
Abstract
It is widely accepted that predator recognition and avoidance are important behaviors in allowing prey to mitigate the impacts of their predators. However, while prey species generally develop anti-predator behaviors through coevolution with predators, they sometimes show accelerated adoption of these behaviors under strong selection pressure from novel species. We used a field manipulation experiment to gauge the ability of the common ringtail possum (Pseudocheirus peregrinus), a semi-arboreal Australian marsupial, to recognize and respond to olfactory cues of different predator archetypes. We predicted that ringtails would display stronger anti-predator behaviors to cues of the invasive European red fox (Vulpes vulpes) in areas where fox impacts had been greatest, and to cues of the native lace monitor (Varanus varius) in areas of sympatry compared with allopatry. We found that ringtails fled quickly and were more alert when exposed to the fecal odors of both predators compared to neutral and pungent control odors, confirming that predator odors are recognized and avoided. However, these aversive responses were similar irrespective of predator presence or level of impact. These results suggest that selection pressure from the fox has been sufficient for ringtails to develop anti-predator behaviors over the few generations since foxes have become established. In contrast, we speculate that aversive responses by ringtails to the lace monitor in areas where this predator is absent reflect recent coexistence of the two species. We conclude that rapid evolution of anti-predator behaviors may occur when selection is strong. The maintenance of these behaviors should allow re-establishment of predator–prey relationships if the interactants regain sympatry via range shifts or management actions to reintroduce them to their former ranges.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The adoption of anti-predator behavior is a key survival strategy in prey populations, often providing prey with strong mitigation of predator impacts (Sih and Christensen 2001). Anti-predator behaviors include increased vigilance, escape and avoidance responses (Kats and Dill 1998; Apfelbach et al. 2005; Martin et al. 2010), as well as altered habitat use and avoidance of preferred feeding patches (Lima 1998; Hayes et al. 2006). They may develop over ecological or evolutionary timeframes depending on the strength of selection that is imposed (Strauss et al. 2006; Kovacs et al. 2012). However, anti-predator behavior is costly in terms of time and access to resources and may be maintained only if it provides net fitness benefits. Identifying the conditions that allow retention of anti-predator behaviors is important for understanding the selective milieu of prey and crucial also in applied conservation programs. For example, managers increasingly are introducing predators back into their historical ranges to restore natural systems (Miller et al. 1999; Terborgh et al. 2001; Devineau et al. 2010) or control invasive species (Kinnear et al. 1998; Olsson et al. 2005), making it essential to forecast how native prey will respond to their reappearance (Griffin et al. 2000).
Some anti-predator behaviors are heritable, but others are more phenotypically plastic (Blumstein and Daniel 2005). Following the removal of a predator, relaxation of selection can lead to the rapid loss of costly behavioral traits from a prey population (Endler and Houde 1995); in others, however, specific anti-predator responses may be conserved, with some traits persisting long after isolation (Coss and Ramakrishnan 2000; Blumstein 2006; Hollen and Manser 2007). In moose (Alces alces), for example, behavioral adjustments to the presence of wolves (Canis lupus) occurred within one generation following the re-establishment of encounters with the recolonizing predator (Berger et al. 2001). Likewise, yellow-bellied marmots (Marmota flaviventris) retain the ability to discriminate between extirpated predators and associated risks in populations exposed to multiple predators (Blumstein et al. 2009). The ‘multipredator hypothesis’ suggests that species may retain behavioral traits towards predators they no longer coexist with if subject to persistent pressure from other predator types (Blumstein et al. 2004; Blumstein 2006).
These observations suggest that prey which accurately judge predation risk and calibrate their responses accordingly should maximize the benefit-to-cost ratio of their behavior and increase their fitness. For prey that are active by day and detect predators visually, judging both immediate danger and general levels of predation risk may be relatively simple. For example, thirteen-lined ground squirrels (Spermophilus tridecemlineatus) spend more time in alert postures if their long-range view is occluded than if unrestrained, allowing them to markedly reduce their reaction times when exposed to birds of prey (Arenz and Leger 1997). For prey that are active by night or that occupy structurally complex habitats, by contrast, visual assessment of predation risk may not be reliable. Some nocturnal prey species use auditory cues to detect the immediate presence of a predator (e.g. many flying insects; Triblehorn and Yager 2005), but many use the more remote but enduring cues provided by predator odor (Kats and Dill 1998). For example, many mammalian predators deposit urinary and fecal odors throughout their home ranges and on range boundaries (Macdonald 1980), thus providing cues to prey animals about their presence, identity and potential risk of encounter (Muller-Schwarze 2006).
