
Abstract
Fermentations carried out at 450-L and 20-L scale to produce Fab’ antibody fragments indicated a serious problem to control levels of dissolved oxygen in the broth due to the large oxygen demand at high cell densities. Dissolved oxygen tension (DOT) dropped to zero during the induction phase and it was hypothesised that this could limit product formation due to inadequate oxygen supply. A gas blending system at 20-L scale was employed to address this problem and a factorial 22 experimental design was executed to evaluate independently the effects and interaction of two main engineering factors: agitation rate and DOT level (both related to mixing and oxygen transfer in the broth) on Fab’ yields. By comparison to the non-gas blending system, results in the gas blending system at same scale showed an increase in the production of Fab’ by 77% independent of the DOT level when using an agitation rate of 500 rpm level and by 50% at an agitation rate of 1,000 rpm with 30% DOT. Product localisation in the cell periplasm of >90% was obtained in all fermentations. Results obtained encourage further studies at 450-L scale initially, to evaluate the potential of gas blending for the industrial production of Fab’ antibody fragments.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Oxygen availability is very critical for the aerobic growth of Escherichia coli in fermentations and, at the same time, difficult to achieve, due to the poor solubility of oxygen [16]. In industrial fed-batch fermentations, production often begins with an exponential feed phase of a limiting carbon source, until the limit of adequate oxygen transfer is reached [38].
Gas blending has been proposed as a technique to address the problems of inadequate oxygen supply. Flores et al. [14] reported that this led to increases in product yields for xanthan gums during cultivation of Xanthomas campestris where the proportion of high molecular weight polymers was higher as DOT levels were increased. Changes in agitation and shear rate are also linked to oxygen mass transfer rates and consequently, it is not always possible to identify the independent effect of oxygen levels on the fermentation, and the levels of metabolite production achieved. For example, Aurebasidium pullulans was found to be sensitive to high levels of oxygen independent of the agitation rate, and an increase of polysaccharide yield was possible by maintaining a low level of DOT at 1,000 rpm [15].
In another study, gas blending was used to separate the effects of DOT from agitation in a fungal fermentation for the production of pneumocandins. In this case, the effect of dissolved oxygen was independent of agitation rate within a power range of 2–15 kW/m3, helping to define operating boundary conditions for successful scale up to 19 m3 [35]. Also, the effect of agitation on the interaction between the extent of mixing in Xanthan fermentation broths and the rate of oxygen transfer was investigated. Findings showed that the biological performance of the culture was independent of agitation rate as long as broth homogeneity could be assured and that critical values of DOT of 6–10% were maintained in the production phase [2].
Information on the effect of DOT on recombinant protein production by E. coli is relatively scarce and no general rules can be derived. DOT has been shown to have variable effects on the production of different recombinant proteins. For example, Li et al. [28] examined four recombinant strains of E. coli for the effects of the dissolved oxygen level on the level of biomass, the plasmid content, and the levels of chloramphenicol acetyltransferase and beta-galactosidase. The optimal dissolved oxygen concentration for the specific activity of recombinant proteins was found to be dependent upon the strain. In another study by De León et al. [11], 1-L bioreactors were used to determine the effect of DOT on the production of penicillin acylase by E. coli and maximum activity was obtained at 1% DOT. Another work [6] studied over-expression of a target protein (MspI methylase) in recombinant E. coli. Results indicated that under oxygen-deficient conditions, the level of target protein decreased drastically.
This study is concerned with maximising the production titres of antibody fragments through control of DOT levels. The importance of Fab’ and other antibody fragments is based on two properties of particular interest: (a) highly specific (involving multiple non-covalent contacts between antigen and antibody based upon complementary in shape and charge) and, (b) high binding strengths (with affinities between 108/M and 1011/M) [17]. The prospect of being able to exploit these properties and generate molecules of high binding affinity against any chosen target led Paul Ehrlich to predict (in his Croonian Lecture to the Royal Society in 1900) an “age of magic bullets” providing great applications in human therapeutics. At present, there are approximately 200 recombinant antibodies that are either approved for use or in a late stage of clinical trials for the treatment of large number of diseases. For example, three different humanised Fab’ fragments are in phase II and III of drug development for cancer, rheumatoid arthritis, and age-related macular degeneration. Furthermore, a chimeric Fab’ has been approved for prevention of blood clotting and refractory unstable angina [9, 36].
