 14 km from the town of Tolhuin. These plots are part of the PEBANPA network (Peri et al. 2016). The forest type corresponds to N. pumilio pure-type based on the classification of Uriarte and Grosse (1991) and to site classes II and III according to the classification proposed by Martínez-Pastur et al. (1997), where dominant heights fluctuate between 22 and 26 m.

(a) Map showing the location of study site in Tierra del Fuego, Argentina. On the right, photos (taken in 2010) of the different treatments analyzed in this study: (b) one high-intense thinning (1 T, only thinned in 1984–1985), (c) two thinnings (2 T, thinned in 1984–1985 and 1999) and (d) two thinnings with pruning (2 T + P, thinned in 1984–1985 and 1999 plus pruning)
Between 1965 and 1967, the National Forestry Administration and the University of Buenos Aires installed a 16-hectare trial in the Aguas Blancas experimental forest where single-tree selective cutting was carried out previously. At that moment, strip clearcuts were performed on alternate strips of 30–40 m wide and 400 m long. Natural regeneration was installed homogeneously and abundantly in all the strips. In the 1984–1985 season, the former National Forest Institute (IFONA) performed different pre-commercial low thinning systems on the regeneration (saplings of approximately 2 m height, 2.5 cm at DBH and 80,000–100,000 ind.ha−1). In 1999, the Centro Austral de Investigaciones Científicas (CONICET) and the Provincial Forest Administration (Dirección de Bosques) of Tierra del Fuego evaluated the first trial on a 2350 m2 area where 5% of the original basal area (BA) had been left. Subsequently, new treatments were installed on this thinned forest and unmanaged sector within the same strip. A second low-intense thinning was performed leaving 68–75% of the BA left by the thinning carried out in 1984–1985. In addition, the same low-intensity thinning with pruning up to 4–5 m in height (2/3 of the living crown) was applied on another sector of the same strip. In consequence, in this study, the following treatments were analyzed: (i) one high-intense thinning (1 T, only thinned in 1984–1985), (ii) two thinnings (2 T, thinned in 1984–1985 and 1999), and (iii) two thinnings with pruning (2 T + P, thinned in 1984–1985 and 1999 plus pruning). In each of these treatments, ring width patterns of 20 individuals were analyzed by means of transverse sections taken at breast height (1.30 m) during an intervention in 2011 (not analyzed in this study). The mean DBH (SE) of the 20 sampled in each treatment was 9.7 cm (0.5) for 1 T, 10.8 cm (0.3) for 2 T and 11.6 cm (0.5) for 2 T + P.
2.2 Dendrochronological Methods
The processing of cross-sections followed standard dendrochronological procedures (Stokes and Smiley 1968). Samples were air-dried and sanded to create polished surfaces to facilitate the identification of the annual rings under a stereomicroscope (10x–50x). Two perpendicular radii were analyzed on each sample and the growth rings were identified and visually cross-dated. According to Schulman’s (1956) convention for the Southern Hemisphere, calendar years of annual rings were assigned to the year in which ring formation begun. The ring widths (RW) were measured using a Velmex UniSlide tablet connected to a Metronics Quick-Check QC-1000 digital counter with an accuracy of 0.01 mm. COFECHA software (Holmes 1983) was used to statistically verify the dating quality of the series and to calculate the correlation between series and mean sensitivity. Basal area increment (BAI) was calculated from RW series assuming circular cross sections after averaging the two radii for each tree. BAI is usually preferred over the RW series since this variable provides a more precise quantification of wood production due to the ever increasing diameter of a living tree (Visser 1995; Pedersen 1998). Then, individual records of both variables were combined to obtain a mean chronology per treatment. In addition, the methodology developed by Nowacki and Abrams (1997) was used to detect growth releases. This method calculates the percentage of growth change (PGC) year by year over the series, in this case individual and average per treatment, using the following equation: [(M2-M1)/M1] x 100 where M1 is the average ring width of the previous 5 years including the calculated year, and M2 is the average of the following 5 years following Fang et al. (2018). A PGC that exceeded 50% and was sustained for at least 3 consecutive years was considered a release.