Although odor cues such as feces and urine provide ostensibly reliable information about predation risk, the responses of prey can be very variable (Kats and Dill 1998). Weak responses may arise if the odors are old and prey animals perceive the risk of encounter with the predator to be low (Dickman and Doncaster 1984), if the prey are young or otherwise inexperienced (Kovacs et al. 2012), if the predator has weak selective effects on the prey (Sih et al. 2010), if the predator is novel and thus not recognized by the prey (Banks and Dickman 2007) or, in experimental studies, if predator odors are presented out of context. In some studies of small mammalian prey, for example, predator odors that are presented on metal traps may conflate the effects of the odors with effects of the novel objects (Dickman and Doncaster 1984; Apfelbach et al. 2005). In contrast, strongly aversive prey responses have been reported towards the odors of both familiar and unfamiliar predators, perhaps reflecting the presence of chemical cues that occur commonly in predator wastes (Stoddart 1982; Fendt et al. 2005).
In the study reported here, our broad objective was to investigate the response of an Australian marsupial, the common ringtail possum (Pseudocheirus peregrinus), hereafter referred to as ‘ringtail’, to the odors of two disparate predators, namely, the introduced European red fox (Vulpes vulpes) and the native lace monitor (Varanus varius). The direct consumptive effects of introduced predators on prey are generally greater in Australia than in other parts of the world (Salo et al. 2007, 2010), perhaps because Australian native prey species are naïve to the predator archetypes represented by the newcomers or because their anti-predator behaviors are less effective against the invaders than against native species (Banks and Dickman 2007). Several naïve local species have been extirpated in Australia as an apparent consequence of the introduction of novel predators (Saunders et al. 2010), presumably because the timeframe was too short for effective anti-predator responses to evolve. For remaining prey species, however, predator avoidance appears to be variable: some species show marked discrimination of predator odors (Gresser 1996; Mella et al. 2010), while others show no recognition or recognition at some times and places only (Banks 1998; Banks et al. 2003; Russell and Banks 2007; McEvoy et al. 2008).
The anti-predator behavior of the ringtail has been little studied. However, it is preyed upon by both the red fox and lace monitor (Triggs et al. 1984; Jessop et al. 2010), and its use of dreys—nest-like constructions of leaves and twigs—for shelter has been interpreted as both a warning to and escape strategy against arboreal predators, such as the monitor (Russell et al. 2003; Lindenmayer et al. 2008). As ringtail occupancy and density are unaffected by fox presence over large areas (J. Anson, unpublished data), it has clearly survived the initial impact of fox arrival and has probably been subject to strong subsequent selection to distinguish and avoid this predator (Kovacs et al. 2012). The ringtail now persists with both predators over most of its range on the Australian mainland, but it occurs in the absence of the monitor over much of western Victoria. Fecal odors likely provide useful cues to the presence and predation risk posed by both predators because both predators defecate throughout their ranges rather than at discrete latrines (Macdonald 1980; King and Green 1999), while the ringtail has well-developed olfactory lobes and uses olfaction to distinguish among individuals (Walker and Croft 1990).
In the light of these observations, our specific aims are to test three hypotheses. We predict that the common ringtail possum will show:
-
1.
generally aversive responses to the fecal odors of the red fox and the lace monitor, but not to other pungent odors or to neutral odors;
-
2.
increasing aversion to the odor of the fox in localities of increasing fox impact;
-
3.
aversion to the odor of the lace monitor in localities where the two species co-occur, but little or no aversion where they are allopatric.
We assumed that fox impact—and selection pressure—on the ringtail would be related to the length of time the predator had occurred in a locality and to its density while present.
