
Abstract
While both plant–soil feedbacks and allelochemical interactions are key drivers of plant community dynamics, the potential for these two drivers to interact with each other remains largely unexplored. If soil microbes influence allelochemical production, this would represent a novel dimension of heterogeneity in plant–soil feedbacks. To explore the linkage between soil microbial communities and plant chemistry, we experimentally generated soil microbial communities and evaluated their impact on leaf chemical composition and allelopathic potential. Four native perennial old-field species (two each of Aster and Solidago) were grown in pairwise combination with each species’ soil microbial community as well as a sterilized inoculum. We demonstrated unequivocally that variation in soil microbial communities altered leaf chemical fingerprints for all focal plant species and also changed their allelopathic potential. Soil microbes reduced allelopathic potential in bioassays by increasing germination 25–54% relative to sterile control soils in all four species. Plants grown with their own microbial communities had the lowest allelopathic potential, suggesting that allelochemical production may be lessened when growing with microbes from conspecifics. The allelopathic potential of plants grown in congener and confamilial soils was indistinguishable from each other, indicating an equivalent response to all non-conspecific microbial communities within these closely related genera. Our results clearly demonstrated that soil microbial communities cause changes in leaf tissue chemistry that altered their allelopathic properties. These findings represent a new mechanism of plant–soil feedbacks that may structure perennial plant communities over very small spatial scales that must be explored in much more detail.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
There is now compelling evidence that plant-soil microbe interactions can mediate plant performance to such an extent that they can shift the predominate form of competition between interspecific and intraspecific (Pendergast et al. 2013). Studies have repeatedly demonstrated that plants can generate soil microbial communities that are highly species specific, even among species that commonly co-occur in the field (Buyer et al. 2002; Kourtev et al. 2002; Reynolds et al. 2003; Pendergast et al. 2013). Indeed, there is now little doubt that soil microbial communities are a dominant force in structuring the composition and dynamics of plant communities (Bever 1994; Van Der Heijden et al. 2008; Bever et al. 2010; van der Putten et al. 2013).
However, the effects of rhizosphere interactions on the physiological and chemical properties of plants and its impacts on plant–plant interactions remain unclear and poorly explored. Leaf chemistry is a key factor in many plant-herbivore, and potentially, plant–plant interactions. If soil microbes alter leaf tissue chemistry this will likely cascade up to impact host choice and foraging behavior of herbivores, particularly phytophagous insects (Liu et al. 2007; Sikes 2010; Schittko and Wurst 2014; Kos et al. 2015a, b). Moreover, plant-soil microbe feedbacks that alter plant chemistry could also mediate plant–plant interactions via allelochemicals released into the soil (Rice 1974; Wardle et al. 1998; Inderjit et al. 2008, 2011). Because interactions with the soil microbial community can dramatically affect plant growth and chemistry (Bennett et al. 2009; Adesemoye et al. 2009; Jung et al. 2012; Vannette and Hunter 2013), we hypothesize that soil microbes may potentially change the strength of plant–plant allelopathic interactions. While the importance of soil microbes in mitigating the impacts of allelochemicals within the soil has received some attention (e.g., Lankau 2010; Cipollini et al. 2012; Shannon-Firestone and Firestone 2015), the potential for soil microbes to influence a plant’s production of allelochemicals has not been evaluated. We argue that this feedback may be common because of the ubiquity of plant–soil feedbacks and because soil microbes are known to alter plant defensive chemistry (e.g., Badri et al. 2013; Giron et al. 2013).
Though species often vary in their response to allelochemicals, allelopathy is typically treated as a binary plant characteristic rather than one that varies conditionally (Meiners et al. 2012). This all or nothing approach to allelochemical production lacks context dependency whereby ecological interactions and abiotic conditions (e.g., nutrient availability) alter the degree to which a given species is allelopathic. Nonetheless, a few studies have demonstrated that the strength of allelopathic interactions varies with nutrient availability, herbivores, and competitors (e.g., Kong et al. 2002, 2004; Rivoal et al. 2011; Ladwig et al. 2012). Studies evaluating the context dependency of allelopathy remain few and there appears to be a great potential for microbial communities to alter allelochemical production on a very local scale.
Allelopathy may function directly by inhibiting plant growth (Rice 1974; Kong et al. 2004; Thorpe et al. 2009; Greer et al. 2014), or indirectly through altering the abundance of soil mycorrhizal fungi or other microbial constituents (Roberts and Anderson 2005; Hale et al. 2011; Cipollini et al. 2012; Wang et al. 2012). Accordingly, we expect allelochemicals to both respond to and generate important changes in soil microbial communities. In addition, variation in the composition of soil microbial communities among plant species can respond to phylogenetic distance, with more closely related plant species generating more similar soil microbial communities and having more similar responses to soil microbes (Anacker et al. 2014; Burns et al. 2015). Consequently, we also expect phylogenetic distance between plant species to influence the impacts of their microbial communities on allelochemical production, with microbial communities from more closely related species producing similar effects on allelopathic potential.
To determine whether interactions with different soil microbial communities can generate strong conditionality in allelopathic interactions, we grew four old-field species from two genera in the Asteraceae in a suite of experimentally cultured microbial communities (Pendergast et al. 2013). This previous experiment demonstrated two central tenets of plant feedback theory: (1) plant species generate distinct soil microbial communities and (2) these differences in microbial communities alter plant performance sufficiently to change the nature of pairwise competitive interactions. Here, we used tissues from plants grown in distinct soil microbial communities to explore the potential for context dependency in allelopathy (Meiners et al. 2012). We used HPLC characterization of leaf metabolites followed by a series of allelopathy bioassays to test the following hypotheses: (1) Soil microbial communities will alter leaf chemistry, and this effect will vary with microbial community identity. (2) Soil microbial communities will influence the allelopathic potential of plants. (3) The degree of relatedness among plant species will influence the strength of their microbial communities’ impacts on allelopathic potential. This study represents a first attempt to integrate plant-soil microbial feedbacks into subsequent allelopathic interactions.
