
Abstract
This work describes potential opportunities for utilization of agro-industrial residues to produce green biodegradable plastics of poly(3-hydroxybutyrate) (PHB). Wheat straws were examined with good efficacy of carbon substrates using Cupriavidus necator. Production was examined in separate hydrolysis and fermentation (SHF) in the presence and absence of WS hydrolysis enzymes, and in simultaneous saccharification and fermentation (SSF) with enzymes. Results showed that production of PHB in SSF was more efficient in terms of viable cell count, cell dry weight, and PHB production and yield compared to those of SHF and glucose-control cultures. While glucose control experiment produced 4.6 g/L PHB; SSF produced 10.0 g/L compared to 7.1 g/L in SHF when utilizing enzymes during WS hydrolysis. Results showed that most of sugars produced during the hydrolysis were consumed in SHF (~98 %) compared to 89.2 % in SSF. Results also demonstrated that a combination of glucose and xylose can compensate for the excess carbon required for enhancing PHB production by C. necator. However, higher concentration of sugars at the beginning of fermentation in SHF can lead to cell inhibition and consequently catabolite repressions. Accordingly, results demonstrated that the gradual release of sugars in SSF enhanced PHB production. Moreover, the presence of sugars other than glucose and xylose can eliminate PHB degradation in medium of low carbon substrate concentrations in SSF.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Most plastic materials currently used on daily basis are non-biodegradable polymers derived from fossil fuels. In view of the environmental impact that could arise from non-biodegradable plastics in future, various types of biodegradable plastics are being developed. Plastics are made with various polymers that fall under the following groups: polynucleotides, polyamides, polysaccharides, polyoxoesters, polythioesters, polyphosphates, polyisoprenoides, and polyphenols [1]. Poly(-3-hydroxybutyrate) (PHB) is one of the promising biodegradable natural polymers that fall under the polyoxoesters group. It has wide economic interest considering its material properties that are closely related to those of propylene [2–4]. PHB can be synthesized by polymerizations using monomers under mild conditions with minimal impact to the environment [3, 4]. Furthermore, PHB is one of the biopolymers, which has been known to undergo biodegradation under aerobic and anaerobic environments without forming any toxic products [5, 6]. When hydrolyzed, PHB produces (R)-3-hydroxybutyric acid monomers, which has lots of industrial and medical applications [7, 8]. PHB is a biopolymer that can be produced through fermentation processes by different bacterial strains. Most of these bacteria have been reported to accumulate PHB within their cells as carbon and energy storage materials under certain nutrient requirements in their culture broth. Among some of these PHB producers, Cupriavidus necator (C. necator) and Azohydromonas lata (A. lata) are the most widely studied strains [4, 9, 10]. PHB has potential applications in regenerative medicine considering its biodegradability (e.g., bone regeneration, dressing of wounds, sutures, etc.) [11]. The use of PHB as packaging materials has also been reported [12]. There is also some evidence that PHB could serve as micro-particulate carrier of drugs [13]. Although a lot of work has been extensively carried out on PHB production, one of the major challenges in the commercialization is the cost of production, which is still far higher than those of the petroleum-based plastics. It is evidently clear that the cost of producing PHB has to be considerably reduced before the polymer can completely replace plastics derived from petroleum products.
PHB is a promising green biomaterial that can be synthesized from renewable low value feed-stocks by fermentative processes. Another interesting aspect of PHB is that most bacteria that produce it have wide substrate specificities. The growing interest in the use of renewable and sustainable agricultural residues as carbon substrates to produce green products is quite obvious. As a way of reducing the cost of production, agricultural residues such as molasses, potato waste, glycerol, and soy cake have been investigated [14–17]. However, most starchy renewable resources would serve better as foods or feeds rather than as feedstock for production of green chemicals. This justifies the need to shift from starchy foods to non-edible resources (such as cellulose) which are more abundant and cheaper. Among agricultural residues, wheat straw (WS) is one of the abundant biomass that can offer a lot of advantages in the production of PHB, a green biomaterial. WS is one of the cellulosic biomass which is generated at wheat processing industries. Statistics presented by Food and Agriculture Organization shows that Canada alone produced 29 million metric tons of wheat in 2008, implying that Canada is one of the largest producers of WS in the world [17]. Furthermore, it was reported that about 21 % of the world’s food depends on the wheat crop [18]. All these evidence have indicated the abundance of WS for possible exploitation in production of fine chemicals and high-value products. Although WS has some applications in farming industries such as in making fertilizers or as animal feeds, the increasingly accumulation of WS requires that other alternative ways of utilizing it should be implemented.