Methods
Study species
The common ringtail is a small (700–900 g) folivorous possum that feeds largely on eucalyptus leaves. The species is primarily arboreal, but animals are often active on or near the forest floor and occupy small home ranges covering 0.02–0.05 ha (Smith et al. 2003; Van Dyck and Strahan 2008). The lace monitor is a carnivorous tree goanna that hunts by day in both the terrestrial and arboreal environments (Weavers 1989). Weighing on average 7 kg, but reaching up to 14 kg in an altered habitat with human food subsidies, this species is the second largest carnivore in eastern Australia (Weavers 1988; Guarino 2001). The ringtail is a key component of its diet, representing over 60 % by volume of ingested prey in some areas (Jessop et al. 2010); monitors can remove nesting ringtails directly from tree hollows and dreys. The red fox (Vulpes vulpes) was first introduced to southern Victoria for hunting purposes in 1845, but did not become established in the state more broadly until about 1880 (Abbott 2011). The fox has therefore co-existed with ringtails in some parts of the latter species’ range for approximately 130 years, or >60 ringtail generations. It achieved formal pest status in Victoria in 1894 and is now a serious agricultural pest over most southern parts of the continent (Saunders et al. 2010). This novel predator has broadly negative impacts on native vertebrates via direct predation and, probably, competition (Banks et al. 2000; Short et al. 2002; Glen and Dickman 2005, 2008; Olsson et al. 2005; Glen et al. 2009; Ritchie and Johnson 2009), with the ringtail forming a large part of its diet in some areas (Triggs et al. 1984). Both predators are mobile and occupy ranges that potentially overlap those of many ringtails.
Study sites
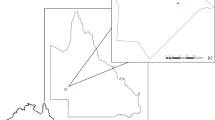
We chose three coastal study sites in south-eastern Australia with populations of common ringtail possums: Cape Conran (East Gippsland), Point Lonsdale (Bellarine Peninsula) and Phillip Island (Western Port) (Fig. 1). Sites were chosen to represent the different geographical ranges of the two predators and allow testing of the three hypotheses. These sites also have very few or no native mammalian predators; the spotted-tailed quoll (Dasyurus maculatus) may occur in small numbers at Cape Conran but not at the other two sites.

Map of western Victoria, Australia, showing study sites at Cape Conran Coastal Park, Point Lonsdale, and Phillip Island. Inset Location of the study region in Australia
Cape Conran Coastal Park (CCCP) is located in far East Gippsland (37°48′S, 148°52′E). Our 13,000-ha site was established as a protected area in 1997 and is a mosaic of coastal heath, banksia woodland and lowland forest. Both lace monitors and foxes are present, the fox probably arriving and establishing after 1900 (Abbott 2011). Fox baiting has occurred since 1998 throughout parts of the park, with ‘1080’ (sodium monofluoroacetate) baits deployed on a regular basis by the Southern Ark fox management program (Murray et al. 2006). Due to its widespread distribution, baiting is aimed at fox suppression rather than complete eradication, so that densities are kept well below 1 fox km−2 (Diment 2010).
Point Lonsdale is a coastal community located on a rocky outcrop at the south-eastern end of the Bellarine Peninsula, approximately 100 km south of Melbourne (38°17′6″S, 144°36′8″E). The site covers 4,000 ha and is bordered by both semi-urban and agricultural land. The habitat is fragmented due to urbanization but retains substantial remnant vegetation and dense clusters of coastal tea tree (Leptospermum laevigatum) that provide a good habitat for ringtails (How et al. 1984). Point Lonsdale is outside the geographical range of the lace monitor. Foxes were first recorded here in 1845 but probably established around 1878–1881 (Abbott 2011); they are now abundant (>1 fox km−2) and widespread and have not been subject to any management.
Phillip Island is located 140 km south-east of Melbourne (38°28′60S, 145°13′60E). This island covers 10,000 ha and is located in Bass Strait (south and west coast) and Western Port Bay (north coast). The landscape consists of cleared grazing land, remnant and revegetated forest, estuarine vegetation, coastal heath and urban areas. Phillip Island is connected to the mainland via a permanent artificial bridge. The lace monitor is not present as Phillip Island is outside its geographical range. The red fox was first recorded on the island in 1907 (Gliddon 1968) and has been subject to control efforts since the 1930s. Baiting maintains the population island-wide at very low levels of just 15–30 breeding pairs (Berry and Kirkwood 2010).
