
Abstract
Drought, defined as a marked deficiency of precipitation relative to normal, occurs as periods of below-average precipitation or complete failure of precipitation inputs, and can be limited to a single season or prolonged over multiple years. Grasslands are typically quite sensitive to drought, but there can be substantial variability in the magnitude of loss of ecosystem function. We hypothesized that differences in how drought occurs may contribute to this variability. In four native Great Plains grasslands (three C4- and one C3-dominated) spanning a ~ 500-mm precipitation gradient, we imposed drought for four consecutive years by (1) reducing each rainfall event by 66% during the growing season (chronic drought) or (2) completely excluding rainfall during a shorter portion of the growing season (intense drought). The drought treatments were similar in magnitude but differed in the following characteristics: event number, event size and length of dry periods. We observed consistent drought-induced reductions (28–37%) in aboveground net primary production (ANPP) only in the C4-dominated grasslands. In general, intense drought reduced ANPP more than chronic drought, with little evidence that drought duration altered this pattern. Conversely, belowground net primary production (BNPP) was reduced by drought in all grasslands (32–64%), with BNPP reductions greater in intense vs. chronic drought treatments in the most mesic grassland. We conclude that grassland productivity responses to drought did not strongly differ between these two types of drought, but when differences existed, intense drought consistently reduced function more than chronic drought.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Drought defined meteorologically as a “prolonged absence or marked deficiency of precipitation” (World Meteorological Organization 1992; Slette et al. 2019) affects ecological patterns and processes in most terrestrial biomes. While drought is often quantified in terms of a reduction in the amount of precipitation, there are additional dimensions of drought that can modify its impact on ecosystem structure and function (Luo et al. 2020). A given magnitude of drought may have varying impacts depending on drought timing (Jentsch et al. 2011; Zeiter et al. 2016; Denton et al. 2017) and duration (Hoover et al. 2014), as well as antecedent conditions (Sala et al. 2012; Gong et al. 2020). Other studies have concluded that there can be strong interactions between drought responses and the precipitation patterns occurring during drought years (Cherwin and Knapp 2012; Raz-Yaseef et al. 2010, 2012). Indeed, Felton et al. (2019) reported that precipitation pattern (event size, variability in the timing of events, etc.) affects ecosystem function primarily during years with below-average precipitation. In other words, precipitation patterns matter most when water is limiting.
In an analysis of precipitation regimes for extreme wet and dry years across major terrestrial biomes, Knapp et al. (2015) reported that, beyond precipitation amount, two precipitation attributes: (1) a lack of large rain events and (2) an increase in the average length of the dry periods between precipitation events were most important for distinguishing extreme wet and average years from extreme dry years. Thus, natural droughts tend to occur when reduced precipitation amounts are a result of smaller event sizes and longer dry periods relative to average or wet years. Disentangling the relative importance of these two dimensions of drought on ecosystem responses is a challenge under natural conditions because these attributes are often correlated (Knapp et al. 2015). However, with the intensification of the hydrological cycle resulting in an increase in large precipitation events and longer intervening dry periods (Easterling et al. 2000; Zobel et al. 2018; Papalexiou and Montanari 2019), there is a need to know how variations in these two precipitation attributes may alter ecosystem sensitivity to drought.
To assess how responsive ecosystems are to different types of drought, we experimentally imposed extreme drought for 4 years by reducing precipitation inputs into native grasslands either by (1) reducing each event by 66% during the entire growing season (hereafter chronic drought) or (2) completely excluding all rain events for a shorter portion of the growing season (hereafter intense drought). While both types of drought excluded similar total amounts of rainfall, the chronic drought treatment primarily reduced event size, whereas the intense drought treatment extended the length of dry periods between events. Both theory and previous research on how precipitation regimes affect ecosystem function suggest that mesic grasslands should be more sensitive to extended dry periods, but that arid grasslands respond more strongly to changes in event size (Knapp et al. 2008; Heisler-White et al. 2009). Thus, we expected that grasslands spanning a precipitation gradient would respond differently to these two types of drought. We tested this hypothesis by imposing chronic and intense drought in four grasslands that spanned a mean annual precipitation (MAP) range of 380–880 mm. We imposed these droughts over four consecutive years, allowing us to assess how a third dimension of drought—duration—influenced responses to these two drought treatments. Here, we report responses in annual aboveground net primary production (ANPP) throughout this multi-site 4-year experiment and contrast annual ANPP responses with the cumulative effect of drought on belowground net primary production (BNPP) measured during the final year of the experiment.