While there has been lots of interest recently in utilizing agricultural residues such as WS in green biofuel production [19, 20], there has been no studies where they were utilized to produce PHB. Literature search showed that most PHB-producing bacteria do not metabolize WS. However, WS can be hydrolyzed to various sugars such as glucose, xylose, galactose, arabinose, and mannose [21, 22]. Bioconversion of WS has been favored because of its relatively low lignin content (<20 %, w/w) and high carbohydrate content [hemicellulose (50 %) and cellulose (30 %)] [23]. Interestingly, efficient utilization of WS would not only help in the production of fine chemicals and high-value products but also would ensure the disposal of huge accumulation of WS at the wheat processing factories. To utilize WS in fermentation industries, pretreatment following well-defined methods should be implemented initially [22–24]. The objective of the pretreatment is to break down the shield formed by lignin and hemicellulose, disrupt the crystalline structure and reduce the degree of polymerization of cellulose. WS hydrolysis can be influenced by porosity (accessible surface area) of lignocellulosic biomass, cellulose fiber crystallinity, and lignin and hemicellulose content [25]. A pretreatment process is, therefore, essential to remove lignin and hemicellulose, reduce cellulose crystallinity, and increase the porosity of the materials [26]. Pretreatment of cellulosic materials are generally done using acid [24, 27], or alkali [28, 29]. It was reported that the use of dilute sulfuric acid is advantageous in the hydrolysis of cellulosic biomass as it hydrolyzes much of the hemicelluloses to pentose sugars [19]. Even by pretreatment with dilute sulfuric acid and heating, concentrations of sugars achievable from the hydrolysis are still low. To generate more sugars from WS, enzymatic hydrolysis is usually utilized that leads to a higher cost of hydrolysis.
Although most fermentative production of PHB has been extensively done using starchy and soluble sugars, very little information is available regarding the use of cellulosic materials such as carbon substrates [30]. Previous study by Page and Knosp [31] reported that beet molasses and malt extract promoted higher PHB production of 2.74 and 2.80 g/L, respectively. Moreover, Hori et al. [32] demonstrated the ability of Bacillus megaterium to accumulate PHB with cell content up to 32 % of the cell dry weight. Later, Chaijamrus and Udpuay [30] achieved highest PHB production of 43 % cell content after 45 h of growth when 4 % molasses and 4 % CSL were used. In the present study, cost-competitive and environmentally friendly approach to produce green biodegradable bioplastics using bacteria strain fed on sugars produced from agro-industrial waste was presented. PHB was produced in simultaneous saccharification and fermentation (SSF) and separate hydrolysis and fermentation (SHF) using C. necator solely from WS as a renewal resource of agricultural residue. WS were chosen for its low cost and high availability especially within central Europe and North America. A comparison is established between hydrolysis with and without enzymes and their effect on the PHB production in SHF. C. necator generally assimilates glucose very well and relatively high yields of PHB have been reported in previous studies with glucose substrate [33–36]. Therefore, glucose culture was used as the control culture to understand glucose role in the production of PHB and if sugar mixtures can be utilized to enhance its production.
Materials
Cupriavidus necator (ATCC 17699) was purchased from American Culture Collection Center, (Manassas, VA 20108, USA). Wheat Straw was collected from a local farm in Barrie, Ontario, Canada. The strain was grown in basal medium containing the following components (per L): 5 g yeast extract, 10 g polypeptone, 200 mg MgSO4·7H2O, 100 mg NaCl, 20 mg CaCl2·2H2O, 10 mg FeSO4·H2O, 0.5 mg Na2MoO4·2H2O, 0.5 mg Na4WO4·2H2O, 0.5 mg MnSO4, 1,600 mg K2HPO4, 200 mg KH2PO4 and 1,000 mg (NH4)2SO4 and wheat straw 12 % (w/v). All chemicals were purchased from Sigma-Aldrich and were used as received.