In terms of testing hypothesis (2), fox impact was expected to be greatest at Point Lonsdale, intermediate at Cape Conran and least on Phillip Island, thus forming a cline in impact strength. The contrasting sympatry of ringtails and monitors at Cape Conran and allopatry at the other sites allowed testing of hypothesis (3).
Experimental design
Experimental manipulations were carried out at each site using four odors: a procedural non-odor control (distilled water); a chemical pungency control (1 part cologne to 5 parts distilled water); odors from the introduced predator (fox) and native predator (lace monitor), respectively. Predator odors were obtained by liquidizing freshly collected fecal material with distilled water in a ratio of 1 part feces to 5 parts water. The chemical pungency control was expected to provide a non-predator odor and allow us to distinguish whether odor-responses by ringtails were specific to predator odors or simply responses to strong odor per se.
Ringtails were located in situ in each study site on branches in low trees and shrubs using hand-held variable 100-W spotlights (Faunatech, Victoria, Australia) from 1 to 6 h after sunset when the animals were active. After discovery, each individual was approached slowly under low light to reduce further disturbance. A 2-m pole with an odor-soaked cloth at the end was raised slowly and held steady in a fixed position at the face level of the animal for 30 s. A pilot trial showed that if individuals responded to an odor it would occur within this timeframe. Separate poles were used for each odor to ensure no cross-contamination of scents, and at all sites each odor was deployed to ten different ringtail possums (n = 40 ringtails per site). To ensure independence, home ranges were not sampled again once an individual had been used in an odor trial. We recorded observations digitally during each response trial and also noted the time, height and vegetation type where each animal was observed. Trials were not conducted during full moon or on nights immediately preceding or following a full moon, or on nights with high wind speeds (Wayne et al. 2005a, b) in order to reduce the possibility that unfavorable environmental conditions might alter behavior through increasing predation risk. All trials were completed between November 2010 and February 2011.
Data analysis
A two-step analysis was used to assess the responses of ringtails to the experimental odor manipulations. These steps quantified ringtails’ responses to the initial approach of the observer, and then to the non-predator (odor control) and predator cues (recognition and response). As our approach to animals could be interpreted as a stimulus, we first wanted to test whether our presence introduced any systematic bias into their subsequent responses to the experimental odors. Digital recordings were used initially to categorize each individual’s posture upon our approach into three escalating levels, termed ‘aware’, ‘alert’ and ‘alarmed’. An individual was categorized as aware if it looked towards the observer and continued to hold its body parallel with the branch it was detected upon and to pursue the activities that it was engaged in when first seen. Individuals were scored as alert if they stopped engaging in an activity when they were first detected and raised their body to an angle of 45° or more, and they were scored as alarmed if they raised their forelimbs and prepared to flee. We used generalized log-linear modeling (GLM) and Pearson χ2 tests of independence to determine whether these initial postures differed between study sites or odor treatments.
Secondly, to describe the responses of ringtails to the four different odors, we again used our digital recordings to score 11 types of behavior in the broad categories of ‘movement’, ‘alertness’ and ‘other’ (Table 1). We used J-Watcher (ver. 1.0) to tally the frequency of discreet behaviors and measure the duration of continuous behaviors (Blumstein and Daniel 2007). To test hypothesis (1) we compared the behavioral responses of ringtails to the four different odors pooled across all study sites. We tested hypotheses (2) and (3) by comparing the responses of ringtails to the odors of fox and lace monitor, respectively, between the study sites. Generalized linear modeling again was used to analyze behavioral responses, with discreet behaviors compared using multinomial GLM and Pearson χ2 tests of independence and the single continuous variable (response time, or time to flee) analyzed using linear GLM with pairwise comparisons for different odors within and between sites. Additionally, we compared discrete behaviors displayed by animals to the two control odors and the two predator odors using multinomial GLM. All tests were carried out using SPSS ver.15.0 (SPSS 2006).