Fed-batch fermentations are preferable to batch operation for the production of antibody fragments since batch methods usually result in low biomass concentrations with reported titres from 40 mg/L [7] to 450 mg/L [25]. On the other hand, fed-batch fermentations using a highly defined medium can result in levels up to 50 g/L of dry cell weight (DCW) [18] and titres up to 1–2 g/L [10]. Titres as high as 3 g/L using an optimised expression vector under non-limited growth conditions have been reported [20].
This study examines the effect of DOT levels and agitation rates on Fab’ production by E. coli at 20-L scale. The principal aim was to design and carry out experiments to test the hypothesis that increased Fab’ titres can be achieved in a fermenter system that is not oxygen limited. The questions tackled by this research have been addressed by comparison of a batch-fed fermentation of an oxygen deprived culture with those growing in environments with constant DOT levels of 30% or 50%. In particular, the effect of DOT and agitation rate on growth kinetics, Fab’ yield and cellular localisation were determined.
Furthermore, the interaction between operating parameters and cell productivity has been evaluated by using statistical design of experiments (DoE). It is hoped that the results will help guide future optimisation studies.
Materials and methods
Strain
Escherichia coli strain W3110 pAC tAC 4D5 Fab’ provided by Celltech Chiroscience Ltd, Slough, UK was used to prepare a working cell bank (WCB) as described elsewhere [8]. The plasmid pAC tAC encoded the light chain and heavy chain Fd’ fragment of the parent antibody under transcriptional control of the E. coli tac promoter. Each antibody chain was preceded by the E. coli ompA signal sequence to direct expression to the periplasmic space.
Inoculum development
Cultures of 0.25 mL were first grown in 250 mL of complex media (2xYT) in shake flasks. The starter cultures were incubated for 8 h in an orbital shaker (200 rpm) at 30°C. Then an aliquot was taken and the optical density (OD) at 600 nm measured. Samples of 25 mL culture (OD600 2.0–3.0) were transferred to defined media flasks of 250 mL working volume. The flasks were incubated for 12 h in an orbital shaker (200 rpm) at 30°C in order to obtain the required OD600 of 1.5–2.0. This provided an inoculum in mid-exponential growth phase, and resulted in an initial OD600 of 0.2–0.3 in the 20-L fermenter.
The fermenter had an initial working volume of 12.5 L and contained the same defined medium as the shake flasks. For the pilot plant scale experiments, a seed fermenter of 15 L working volume was cultivated in order to obtain the required OD600 of 6.0–6.5 before inoculating the 450 L fermenter. The fermenter had an initial working volume of 300 L and contained the same defined medium as the seed fermenter. The preparation of the medium was as previously described [8].
Fermentation protocol and setup
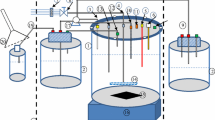
Fermentations were carried out at 20-L and 450-L scale in computer-controlled bioreactors (Applikon, Schiedam, Holland and Chemap AG, Volketswil, Switzerland, respectively). The 20-L vessel was sterilised using a steam jacket and a holding time of 20 min at 121°C. The pH was measured using a pH probe (Broadley Technologies Ltd, Bedford, UK) and maintained at 6.95±0.05 by computer-controlled additions of either sulphuric acid (5% v/v) or ammonia (15% v/v). The temperature was maintained at 30±0.5°C by using a heated jacket. The temperature was reduced to 27°C before induction in order to promote correct product folding. The agitation rate was set at either 500 rpm or 1,000 rpm. DOT was monitored using a DOT probe (Broadley Technologies Ltd). Fermenter exit/inlet gas compositions were monitored by mass spectroscopy (MM8 80 Instrument, VG Gas Analysis Ltd, Middlewich, UK). Data were logged using BioXpert software (Applikon, Schiedam, Holland). The airflow rate was set at 2.5 L/min in all the gas blending experiments. Gas blending was achieved by mixing the inlet air with pure oxygen (BOC Gases, Surrey, UK). The valve releasing oxygen into the inlet stream was set according to a computer control so as to maintain the DOT at 30% or 50%. The oxygen flow rate was varied under the control of the PID controller from 0 L/min to 2.5 L/min.
Samples of 30 mL of fermentation broth were taken in universal bottles every 4–6 h in order to measure OD600 and on which additions of glycerol, cation solution (magnesium sulphate and calcium chloride to enhance cell wall strength [40]), and lactose were based (see Table 1). Two glycerol additions were made during exponential phase after reaching the maximum specific growth rate and a third addition was made prior to addition of lactose (as detailed in Table 1). Lactose acted as the inducer for Fab’ formation and resulting in a reduction of the cell growth. This batch-fed feeding strategy was based on the protocol developed by Celltech Chiroscience Ltd. and is reported elsewhere [8, 23]. Assays were carried out on these samples to determine the concentrations of residual substrates, the levels of product formation and the biomass achieved.