Materials and methods
Study sites
We established four identical drought experiments in native Great Plains grasslands in 2013. The four sites spanned a ~ 500-mm precipitation gradient with varying soil texture (Burke et al. 1989, 1991). Sites included a northern mixed-grass prairie at the High Plains Grasslands Research Center (HPG) near Cheyenne, WY, a semi-arid short-grass steppe at the Central Plains Experimental Range in NE Colorado (SGS), a southern mixed-grass prairie at the Hays Agricultural Research Center (HYS) near Hays KS, and an annually burned tallgrass prairie watershed at the Konza Prairie Biological Station (KNZ; Fig. 1, Table 1). The dominant species in these grasslands shift along the rainfall gradient from Bouteloua gracilis at SGS, to Pascopyrum smithii and B. gracilis at HPG to B. curtipendula, Schizachyrium scoparium, and P. smithii at HYS, and finally to Andropogon gerardii, and Sorghastrum nutans at KNZ. All sites were strongly C4 dominated except HPG where the C3 P. smithii dominates. Physiological differences between C4 and C3 species are well known and were consistent with the traits of these dominant species (see Monson et al. 1986; Taylor et al. 2010; Morgan et al. 2011). Within the C4 grasses, growth rates and productivity increased from the drier to more mesic sites (McCulley et al. 2005), but all grass species rely predominantly on shallow soil layers for water uptake (Dodd and Lauenroth 1997; Nippert and Knapp 2007).

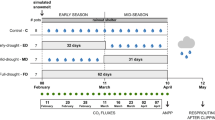
a Site locations within the approximate ranges of shortgrass, mixedgrass, and tallgrass prairie of the Great Plains, U.S.A. b Example of how the two drought treatments differed. Control (gray) and treatment (color) precipitation (mm) by month for (left) intense (full exclusion over a period of the growing season) and (right) chronic (66% reduction throughout the growing season) droughts at SGS in 2017
At each site, 10 blocks, each containing three 6 × 6 m plots (n = 30 plots total per site), were established in an area with relatively homogeneous vegetation and minimal slope. Blocks were separated by at least 5 m, and plots within each block were located at least 2 m apart. Each plot was trenched around the perimeter to 40–50 cm and lined with plastic sheeting to hydrologically isolate them from the surrounding soil. Aboveground, 10 cm of aluminum flashing was partially buried around the perimeter of each plot to limit overland flow. Commercially available greenhouse structures (Stuppy, Inc, North Kansas City, MO) were installed over 20 of the plots. The frames were fitted with clear polycarbonate plastic (Dynaglas, Palram Americas Ltd) roofs designed to deflect either a fixed portion of each rain event (Yahdjian and Sala 2002) or completely exclude all rain. Because there were no side or end walls to impede airflow, the impacts of these roofs on plot microclimatic and plant ecophysiological processes are minimal (Loik et al. 2019). Only the central 5 × 5 m area of the plot was dedicated to sampling to provide a buffer that minimized edge effects and occasional wind-driven rain inputs (Griffin-Nolan et al. 2019). Each 5 × 5 m plot was divided into four 2 × 2 m subplots (each with a 50 cm buffer) and randomly assigned to either plant species composition sampling or destructive sampling measurements. None of the sites were grazed by large ungulates over the study period.
Drought treatments
The experiment consisted of three drought treatments that were randomly assigned to the three plots within each block at each site: ambient precipitation, chronic drought—a 66% reduction in each rainfall event throughout the duration of the growing season (approximately April 1–September 15), and intense drought—a complete exclusion of every rain event for a shorter portion of the growing season (typically late-April, early-May to mid, late-August depending on ambient precipitation patterns). The chronic treatment was achieved using a roof comprised of clear corrugated polycarbonate plastic sheeting cut into 15.25-cm strips spaced to cover 66% of the roof area. The intense treatment was achieved with a complete roof made from panels of clear corrugated polycarbonate sheeting. All roof edges were equipped with rainfall gutters and corrugated tubing to collect and divert the rainfall from each plot. Roofs were installed at the beginning of each growing season and removed at the end of each growing season. The ambient precipitation treatments did not have greenhouse shelters in place, but the widely spaced (1.2 m) metal greenhouse support structures would be expected to have minimal microclimatic effects. Aboveground ambient precipitation and temperature were monitored via a tipping rain gauge and a relative humidity sensor (Campbell Sci CS215). Drought treatment precipitation levels were calculated based on expected reductions from ambient conditions. Rainfall event sizes were compared to and adjusted based on soil moisture sensor data to account for lateral rainfall blowing into the plots. All rainfall events less than 0.3 mm were excluded from calculations and gaps in ambient records were filled using nearby rain gauges.
The two drought treatments were imposed beginning in 2014. Both drought treatments excluded similar amounts of precipitation each growing season, but other precipitation attributes (event size, number, time between events) differed markedly (Fig. 2, Tables S1–S3). We estimated aboveground net primary production (ANPP g m−2) at the end of the growing season in the pre-treatment year (2013) and annually from 2014 to 2017. At the end of each growing season, all aboveground biomass was harvested to the ground surface within three 20 × 50 cm quadrats randomly located within the 2 × 2 m subplot dedicated to biomass sampling in each plot at each site. Quadrats were flagged to prevent subsequent resampling. Biomass was separated in the field into grass, forb, woody (if present), and previous year’s dead material. Biomass samples were dried at 60 °C for 48 h and then weighed to the nearest 0.01 g.