Materials and methods
Four native old-field herbs, Solidago canadensis, S. rugosa, Aster novae-angliae, and A. pilosus (synonymous with Symphyotrichum), were used in this experiment. The S. canadensis/altissima species complex dominates old-field communities for decades after abandonment throughout large portions of the northeastern and Midwestern United States and southern Canada (Werner et al. 1980; Banta et al. 2008; Pisula and Meiners 2010a). The other three focal species are less abundant, but commonly co-occur with S. canadensis throughout the same region (Myster and Pickett 1992; Banta et al. 2008). Solidago canadensis, is also invasive in Europe and Asia (Weber 1998; Abhilasha et al. 2008; Yuan et al. 2012), at least partly attributed to its allelopathic properties. The initial source of microbial communities and leaf tissue samples used in our experiments came from northwestern Pennsylvania, USA where Pendergast et al. (2013) evaluated how contrasting soil microbial communities caused differences in the performance of our focal old-field species (for details on inocula preparation and experiment see Pendergast et al. 2013).
Experimental generation of soil microbial communities
To culture the soil microbes associated with each species, greenhouse and field samples were used to generate a pooled inoculum. In the greenhouse, seedlings of each species were grown in 20 cm × 15 cm pots filled with a 3:1 ratio of homogenized soil from the upper 10 cm of an old-field near the Pymatuning Laboratory of Ecology (PLE) mixed with autoclaved silica sand with no supplemental fertilization. After one year, soil was separated from plant biomass. This generated a soil community trained by an individual plant species (e.g., Bever 1994; Hausmann and Hawkes 2009). Perhaps more importantly, because cultures produced under greenhouse conditions likely lack critical components of the soil microbial community (Sýkorová et al. 2007), we also used inocula produced under field conditions using soil samples from 5-year old monocultures of each focal species at PLE (Pendergast et al. 2013). Soil cores 20-cm deep were collected from these plots and stored at 4 °C until inoculum preparation (Bever 1994). Thus, our microbial communities likely represent a more complete complement of microbes than used in previous studies because they added a critical in situ component lacking in most previous studies.
Soil from both greenhouse and field monocultures were pooled at a 1:1 ratio to create a single inoculum for use in the experiment. The inoculum for each plant species was divided equally into two portions: one stored at 4 °C to use as live soil inoculum, and one autoclaved to use as sterile inoculum. Each live soil inoculum was composed of equal parts of each of the four soil communities, with all but one autoclaved. A sterile control composed of autoclaved soil from all four species was also generated. This inoculum strategy minimizes the biotic variation among inocula, including the pulse of nutrients that is released with autoclaving (Bever 1994). Genetic analyses of these inocula verified that there were strong differences in soil bacteria and fungal community composition (Pendergast et al. 2013).
Generation of plant material grown in association with different microbial communities
The goal of the original greenhouse experiment was to determine whether soil communities altered the performance of each focal species (Pendergast et al. 2013). To do this, 30 cm diameter pots were filled with a mixture of autoclaved field soil and silica sand in a 3:1 ratio. Pooled field and greenhouse soil inocula (described above) from one plant species were mixed into the top 5 cm of each pot in a 1:16 ratio of inoculum to sterile soil/sand. The abundance of autoclaved soil in both the inoculum and pot substrate should further minimize abiotic differences among treatments. Four similarly sized (with 4.08 ± 0.03 leaves), sterile-reared seedlings of a focal species were planted in each pot. Each of the four species were grown in all five soil microbial communities (sterile and from each of the four focal species) and replicated seven times in a greenhouse at PLE. After four months growth without supplemental fertilization, aboveground biomass was harvested and dried at 60 °C. The majority of plants were flowering at harvest (86–97% in each species), reducing the potential influence of developmental stage on plant chemistry (e.g., Filep et al. 2016). The exception was S. rugosa, where only 36% of plants had flowered. The results of this experiment showed that plant performance depended on microbial community identity, with the microbial community of S. canadensis producing the greatest reduction of growth in both itself and the other species (Pendergast et al. 2013). Here, we used leaf tissues from these plants to determine the effects of the soil microbial community on plant chemistry and allelopathic potential using a series of bioassays and HPLC analysis. While drying leaf tissues may result in the breakdown of some leaf chemicals and the loss of some volatile compounds, the consistency of treatment should ensure comparability across microbial communities. Tissues were stored dry until processing for leaf metabolites or allelopathic bioassays.
Chemical analyses
HPLC analysis was used to characterize leaf metabolites and determine whether soil microbial communities induced variation in the amount or type of chemicals produced. Metabolites were extracted using 1 mL of HPLC-grade methanol from 100 mg of leaf tissue that was ground to a fine powder with a pestle and liquid nitrogen. After centrifugation, the supernatant was filtered through a 0.22-µm filter and analyzed using a Hitachi Chromaster HPLC with a 5430 Diode Array detector. The mobile phase was a mixture of acetonitrile:water (v/v) at 20:80 from 0–5 min, a linear gradient of 20:80 to 95:5 from 5–45 min, 95:5 from 45–55 min, a linear gradient of 95:5 to 20:80 for 55–60 min, and 20:80 for 60–70 min. The flow rate was constant at 0.7 mL/min and the sample loading volume was 10 µL. This analytical approach would detect more polar items such as aldehydes simple unsaturated lactones in the first few minutes of the run, though the resolution may be less clear. Non-polar chemicals should have been better resolved than the polar chemicals and could include: aromatic acids, coumarins, quinones, flavonoids, tannins, alkaloids, terpenoids, phenolics and polyacetylenes. This range of detection covers the range of allelopathically active compounds within the Asteraceae (Chon and Nelson 2010; Kim and Lee 2011, Uesugi and Kessler 2013).