To compare growth before and after interventions, we calculated 5-year means (periodic annual increments, PAI5) and statistically analyzed the differences between them for the 2 years of interventions. The mean differences in PAI of pre- and post-intervention periods (ΔPAI5) for each treatment were calculated in the context of pretest-posttest analyses (Dimitrov and Rumrill Jr 2003) and then compared with one-way ANOVAs and HSD Tukey post-hoc tests. Before performing this procedure, the series were checked for normality using the Shapiro-Wilk test. Residuals normality and homoscedasticity were also checked through Kolmogorov-Smirnov and Bartlett tests, respectively. Since the ΔPAI in BAI for the 1984 did not fulfilled this assumption, the non-parametric Kruskal-Wallis test was used to detect differences between means.
To estimate the effect of the second intervention in relation to what was expected with an unaffected control situation, the radial growth pattern of 1 T treatment was used as a reference pattern. Thus, the percentage differences of 2 T and 2 T + P treatments in relation to 1 T were calculated. In addition, paired student’s t tests were performed between 1 T, 2 T and 2 T + P mean annual increments.
All statistical analyses were conducted on R version 3.5.3(R Development Core Team 2019).
3 Results
3.1 Radial Growth Patterns
To analyze the effect of thinning and pruning on radial growth in the experimental long-term stand at Aguas Blancas, ring widths were measured for each radii to create individual RW and BAI series and then combined into treatment mean series (Table 14.1 and Fig. 14.2). Firstly, all mean series had almost the same starting year, which corroborated the existence of a post-harvesting cohort in the three treatments.

Average ring width (RW, left panels) and basal area increment (BAI, right panels) for one thinning (1 T; a and b), two thinnings (2 T; c and d) and two thinnings with pruning (2 T + P; e and f) treatments. The central axis indicates the number of trees (n). The vertical dotted lines indicate the years of the two interventions (1984 and 1999). The grey shaded areas at the bottom of each panel represent the sample size (average series per tree) per year
The first thinning in 1984, which was very intense since it reduced the occupation level to 6% of the initial density, generated an increase in the average annual increment for the three treatments. This was reflected by large peaks in the radial growth patterns associated to this event, both in ring width and basal area increment in all three treatments. The second intervention of 1999, which left 56% of the density of individuals in 2 T and 2 T + P treatments, could only be seen as a peak, smaller than that of 1984, in the RW and BAI series of 2 T + P treatment (Fig. 14.2e, f), but with almost no changes in 2 T (Fig. 14.2c, d).
In terms of common variability between individual series per treatment, the average inter-correlation per individual series before the first intervention was relatively low (r = 0.274 ± 0.031) and it increased to 0.719 ± 0.023 between-interventions period (1985–1999). Then, after the second intervention, the 2 T and 2 T + P treatments showed a greater increase, reaching r values of 0.733 and 0.749 respectively. The 1 T treatment, which had no intervention in 1999, showed a slight decrease in the mean inter-correlation from 0.764 to 0.722. On the other hand, the mean sensitivity, as a measure of the degree of inter-annual variation, also showed an increase associated with the application of the intermediate treatment.
To evaluate the annual variation in each treatment and to quantify growth releases, the annual percentages of growth change (PGC) in RW and BAI series were calculated (Fig. 14.3). For the 1984 intervention, the average PGC between treatments was 129% (SE = 20%), and it was recorded as individual releases on 57% of the trees in each treatment. However, the PGC for BAI increased to 419% (SE = 39%) at same year. Neither RW nor BAI showed a PGC greater than 50% nor individual release associated with the interventions carried out in 1999 were detected. Only a light growth release was found in 1996 for the three treatments.

Percentages of growth change (PGC) in mean ring width (RW, left panels: a, c and e) and basal area increment (BAI, right panels: b, d and f) (black lines) with their respective standard errors (grey lines) for the three treatments: 1 T, one thinning (a, b); 2 T, two thinnings (c, d) and 2 T + P, two thinnings with pruning (e, f). Cut lines indicate 50% PGC threshold for release detection. The bars show the individual trees released (PGC >50%) in each treatment. The vertical dotted lines indicate the two interventions (years 1984 and 1999)
3.2 Differences in Periodic Growth Due to the Interventions
Growth changes associated with the two interventions were analyzed by calculating 5-year periodic annual differences (ΔPAI5) and by comparing them between treatments (Fig. 14.4). In the case of the 1984 strong thinning, significant differences were found in RW (F = 7.994, p < 0.001), but not for BAI (H = 5.318, p = 0.07). The paired comparison of means indicated that 1 T treatment was not different from 2 T + P and 2 T, respectively, with the highest and lowest growth which were significantly different from each other. In opposition, the 1999 intervention generated significant differences in both RW (F = 14.008, p < 0.001) and BAI (F = 16.729, p < 0.001). In this last intervention, 1 T presented negative differences and differed from 2 T and 2 T + P in both RW and BAI. The treatments 2 T and 2 T + P had positive values in ΔPAI5, but differed significantly from each other with a greater increase in 2 T + P treatment.