Biomass measurement by optical density
The growth profile of the strain was followed by optical density measurements of the broth, measured at a wavelength of 600 nm with a spectrophotometer (DU-Spectrophotometer, Beckman Instruments Ltd., High Wycombe, UK). Samples were diluted with RO water (reverse osmosis purified) in order that the absorbance measurement was in the linear range of the instrument, i.e. 0–0.8 absorbance units.
Dry cell weight
Samples of 1 mL were placed into pre-weighed eppendorfs, and then spun down in a mini-centrifuge (Eppendorf Centrifuge 5415R, Hamburg) at 13,000 rpm for 5 min. The supernatant was discarded and the recovered pellets were dried to constant weight in an oven at 100°C for 24 h. The eppendorfs were then weighed again and the difference between the final weight and the empty eppendorfs taken as the DCW.
Glycerol, lactose and acetate analysis by HPLC
Glycerol, lactose and acetate concentrations were determined via high pressure liquid chromatography (HPLC). Samples of fermentation broth, 1 mL, were centrifuged at 13,000 rpm for 5 min, and the supernatant filtered using a 0.22-μm filter (Sigma-Aldrich, Dorset, UK). Glycerol, lactose and acetate in each sample were separated with an Aminex HPX-87H column (300 mm×7.8 mm) on a Summit HPLC system (Dionex Corp., CA, USA), with 5 mM sulphuric acid as the mobile phase. Glycerol and acetate concentration were determined using UV detection at 215 nm and lactose concentration was measured using a RI (refractive index) detector (Dionex Solution GmbH, Germering, Germany). The system was calibrated with known concentrations of glycerol, lactose and acetate each time the assay was performed.
Fab’ concentration measurement by ELISA
Fab’ concentration in the supernatant was measured by an enzyme linked immuno assay (ELISA) following periplasmic lysis as described in Bowering et al. [8]. ELISA plates were coated overnight with HP6045 (a mouse antihuman monoclonal antibody supplied by Celltech Chiroscience, Ltd.) in phosphate-buffered saline (PBS). After washing four times with distilled water, serial dilutions of samples and standards were performed on the plate in 100 μL sample/conjugate buffer and left to shake at 250 rpm, and room temperature for 1 h. After washing four times with dH2O (distilled water), the revealing antibody GD12 peroxidase (The Binding Site, Birmingham, UK) was added and diluted 1/1,000 in sample/conjugate buffer and left to shake at 250 rpm, and room temperature for 1 h. After washing four times with dH2O, tetramethyl benzidine (TMB) substrate was added and the absorbance at 630 nm was recorded using an automated plate reader. The concentration of Fab’ was calculated by comparison with purified Fab’ standards of the appropriate isotype. Assays were run at least in duplicate to ensure reproducible results.
Total protein assay
The Total protein assay was performed using a BioRad diagnostic kit (BioRad Laboratories GmbH, München, Germany) according to the manufacturer’s instructions. Assays were run at least in duplicate to ensure reproducible results within a range of ±4%.
Experimental design
Fermentations in the two different reactor systems were carried out:
-
1.
Fermentation at 450-L scale without gas blending. A single fermentation was performed to evaluate O2 levels at pilot scale using off-gas analysis.
-
2.
Fermentation at 20-L scale without gas blending. Experiments were performed in duplicate to evaluate O2 levels at bench scale based on off-gas analysis.
-
3.
Fermentation at 20-L scale with gas blending. A factorial 22 experiment [32] was performed to evaluate effects and interactions of agitation rate and DOT level on Fab’ yields and cell growth. As detailed in Table 2, four fermentations with different O2 transfer environments were set and analysed. All fermentations were carried out in duplicate.
Statistical analysis of the results obtained was carried using the data analysis and graphical data presentation program MINITAB (Minitab Ltd., Coventry, UK). A standard analysis of variance (ANOVA) was employed to identify and quantify effect and interactions of agitation rate and DOT level on the production of Fab’ fragments.
Results and discussion
Both the fermentations at 20-L and 450-L scale without gas blending had similar initial K La values. Overall volumetric mass transfer coefficients (K La) were estimated using the oxygen-balance technique, as described in Bailey et al. [5]. Furthermore, both fermentations were operated under cascade control to maintain a DOT level of 30%. Nevertheless, in both cases the DOT level dropped close to zero before induction. By contrast in fermentations which employed gas blending a constant level of DOT (ca. 30%) was maintained throughout the fermentation (see Fig. 1).