Experimental methods
Bacterial strain and culture growth conditions
Cupriavidus necator (ATCC 17699) was activated, in accordance with ATCC guidelines, using nutrient agar. Liquid culture was prepared by transferring dry bacterial powder into nutrient broth and statically incubated (Symphony 8.5A, VWR) at 30 °C and initial pH 7 (Easy Seven, Mettler Toledo) for 3 days. Cultivation on Agar plates was done by transferring liquid culture aseptically into Petri plates, containing nutrient agar, and incubated at 30 °C and initial pH 7.0 for 7 days. Inoculum solution was prepared from the 7-day-old agar-plates. These cultures were aseptically flooded with 20 mL sterile distilled water and gently suspending the culture with a cell spreader on agar-plates. Final solution was transferred to sterile inoculum tubes and mixed thoroughly using a VWR Analogue Vortex Mixer.
Wheat straw hydrolysis
Initial weight of 120 g of WS was soaked in 1 L of 1 % (v/v) sulfuric acid for 1 h in flasks with gentle shaking. The mixture was heated at 121 °C for 30 min in an autoclave. Upon cooling, the mixture was adjusted to pH 7 using 10 M NaOH. Thereafter, the WS hydrolysates were filtered to remove the WS debris in the case of SHF with only acidic hydrolysis, leaving a clear solution to which mineral salts and other ingredients were added. Further sterilization was done at 121 °C for 20 min before inoculating the seed culture. In the case of acidic hydrolysis with enzymes in SHF, 3.75 mL each of cellulase and β-glucosidase was added per 120 g of WS suspension solution after acidic pretreatment. After the addition of enzymes, solutions were incubated at 45 °C for 72 h under mild agitation (100 rpm). The seed culture was then added to the clear WS medium filtrate after removal of debris in the case of SHF. However, seed culture was added simultaneously with the addition of enzymes to the suspension solution of WS above in the case of SSF. All PHB fermentation production experiments were conducted for 72 h, when most of the produced sugars were consumed in all fermentation experiments.
PHB production experiments
Fermentation experiments were conducted in 500 mL shaking flasks each containing 200 mL of fermentations medium. The media composition was as follows: 200 mL of WS hydrolysate solution (Carbon source), 5 g/L yeast extract (nitrogen source), 10 g/L polypeptone, 200 mg/L MgSO4·7H2O, 100 mg/L NaCl, 20 mg/L CaCl2·2H2O, 10/L mg FeSO4·H2O, 0.5/L mg Na2MoO4·2H2O, 0.5/L mg Na4WO4·2H2O, 0.5/L mg MnSO4, 1,600 mg/L K2HPO4, 200 mg/L KH2PO4 and 1,000 mg/L (NH4)2SO4.
Fermentation broths were aseptically added to the hydrolysate additive mixture and sterile distilled water was added to reach final volume. When the solutions reached room temperature, each flask was aseptically inoculated using 2 mL of the inoculum and then incubated at 30 °C for 3 (72 h) days with shaking speed of 250 rpm. At the end of the 3 days, the pH of each flask was recorded. During fermentation time, viable cell concentrations, sugar concentrations, and PHB production were analyzed. Samples were initially taken before and after inoculation and then were periodically thereafter until the end of fermentation. These samples were stored at −80 °C until analyzed. All samples were sterilized or autoclaved before disposal. Experimental results reported here are the averages of three biological replicates for all fermentation experiments.
Cell growth measurements
One milliliter of culture broth suspended in Eppendorf tube was centrifuged, washed with distilled water and then dried in the oven at 105 °C for 24 h. In the case of SSF cultures containing cells and WS debris, dry weight of cell was obtained after deducting the weight of WS from the total weight of the sample. Growth of C. necator was monitored by plating out the cultures in agar media after serial dilutions. Agar cultures were incubated for 24–48 h at 30 °C. Colonies were enumerated in terms of colony-forming unit per mL of culture medium (CFU/mL), in addition to bacterial cells dry weight concentration (g/L).
Extraction and quantification of PHB
PHB was extracted following the method described by Ugwu et al. [37]. One gram of lyophilized cells was extracted with 50 mL chloroform in a reflux condenser under heating in an oil bath (75 °C) for 8 h. Cell debris was removed by filtration through disposable membrane filters with 0.45 μm pore size. The chloroform/PHB solution was then precipitated with ethanol (volume of ethanol was 4 times that of the chloroform). PHB was recovered from the solvents (i.e., chloroform/ethanol mixture) by filtration followed by drying to a constant weight. The weight of PHB was calculated gravimetrically. Final PHB production was quantified as cell contents (wt%) and as a concentration (g/L).