Differences in periodic annual increment (ΔPAI) for 5 years before and after the 1984 (left panels: a and c) and 1999 (right panels: b and d) interventions in ring width (RW, upper panels: a and b) and basal area increment (BAI, lower panels: c and d) for each treatment (1 T, one thinning; 2 T, two thinnings and 2 T + P, two thinnings with pruning). Bars represent mean difference with standard error bars. Means with the same letter are not significantly different at p < 0.05 based on ANOVA and HSD Tukey post-hoc tests. Kruskal-Wallis test was used to detect differences between medians in panel c (H = 5.318, p = 0.07)
In order to evaluate the effect of the second intervention compared to what was expected without any type of treatment, the percentage differences in the annual growth of the 2 T and 2 T + P treatments in relation to the 1 T treatment were analyzed (Fig. 14.5). No percentage changes lower than −25% or greater than 15% were found for 2 T and 2 T + P during the years prior to the 1999 intervention (Fig. 14.5a, b). After the intervention, positive differences were found with peaks in 2001 with greatest percentage difference of 96% and 121% in RW and BAI, respectively. The difference corresponding to 2 T + P was found to be greater than 2 T in all situations. Paired t tests evidenced that these differences were of lesser magnitude and limited to 2 years for 2 T treatment (Fig. 14.5c, d) while in 2 T + P the increase in growth lasted until 10 years after the last intervention (Fig. 14.5e, f).

Annual average differences of the two-thinning (2 T, blue lines and blue bars) and two-thinning plus pruning (2 T + P, red lines and red bars) treatments with respect to the one thinning treatment (1 T) for ring width (RW, left panels) and basal area increment (BAI, right panels) analyzed for 10 years before and after the second intervention. Differences are expressed as a percentage of the 1 T treatment (a, b) or in the same units (c–f). The vertical dotted lines indicate the date of the second intervention (1999). On the lower plots (c, d, e and f), segments capped with an asterisk correspond to intervals with index differences statistically different (p < 0.05) from the long-term mean based on paired t tests
4 Discussion
This long-term case study showed that the dendrochronological techniques widely and mainly used to detect growth releases due to disturbances can be applied to quantify and to reconstruct the effect of intermediate treatments in N. pumilio forests. In addition, as it was expected, higher growth rates were found as a consequence of highly intensive thinning interventions. On the other hand and using dendrochoronological methods for the first time in this species under silvicultural management, a significant increase in radial growth was detected as a response to the combined effect of thinning and pruning. Moreover, BAI appears to be a more sensible parameter than RW to detect the effect of intermediate treatments.
The effect of the intermediate treatments performed in 1984 and 1999 were verified both visually through peaks in their growth patterns and increases in the PGC for the three treatments. In the three treatments analyzed, in terms of trends, considerable increases in RW and BAI were observed as a consequence of the heavy thinning that these plots had in 1984 (Fig. 14.2) These radial growth releases are due to the fact that thinning, as an intermediate type of silvicultural treatments, produces the release of resources (Smith et al. 1998) and, consequently, has a positive effect on the growth of the remaining trees. In relation to the 1999 intervention, the plot that received thinning and pruning (2 T + P) showed a higher growth, in relation to the 1 T treatment, 2 years after the intervention (2001). The treatment that only received thinning on 1999 (2 T) also showed an increase in the percentage difference with respect to treatment without intervention. However, this increase did not reach the magnitude detected in 2 T + P. This could be due to the fact that the elimination of basal branches coincided with those less efficient in terms of photosynthetic balance and, consequently, leaving remnant branches with greater photosynthetic capacity that use more efficiently the higher amount of light irradiance resulting from the simultaneous thinning of individuals.