DOT profiles of Fab’ antibody fragment fermentation in a (filled diamond) 450-L scale with no gas blending and two 20-L scale fermentation dashed line with and solid line without gas blending. The arrow indicates the point of induction
The 450-L fermentation data were only used to assess whether a 20-fold increase in scale has an effect on the fermentation performance. Results at 450-L scale indicated total Fab’ levels of 75 mg/L, which represents 0.0018 mg of Fab’/L/g of lactose consumed. This value was found to be ~95% less when compared to a total Fab’ yield of 35 mg/L at 20-L scale, which represents 0.055 mg of Fab’/L/g of lactose utilised.
Results at pilot scale strengthened the hypothesis that O2 limitation could limit product formation as the culture at this scale observed longer periods with DOT levels near to zero resulting in lower product formation per lactose consumed than that at 20-L scale. It is conceivable that the long period of O2 depletion prior to induction (ca. 10 h) may have lead to a reduction in viable cells and this contributed to the reduced Fab’ production obtained compared to that at 20-L scale.
This study focuses on comparison of fermentations with and without gas blending at 20-L scale to allow the effect of gas blending on final Fab’ yield, growth kinetics and cellular localisation of the product to be identified. Furthermore, the effect of mixing and oxygen transfer on biomass levels and product yields were evaluated using a factorial design 22 . In Table 3, the performances of these five different fermentations are summarised. As can be seen, an increase in the production of Fab’ by 77% was achieved using a low agitation rate independent of the DOT level. Furthermore, similar biomass yields on glycerol (Y X/glycerol of 0.35 ± 0.01) were observed in all fermentations with and without gas blending, except GB-4 (+ rpm and + DOT) that exhibited reduced biomass and product yields. Results suggest that acetate levels be related to biomass yields as GB-4 observed the highest final acetate level and the lowest maximum biomass yield. Furthermore, specific oxygen uptake rates seem to be related to Fab’ yield as the non-GB and GB-4 observed the lowest specific oxygen uptake rates and the lowest Fab’ yields. The following sections discuss each of these aspects in turn.
Effect of gas blending on biomass levelsss
A maximum specific growth rate of 0.17±0.02/h was obtained for all fermentations with and without gas blending. In Fig. 2a, biomass accumulation profiles of fermentations GB-1 (−rpm and −DOT), GB-4 (+ rpm and + DOT) and the non-GB fermentation are presented. Fermentations GB-2 and GB-3 exhibited very similar profiles to GB-1 (results not shown). Biomass levels of ca. 30 g/L were obtained in all

Effect of gas blending on final biomass levels (a) and glycerol uptake rates (b) in two different fermentation systems at 20-L scale (from left to right): (open circle) non-gas blending, gas blending: (filled circle) GB-1 (−rpm, −DOT), and (filled square) GB-4 (+rpm, +DOT). Error bars represent standard deviations of duplicate fermentations
20-L fermentations except for that when the most extreme conditions of DOT and agitation speed were used (GB-4). For the fermentation GB-4 (+ rpm and + DOT) the biomass levels produced were 17% lower than the average level of all the other fermentations.
The lower level of biomass in GB-4 was unexpected and the protein concentration was therefore measured in the supernatant, to determine whether the reduced biomass levels could be attributed to increased levels of cell lysis or shear damage under conditions of high agitation and aeration. Results presented in Table 3 showed total protein values of 25–30 mg/L were obtained in supernatants of all fermentations, effectively ruling out cell lysis as a reason for the differences in biomass levels.
As can be seen in Table 3, GB-4 (+ rpm and + DOT) exhibited the highest concentration of acetate at the end of the fermentation. Although cycles of excretion and re-consumption could be expected throughout the fermentation, it is an indication that the acetate levels reached in GB-4 most probably where higher than those of the other fermentations. This may explain the low biomass level achieved as it is known acetic acid leads to growth inhibition of E. coli [24]. For example, it has been shown that in glucose-feedback-controlled fed-batch fermentations, biomass concentrations up to 30 g/L were produced while accumulating 2 g/L of acetate. When 8 g/L of acetate was accumulated the biomass concentration was less than 10 g/L. The acetate level realised was strain specific [30]. The excretion of acetate under fully aerobic conditions was not expected because it is generally accepted that E. coli does not produce acetate when growing on carbon sources such as glycerol [19]. This was explained to be an effect of the restricted substrate uptake and the maximal uptake rate was not suppose to reach the threshold required to trigger acetate excretion [20]. However, a study by Korz et al. [26] has shown that acetate is produced up to 3.3 g/L in high cell density fed batch cultures grown on glycerol. Based on their findings the authors concluded that the critical growth rate with regard to acetate formation is ca. 0.17/h . This was supported by the work of Macaloney et al. [31], who also found acetate excretion in fed-batch cultures of E. coli grown on glycerol when a critical specific growth rate was exceeded. Since the specific growth rates obtained in all GB and non-GB fermentations are very close to the reported threshold value the excretion of acetic acid during the growth phase is likely.