Chromatographic peaks that were discernable from the baseline (>75 µV s) were integrated and aligned visually. Peaks that occurred in five or fewer individuals within a species across all microbial communities were omitted. For each focal species, a principal components analysis was performed based on a correlation matrix using PC-Ord 6 (McCune and Grace 2002). Informative axes were determined by randomization tests, resulting in varying numbers of axes retained per species. PCA scores were then used in a one-way MANOVA comparing each inoculum treatment, followed by a series of univariate ANOVAS for each PCA axis to determine which axes were associated with microbial community differences. These results were also used to select axes for biplots to illustrate changes in chemistry associated with microbial community identity. Univariate analyses of Solidago rugosa only identified one significant PCA axis, so the first principal component was also included in the biplot, despite being non-significant.
Bioassays to determine microbial community effects on allelopathic potential
A standardized germination bioassay was employed to determine the effects of each soil microbial community on the allelopathic potential of each of the four focal plant species following the methods of Butcko and Jensen (2002). For each focal species, dried leaves from all replicates of the greenhouse experiment were pooled by species and soil community. Extracts were made from 12.5 g of dried leaf tissue in 500 mL of deionized water. This ratio of biomass to water generates plant extracts that affect germination of target species and allows for differentiation among allelopathic species (Butcko and Jensen 2002; Pisula and Meiners 2010b). Leaves were kept whole to prevent the release of compounds that may not be released under natural circumstances (Inderjit and Dakshini 1995). The mixture was placed on a magnetic stirrer for 24 h at room temperature and strained through cheesecloth to remove particulates. This coarse extract preparation may allow some compounds to break down, but is more representative of what would be naturally available than a solvent based extraction. Though root tissues may also be important local contributors of allelochemicals, we focus on leaves here for their ease of collection and their larger spatial range of potential impact in communities.
Dilutions of each extract, ranging from 0 to 100% in 10% increments, were made. Filter paper was placed in 90-mm Petri plates with 20 seeds of radish (Raphanus sativus L. ‘Early Scarlet Globe’; Bay Farm Services, Inc., Bay City, MI, USA), which served as the target species in all trials. We used this species to evaluate allelopathic potential because it germinates quickly, is commonly used in allelopathic studies, and is sensitive to allelopathic inhibition (Butcko and Jensen 2002; Pisula and Meiners 2010b). Previous work with Aster and Solidago has found germination rates too variable and slow for utility in such bioassays (Meiners, unpublished data). Five replicates were run at each dilution for each focal species/microbial community combination. Four mL of extract were added to each plate and incubated at 25 °C with a 12/12 h light/dark cycle. Petri plates were placed in plastic zip-lock bags to retain moisture during incubation. The plates were removed after four days and germinated seeds counted.
Logistic modeling was used to measure the overall effect of plant extract concentration (continuous) and soil community identity (categorical) on germination in each focal species (SAS 9.1;SAS Institute Inc., Cary, NC). This analysis was followed by individual logistic regressions of germination as a function of extract concentration for each species and each soil microbial community. Coefficients (β values) from these regressions were used to compare the relative strength of plant extracts from each soil community (Meiners 2014). Finally, all data were combined into a single logistic analysis to determine the influence of focal species relatedness on the impact of the microbial community. In this analysis, soil communities were categorized as self (same species), congener (different species, same genus), confamilial (both non-congeners) and sterile. Focal species identity was included to account for overall differences in allelopathic activity among plants.
The bioassay approach is valuable because it allows for the uniform testing of multiple plant species, which may differ dramatically in the allelochemicals produced. This technique has also proven highly valuable because it can simultaneously compare relative allelopathic potential among plant species (Pisula and Meiners 2010b; Meiners 2014) as well as detect differences within species in plants grown under different environmental conditions (Ladwig et al. 2012). A drawback of this approach is that it does not account for any microbial processing of plant extracts and interactions that might occur following senescence when leaf material comes in contact with the soil matrix (Inderjit and Dakshini 1995; Gibson 2002). Because our focus was on detecting within- and between-species variation, we chose the bioassay approach to quantify allelopathic potential and to isolate the effects of plant growing conditions.
Results
Soil microbial communities caused substantial and species-specific changes in leaf chemistry
HPLC analysis of leaf extracts resulted in chemical fingerprints that ranged from 49 to 91 separate metabolites (peaks) for each species. In support of our first hypothesis, inoculating experimentally generated microbial communities into the soil caused significant changes in the chemical fingerprints of leaves for all four species. PCA analyses of plant metabolite compositions resulted in four to six informative axes that explained 48–58% of the variation in chemical composition of the leaf extracts of each focal plant. Based on these PCA scores, our results demonstrate unequivocally that leaf metabolites differed significantly across soil communities for all four focal plant species (MANOVAs—Table 1; Fig. 1).