Precipitation attributes (mean ± SE) for ambient conditions and the two drought treatments combined across the four grasslands (Fig. 1). Different letters indicate significant differences using Tukey adjusted pairwise comparisons: α = 0.05. a Growing season (April 1–September 15) precipitation (mm); b Average number of rain events > 0.3 mm during the growing season; c Average size (mm) of a rain event during the growing season; d length (days) of average dry period experienced between rain events during the growing season. See Table S4 for summary statistics for all sites pooled together as shown above
In 2017, the last year of the imposed drought, we estimated belowground net primary production (BNPP g m−2) using root ingrowth cores to measure fine root mass production. At the start of the growing season, three soil cores (5-cm diameter, 20-cm deep) were taken from the same subplot within each plot at each site that was not otherwise allocated for community composition or ANPP. A 20-cm cylindrical mesh basket filled with sieved, root-free soil (previously collected adjacent to the site) was placed into each of the holes created by the soil cores. Any space between the ingrowth core and the intact soil was carefully filled with sieved soil. All ingrowth cores were removed at the end of the growing season, placed in plastic bags, and stored at 4 °C until processing. Each core was cut in half at a depth of 10 cm, yielding two depth increments (0–10 cm and 10–20 cm below soil surface) that were each processed individually. All soil was washed off the roots by first wet sieving the ingrowth core depth increment to remove excess soil and then submerging the remaining sample in a shallow bowl of water and carefully picking out roots and removing any attached soil. Roots were dried at 60 °C for at least 48 h and were weighed to the nearest 0.0001 g.
Differences in drought characteristics (growing season precipitation (GSP), event number, event size, and average number of dry days between events) across all sites were determined using analysis of variance (ANOVA) with site, treatment, and their interaction as fixed effects for each drought variable modeled separately. All years were pooled together as the actual drought characteristics of each year were independent through time. Drought impacts on ANPP were assessed with separate repeated measures mixed-model ANOVAs for each site with year, treatment, and their interaction as fixed effects and sampling block as a random effect due to repeated measurements in the same plots over time. Each site was assessed independently allowing us to focus on responses to the two types of drought within each site, rather than on differences driven by the fivefold range in productivity across these different grasslands (Table 1). Lastly, the single-year estimates of BNPP and root depth distribution ratios were assessed by mixed-model ANOVA with site, treatment, and interactions as fixed effects and block as a random effect. These analyses were followed by Tukey HSD pairwise comparisons. All calculations assessed significance based on α = 0.05 and analyses were performed with R version 3.5.1 and the lme4 package (Bates et al. 2015).
Results
Drought attributes
Across all sites, intense and chronic droughts together reduced annual precipitation by ~ 48% (from 44% at KNZ to 51% at CHY) and growing season precipitation by ~ 62% (57% at HYS to 68% at SGS) compared to ambient conditions over the course of the experiment (F = 31.34, p < 0.001; Fig. 2; Tables S1–S5). Not surprisingly, event size was significantly reduced in the chronic treatment (F = 11.13, p < 0.001), with this effect also varying by site (F = 14.19, p < 0.001). Number of events differed significantly by treatment (F = 67.69, p < 0.001) but not site, with the intense drought reducing event number more than the chronic treatment. Lastly, the average number of dry days between rainfall events was, as expected, significantly different by treatment (F = 37.09, p < 0.001) but not site, with the intense drought treatment imposing a greater number of dry days than either the chronic or ambient treatments. In sum, both drought treatments reduced precipitation from ambient by similar amounts, with the chronic treatment primarily reducing event size and the intense treatment reducing event number and increasing the length of dry periods.
ANPP responses to drought
Prior to the installation of the rainfall exclusion infrastructure, overall levels of ANPP were not statistically different across treatments at any site in 2013 (p > 0.05). In 2013, ANPP at HPG was lower than expected due to previously identified negative legacy of the substantial 2012 drought in the Great Plains (Griffin-Nolan et al. 2018) which impacted this site more than the other three included in this study. Once the drought treatments were initiated, sites did differ dramatically in how ANPP responded to drought. Most notable was the overall lack of response of the C3-dominated HPG site to drought (ANPP was reduced by only 6%), with significant drought effects only evident in the first year (Fig. 3). A lack of ANPP reductions in response to summer drought has been noted previously in C3-dominated Northern Mixed grasslands in the central US-based largely on the timing of growth and water availability (Knapp et al. 2020; Frank 2007, see Discussion). In contrast, ANPP reductions for the C4-dominated sites averaged 33% (SGS = 28%; HYS = 37%; KNZ = 35%).

Aboveground net primary production (ANPP, mean ± SE) responses to 4 years of ambient (gray), chronic rainfall exclusion (orange), and intense rainfall exclusion (red) treatments in a–d four field sites (Fig. 1a). Included is the pre-treatment year (2013) and four treatment years (2014–2017). Different letters indicate significant differences using Tukey adjusted pairwise comparisons: α = 0.05
Beginning in 2014, each of the three C4-dominated sites responded differently to the drought treatments over time with most consistent statistically significant reductions in ANPP for the more mesic sites (Fig. 3; Table S6). Plots at the most xeric site (SGS), experienced two years (2015 and 2017) of significant declines in ANPP, but no significant effect in 2014 or 2016. At HYS and KNZ, drought treatments in 2014 resulted in significant declines in the intense treatment only; while for all subsequent years, both types of drought resulted in significant declines in ANPP (Fig. 3, all p < 0.05).