HPLC analysis
Sugar concentrations were analyzed using pre-calibrated high-performance liquid chromatography (HPLC; Perkin Elmer). This instrument was equipped with an Ion Exchange column (Aminex HPX-87H, Biorad, Hercules, USA), a pump Series 200 (Perkin Elmer), Auto sampler Series 200 (Perkin Elmer) and a refractive index detector (HP1047A, Hewlett Packard). Samples of 50 μL were diluted 20-fold with deionized water and filtered (0.45 μm Gelman Acrodisc CR PTEF, Millipore). Total of 50 μL from each diluted sample was injected into the column and circulated for 15 min at a flow rate of 0.6 mL/min using filtered (0.2 μm nylon Millipore) and degassed mobile phase of 5 mM H2SO4 [23]. The column temperature was maintained at 60 °C using the column heater CH-30 controlled by an Eppendorf TC 50. Sugar concentrations were quantified from calibration curves that were constructed from standard sugar solutions of known concentrations (10–100 μg/mL).
Results and discussion
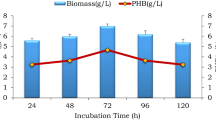
Figure 1a shows the growth profiles of C. necator in CFU/mL, while Fig. 1b shows the bacterial cell dry weight concentration for the different fermentation experiments conducted in the present study.

Time course of viable cells in SSF, SHF (with and without enzymes) in comparison with glucose substrate in the control experiment
As shown in Fig. 1a, b, the growth of C. necator concentrations increased gradually over the fermentation time, resulting in highest cell concentration reached at 72 h of fermentation time. The highest growth of C. necator was observed in the SSF compared to those of SHF and control fermentation with glucose feedstock in Figs. 1a, b. It is also clear from Fig. 1 that C. necator cell growth in the fermentation broth with glucose was higher than that of the SHF without enzymes. SHF with enzymatic hydrolysis showed higher cell growth compared to that of SHF without enzymes and glucose control culture throughout all fermentation period. The viable cell counts achieved at the end of SSF, SHF with enzymes, and glucose-controlled cultures were 63 × 107, 49 × 107, and 38 × 107 CFU/mL, respectively (results are listed in Table 1). This implies that the viable cell count in SSF was ~29 % higher than that of SHF with enzymes and ~66 % higher than that of glucose-controlled cultures. Apparently, the SSF enabled gradual release of sugars in a manner that promoted the growth of the cells. Similar trend was obtained with the cell dry weight concentrations in Fig. 1b, with final cell concentrations for SSF, SHF with enzymes, and glucose-controlled cultures of 15.3, 12.2, and 10.5 g/L, respectively (Table 1). This is equivalent to ~25 and ~46 % higher dry weight cell concentration in SSF than that in SHF with enzymes and glucose-control, respectively.
Typically, growth of C. necator in the different fermentations is dependent on the availability and consumption of sugar in the culture media. Figure 2 shows total sugar concentration profile during the fermentation period. Although the total sugars available at the beginning of the SHF were easily determined, the total sugars produced during the SSF process can only be estimated from equivalent WS hydrolysis control experiment. This is due to the fact that production and consumption of sugars through SSF occur simultaneously.

Total sugars concentration profiles during SSF and SHF (with and without enzymes)
Results in Fig. 2 demonstrate that the majority of sugars produced during the hydrolysis were consumed during the fermentation experiments. Table 1 shows that in general, the total sugars produced were higher when enzymes were utilized during the hydrolysis of WS in both SHF and SSF than in the absence of the enzymatic hydrolysis in SHF (i.e., ~56.1 to 56.3 compared to 48.9 g/L, respectively). This is, respectively, equivalent to 97.1 and 98.5 % in SHF with and without enzymes. Moreover, the total amount of glucose culture that was used in the control experiment (i.e., 30 g/L) was consumed towards the end of fermentation. SSF sugar consumption recorded a slightly lower consumption value of ~89.2 %. Concentration profiles of individual sugars in Fig. 3a, b show steep consumption until full consumption was reached after 72 h of fermentation time. Further examination of this Figure reveals that consumption of glucose and xylose were almost similar in both SHF experiments as defined by the inclination of the concentration profile curves. Concentration profiles of all other individual sugars were similar with clearly lower initial concentrations and lower consumptions compared to that of glucose and xylose.