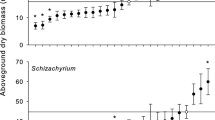
Effect of microbial community identity on the metabolite fingerprint of Aster and Solidago species. Data plotted are mean ± SE for two PCA axes selected for their ability to distinguish among soil communities (Table 1). Sample sizes are seven per species/soil combination with the exception ASPI in Self soil; SOCA in SORU and Sterile; and SORU in ASNE, SOCA and Sterile, which had six replicates. Only four replicates were available for SORU in self soil. Soil communities are abbreviated by the first two letters of the genus and species
While experimental manipulation of microbial communities caused tremendous variation in the leaf metabolite fingerprints of our focal species (Electronic supplementary material), the relationships among microbial communities also varied in a species-specific manner. With the exception of A. pilosus, sterile soils produced a leaf metabolite fingerprint similar to those produced in self soil. Aster pilosus leaf metabolites from self soil clustered with A. novae-angliae and S. canadensis, and were distinct from plants grown in sterile soils or with microbes from S. rugosa. Similarity of leaf metabolite fingerprints from self soil to other microbial communities also varied among species. For example, metabolite fingerprints of A. novae-angliae were similar in all soil communities except those from plants grown with S. canadensis microbes. In contrast, the metabolite composition of S. rugosa was similar for all heterospecific soil communities. Overall, each microbial community caused substantial changes in leaf metabolite composition and this varied strongly among our focal species (Fig. 1).
Changes in the metabolite fingerprints of leaves can come about in two ways, through changes in the relative abundances of chemicals or through changes in the identities of the chemicals produced. Similarity of metabolite composition across microbial inoculants for each focal plant species averaged 70.4% based on presence–absence data, but only 51.3% when relative abundances (area of each HPLC peak) of metabolites were included. This suggests that microbial-induced variation in metabolite composition was primarily driven by changes in the amount of each metabolite rather than the identities of metabolites produced.
Soil microbial communities influenced allelopathic potential
The degree to which plant extracts inhibited germination was entirely dependent upon the experimentally created soil microbial communities. Overall, plant extracts inhibited germination by as much as 80% (range 25–80%). As hypothesized, microbial community identity caused substantial variation in the allelopathic potential of focal species by generating large intraspecific variation in the degree that plant extracts inhibited target plant germination (Fig. 2; Table 2). Similar to the variation seen in leaf metabolite production, species varied greatly with respect to which microbial community produced the greatest allelopathic potential (Table 3). In A. novae-angliae and S. canadensis, sterile soils produced plants with the greatest allelopathic activity. In contrast, A. pilosus and S. rugosa had the greatest allelopathic potential when grown in microbial communities from A. novae-angliae. The lowest allelopathic potential occurred in a different microbial community for each focal plant species, demonstrating unequivocally that allelopathic potential is not only highly species specific, but also specific to the soil microbial community focal plants were treated with.

Bioassay results for Aster and Solidago species grown with differing soil microbial communities. Plots are model predictions from logistic regressions of Raphanus sativus germination as a function of extract concentration from each focal species. The sterile soil is represented by the solid line (as a reference) in all of the panels. Statistical analysis of these patterns presented in Table 2
The relationship of the soil community to the target plant mediated the strength of microbial feedbacks on allelopathy
In addition to the species- and inoculum-specific nature of the plant-microbial community interactions described above, more consistent patterns in allelopathic potential emerged when microbial communities were aggregated based on relatedness to the focal plant (Fig. 3). Because the focal species differed in their baseline allelopathic potentials, controlling for these differences was necessary in the pooled analysis (Table 4). After accounting for differences in focal species, there was significant variation in allelopathic potential associated with the focal plants relatedness to the plant that generated the soil microbial community. As an overall baseline, soil microbes reduced allelopathic potential because plants grown with sterilized soil inocula generally produced the greatest allelopathic activity relative to all live inocula. Within live soil inocula, plants grown with the most closely related microbial communities, that is conspecifics, had the lowest allelopathic potential. In contrast, both heterospecific soil microbial communities, congeners and confamilials, had greater allelopathic potentials that were intermediate between self and sterile soil inocula. However, there was no difference in allelopathic potential between plants inoculated with the more closely related congener microbial communities and those inoculated with the less related confamilials.

Effect of relatedness to the soil microbial community on the allelopathic potential expressed by Aster and Solidago species. Soil-species combinations are pooled into four groups—sterile soils, self soil, congener soil (same genus, other species), and confamilial (both species from the other genus). Plots are model predictions from logistic regressions of germination as a function of extract concentration (see Table 4 for analysis details)
Discussion
We found strong support for two of our hypotheses and limited support for a third. Specifically, experimentally cultured soil microbial communities caused substantial changes in leaf chemistry and the strength of this feedback varied strongly among coexisting plant species. Variation occurred to a lesser degree with the relationship of the plant to the soil microbial community. While soil microbes have been shown to affect leaf chemistry previously (Badri et al. 2013; Giron et al. 2013; Kos et al. 2015a; Schweiger and Müller 2015), our results demonstrate for the first time, to our knowledge, that different soil communities can influence allelopathic potential. Though we focused on a group of plants (Asteraceae) known to have high allelopathic activity (Chon and Nelson 2010; Kim and Lee 2011, Uesugi and Kessler 2013), we see no reason that microbial influences on allelopathic potential should not be a general phenomenon.
Our results suggest that there may be ongoing reciprocal feedbacks whereby soil microbial communities alter leaf chemistry, which upon plant inputs to soil (litter, root turnover, exudates) simultaneously alter microbial communities and plant–plant interactions at the scale of plant neighborhoods. While speculative, we suggest that such fine scale spatial dynamics occurring in the soil could create extremely patchy habitats where some species are favored over others, based upon these species-specific reciprocal feedbacks and their influence on competitive dynamics (Pendergast et al. 2013) and allelopathic interactions. If so, this suggests the potential for a fairly cryptic axis of niche differentiation at very small spatial scales premised upon species-specific plant–microbe interactions (Griffin et al. 2016). Regardless, these types of ongoing reciprocal feedbacks remain poorly explored in plant community ecology.