In almost all years, ANPP was reduced in droughted plots compared to ambient, but the different types of drought often did not differ from one another. However, responses to these two types of drought were not identical. In four cases, HPG in 2014, HYS in 2014, KNZ in 2014 and 2017, ANPP in the chronic treatment was not different from ambient, but ANPP was significantly reduced by the intense drought treatment. Finally, at the wettest site (KNZ), the two drought treatments differed from one another in 2015, with the intense treatment resulting in significantly lower ANPP than the chronic treatment. While this was the sole example of differential sensitivity between the two drought types, the trend in nearly all cases was for greater declines in ANPP in the intense than in the chronic drought treatment. Lastly, we observed little evidence of drought duration affecting ANPP, with only KNZ experiencing a significant treatment by year interaction (Table S6). The significant interaction at this site can likely be attributed to the differences between the intense and chronic droughts in 2015; however, this was not a directional response as the later years of the experiment did not experience the largest drought effects.
BNPP responses to drought
After 4 years of drought in 2017, BNPP to 20-cm depth declined significantly at each of the C4-dominated sites compared to ambient, although the decline was not statistically significant at HPG (Fig. 4; Tables S7). The relative magnitude of these responses (averaged across drought types) varied from 30–35% reductions at HPG and HYS to 50–65% reductions at SGS and KNZ, and in all cases, BNPP was reduced more than ANPP in the fourth year of the treatments (ANPP responses were 35–42% in year 4 in the C4 grasslands, Fig. 3). Similar to the aboveground patterns, greater reductions in BNPP were observed in the intense plots primarily at KNZ, suggesting that the intense drought differentially impacted both above and belowground productivity. In contrast, the ratio of fine root production in the shallow (0–10 cm) vs. deeper (10–20 cm) soil layers was significantly reduced at only the C3-dominated semi-arid site (HPG) with no shift to deeper root allocation in the C4-dominant, more mesic sites (Table S8).

a Response of belowground net primary production (BNPP, 0–20 cm, mean ± SE) in the final year (2017) of a 4-year period of either intense or chronic drought. Ambient treatments (gray), chronic rainfall exclusion (orange) and intense rainfall exclusion (red) with different letters indicating significant differences within each site. b The ratio of BNPP for shallow (0–10 cm) vs. deeper (10–20 cm) depths. Smaller values indicate more BNPP was allocated at the deeper soil depth
Discussion
Variability in grassland responses to extreme drought
Based on the previous studies, drought-induced reductions in productivity are expected to be greater (relative to ambient treatments) in more xeric than mesic ecosystems (Huxman et al. 2004; Vicente-Serrano et al. 2013; Knapp et al. 2015; Gherardi and Sala 2018; Maurer et al. 2020), although some exceptions to this pattern have been noted (Liu et al. 2013; Tielbörger et al. 2014). In our study, distinct differences in drought sensitivity related to mean annual precipitation (MAP) were not evident, perhaps due to the modest (~ 500 mm) precipitation gradient encompassed by these four grasslands. By comparison, Huxman et al. (2004) concluded that drought sensitivity decreased as MAP increased by comparing ecosystems spanning a > 2500-mm precipitation gradient. Average drought-induced reductions in ANPP for the three C4 grasslands in this experiment ranged from 28% at SGS to 35 and 37% at KNZ and HYS, respectively. The magnitude of this drought response compares favorably to responses to the natural 2012 extreme drought observed at these same sites (24–53%, Knapp et al. 2015). However, SGS was most sensitive to the 2012 drought; whereas, HYS was most responsive to the drought treatments in our study.
Somewhat surprisingly, in three of the four treatment years, there was no drought-induced reduction in ANPP in the C3-dominated grassland (HPG). This contrasts with the response of this grassland to the 2012 drought (Knapp et al. 2015) but is consistent with lack of ANPP response to drought measured in other C3-dominated grasslands (Frank 2007; Kreyling et al. 2008). Further, in experiments comparing responses of C3 vs. C4 grasslands to alterations in summer precipitation inputs, C3 grasslands were less responsive (Wilcox et al. 2015). It is likely that the timing of drought compared to when C3 northern mixed grassland species undergo biomass accumulation explains this variability in response (see Knapp et al. 2020). Drought that develops during the warm summer months is likely to have the least impact on ANPP in ecosystems dominated by C3 grasses that are more active in cooler months. Indeed, extensive ecophysiological studies comparing the C3 grass P. smithii (comprising as much as 70% of aboveground biomass at HPG) to the C4 grass B. gracilis (dominant at the SGS) have documented clear photosynthetic adaptations to cooler temperatures for P. smithii (Monson and Williams 1982, Monson et al. 1982, 1983), and greater water-use efficiency in B. gracilis, particularly in warmer months (Monson et al. 1986). Despite considerable overlap in seasonal growth dynamics between these two grasses, due to the growth habit of B. gracilis in warm microenvironments next to the ground surface (Monson et al. 1986), the accumulation of ANPP generally occurs earlier in the growing season in C3 grasslands, driven by winter and early spring precipitation, reducing the sensitivity of these grasslands to summer drought (Frank 2007).