Concentration profiles of individual sugars during a SHF with enzymes, and b SHF without enzymes
The increase in individual sugar concentrations (clear for glucose and xylose concentration profiles) in the first 24 h shown in Fig. 4 for individual sugar concentration profile in SSF culture is explained by the accumulation of produced sugars during pretreatment and due to the lower metabolism at the early stage of fermentation [19, 20]. Similarly, glucose and xylose had the highest consumption compared to other sugars in both SHF and SSF cultures. Generally, consumption rates of both sugars were comparable to those observed in the SHF in Fig. 3a, b.

Concentration profiles of individual sugars during SSF
It is generally known that C. necator gives better growth with glucose compared to other substrates. This is why most studies reported in literature with C. necator were conducted with glucose [30–32]. This explains the higher growth obtained in the present study with enzymes that produced higher glucose followed by the glucose culture experiment (see Fig. 4 and Table 1). In the case of SSF, higher cellular growth was observed with comparable sugar consumption trends and higher final residual sugars of ~2 g/L compared to that in SHF. The reason for the higher growth is obviously the gradual release of sugars to the cells during the SSF process. Results in Table 2 demonstrate that except the mannose and arabinose, higher individual sugar production was achieved when utilizing enzymes during the hydrolysis of WS in both SHF and SSF. Mannose and arabinose that are presented at comparable concentration in Table 2, exist in the hemicellulose, which can be hydrolyzed without the need to utilize enzymes [38]. Examining sugar mixture compositions in Table 2 reveals that concentration of glucose in the WS hydrolysate was the highest in both SHF (i.e., 31 and 25 g/L), followed by the xylose (18 and 14 g/L). This represents that glucose formed more than 50 % of total sugars in both SHF experiments. Moreover, both glucose and xylose formed most of the sugar mixture (i.e., total content of 87.66 and 79.75 % for SHF with and without enzymes, respectively). Results in this Table show that glucose and xylose were both fully consumed in both cases of SHF experiments. Consumption of all other individual sugars (i.e., mannose, galactose, and arabinose) was in the range of 76–95 %. As shown in Figs. 3a and 4, glucose and xylose consumption in SSF was slightly lower than that of SHF apparently due to the gradual formation (i.e., 94 and 100 %, respectively). However, consumption of galactose in both cultures was the same (i.e., 76 % for each culture). Consumption of arabinose was comparable in all fermentation experiments as well, mostly due to its low initial concentration. Based on the results above, individual sugars that were greatly consumed by C. necator were glucose and xylose. This demonstrates that a combination of the two sugars would compensate for the increasingly requirement of carbon at the exponential growth stage of the cells as shown in Fig. 1a, b. Although using enzymes in a separate hydrolysis (i.e. SHF) prior to fermentation would yield higher concentration of sugars (as seen in this case) at the beginning of the fermentation, cell inhibition and consequently, catabolite repression are usually encountered [22]. This could be the possible reason for lower growth of C. necator in SHF compared to that of SSF with the gradual sugar production. Figure 5 shows the concentration profile of glucose in the SSF and SHF (with and without enzymes) compared to that in the glucose control experiment. At comparable initial glucose concentrations, Fig. 3a shows that glucose concentration profiles were similar for the three different cultures examined.

Concentration profiles of glucose substrate in SSF and SHF (with and without enzymes) in comparison with glucose control experiments
Results in Fig. 5 demonstrate that glucose was consumed at similar rates when it was used as single sugar in the feedstock or was part of sugar mixture as was obtained from WS hydrolysis (Table 2). Full consumption of glucose demonstrates that C. necator is selective for glucose as an important substrate for PHB production. Furthermore, xylose appears in the present study to be important element as well as glucose, since their consumption was complete during the PHB production (as shown in Figs. 3, 4). This demonstrates that a combination of glucose and xylose was selectively consumed by the cells compared to other sugars (i.e. mannose, galactose, and arabinose).