One of our key findings is that experimentally created variation in microbial communities caused substantive changes in leaf metabolite composition in all focal species. This confirms that local soil microbes can induce important chemical changes in leaves among long-lived and co-occurring plant species (see also Moore et al. 2003; Giron et al. 2013; Kos et al. 2015a). The changes in the leaves were sufficient to cause substantial differences in the degree that these leaves were allelopathic as determined by our bioassays. Microbes are known to change other aspects of plant phenotypes (Friesen et al. 2011; Griffin and Carson 2015; Hardoim et al. 2015) that may have contributed indirectly to the differences in allelopathy that we observed. Indeed, the composition of S. canadensis leaf metabolite fingerprints from plants grown in sterile and self soil were quite similar, yet the allelopathic potential of leaves from sterile inocula was much greater. This suggests that the microbes were altering plant chemistry in ways other than just those responsible for allelopathic interactions, but only further research that identifies the role of individual chemical constituents can parse this out.
Plants grown in the absence of a live microbial community had greater allelopathic potential, suggesting that interactions with soil microbes cause plants to reduce allelochemical production. Mechanistically, this reduced production could be the result of carbon allocation to defending against pathogens, reduced soil resource uptake, or an overall reduction in plant performance (e.g., Bennett et al. 2015). However, there were some cases (25% of all tested) where plants grown with specific soil communities had significantly greater allelopathic potential than the sterile controls. This occurred when A. pilosus was grown in soil from A. novae-angliae and S. rugosa was grown in soil with any microbial community other than its own. These instances appear to represent the induction of allelochemical production via a microbial interaction. Based on these data, soil microbes may be functioning as signals of the local competitive environment (Bais et al. 2004) that result in the alteration of allelopathic interactions.
Overall, the contrasting microbial communities created leaf metabolite fingerprints and allelopathic potentials that varied substantially depending upon plant species, whether they were grown with their own microbial community, a sterile community, or the microbial community cultured by a heterospecific. This means that microbe–plant interactions will likely occur at the scale of local plant neighborhoods, creating a potentially pervasive source of important heterogeneity in plant–plant interactions. Given the degree to which microbes can change leaf chemistry, this would likely cascade up to alter insect damage (Barbosa et al. 1991; Biere and Bennett 2013; Giron et al. 2013; Schittko and Wurst 2014), and cascade down to effect brown food webs (e.g., rates of decomposition, Inderjit et al. 2011).
There was limited support for the hypothesis that the relationship of the focal plant to the species that generated the microbial community should affect the strength of feedbacks. Leaf metabolite fingerprints were largely similar between sterile and self soils, suggesting that the presence of a species’ own microbial community may not induce changes in leaf metabolites, representing some level of self-recognition (Kigathi et al. 2013) relative to the microbial communities of other species. In contrast, all heterospecific combinations generated statistically equivalent allelopathic potentials in pooled data, so variation beyond self soil was not related to the strength of feedback in this system. Though the soil microbial communities were overall distinct among all focal species (Pendergast et al. 2013), the initial bacterial and AM fungal communities were more similar between Solidago species than the Aster species. More similar microbial communities did not generate similarity in the allelopathy bioassays, suggesting that the chemical responses may be to specific microbial constituents rather than overall microbial community similarity. Within the Asteraceae, Aster and Solidago are closely related. While this allowed us to focus on commonly co-occurring plants known to be allelopathic with likely similar chemical fingerprints, it represents a narrow phylogenetic distance to fully evaluate how relatedness mitigates microbial feedbacks on allelopathy. As plant-soil feedbacks may have a strong phylogenetic signal across a broader range of species (Anacker et al. 2014), studies that include multiple families may reveal a much stronger phylogenetic signal in feedbacks on allelopathy.
The patterns documented here are based on relatively simplistic germination bioassays and analyses of chemical constituents of a suite of plant species whose allelochemicals have not been well characterized (Chon and Nelson 2010). The usage of R. sativus as a model target species in the germination bioassays allowed separation of plants grown under different microbial conditions, but lacks an appropriate ecological context to allow full translation to a field setting. Plant species often vary dramatically in their response to allelochemicals, so we would expect variation in species’ responses in natural communities that must be addressed in future work. Similarly, we were not able to quantify individual chemical responses to the soil microbial context and their contribution to allelopathic impacts. Regardless, we think that the analyses presented argue strongly for detailed analyses of better characterized allelopathic model plants under ecologically realistic settings. This approach will be necessary to fully assess the ecological importance of microbial influence on allelopathic interactions.
The microbially influenced allelopathic interactions that we documented represent another mechanism by which plant–microbe interactions can shape plant community structure. Specificity in plant–microbe interactions appears common (Klironomos 2002; Kardol et al. 2006; Busby et al. 2011; Pendergast et al. 2013), suggesting that microbial mediation of allelochemicals will likely be common and also species and context dependent. While conditionality makes the importance of allelopathy in plant communities difficult to judge, it is consistent with the complexity and context dependence exhibited by many ecological interactions (Meiners et al. 2012). Therefore, it should not be surprising to find similar processes operating within allelopathic interactions. While it will be difficult to separate microbially mediated competitive effects from microbially influenced changes in allelopathic effects (Inderjit and del Moral 1997), our results suggest that this may be an important pathway in understanding plant–plant interactions that should be more fully explored. The context dependency of plant–soil interactions (Kardol et al. 2013), and their potential to cascade throughout systems represents a hidden, and potentially critical source of heterogeneity in species interactions.