In order to help to explain our findings, glycerol utilisation profiles were determined. In Fig. 2b, profiles of fermentations GB-1 (−rpm and −DOT), GB-4 (+ rpm and + DOT) and the non-GB fermentation are presented. Fermentations GB-2 and GB-3 exhibited similar profiles to GB-4 and GB-1, respectively (results not shown). Values of specific glycerol uptake rate could not be estimated due to the substrate oscillations related to the batch-fed feeding strategy. Nevertheless, it is noticeable that a twofold higher glycerol level at the time of induction could be seen in the fermentations with high agitation rate (GB-2 and GB-4) compared to the fermentations with low agitation rate (GB-1 and GB-3). The difference in agitation rate and therefore mass transfer conditions could have affected the fermentation performance with regard to glycerol utilisation and acetate formation and thus could have caused an additional strain on the cells leading to a reduction in glycerol consumption after induction. It has been reported that acetate is produced when carbon flux exceeds the biosynthetic demands of the capacity for energy generation within the cell in a glucose-based system [12, 27, 37]. Therefore, a similar phenomenon in this glycerol-based system could have occurred and the carbon flux was exceeded in GB-2 and GB-4 due to the high mass transfer as a result of the higher agitation rate. On the other hand, Table 3 shows that the final acetate level in the non-GB fermentation was similar to that observed in GB-2 despite using gas blending. Furthermore, the acetate level in GB-2 was 47% lower than that in GB-4. This last result suggests that oxygen has a significant effect on acetate formation, as the main difference between GB-2 and GB-4 is the DOT level.
It is known that high concentrations of oxygen can be toxic, highly reactive and can spontaneously cause unwanted oxidation reactions within the cell [29]. On the other hand, levels up to 100% air saturation in E. coli fermentations using DOT control have been reported with no inhibitory effect on biomass formation [28]. All fermentations in this work used oxygen levels up to 90% air saturation to maintain a constant DOT of 30% or 50%. Nevertheless, GB-4 (+ rpm and + DOT) observed the highest concentrations of dissolved oxygen at the highest agitation rate. It has been found that the catabolic activities of E. coli are affected by the actual oxygen availability per unit of biomass rather than by the residual dissolved oxygen concentration of the culture [1]. Therefore, as GB-4 was the culture with highest DOT level at the highest agitation rate, it is speculated that toxic concentrations of oxygen in the cell could have been reached in this culture due to high transfer rates of O2. For example, oxygen enriched E. coli batch cultures showed that an increase of oxygen supply decreased both the glucose based biomass yield and maximum specific growth rate in addition to increasing excretion of the by-product acetate [34]. It was only hypothesised that the intermediates produced as a result of the reduction of an oxygen molecule were themselves toxic. Further work is needed to evaluate whether similar behaviour is observed when glycerol is used as the main C-source and a mechanistic explanation can be proposed.
Effect of gas blending on Fab’ production
In this system, lactose acts as inducer of product formation and therefore a reduction of growth is expected due to the switch of metabolism to product expression leading to a redirection of carbon and energy away from cell growth and towards product formation. Results shown in Fig. 3 indicate the switch of metabolism after induction based on the pH profiles. A pH increase in the culture broth is likely due to the consumption of amino acids for energy generation and the production of ammonia ions associated with the proton symport for lactose uptake [39]. As cells adapted to lactose metabolism, medium pH dropped again. Fermentation GB-4 observed the latest switch of metabolism (~40 h), possibly due to the effect of the high glycerol concentration on lactose consumption. It has been shown that excessive uptake of carbon sources such as glycerol or lactose can be deleterious to E. coli cells as a consequence of the formation of high levels of the toxic compound methylglyoxal in the case of glycerol or due to the collapse of the proton motive force when exposed to high levels of lactose [13, 21]. The deleterious effects of both glycerol and lactose are in agreement with the low levels of Fab’ seen in fermentation GB-2 and GB-4 and the high levels of GB-1 and GB-3 (these fermentation observed low levels of glycerol at the point of induction). Nevertheless, fermentation GB-2 observed a similar high level of glycerol at the point of induction, but the time of switch of metabolism was similar to that of the other fermentations. This suggests again that the effect of oxygen available to the cell is relevant as the main difference between GB-2 and GB-4 is the DOT level.