Intense vs. chronic drought
Although the magnitude of precipitation reduction was similar for the chronic and intense drought treatments, they differed distinctly in two ways: (1) timing: the intense drought began later in the growing season and ended earlier than the chronic drought treatment, and (2) attributes: rain event number was much lower and the time between events (consecutive dry days) much longer in the intense drought treatment, whereas the average rain event size was much smaller in the chronic drought treatment (Fig. 2). As noted above, C3 grasslands would be expected to be most affected by differences in the timing of drought initiation in the spring. The lack of ANPP responses to both types of drought suggests that most aboveground growth had already occurred prior to the onset of significant water stress in both treatments. Across the three C4 grasslands, theory (Knapp et al. 2008) and past experiments (Knapp et al. 2001; Heisler-White et al. 2009; Thomey et al. 2011; Cherwin and Knapp 2012; Baez et al. 2013) suggest that xeric ecosystems should be more sensitive to changes in event size, whereas mesic systems should respond more to a lengthening of time between events. Our results do not support these predicted differences. When there was evidence of differences in sensitivity between the two types of drought, ANPP was always reduced more in the intense drought treatment (Fig. 3). This suggests that these grasslands were more sensitive to drought characterized by extended periods of no rain (intense) than to reductions in event size (chronic).
Duration of drought
Grassland productivity in any given year is well known to be sensitive to antecedent conditions from previous years (Seastedt and Knapp 1993; Oesterheld et al. 2001; Sala et al. 2012; Griffin-Nolan et al. 2018). We anticipated that as drought duration increased, reductions in ANPP relative to ambient treatments would increase, regardless of drought type (Orth et al. 2020). We found, however, little evidence of cumulative drought effects (Fig. 3), with the possible exception of KNZ. In this case, increased sensitivity to the intense drought treatment occurred in the second year of the experiment, but this pattern did not continue in later years, as would be expected if the effects of drought duration were compounding. Interannual variability in precipitation and ANPP is substantial throughout the Great Plains and Southwestern US (Knapp and Smith 2001; Maurer et al. 2020). During this experiment ANPP at each site varied 1.5- to twofold among years driven by 34–90% differences in growing season precipitation (Fig. 3, Table S2). Such variability can make identifying temporal trends difficult when drought responses are quantified relative to ambient conditions (Hoover et al. 2018), and can lead to multiple ecosystem responses to lower ambient precipitation based on drought magnitude, duration, and timing (Hoover and Rogers 2016).
Belowground responses
Overall, BNPP (0–20 cm) was more sensitive to drought than ANPP in the final year of the 4-year experiment (Fig. 4). This pattern was particularly striking in the C3 grassland (HPG) where ANPP was not affected by either drought treatment, but BNPP was reduced by 32%, albeit this was not a statistically significant decline. Similarly, Frank (2007) reported that ANPP in C3 grasslands in Yellowstone National Park was unresponsive to summer drought, while BNPP (0–20 cm) was reduced by 39%. Belowground, evidence that intense drought reduced productivity more than chronic drought was clear in the most mesic grassland (KNZ), consistent with the theory that mesic grasslands are more sensitive to prolonged dry periods than to reduced precipitation event size. Finally, there is some evidence that during drought, root systems allocate proportionally more biomass to deeper soils (i.e., Arndal et al. 2018). This was evident in the two semi-arid grasslands where shallow soils were likely driest during drought, but not in the more mesic sites (Fig. 4).
Conclusions
Grassland productivity responses to drought can vary significantly and we hypothesized that this variability could be attributed, in part, to key characteristics (event size, event number, duration of dry periods) and/or duration of drought. However, after imposing drought either by reductions in rainfall event size (chronic drought) or the complete exclusion of precipitation (intense drought) in four distinct grassland types, we have only limited support for this hypothesis. In general, there was no consistent impact on either ANPP or BNPP attributable to the different types of drought imposed or to the duration of the drought period. However, when there were differences in productivity responses to these two drought types, intense drought generally resulted in greater reductions in productivity relative to chronic drought, both above- and belowground. Further, in all but one instance, the more negative effect of intense than chronic drought occurred in the two most mesic grasslands, consistent with theory predicting that mesic systems are particularly sensitive to extended dry periods. Finally, drought caused consistently greater magnitude reductions in BNPP (statistically significant in 3 of 4 sites) than in ANPP, for both xeric and mesic grasslands. If the observed drought-induced reductions in root biomass persist in future years, this drought legacy provides a mechanism for the delayed recovery of grassland ANPP following extended drought (Sala et al. 2012).