References
Abhilasha D, Quintana N, Vivanco J, Joshi J (2008) Do allelopathic compounds in invasive Solidago canadensis s.l. restrain the native European flora? J Ecol 96:993–1001
Adesemoye AO, Torbert HA, Kloepper JW (2009) Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb Ecol 58:921–929. doi:10.1007/s00248-009-9531-y
Anacker BL, Klironomos JN, Maherali H, Reinhart KO, Strauss SY (2014) Phylogenetic conservatism in plant-soil feedback and its implications for plant abundance. Ecol Lett 17:1613–1621. doi:10.1111/ele.12378
Badri DV, Zolla G, Bakker MG, Manter DK, Vivanco JM (2013) Potential impact of soil microbiomes on the leaf metabolome and on herbivore feeding behavior. New Phytol 198:264–273. doi:10.1111/nph.12124
Bais HP, Park S-W, Weir TL, Callaway RM, Vivanco JM (2004) How plants communicate using the underground information superhighway. Trends Plant Sci 9:26–32. doi:10.1016/j.tplants.2003.11.008
Banta J, Stark S, Stevens M, Pendergast T, Baumert A, Carson W (2008) Light reduction predicts widespread patterns of dominance between asters and goldenrods. Plant Ecol 199:65–76. doi:10.1007/s11258-008-9412-3
Barbosa P, Krischik VA, Jones CG (eds) (1991) Microbial mediation of plant-herbivore interactions. Wiley, New York
Bennett AE, Bever JD, Deane Bowers M (2009) Arbuscular mycorrhizal fungal species suppress inducible plant responses and alter defensive strategies following herbivory. Oecologia 160:771–779. doi:10.1007/s00442-009-1338-5
Bennett AE, Grussu D, Kam J, Caul S, Halpin C (2015) Plant lignin content altered by soil microbial community. New Phytol 206:166–174. doi:10.1111/nph.13171
Bever JD (1994) Feeback between plants and their soil communities in an old field community. Ecology 75:1965–1977. doi:10.2307/1941601
Bever JD, Dickie IA, Facelli E, Facelli JM, Klironomos J, Moora M, Rillig MC, Stock WD, Tibbett M, Zobel M (2010) Rooting theories of plant community ecology in microbial interactions. Trends Ecol Evol 25:468–478. doi:10.1016/j.tree.2010.05.004
Biere A, Bennett AE (2013) Three-way interactions between plants, microbes and insects. Funct Ecol 27:567–573. doi:10.1111/1365-2435.12100
Burns JH, Anacker BL, Strauss SY, Burke DJ (2015) Soil microbial community variation correlates most strongly with plant species identity, followed by soil chemistry, spatial location and plant genus. AoB Plants 7. doi:10.1093/aobpla/plv030
Busby RR, Gebhart DL, Stromberger ME, Meiman PJ, Paschke MW (2011) Early seral plant species’ interactions with an arbuscular mycorrhizal fungi community are highly variable. Appl Soil Ecol 48:257–262. doi:10.1016/j.apsoil.2011.04.014
Butcko VM, Jensen RJ (2002) Evidence of tissue-specific allelopathic activity in Euthamia graminifolia and Solidago canadensis (Asteraceae). Am Midl Nat 148:253–262. doi:10.1674/0003-0031(2002)148[0253:eotsaa]2.0.co;2
Buyer JS, Roberts DP, Russek-Cohen E (2002) Soil and plant effects on microbial community structure. Can J Micro 48:955–964. doi:10.1139/w02-095
Chon SU, Nelson CJ (2010) Allelopathy in Compositae plants: a review. Agron Sustain Dev 30:349–358
Cipollini D, Rigsby CM, Barto EK (2012) Microbes as targets and mediators of allelopathy in plants. J Chem Ecol 38:714–727. doi:10.1007/s10886-012-0133-7
Filep R, Pal RW, Balázs VL et al (2016) Can seasonal dynamics of allelochemicals play a role in plant invasions? A case study with Helianthus tuberosus L. Plant Ecol 217:1489–1501. doi:10.1007/s11258-016-0662-1
Friesen ML, Porter SS, Stark SC, von Wettberg EJ, Sachs JL, Martinez-Romero E (2011) Microbially mediated plant functional traits. Ann Rev Ecol Evol Syst 42:23–46. doi:10.1146/annurev-ecolsys-102710-145039
Gibson DJ (2002) Methods in comparative plant population ecology. Oxford University Press, Oxford
Giron D, Frago E, Glevarec G, Pieterse CMJ, Dicke M (2013) Cytokinins as key regulators in plant–microbe–insect interactions: connecting plant growth and defence. Funct Ecol 27:599–609. doi:10.1111/1365-2435.12042
Greer M, Wilson GT, Hickman K, Wilson S (2014) Experimental evidence that invasive grasses use allelopathic biochemicals as a potential mechanism for invasion: chemical warfare in nature. Plant Soil 385:165–179. doi:10.1007/s11104-014-2209-3
Griffin EA, Carson WP (2015) The ecology and natural history of foliar bacteria with a focus on tropical forests and agroecosystems. Bot Rev 81:105–149. doi:10.1007/s12229-015-9151-9
Griffin EA, Traw MB, Morin PJ, Pruitt JN, Wright SJ, Carson WP (2016) Foliar bacteria and soil fertility mediate seedling performance: a new and cryptic dimension of niche differentiation. Ecology 97:2998–3008. doi:10.1002/ecy.1537
Hale AN, Tonsor SJ, Kalisz S (2011) Testing the mutualism disruption hypothesis: physiological mechanisms for invasion of intact perennial plant communities. Ecosphere 2:12