In relation to the second intervention that took place in 1999, in the 2 T treatment, which only had a low intense thinning, no changes were observed in RW growth, though there was a slight increase in BAI. On the contrary, treatment with thinning and pruning (2 T + P) showed an increase in both variables. Therefore, the repetition of thinnings 15 years apart did not translate into significant changes in radial growth. However, the addition of pruning produced the increase of radial growth, both measured in RW and BAI, for 10 years after the intervention in comparison to the control situation. This fact would be explained by a greater accumulation of biomass in the stem, possibly due to a greater efficiency in the capture of resources by the remaining individuals who suffered a rejuvenation of the crown. Other authors also detected an increase in radial growth due to the application of pruning in other species from the northern hemisphere (Stein 1955; Staebler 1963). Recently, Giantomasi et al. (2015) also observed that Prosopis flexuosa trees increased radial growth after pruning in the Monte region of western-central Argentina.
The growth rate recorded before the first intervention in the three treatments is similar to that observed by Barrera et al. (2000) in self-thinning young forests located in Valle de Andorra, Tierra del Fuego. The percentage of growth change quantified in this study (with maximum values of 155% in RW and 449% in BAI) was parallel to those reported by Franco (2015) (360% in RW and 431% in BAI) for thinned N. betuloides trees in Tierra del Fuego, and even surpassed the value (80% in BAI) reported by Pérez-de-Lis et al. (2011) for thinned Pinus canariensis trees. However, these values are similar to those reported in other thinning studies on N. pumilio conducted in Southern Patagonia (Peri et al. 2002, 2013).
On another subject, after the first intervention, the mean inter-correlation values, as an expression of the degree of agreement and common signal between ring width series, in comparison to those values reported by Mundo et al. (2007) and Massaccesi et al. (2008) were very high. This would indicate that the application of thinning and pruning treatments had a very homogeneous effect on the remaining trees, they were not damaged and they responded very homogeneously. In addition, the increase in average sensitivity after the first intervention in the three treatments would indicate that these trees had a quite constant radial growth before the opening of the canopy and that once competition had been reduced due to the thinning ; the remaining individuals began to show radial patterns with greater inter-annual variability more linked to climatic annual variability than to the competition for resources.
Finally, this research could be used as reference for future studies to quantify the intensity of past interventions using these same dendrochronological methods presented in this chapter in complementation with reports of exploitation or measurements of forest structure. In this way, by calibrating different thinning intensities with releases in RW and BAI, it would be possible to estimate past thinning and pruning intensities on stands where this type of information is unknown. In addition, it would be very useful to carry out anatomical studies on the samples analyzed in this study or in other sites with known intermediate treatments to better understand the response of N. pumilio in terms of xylem transportation and ecophysiology linked to silvicultural management.
References
Barg AK, Edmonds RL (1999) Influence of partial cutting on site microclimate, soil nitrogen dynamics, and microbial biomass in Douglas-fir stands in western Washington. Can J For Res 29:705–713. https://doi.org/10.1139/x99-045
Barrera MD, Frangi JL, Richter LL et al (2000) Structural and functional changes in Nothofagus pumilio forests along an altitudinal gradient in Tierra del Fuego, Argentina. J Veg Sci 11:179–188. https://doi.org/10.2307/3236797
Boninsegna JA, Keegan J, Jacoby GC et al (1989) Dendrochronological studies in Tierra del Fuego, Argentina. Quat S Am A 7:305–326
Caldentey J, Schmidt H, Ibarra M et al (1998) Modificaciones microclimáticas causadas por el uso silvícola de bosques de lenga (Nothofagus pumilio), en Magallanes, Chile. In: Actas Primer Congreso Latinoamericano de IUFRO, IUFRO-CONAF, Valdivia, p 12