References
Arndal MF, Tolver A, Larsen KS, Beier C, Schmidt IK (2018) Fine root growth and vertical distribution in response to elevated co2, warming and drought in a mixed heathland–grassland. Ecosystems 21:15–30. https://doi.org/10.1007/s10021-017-0131-2
Baez S, Collins SL, Pockman WT, Johnson JE, Small EE (2013) Effect of experimental rainfall manipulations on Chihuahuan Desert grassland and shrubland plant communities. Oecologia 172:1117–1127. https://doi.org/10.1007/2Fs00442-012-2552-0
Bates DM, Mäechler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Burke IC, Yonker CM, Parton WJ, Cole CV, Flach K, Schimel DS (1989) Texture, climate, and cultivation effects on soil organic matter content in US grassland soils. Soil Sci Soc Am J 53:800–805. https://doi.org/10.2136/sssaj1989.03615995005300030029x
Burke IC, Kittel TG, Lauenroth WK, Snook P, Yonker CM, Parton WJ (1991) Regional analysis of the central Great Plains. BioScience 41:685–692. https://doi.org/10.2307/1311763
Cherwin K, Knapp AK (2012) Unexpected patterns of sensitivity to drought in semi-arid grasslands. Oecologia 169:845–852. https://doi.org/10.1007/2Fs00442-011-2235-2
Denton EM, Dietrich JD, Smith MD, Knapp AK (2017) Drought timing differentially affects above- and belowground productivity in a mesic grassland. Plant Ecol 218:317–328. https://doi.org/10.1007/2Fs11258-016-0690-x
Dodd MB, Lauenroth WK (1997) The influence of soil texture on the soil water dynamics and vegetation structure of a shortgrass steppe ecosystem. Plant Ecol 133:13–28. https://doi.org/10.1023/A:1009759421640
Easterling DR, Meehl GA, Parmesan C, Changnon SA, Karl TR, Mearns LO (2000) Climate extremes: observations, modeling, and impacts. Science 289:2068–2074. https://doi.org/10.1126/science.289.5487.2068
Felton AJ, Slette IJ, Smith SM, Knapp AK (2019) Precipitation amount and event size interact to reduce ecosystem functioning during dry years in a mesic grassland. Glob Change Biol 26:658–668. https://doi.org/10.1111/gcb.14789
Frank DA (2007) Drought effects on above- and belowground production of a grazed temperate grassland ecosystem. Oecologia 152:131–139. https://doi.org/10.1007/s00442-006-0632-8
Gherardi LA, Sala OE (2018) Effect of interannual precipitation variability on dryland productivity: a global synthesis. Glob Change Biol 25:269–276. https://doi.org/10.1111/gcb.14480
Gong YH, Zhao DM, Ke WB, Fang C, Pei JY, Sun GJ, Ye JS (2020) Legacy effects of precipitation amount and frequency on the aboveground plant biomass of a semi-arid grassland. Sci Total Environ. https://doi.org/10.1016/j.scitotenv.2019.135899
Griffin-Nolan RJ, Carroll CJW, Denton EM, Johnston MK, Collins SL, Smith MD, Knapp AK (2018) Legacy effects of a regional drought on aboveground net primary production in six central US grasslands. Plant Ecol 219:505–515. https://doi.org/10.1007/s11258-018-0813-7
Griffin-Nolan RJ, Blumenthal DM, Collins SL, Farkas TE, Hoffman AM, Mueller KE, Ocheltree TW, Smith MD, Whitney KD, Knapp AK (2019) Shifts in plant functional composition following long-term drought in grasslands. J Ecol 107:2133–2148. https://doi.org/10.1111/1365-2745.13252
Heisler-White JL, Blair JM, Kelly EF, Harmoney K, Knapp AK (2009) Contingent productivity responses to more extreme rainfall regimes across a grassland biome. Glob Change Biol 15:2894–2904. https://doi.org/10.1111/j.1365-2486.2009.01961.x
Hoover DL, Rogers BM (2016) Not all droughts are created equal: the impacts of interannual drought pattern and magnitude on grassland carbon cycling. Glob Change Biol 22:1809–1920. https://doi.org/10.1111/gcb.13161
Hoover DL, Knapp AK, Smith MD (2014) Resistance and resilience of a grassland ecosystem to climate extremes. Ecology 95:2646–2656. https://doi.org/10.1890/13-2186.1
Hoover DL, Wilcox KR, Young KE (2018) Experimental droughts with rainout shelters: a methodological review. Ecosphere. https://doi.org/10.1002/ecs2.2088
Huxman TE, Smith MD, Fay PA, Knapp AK, Shaw MR, Loik ME, Smith SD, Tissue DT, Zak JC, Weltzin JF, Pockman WT, Sala OE, Haddad BM, Harte J, Koch GW, Schwinning S, Small EE, Williams DG (2004) Convergence across biomes to a common rain-use efficiency. Nature 429:651–654. https://doi.org/10.1038/nature02561
Jentsch A, Kreyling J, Elmer M, Gellesch E, Glaser B, Grant K, Hein R, Lara M, Mirzae H, Nadler SE, Nagy L, Otieno D, Pritsch K, Rascher U, Schädler M, Schloter M, Singh BK, Stadler J, Walter J, Wellstein C, Wöllecke J, Beierkuhnlein C (2011) Climate extremes initiate ecosystem-regulating functions while maintain productivity. J Ecol 99:689–702. https://doi.org/10.1111/j.1365-2745.2011.01817x