Effect of gas blending on metabolic switch from glycerol to lactose based on pH profiles in two different fermentation systems at 20-L scale with gas blending (GB) and without gas blending (non-GB): a GB-1 (−rpm, −DOT); b GB-2 (+rpm, −DOT); c GB-3 (−rpm, +DOT); d GB-4 (+rpm, +DOT); and e non-GB
After induction, cell growth and metabolic activity are often influenced by the on-going expression of recombinant proteins within the cell [3, 4, 33]. This metabolic activity could be related to respiratory data such as RQ values and specific oxygen uptake rates. For example, values of RQ were in agreement with the switch of metabolism. A switch of RQ from ca. 0.7 to 1.0 was observed at the same time as that of the pH peak in all fermentations. Furthermore, when comparing specific oxygen uptake rates obtained from off-gas analysis during the induction period (see Table 3), values of 3 and 4 mmol/g/h for the non-GB and GB-4 fermentations, respectively were determined. Both fermentations exhibited similar low Fab’ yields. By contrast, fermentations GB-1 and GB-3 that both yielded higher Fab’ titres observed values of ~10 mmol/g/h . It appears that the higher the oxygen consumption is by the cells the higher are metabolic activity and therefore product formation. Nevertheless, at this point it is not possible to determine what percentage of oxygen is related to product metabolism as opposed to cell maintenance and therefore further work is needed to establish if a direct correlation between specific oxygen uptake rate and product yield exist.
In Table 4, the effect of gas blending on product formation is summarised. The Fab’ titres were measured at the time of harvest (~46 h) for all fermentations. Based on these results, a 77% increase in Fab’ yield was achieved when comparing the gas blending system with low agitation rate with the non-gas blending fermentation. However, it can also be suggested that high agitation rate had a detrimental effect regardless of the DOT level used in the gas blending system. In the case of the fermentations without gas blending, the cells ran out of oxygen after 30 h (i.e. during the induction period). In this case, expression of Fab’ possibly occurred under conditions of low oxygen, which might have limited product synthesis. In terms of economics, low agitation and low DOT (GB-1) is a less expensive system as it requires less oxygen while obtaining similar levels of Fab’ and biomass when compared to low agitation and high DOT (GB-3). This will become an important consideration at large-scale. Furthermore, in all gas blending fermentations, the level of periplasmic product localisation was high, with >85% being retained in the periplasm.
A comparison of the final volumetric Fab’ concentrations in Fig. 4 shows that conditions of low agitation and low DOT (GB-1) and low agitation and high DOT (GB-3) gave threefold higher concentrations of Fab’ compared to those of the other fermentations.

Final periplasmic and extracellular Fab’ antibody fragment concentration in two different fermentation systems: 20-L scale with gas blending according to the 22 factorial DoE (see Table 1), and 20-L scale without gas blending. Error bars represent standard deviations of duplicate fermentations
Statistical analysis
DoE methodology [32] allowed obtaining further information towards the optimisation of this fermentation process. In Table 5, the ANOVA to evaluate the effects and interactions of DOT and agitation speed on Fab’ production is summarised. It can be observed, that the effect of A (agitation speed) is significant for Fab’ titres as the P value is 0.001. This would imply that there is 95% probability that agitation rate has a large impact on Fab’ yield when considering experimental error from duplicates. The effect of B (DOT) and its interaction with A appeared to be small relative to the main effect of A and seems to be not statistically significant (P>0.01). Results suggest that from the two engineering factors studied, agitation rate is the main factor to focus on towards optimisation of this fermentation process. Nevertheless, further work is needed to evaluate different levels of agitation rate to study the effect on Fab’ yields and by-product formation in more detail.
Conclusions
The results reported in this work suggest a fermentation operating strategy that employs gas blending can increase Fab’ titres in bioreactor systems that are not oxygen limited when appropriate mass transfer conditions are achieved. Gas blending was successful in maintaining constant levels of DOT at 20-L scale with a resulting increase in the production of Fab’ of 77% at an agitation rate of 500 rpm independent of the DOT level, compared to operation at the same scale, but without gas blending. In all cases, a level of product localisation in the periplasm of 84–93% was also obtained.
An increase in Fab’ yields due to gas blending suggests that protein synthesis is related to oxygen consumption, whereby a constant oxygen supply at an appropriate agitation rate supports higher product titres. Results presented here suggest an alternative approach to increase Fab’ yields. However, further work should be carried out towards improving the feeding strategy partly because the current batch-fed feeding strategy leads to glycerol oscillations that might be limiting Fab’ productivity.