Daniel TW, Helms JA, Baker FS (1982) Principios de silvicultura. McGraw-Hill, México DF
Dimitrov DM, Rumrill PD Jr (2003) Pretest-posttest designs and measurement of change. Work 20:159–165
Fang O, Alfaro RI, Zhang QB (2018) Tree rings reveal a major episode of forest mortality in the late 18th century on the Tibetan Plateau. Glob Planet Change 163:44–50. https://doi.org/10.1016/j.gloplacha.2018.02.004
Fayle DCF (1983) Differences between stem and root thickening at their junction in red pine. Plant Soil 71:161–166. https://doi.org/10.1007/BF02182651
Franco MG (2015) Respuesta del crecimiento radial de Nothofagus betuloides “guindo” a diferentes intensidades de raleo en Tierra del Fuego, Argentina. Undergraduate thesis, Universidad Nacional de La Plata, Argentina
Fritts HC, Swetnam TW (1989) Dendroecology: a tool for evaluating variations in past and present forest environments. Adv Ecol Res 19:111–188. https://doi.org/10.1016/S0065-2504(08)60158-0
Giantomasi MA, Alvarez JA, Villagra PE et al (2015) Pruning effects on ring width and wood hydrosystem of Prosopis flexuosa DC from arid woodlands. Dendrochronologia 35:71–79. https://doi.org/10.1016/j.dendro.2015.07.002
Gutiérrez E (1992) Dendrochronological analysis of Nothofagus pumilio along an altitudinal gradient in a Nothofagus forest in Tierra del Fuego. Argentina. Lundqua Rep 34:137–141
Holmes RL (1983) Computer-assisted quality control in tree-ring dating and measurement. Tree-Ring Bull 43:69–78
Liu C, Ruel JC, Zhang SY (2007) Immediate impacts of partial cutting strategies on stand characteristics and value. For Ecol Manag 250:148–155. https://doi.org/10.1016/j.foreco.2007.05.010
Martínez-Pastur G, Peri PL, Vukasovic RF et al (1997) Site index equation for Nothofagus pumilio Patagonian forest. Phyton 61:55–60
Martínez-Pastur G, Cellini JM, Peri PL et al (2000) Timber production of Nothofagus pumilio forests by a shelterwood system in Tierra del Fuego (Argentina). For Ecol Manag 134:153–162. https://doi.org/10.1016/S0378-1127(99)00253-4
Martínez-Pastur G, Cellini JM, Lencinas MV et al (2001) Modificación del crecimiento y de la calidad de fustes en un raleo fuerte de un rodal en fase de crecimiento óptimo inicial de Nothofagus pumilio (Poepp. et Endl.). Ecol Austral 11:95–104
Martínez-Pastur G, Lencinas MV, Cellini JM et al (2009) Timber management with variable retention in Nothofagus pumilio forests of Southern Patagonia. For Ecol Manag 258:436–443. https://doi.org/10.1016/j.foreco.2009.01.048
Martínez-Pastur GJ, Cellini JM, Lencinas MV et al (2011) Environmental variables influencing regeneration of Nothofagus pumilio in a system with combined aggregated and dispersed retention. For Ecol Manag 261:178–186. https://doi.org/10.1016/j.foreco.2010.10.002
Martínez-Pastur G, Favoretti S, Paredes D et al (2013) Propuestas de cosecha para bosques secundarios y floreados de Nothofagus pumilio en Tierra del Fuego. In: Actas de las II Jornadas Forestales de Patagonia Sur y II Congreso Internacional Agro-forestales Patagónico. Calafate, p 14
Massaccesi G, Roig FA, Martínez-Pastur GJ et al (2008) Growth patterns of Nothofagus pumilio trees along altitudinal gradients in Tierra del Fuego, Argentina. Trees-Struct Funct 22:245–255
Misson L, Vincke C, Devillez F (2003) Frequency responses of radial growth series after different thinning intensities in Norway spruce (Picea abies (L.) Karst.) stands. For Ecol Manag 177:51–63. https://doi.org/10.1016/S0378-1127(02)00324-9
Mundo IA, Barrera MD, Roig FA (2007) Testing the utility of Nothofagus pumilio for dating a snow avalanche in Tierra del Fuego, Argentina. Dendrochronologia 25:19–28. https://doi.org/10.1016/j.dendro.2007.01.001
Mutarelli E, Orfila E (1973) Algunos resultados de las investigaciones de manejo silvicultural que se realizan en los bosques Andino-Patagónicos de Argentina. Rev For Argent 17:69–75
Nowacki GJ, Abrams MD (1997) Radial-growth averaging criteria for reconstructing disturbance histories from presettlement-origin oaks. Ecol Monogr 67:225–249. https://doi.org/10.1890/0012-9615(1997)067[0225:RGACFR]2.0.CO;2
Otero L, Contreras A, Barrales L (1998) Cortas de protección en claros de bosque: efectos microclimáticos. In: Actas Primer Congreso Latinoamericano de IUFRO. IUFRO-CONAF, Valdivia, p 8