Knapp AK, Smith MD (2001) Variation among biomes in temporal dynamics of aboveground primary production. Science 291:481–484. https://doi.org/10.1126/science.291.5503.481
Knapp AK, Briggs JM, Koelliker JK (2001) Frequency and extent of water limitation to primary production in a mesic temperate grassland. Ecosystems 4:19–28. https://doi.org/10.1007/s100210000057
Knapp AK, Beier C, Briske DD, Classen AT, Luo Y, Reichstein M, Smith MD, Smith SD, Bell JE, Fay PA, Heisler JL, Leavitt SW, Sherry R, Smith B, Weng E (2008) Consequences of more extreme precipitation regimes for terrestrial ecosystems. Bioscience 58:811–821. https://doi.org/10.1641/B580908
Knapp AK, Hoover DL, Wilcox KR, Avolio ML, Koerner SE, La Pierre KJ, Loik ME, Luo Y, Sala OE, Smith MD (2015) Characterizing differences in precipitation regimes of extreme wet and dry years: implications for climate change experiments. Glob Change Biol 21:2624–2633. https://doi.org/10.1111/gcb.12888
Knapp AK, Chen A, Griffin-Nolan RJ, Baur LE, Carroll CJW, Gray JE, Hoffman AM, Li X, Post AK, Slette IJ, Collins SL, Luo Y, Smith MD (2020) Resolving the Dust Bowl paradox of grassland responses to extreme drought. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.1922030117
Kreyling J, Wenigmann M, Beierkuhnlein C, Jentsch A (2008) Effects of extreme weather events on plant productivity and tissue die-back are modified by community composition. Ecosystems 11:752–763. https://doi.org/10.1007/s10021-008-9157-9
Liu G, Liu H, Yin Y (2013) Global patterns of NDVI-indicated vegetation extremes and their sensitivity to climate extremes. Environ Res Lett. https://doi.org/10.1088/1748-9326/8/2/025009
Loik ME, Lesage JC, Brown TM, Hastings DO (2019) Drought-Net rainfall shelters did not cause nondrought effects on photosynthesis for California central coast plants. Ecohydrology 12:e2138. https://doi.org/10.1002/eco.2138
Luo W, Zuo X, Griffin-Nolan RJ, Xu C, Sardans J, Yu Q, Wang Z, Han X, Penuelas J (2020) Chronic and intense droughts differentially influence grassland carbon-nutrient dynamics along a natural aridity gradient. Plant Soil. https://doi.org/10.1007/2Fs11104-020-04571-8
Maurer GE, Hallmark AJ, Brown RF, Sala OE, Collins SL (2020) Sensitivity of primary production to precipitation across the United States. Ecol Lett 23:527–536. https://doi.org/10.1111/ele.13455
McCulley RL, Burke IC, Nelson JA, Lauenroth WK, Knapp AK, Kelly EF (2005) Regional patterns in carbon cycling across the Great Plains of North America. Ecosystems 8:106–121. https://doi.org/10.1007/s10021-004-0117-8
Monson RK, Williams GJ (1982) A correlation between photosynthetic temperature adaptation and seasonal phenology patterns in the shortgrass prairie. Oecologia 54:58–62. https://doi.org/10.1007/BF00541108
Monson RK, Stidham MA, Williams GJ, Edwards GE, Uribe EG (1982) Temperature dependence of photosynthesis in Agropyron smithii Rydb.: I. Factors affecting net CO2 uptake in intact leaves and contribution from Ribulose-1, 5-Bisphosphate carboxylase measured in vivo and in vitro. Plant Physiol 69:921–928. https://doi.org/10.1104/pp.69.4.921
Monson RK, Littlejohn RO, Williams GJ (1983) Photosynthetic adaptation to temperature in four species from the Colorado shortgrass steppe: a physiological model for coexistence. Oecologia 58:43–51. https://doi.org/10.1007/BF00384540
Monson RK, Sackschewsky MR, Williams GJ III (1986) Field measurements of photosynthesis, water-use efficiency, and growth in Agropyron smithii (C3) and Bouteloua gracilis (C4) in the Colorado shortgrass steppe. Oecologia 68:400–409. https://doi.org/10.1007/2FBF01036746
Morgan JA, LeCain DR, Pendall E, Blumenthal DM, Kimball BA, Carrillo Y, Williams DG, Heisler-White J, Dijkstra FA, West M (2011) C4 grasses prosper as carbon dioxide eliminates desiccation in warmed semi-arid grassland. Nature 476:202–205. https://doi.org/10.1038/nature10274
Nippert JB, Knapp AK (2007) Linking water uptake with rooting patterns in grassland species. Oecologia 153:261–272. https://doi.org/10.1007/2Fs00442-007-0745-8
Oesterheld M, Loreti J, Semmartin M, Sala OE (2001) Interannual variation in primary production of a semi-arid grassland related to previous-year production. J Veg Sci 12:137–142. https://doi.org/10.1111/j.1654-1103.2001.tb02624.x
Orth R, Destouni G, Jung M, Reichstein M (2020) Large-scale biospheric drought response intensifies linearly with drought duration in arid regions. Biogeosciences 17:2647–2656. https://doi.org/10.5194/bg-2019-442
Papalexiou SM, Montanari A (2019) Global and regional increase of precipitation extremes under global warming. Water Resour Res 55:4901–4914. https://doi.org/10.1029/2018WR024067