During the course of the fermentation, pH of the culture media was monitored to understand if the pH could be a limiting factor for the growth of C. necator. Values for the pH of the broths at the end of the fermentations were in the range of 5.4–6.2 as listed in Table 1. This demonstrates no major changes from the initial pH of 7. According to these results, glucose-control culture had the least reduction in pH, whereas acidic pretreatment effects might have caused the slight drop in pH in SSF and SHF cultures. Nevertheless, the presence of furfuric acid in the WS hydrolysates cannot be completely ruled out given the moderate temperature used for the hydrolysis. Reduction in pH can occur due to bioconversion process during fermentations [37]. This may lead to the breakage of PHB polymer to its monomeric compounds such as (R)-3-hydroxybutyric acid [8, 35].
Results in Table 1 demonstrate that SSF gave not only the highest cell growth but also the highest production of PHB compared to SHF and glucose-controlled cultures. Final PHB concentration achieved with SSF, SHF (with enzymes), and glucose-controlled cultures at 72 h were 10, 7.08, and 4.60 g/L, respectively. This implies that the PHB concentration achieved in SSF was 41 % higher than that of SHF and 117 % higher than that of glucose-controlled culture. Accordingly, PHB cell contents were 65, 58, and 44 % for SSF, SHF and glucose-controlled cultures, respectively. Table 1 also shows the percentage yield of PHB per total sugars produced by the three fermentation processes. SSF had the highest yield of 16 %, followed by glucose-controlled cultures at 15 %, while SHF had the lowest yield of 13 %.
The PHB production obtained with SSF in this study (i.e., cell content of 65 %) is higher than those results reported in the literatures for various agro-industrial residues and C. necator. Oliviera et al. [16] reported a PHB content of 39.2 % for cultures of C. necator using soy cake enriched with molasses in solid state fermentation. Also with the same C. necator, PHB content of 38 % was recently reported for SSF process with broken rice [37]. Studies have shown that both synthesis and degradation of PHB can occur inside the cells during fermentation processes, depending on the availability of nutrients [25, 26]. According to these studies, synthesis of PHB by C. necator generally occurs under excess carbon supply, since PHB degrades in medium of low carbon substrate concentrations. However, higher concentration of sugars (as demonstrated above) at the beginning of the fermentation can lead to cell inhibition and consequently, catabolite repressions are usually encountered [22]. Therefore, it is essential to produce PHB in the presence of sugar mixtures in fermentation media with gradual release of sugars such as in SSF.
Conclusions
PHB was produced in SSF and SHF using C. necator solely from WS as a renewal resource of agricultural residue as a cost-competitive and environmentally friendly approach. Production was examined for acidically hydrolyzed WS, with and without enzymatic hydrolysis, in comparison with pure glucose as control. In general, enzymatic hydrolysis resulted in higher cell growth in both SSF and SHF, which was associated with higher total sugar production. Apparently, sugar hydrolysate from WS contains different types of sugars which would be more beneficial to the growth of bacterial strains that have wide substrate specificities. Of all the three processes, SSF was the best in terms of cellular growth in addition to PHB production and yield. PHB cell contents were 65 % obtained from SSF compared to 58 and 43 % from SHF with enzymes and glucose control, respectively. Highest yield of PHB based on total sugars consumed was obtained in SSF, which was comparable to the glucose control experiment (i.e., 15–16 %).
Results demonstrate that a combination of glucose and xylose can compensate for the excess carbon required for enhancing PHB production by C. necator. Generally, the full consumption of glucose observed demonstrates that C. necator is selective for glucose as an important substrate for PHB production. Furthermore, results also showed that xylose appears to be equivalently important substrate. Moreover, the higher sugar concentration obtained at the beginning of fermentation in SHF caused lower cellular growth compared to that of SSF. This may be related to the catabolite repressions. This demonstrates that the gradual release of sugars in SSF enhanced PHB production, while the presence of WS sugar hydrolysate mixture eliminated PHB degradation in the low sugar substrate concentrations.