Pamerleau-Couture É, Krause C, Pothier D et al (2015) Effect of three partial cutting practices on stand structure and growth of residual black spruce trees in North-Eastern Quebec. Forestry 88:471–483. https://doi.org/10.1093/forestry/cpv017
Pedersen BS (1998) The role of stress in the mortality of midwestern oaks as indicated by growth prior to death. Ecology 79:79–93. https://doi.org/10.1890/0012-9658(1998)079[0079:TROSIT]2.0.CO;2
Pérez-de-Lis G, García-González I, Rozas V et al (2011) Effects of thinning intensity on radial growth patterns and temperature sensitivity in Pinus canariensis afforestations on Tenerife Island, Spain. Ann For Sci 68:1093–1104. https://doi.org/10.1007/s13595-011-0125-8
Peri PL, Martínez-Pastur G, Vukasovic R et al (2002) Thinning schedules to reduce risk of windthrow in Nothofagus pumilio forests of Patagonia, Argentina. Bosque 23:19–28
Peri PL, Martínez-Pastur G, Monelos L (2013) Natural dynamics and thinning response of young lenga (Nothofagus pumilio) trees in secondary forests of Southern Patagonia. Bosque 34:273–279
Peri PL, Lencinas MV, Bousson J et al (2016) Biodiversity and ecological long-term plots in Southern Patagonia to support sustainable land management: the case of PEBANPA network. J Nat Conserv 34:51–64. https://doi.org/10.1016/j.jnc.2016.09.003
R Development Core Team (2019) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. https://www.R-project.org/
Ruano I, Rodríguez-García E, Bravo F (2013) Effects of pre-commercial thinning on growth and reproduction in post-fire regeneration of Pinus halepensis mill. Ann For Sci 70:357–366. https://doi.org/10.1007/s13595-013-0271-2
Schulman E (1956) Dendroclimatic changes in semiarid America. University of Arizona Press, Tucson
Schweingruber FH (1996) Tree rings and environment: dendroecology. Haupt, Vienna
Smith DM, Larson BC, Kelty MJ et al (1998) The practice of Silviculture: applied forest ecology, 9th edn. Wiley, New York
Staebler GR (1963) Growth along the stems of full crowned Douglas-fir trees after pruning to specified heights. J For 61:124–127
Stein WI (1955) Pruning to different heights in young Douglas-fir. J For 53:352–355
Stokes M, Smiley T (1968) An introduction to tree-ring dating. University of Chicago Press, Chicago
Thibodeau L, Raymond P, Camiré C et al (2000) Impact of precommercial thinning in balsam fir stands on soil nitrogen dynamics, microbial biomass, decomposition, and foliar nutrition. Can J For Res 30:229–238. https://doi.org/10.1139/x99-202
Thorpe HC, Thomas SC, Caspersen JP (2007) Residual-tree growth responses to partial stand harvest in the black spruce (Picea mariana) boreal forest. Can J For Res 37:1563–1571. https://doi.org/10.1139/X07-148
Uriarte A, Grosse H (1991) Los bosques de lenga. Una orientación para su uso y manejo. Recopilación bibliográfica. CORFO-INFOR, Concepción
Venturoli F, Franco AC, Fagg CW (2015) Tree diameter growth following silvicultural treatments in a semi-deciduous secondary forest in Central Brazil. Cerne 21:117–123. https://doi.org/10.1590/01047760201521011204
Vincent M, Krause C, Zhang SY (2009) Radial growth response of black spruce roots and stems to commercial thinning in the boreal forest. Forestry 82:557–571. https://doi.org/10.1093/forestry/cpp025
Visser H (1995) Note on the relation between ring widths and basal area increments. For Sci 41:297–304. https://doi.org/10.1093/forestscience/41.2.297
Acknowledgments
This chapter was partly funded by SECYT-Universidad Nacional de La Plata (award code A282). The authors are very grateful to the editors of this book for their invitation to write a chapter. We also thank two anonymous reviewers for helping to improve the final version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Copyright information
© 2020 Springer Nature Switzerland AG
About this chapter

Cite this chapter
Mundo, I.A., Palazzini, D.A., Barotto, A.J., Martínez-Pastur, G.J., Barrera, M.D. (2020). Dendroecology Applied to Silvicultural Management in the Southern Patagonian Forests: A Case Study from an Experimental Forest in Tierra del Fuego, Argentina. In: Pompa-García, M., Camarero, J. (eds) Latin American Dendroecology. Springer, Cham. https://doi.org/10.1007/978-3-030-36930-9_14
Download citation
DOI: https://doi.org/10.1007/978-3-030-36930-9_14
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-030-36929-3
Online ISBN: 978-3-030-36930-9
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)