Results
Initial responses
The initial posture of individuals upon approach suggested high awareness but very little perception of risk. Thus, 76 % of individuals were scored as ‘aware’ and 24 % as ‘alert’, with no individuals recorded in the ‘alarm’ posture. The distribution of these postures was consistent among trials, with no differences detected between sites (GLM: \( \chi_{2}^{2} \) = 2.95, P = 0.229) or odors (\( \chi_{3}^{2} \) = 0.656, P = 0.418).
Responses to odors
In terms of the time taken for them to flee, ringtails responded to the four odors very differently (GLM: \( \chi_{3}^{2} \) = 134.25, P < 0.001). While the mean time to flee was slow and almost identical for the two control odors (26.6 s; P = 0.95), by (Varanus varius); comparison animals fled 3.8-fold more rapidly when exposed to fox odor and 3.1-fold faster in response to the odor of lace monitors (Fig. 2). Comparisons across sites revealed no difference in the time taken for ringtails to respond to fox odor (GLM: \( \chi_{2}^{2} \) = 3.332, P = 0.189) (Fig. 3). There was also no difference in response time to lace monitor odor between sites (GLM: \( \chi_{2}^{2} \) = 0.773, P = 0.679) or in the time that ringtails took to respond to the two predator odors within each site (CCCP: GLM: \( \chi_{1}^{2} \) = 0.278, P = 0.598; Phillip Island: GLM: \( \chi_{1}^{2} \) = 1.102, P = 0.294; Point Lonsdale (GLM \( \chi_{1}^{2} \) = 2.667, P = 0.102).

Pooled response times [seconds ± standard error (SE)] of common ringtail possums (Pseudocheirus peregrinus; n = 120) across Victoria to each of four olfactory cues (n = 30): chemical pungency control, procedural control and fox (Vulpes vulpes) and lace monitor (Varanus varius; goanna) fecal odor

Response times (seconds ± SE) of common ringtail possums (n = 60) to lace monitor (goanna) and fox odors at three sites that vary in coexistence histories with these two predator archetypes with respect to fox introduction and management and geographical range of the goanna: Cape Conran Coastal Park (fox suppression and goanna sympatry), Phillip Island (fox suppression and goanna allopatry) and Point Lonsdale (no fox suppression and goanna allopatry)
In terms of discreet behavioral responses by the ringtail to odor, there was only one difference in all the between-site comparisons (Table 2). A significant difference was found between sites in the posture of ringtails at the cessation of their exposure to fox odor (GLM: \( \chi_{4}^{2} \) = 19.741, P = 0.001), with 60 % of individuals at CCCP in the ‘alarmed’ category after exposure to fox odor, compared with no individuals at either Point Lonsdale or Phillip Island. There were several differences in discrete behaviors between the two control odors and the two predator odors, with very marked increases observed in escape behaviors in the presence of predator odor, including the direction and speed of movement and alertness at cessation of exposure to the odor (Table 3).
Discussion
The results of our study suggest that common ringtail possums have retained specific anti-predator responses towards the olfactory cues of an ancestral predator and that they have developed anti-predator responses towards the olfactory cues of an evolutionarily novel predator. Upon discovery, ringtails were either aware of, or alert to, our presence; while animals looked towards us, perhaps to monitor the level of threat that we represented, none fled. Upon presentation with the two odor-control stimuli, ringtails continued to exhibit very little response. Most animals simply ceased foraging and maintained their initial posture or oriented themselves towards the stimulus while remaining apparently unthreatened. In several instances, ringtails continued to forage or showed investigative behavior, holding onto the pole or the cloth, sniffing and even on occasion licking the pole. In contrast, the odors of both lace monitor and fox elicited strong anti-predator responses. In almost all instances, ringtails immediately stopped foraging to engage in vigilance and escape strategies, with alarm calling employed on three occasions in addition to these other behavioral responses (Table 3). There was little difference in behavioral response to the odor of either predator, even though ringtails might be expected to escape foxes most readily by climbing up and to escape monitors by freezing or moving to thin twigs that could support their weight but not that of the more massive predator. Taken together, these findings provide strong support for our first hypothesis.