References
Steinbuchel A (2001) Macromol Biosci 1:1–24
Baei MS, Najafpour GD, Younesi H, Tabandeh F, Eisazadeh H (2009) World Appl Sci J 7:157–161
Tokiwa Y, Ugwu CU (2007) J Biotechnol 132:264–272
Tokiwa Y, Calabia BP (2008) Can J Chem 86:548–555
Nishida H, Tokiwa Y (1993) J Environ Polym Degrad 1:227–233
Nishida H, Tokiwa Y (1993) J Polym Degrad 1:65–80
Gao HJ, Wu Q, Chen GQ (2002) Microbiol Lett 213:59–65
Ugwu CU, Tokiwa Y, Ichiba T (2011) Bioresour Technol 102:6766–6768
Tsuge T (2002) J Biosci Bioeng 94:579–584
Khanna S, Srivastava AK (2005) Process Biochem 40:607–619
Bernd HE, Kunze C, Freier T, Sternberg K, Kramer S, Behrend D, Prall F, Donat M, Kramp B (2009) Acta Otolaryngol 129:1010–1017
Bucci DZ, Tavares LB, Sell I (2007) Polym Test 26:908–915
Cheng C, Chung Him Y, Yin Chung C, Peter HF, Man Ken C (2006) Biomaterials 27:4804–4814
Castilho L, Mitchel D, Freire D (2009) Bioresour Technol 100:5996–6009
Rusendi D, Sheppard J (1995) Bioresour Technol 54:191–196
Oliviera F, Dias M, Castilho L, Friere D (2007) Biosour Technol 98:633–638
FAOSTAT (2011) Production-Crops, 2010 data, Food and Agriculture Organization of the United Nations
Ortiz R, Sayre K, Govaerts B, Gupta R, Subbarao G, Ban T, Hodson D, Dixon J, Ivan Ortiz-Monasterio J, Reynolds M (2008) Agric Ecosyst Environ 126:46–58
Qureshi N, Saha B, Hector R, Hughes S, Cotta M (2008) Biomass Bioenergy 32:168–175
Dahman Y, Jayasuriya KE, Kalis M (2010) Appl Biochem Biotechnol 162:1647–1659
Qureshi N, Ezeji T (2008) Biofuels Bioprod Biorefin 2:319–330
Saha B, Nichols N, Cotta M (2011) Bioresour Technol 102:10892–10897
Harper S, Lynch J (1981) J Sci Food Agric 32:1057–1062
Al-Abdallah W, Dahman Y (2013) J Chem Technol Biot 36:1735–1743
Zabihi S, Alinia R, Esmaeilzadeh F, Kalajahi J (2010) Biosyst Eng 105:288–297
Karimi K, Emtiazi G, Taherzadeh MJ (2006) Microb Technol 40:138–144
Marzialetti T, Olarte MB, Sievers C (2008) Ind Eng Chem Res 47:7131–7140
Chang VS, Nagwani M, Kim CH (2001) Appl Biochem Biotechnol 94:1–28
Silverstein R, Chen Y, Sharma-Shivappa R (2007) Bioresour Technol 98:3000–3011
Chaijamrus S, Udpuay N (2008) CIGR J 10:1–11
Page WJ, Knosp O (1989) Appl Environ Microbiol 55:1334–1339
Hori K, Kaneko M, Tanji Y, Xing XH, Unno H (2002) Appl Microbiol Biotechnol 59:211–216
Kim B, Lee S, Lee S, Chang H, Chang Y, Woo S (1994) Biotechnol Bioeng 43:892–898
Hahn S, Chang Y, Lee S (1995) Appl Environ Microbiol 61:34–39
Metabolix I, Tepha I (2003) Chem Biol 10:893–894
Lee SY, Lee Y, Wang F (1999) Biotechnol Bioeng 65:363–368
Ugwu C, Tokiwa Y, Aoyagi H (2012) J Polym Environ 20:254–257
Thirmal C, Dahman Y (2012) Can J Chem Eng 90:745–761
Acknowledgments
Authors would like to acknowledge financial support from Agriculture and Agri-Food Canada, the Natural Sciences and Engineering Research Council of Canada (NSERC), and the Faculty of Engineering and Architectural Science at Ryerson University in Toronto, Canada.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Dahman, Y., Ugwu, C.U. Production of green biodegradable plastics of poly(3-hydroxybutyrate) from renewable resources of agricultural residues. Bioprocess Biosyst Eng 37, 1561–1568 (2014). https://doi.org/10.1007/s00449-014-1128-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-014-1128-2