References
Alexeeva S, Hellingwerf KJ, Teixeira de Mattos MJ (2002) Quantitative assessment of oxygen availability: perceived aerobiosis and its effect on flux distribution in the respiratory chain of Escherichia coli. J Bacteriol 184(5):1402–1406
Amanullah A, Tuttiet B, Nienow AW (1998) Agitator speed and dissolved oxygen effects in Xanthan fermentations. Biotech Bioeng 57(2):198–210
Andersson L, Yang S, Neubauer P, Enfors SO (1996) Impact of plasmid presence and induction on cellular response in fed-bacth cultures of Escherichia coli. J Biotechnol 46:255–263
Andersson L, Strandberg L, Enfors SO (1996) Cell segregation and lysis have profound effects on the growth of Escherichia coli in high cell density fed batch cultures. Biotechnol Prog 12:190–195
Bailey JE, Ollis DF (1986) Biochemical engineering fundamentals, 2nd edn. McGraw-Hill International editions, Singapore
Bhattacharya SK, Dubey AK (1997) Effects of dissolved oxygen and oxygen mass transfer on overexpression of target gene in recombinant E. coli. Enzyme Microb Technol 20:355–360
Berry MJ, Wattam TAK, Willets J, Linder N, De Graaf T, Hunt T, Gani M, Davis PJ, Porter P (1994) Assay and purification of Fv fragments in fermenter cultures: design and evaluation of generic binding reagents. J Immunol Methods 167:173–182
Bowering LC, Bracewell DG, Keshavarz-Moore E, Hoare M, Neil A, Weir C (2002) Comparison of techniques for monitoring antibody fragment production in E. coli fermentation cultures. Biotechnol Prog 18:1431–1438
Brekke OH, Loset GA (2003) New technologies in therapeutic antibody development. Curr Opin Pharmacol 3:544–550
Carter P, Kelly RF, Rodriguez ML, Snedcor B, Covarrubias M, Velligan MD, Wong WLT, Rowland AM, Kotts CE, Carver ME, Yang M, Bourrel JH, Shepard HM, Henner D (1992) High level E. coli expression and production of a bivalent huminizes antibody fragment. Bio Technol 10:163–167
De Leon A, Hernandez V, Galindo E, Ramirez OT (2003) Effects of dissolved oxygen tension on the production of recombinant penicillin acylase in Escherichia coli. Enzyme Microb Tech 33:689–697
El-Mansi EM, Holms WH (1989) Control of carbon flux to acetate excretion during growth of Escherichia coli in batch and continuous cultures. J Gen Microbiol 135(Pt 11):2875–2883
Ferguson GP, Tötemeyer S, MacLean MJ, Booth IR (1998) Methylglyoxal production in bacteria: suicide or survival? Arch Microbiol 170:209–219
Flores F, Torres LG, Galindo E (1994) Effect of the dissolved-oxygen tension during cultivation of Xanthomonas campestris on the production and quality of Xanthan gum. J Biotech 34(2):165–173
Gibbs PA, Seviour RJ (1996) Does agitation rate and/or oxygen saturation influence exopolysaccharide production by Aurebasidium pullulans in batch culture? Appl Microbiol Biotechnol 46:503–510
Gupta A, Rao G (2003) A study of oxygen transfer in shake flasks using a non-invasive oxygen sensor. Biotechnol Bioeng 5(843):351–358
Harris B (1999) Exploiting antibody-based technologies to manage environmental pollution. Trends Biotech 17(7):290–296
Harrison JS, Keshavarz-Moore E (1996) Production of antibody fragments in Escherichia coli. Recombinant DNA biotechnology III: the integration of biological and engineering sciences 182
Holms WH (1986) The central metabolic pathways of E. coli relationship between flux and control at a branch point, efficiency of conversion to biomass, and excretion to acetate. Curr Top Cell Reg 28:69–105
Holms WH (1996) Flux analysis and control of the central metabolic pathways in E. coli. Microb Rev 19:85–116
Hogema BM, Arents JC, Baden R, Postma P (1999) Autoregulation of lactose uptake through the LacYpermease by enzyme IIA Glc of the PTS in Escherichia coli K12. Mol Microb 31:1825–1833
Horn U, Strittmatter W, Krebber A, Knupfer U, Kujau M, Wenderoth R, Muller K, Matzku S, Pluckthun A, Riesenberg D (1996) High volumetric yields of functional dimeric antibodies miniantibodies in Escherichia coli, using an optimized expression vector and high-cell-density fermentation under non-limited growth conditions. Appl Microbiol Biotechnol 46:524–532