Raz-Yaseef N, Yakir D, Rotenberg E, Schiller G, Cohen S (2010) Ecohydrology of a semi-arid forest: partitioning among water balance components and its implications for predicted precipitation changes. Ecohydrology 3:143–154. https://doi.org/10.1002/eco.65
Raz-Yaseef N, Yakir D, Schiller G, Cohen S (2012) Dynamics of evapotranspiration partitioning in a semi-arid forest as affected by temporal rainfall patterns. Agric For Meteorol 157:77–85. https://doi.org/10.1016/j.agrformet.2012.01.015
Sala OE, Gherardi LA, Reichmann L, Jobbágy E, Peters D (2012) Legacies of precipitation fluctuations on primary production: theory and data synthesis. Philos Trans R Soc B 367:3135–3144. https://doi.org/10.1098/rstb.2011.0347
Seastedt TR, Knapp AK (1993) Consequences of non-equilibrium resource availability across multiple time scales: the transient maxima hypothesis. Am Nat 141:621–633. https://doi.org/10.1086/285494
Slette IJ, Awad M, Even T, Post AK, Punzalan A, Williams S, Smith MD, Knapp AK (2019) How ecologists define drought, and why we should do better. Glob Change Biol 25:3193–3200. https://doi.org/10.1111/gcb.14747
Taylor SH, Hulme SP, Rees M, Ripley BS, Woodward FI, Osborne CP (2010) Ecophysiological traits in C3 and C4 grasses: a phylogenetically controlled screening experiment. New Phytol 185:780–791. https://doi.org/10.1111/j.1469-8137.2009.03102.x
Thomey ML, Collins SL, Vargas R, Johnson JE, Brown RF, Natvig DO, Friggens MT (2011) Effect of precipitation variability on net primary production and soil respiration in a Chihuahuan Desert grassland. Glob Change Biol 17:1505–1515. https://doi.org/10.1111/j.1365-2486.2010.02363.x
Tielbörger K, Bilton MC, Metz J, Kigel J, Holzapfel C, Lebrija-Trejos E, Konsens I, Parag HA, Sternberg M (2014) Middle-Eastern plant communities tolerate 9 years of drought in a multi-site climate manipulation experiment. Nat Commun 5:5102. https://doi.org/10.1038/ncomms6102
Vicente-Serrano SM, Gouveia C, Camarero JJ, Begueria S, Trigo R, Lopez-Moreno JI, Azorin-Molina C, Pasho E, Lorenzo-Lacruz J, Revuelto J, Moran-Tejeda E, Sanchez-Lorenzo A (2013) Response of vegetation to drought time-scales across global land biomes. Proc Natl Acad Sci 110:52–57. https://doi.org/10.1073/pnas.1207068110
Wilcox KR, Von Fischer JC, Muscha JM, Petersen MK, Knapp AK (2015) Contrasting above- and belowground sensitivity of three Great Plains grasslands to altered rainfall regimes. Glob Change Biol 21:335–344. https://doi.org/10.1111/gcb.12673
World Meteorological Organization (1992) International meteorological vocabulary (2nd ed., p 784). Publication no. 182. Geneva, Switzerland: World Meteorological Organization (WMO).
Yahdjian L, Sala OE (2002) A rainout shelter design for intercepting different amounts of rainfall. Oecologia 133:95–101. https://doi.org/10.1007/2Fs00442-002-1024-3
Zeiter M, Scharrer S, Zweifel R, Newbery DM, Stampfili A (2016) Timing of extreme drought modifies reproductive output in semi-natural grassland. J Veg Sci 27:238–248. https://doi.org/10.1111/jvs.12362
Zobel Z, Wang J, Wuebbles DJ, Kotamarthi VR (2018) Analyses for high-resolution projections through the end of the 21st century for precipitation extremes over the United States. Earth’s Future 6:1471–1490. https://doi.org/10.1029/2018EF000956
Acknowledgments
This manuscript is to be published as part of a Special Issue honoring Russ Monson. We appreciate Russ' contributions to the broader ecological community through his long-standing editorial work with Oecologia as well as his foundational research advancing our understanding of C3 and C4 photosynthesis from an ecophysiological and evolutionary perspective. Research support was provided by the National Science Foundation: Macrosystems Biology/Emerging Frontiers Programs (EF‐1137342, EF‐1137378, EF-1137363) and the Long-Term Ecological Research program. Additionally, we want to thank Maddie Shields, Joshua O’Malley, John Dietrich, and Kevin Wilcox.
Author information
Authors and Affiliations
Contributions
CJWC and AKK wrote the initial draft, CJWC analyzed the data, AKK, MDS, IJS, and SLC edited the first draft, AKK, MDS, SLC, YL conceived the experiment, CJWC, IJS, RJG, LEB, AMH, EMD, JEG, AKP, MKJ, and QY established the experiment and collected data. All authors provided editorial advice.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Paul Stoy.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Carroll, C.J.W., Slette, I.J., Griffin-Nolan, R.J. et al. Is a drought a drought in grasslands? Productivity responses to different types of drought. Oecologia 197, 1017–1026 (2021). https://doi.org/10.1007/s00442-020-04793-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-020-04793-8