Hardoim PR, van Overbeek LS, Berg G, Pirttilä AM, Compant S, Campisano A, Döring M, Sessitsch A (2015) The hidden world within plants: ecological and evolutionary considerations for defining functioning of microbial endophytes. Micro Molec Biol Rev 79:293–320. doi:10.1128/mmbr.00050-14
Hausmann NT, Hawkes CV (2009) Plant neighborhood control of arbuscular mycorrhizal community composition. New Phytol 183:1188–1200. doi:10.1111/j.1469-8137.2009.02882.x
Inderjit, Dakshini KMM (1995) On laboratory bioassays in allelopathy. Bot Rev 61:28–44. doi:10.1007/bf02897150
Inderjit, del Moral R (1997) Is separating resource competition from allelopathy realistic? Bot Rev 63:221–230. doi:10.2307/4354300
Inderjit, Seastedt T, Callaway R, Pollock J, Kaur J (2008) Allelopathy and plant invasions: traditional, congeneric, and bio-geographical approaches. Bio Invas 10:875–890. doi:10.1007/s10530-008-9239-9
Inderjit, Wardle DA, Karban R, Callaway RM (2011) The ecosystem and evolutionary contexts of allelopathy. Trends Ecol Evol 26:655–662
Jung SC, Martinez-Medina A, Lopez-Raez JA, Pozo MJ (2012) Mycorrhiza-induced resistance and priming of plant defenses. J Chem Ecol 38:651–664. doi:10.1007/s10886-012-0134-6
Kardol P, Martijn Bezemer T, van der Putten WH (2006) Temporal variation in plant–soil feedback controls succession. Ecol Lett 9:1080–1088. doi:10.1111/j.1461-0248.2006.00953.x
Kardol P, De Deyn GB, Laliberté E, Mariotte P, Hawkes CV (2013) Biotic plant–soil feedbacks across temporal scales. J Ecol 101:309–315. doi:10.1111/1365-2745.12046
Kigathi RN, Weisser WW, Veit D, Gershenzon J, Unsicker SB (2013) Plants suppress their emission of volatiles when growing with conspecifics. J Chem Ecol 39:537–545. doi:10.1007/s10886-013-0275-2
Kim Y, Lee E (2011) Comparison of phenolic compounds and the effects of invasive and native species in East Asia: support for the novel weapons hypothesis. Ecol Res 26:87–94. doi:10.1007/s11284-010-0762-7
Klironomos JN (2002) Feedback with soil biota contributes to plant rarity and invasiveness in communities. Nature 417:67–70
Kong CH, Hu F, Xu XH (2002) Allelopathic potential and chemical constituents of volatiles from Ageratum conyzoides under stress. J Chem Ecol 28:1173–1182. doi:10.1023/a:1016229616845
Kong CH, Hu F, Liang WJ, Wang PW, Jiang Y (2004) Allelopathic potential of Ageratum conyzoides at various growth stages in different habitats. Allelopath J 13:233–240
Kos M, Tuijl MAB, de Roo J, Mulder PPJ, Bezemer TM (2015a) Plant–soil feedback effects on plant quality and performance of an aboveground herbivore interact with fertilisation. Oikos 124:658–667. doi:10.1111/oik.01828
Kos M, Tuijl MAB, de Roo J, Mulder PPJ, Bezemer TM (2015b) Species-specific plant–soil feedback effects on above-ground plant–insect interactions. J Ecol 103:904–914. doi:10.1111/1365-2745.12402
Kourtev PS, Ehrenfeld JG, Häggblom M (2002) Exotic plant species alter the microbial community structure and function in the soil. Ecology 83:3152–3166
Ladwig LM, Meiners SJ, Pisula NL, Lang KA (2012) Conditional allelopathic potential of temperate lianas. Plant Ecol 213:1927–1935. doi:10.1007/s11258-012-0087-4
Lankau R (2010) Soil microbial communities alter allelopathic competition between Alliaria petiolata and a native species. Bio Invas 12:2059–2068. doi:10.1007/s10530-009-9608-z
Liu J, Maldonado-Mendoza I, Lopez-Meyer M, Cheung F, Town CD, Harrison MJ (2007) Arbuscular mycorrhizal symbiosis is accompanied by local and systemic alterations in gene expression and an increase in disease resistance in the shoots. Plant J 50:529–544. doi:10.1111/j.1365-313X.2007.03069.x
McCune B, Grace JB (2002) Analysis of ecological communities. MjM Software Design, Gleneden Beach
Meiners SJ (2014) Functional correlates of allelopathic potential in a successional plant community. Plant Ecol 215:661–672. doi:10.1007/s11258-014-0331-1
Meiners SJ, Kong C-H, Ladwig LM, Pisula NL, Lang KA (2012) Developing an ecological context for allelopathy. Plant Ecol 213:1221–1227. doi:10.1007/s11258-012-0078-5
Moore JC, McCann K, Setälä H, De Ruiter PC (2003) Top-down is bottom-up: does predation in the rhizosphere regulate aboveground dynamics? Ecology 84:846–857. doi:10.1890/0012-9658(2003)084[0846:tibdpi]2.0.co;2
Myster RW, Pickett STA (1992) Dynamics of associations between plants in ten old fields during 31 years of succession. J Ecol 80:291–302
Pendergast TH, Burke DJ, Carson WP (2013) Belowground biotic complexity drives aboveground dynamics: a test of the soil community feedback model. New Phytol 197:1300–1310. doi:10.1111/nph.12105
Pisula NL, Meiners SJ (2010a) Allelopathic effects of goldenrod species on turnover in successional communities. Am Midl Nat 163:161–172