Humphreys DP, Carrington B, Bowering LC, Ganesh R, Sehdev M, Smith BJ, King LM, Reeks DG, Lawson A, Popplewell AG (2002) A plasmid system for optimization of Fab’ production in Escherichia coli: importance of balance of heavy chain and light chain synthesis. Protein Expr Purif 26(2):309–320
Johnston W, Cord-Ruwisch R, Cooney MJ (2002) Industrial control of recombinant E. coli fed-batch culture: new perspectives on traditional controlled variables. Bioprocess Biosystems Eng 25:111–120
King DJ, Byron OD, Mountain A, Weir N, Harvey A, Lawson DG, Proudfoot KA, Baldock D, Harding E, Yarranton GT, Owens RJ (1993) Expression, purification and characterisation of B72.3 Fv fragments. Biochem J 290:723–729
Korz DJ, Rinas U, Hellmuth K, Sanders E A, Deckwer W-D (1995) Simple fed-batch technique for high cell density cultivation of Escherichia coli. J Biotech 39:59–65
Lee SY (1996) High cell-density culture of Escherichia coli. Trends Biotechnol 14:98–105
Li X, Robbins JW Jr, Taylor KB (1992) Effect of the levels of dissolved oxygen on the expression of recombinant proteins in four recombinant Escherichia coli strains. J Ind Microbiol 9(1):1–9
Lodish H, Berk A, Lawrence Zipursky S, Matsudaira P, Baltimore D, Darnell J (2000) Molecular cell biology, 4th edn. Freeman and Company, New York
Luli G, Strohl WR (1990) Comparison of growth, acetate production, and acetate inhibition of Escherichia coli strains in batch and fed-batch fermentations. App Environ Microbiol 56(4):1004–1011
Macaloney G, Hall JW, Rollins MJ, Draper I, Anderson KB, Preston J, Thompson BG, McNeil B (1997) The utility and performance of near-infra red spectroscopy in simultaneous monitoring of multiple components in a high cell density recombinant Escherichia coli production process. Bioprocess Eng 17:157–167
Montgomery DC (2001) Design and analysis of experiments, 5th edn. Wiley, USA
Neubauer P, Lin HY, Mathiszik B (2003) Metabolic load of recombinant protein production: inhibition of cellular capacities for glucose uptake and respiration after induction of a heterologous gene in Escherichia coli. Biotech Bioeng 83:53–64
O’Bierne D, Hammer G (2000) Oxygen availability and the growth of Escherichia coli W3110: a problem exacerbated by scale-up. Bioprocess Biosyst Eng 23:487–494
Pollard DJ, Kirschner TF, Hernandez D, Hunt G, Olewinski R, Salmon PM (2002) Pilot-scale process sensitivity studies for the scale-up of fungal fermentation for the production of pneumocandis. Biotech Bioeng 78(3):270–279
Roque AC, Lowe CR, Taipa M (2004) Antibodies and genetically engineered related molecules production and purification. Biotechnol Prog 20:639–654
Rocha I, Ferreira EC (2002) On-line simultaneous monitoring of glucose and acetate with FIA during high cell density fermentation of recombinant E. coli. Anal Chim Acta 462:293–304
Sanden AM, Prytz I, Tubulekas I, Forberg C, Le H, Hektor A, Neubauer P, Pragai Z, Harwood C, Ward A, Picon A, De Mattos JT, Postma P, Farewell A, Nystrom T, Reeh S, Pedersen S, Larsson G (2003) Limiting factors in Escherichia coli fed-batch production of recombinant proteins. Biotechnol Bioeng 20;81(2):158–166
Straight J, Parulekar S, Jansen N (1989) Bacterial growth on lactose: an experimental investigation. Biotechnol Bioeng 34:705–716
Vaara M (1992) Agents that increase the permeability of the outer membrane. Microbiol Rev 56:395–411
Acknowledgements
Special thanks to Helen Baldascini, and Colin Jacques. Financial support from Consejo Nacional de Ciencia y Tecnología (CONACyT) is gratefully acknowledged. Support for the Innovative Manufacturing Research Centre funded by the Engineering and Physical Sciences Research Council (EPSRC) is also gratefully acknowledged.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
García-Arrazola, R., Dawson, P., Buchanan, I. et al. Evaluation of the effects and interactions of mixing and oxygen transfer on the production of Fab’ antibody fragments in Escherichia coli fermentation with gas blending. Bioprocess Biosyst Eng 27, 365–374 (2005). https://doi.org/10.1007/s00449-005-0414-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00449-005-0414-4