Pisula NL, Meiners SJ (2010b) Relative allelopathic potential of invasive plant species in a young disturbed woodland. J Torrey Bot Soc 137:81–87
Reynolds HL, Packer A, Bever JD, Clay K (2003) Grassroots ecology: plant–microbe–soil interactions as drivers of plant community structure and dynamics. Ecology 84:2281–2291
Rice EL (1974) Allelopathy. Academic Press, New York
Rivoal A, Fernandez C, Greff S, Montes N, Vila B (2011) Does competition stress decrease allelopathic potential? Biochem Syst Ecol 39:401–407
Roberts KJ, Anderson RC (2005) Effect of garlic mustard [Alliaria petiolata (Beib, Cavarra & Grande)] extracts on plants and arbuscular mycorrhizal (AM) fungi. Am Midl Nat 146:146–152
Schittko C, Wurst S (2014) Above- and belowground effects of plant-soil feedback from exotic Solidago canadensis on native Tanacetum vulgare. Bio Invasion 16:1465–1479. doi:10.1007/s10530-013-0584-y
Schweiger R, Müller C (2015) Leaf metabolome in arbuscular mycorrhizal symbiosis. Curr Opin Plant Biol 26:120–126. doi:10.1016/j.pbi.2015.06.009
Shannon-Firestone S, Firestone J (2015) Allelopathic potential of invasive species is determined by plant and soil community context. Plant Ecol 216:491–502. doi:10.1007/s11258-015-0453-0
Sikes BA (2010) When do arbuscular mycorrhizal fungi protect plant roots from pathogens? Plant Signal Behav 5:763–765. doi:10.4161/psb.5.6.11776
Sýkorová Z, Ineichen K, Wiemken A, Redecker D (2007) The cultivation bias: different communities of arbuscular mycorrhizal fungi detected in roots from the field, from bait plants transplanted to the field, and from a greenhouse trap experiment. Mycorrhiza 18:1–14. doi:10.1007/s00572-007-0147-0
Thorpe AS, Thelen GC, Diaconu A, Callaway RM (2009) Root exudate is allelopathic in invaded community but not in native community: field evidence for the novel weapons hypothesis. J Ecol 97:641–645
Uesugi A, Kessler A (2013) Herbivore exclusion drives the evolution of plant competitiveness via increased allelopathy. New Phytol 198:916–924. doi:10.1111/nph.12172
Van Der Heijden MGA, Bardgett RD, Van Straalen NM (2008) The unseen majority: soil microbes as drivers of plant diversity and productivity in terrestrial ecosystems. Ecol Lett 11:296–310. doi:10.1111/j.1461-0248.2007.01139.x
van der Putten WH, Bardgett RD, Bever JD, Bezemer TM, Casper BB, Fukami T, Kardol P, Klironomos JN, Kulmatiski A, Schweitzer JA, Suding KN, Van de Voorde TFJ, Wardle DA (2013) Plant–soil feedbacks: the past, the present and future challenges. J Ecol 101:265–276. doi:10.1111/1365-2745.12054
Vannette RL, Hunter MD (2013) Mycorrhizal abundance affects the expression of plant resistance traits and herbivore performance. J Ecol 101:1019–1029. doi:10.1111/1365-2745.12111
Wang J, Li X, Zhang J, Yao T, Wei D, Wang Y, Wang J (2012) Effect of root exudates on beneficial microorganisms—evidence from a continuous soybean monoculture. Plant Ecol 213:1883–1892. doi:10.1007/s11258-012-0088-3
Wardle DA, Nilsson M-C, Gallet C, Zackrisson O (1998) An ecosystem-level perspective of allelopathy. Biol Rev 73:305–319
Weber E (1998) The dynamics of plant invasions: a case study of three exotic goldenrod species (Solidago L.) in Europe. J Biogeogr 25:147–154. doi:10.1046/j.1365-2699.1998.251119.x
Werner PA, Gross RS, Bradbury IK (1980) The biology of Canadian weeds.: 45. Solidago canadensis L. Can J Plant Sci 60:1393–1409. doi:10.4141/cjps80-194
Yuan Y, Wang B, Zhang S, Tang J, Tu C, Hu S, Yong JWH, Chen X (2012) Enhanced allelopathy and competitive ability of invasive plant Solidago canadensis in its introduced range. J Plant Ecol. doi:10.1093/jpe/rts033
Acknowledgements
We thank Grace S. Lloyd, Siarhei Tsymbalau and Garland P. Waleko for their assistance in the field and greenhouse, Anna J. Herzberger for assistance in the lab, and Sharon N. Dubosky for comments on the manuscript. This work was funded by the G. Murray McKinley Research Fund of the Pittsburgh Foundation, a NSF Doctoral Dissertation Improvement Grant (#0508012), and an Undergraduate Research Grant from Eastern Illinois University
Author contribution statement
TP and WC designed and conducted the initial project, KP and SM conducted the allelopathy work, TC conducted the HPLC analysis and all authors contributed to the writing and editing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Maria J. Pozo.
Data available from the Dryad Digital Repository: http://dx.doi.org/10.5061/dryad.3418j.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Meiners, S.J., Phipps, K.K., Pendergast, T.H. et al. Soil microbial communities alter leaf chemistry and influence allelopathic potential among coexisting plant species. Oecologia 183, 1155–1165 (2017). https://doi.org/10.1007/s00442-017-3833-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-017-3833-4