With respect to our second hypothesis, we predicted that ringtails would show varied levels of response to red fox odor that would be calibrated by the level of fox impact that had been experienced by animals across the study sites. This hypothesis was not supported. On the one hand, there was a trend for animals to flee more quickly when exposed to fox odor at the most strongly fox-impacted site (Point Lonsdale) than at the less impacted sites (Fig. 3), as we had predicted. However, the trend was not significant; post hoc power analysis (Cohen 1988) indicated that a doubling of the sample size would be needed to achieve a significant effect at the 5 % level, suggesting that—even if real—any grading of response by ringtails to fox impact would be weak. On the other hand, the lack of response to fox odor in all but one of the discreet categories of behavior (Table 2) and the high level of alarm shown by ringtails to fox odor at Cape Conran, but not at the other two study sites, were entirely inconsistent with our second hypothesis.
There are several possible reasons why ringtails showed generally invariant levels of response to fox odor across sites. Firstly, ringtails may show a general avoidance of canid odor owing to their relatively long (4,500 year) co-occurrence with the Australian ‘native dog’, or dingo (Canis lupus dingo). Although this predator is rare in Victoria due to continuous persecution from humans, it includes ringtails in its diet at low but consistent frequencies (Brown and Triggs 1990; Glen et al. 2006, 2011). However, Banks et al. (2003) argued that wild dog feces were likely to be too sparse to provide reliable indicators of predator encounter and demonstrated that native Australian bush rats did not avoid the feces of domestic dogs. In addition, the fear-inducing chemical constituent of fox feces—2,4,5-trimethyl-thiazoline—(Vernet-Maury 1980) is absent from the feces of dogs (Arnould et al. 1998), perhaps allowing ringtails to distinguish between the two predators. These observations do not preclude the possibility that ringtails show general avoidance of canid fecal odors, but we consider this explanation unlikely.
Secondly, the impacts of predators can be mediated by habitat structure. Preference by prey for structurally complex habitats can represent general habitat-based avoidance behavior irrespective of predator type (Sutherland and Predavec 1999; Stokes et al. 2004). In our study, however, we are confident that the perception of predation risk by ringtails was not unduly influenced by habitat structure. Habitats were very similar throughout our study sites, with ringtails in each site showing a particular preference for complex mid- and under-storey habitats.
Thirdly, we assumed that animals that had been subject to limited fox impact would be more naïve to this novel predator than animals in sites with a longer fox presence and higher densities. Naiveté towards fox odor has previously been demonstrated in several Australian mammals, including bush rats (Banks 1998; McEvoy et al. 2008), antechinuses (Russell and Banks 2007), hare-wallabies (McLean et al. 1996), quolls (Jones et al. 2004) and tammar wallabies (Griffin et al. 2001), and we therefore expected the ringtail possum also to be relatively fox naïve at sites where its exposure to the fox had been limited. It is possible, however, that fox density and impact on prey may be decoupled due to certain aspects of fox behavior, such as surplus killing (Kruuk 1972). In this case, the per capita effect of foxes on prey increases as fox density falls because prey that are surplus to the predators’ energetic requirements are killed anyway and cached (Short et al. 2002). Even single foxes can rapidly deplete small populations of naïve prey and thus exert strong selection for anti-predator behavior (Short et al. 2002; Dickman 2012). Rapid evolution of behavioral responsiveness to a novel predator has been demonstrated previously, with native black snakes (Pseudechis porphyriacus) developing behavioral adaptations to the invasive cane toad (Bufo marinus) within 70 years of the latter’s introduction to Australia in 1935 (Phillips and Shine 2006). Studies of island populations also have demonstrated potential plasticity or rapid evolution of behavioral traits in response to novel predators. Island foxes Urocyon littoralis subject to predation by invasive golden eagles (Aquila chrysaetos) rapidly shifted their activity patterns (Hudgens and Garcelon 2011), while New Zealand island birds responded quickly to novel predatory mammals with behavioral changes in parental care (Massaro et al. 2008). In all of these studies, prey have responded within a few generations to strong predator-induced selection; we suspect this has occurred here with ringtails.
Our third hypothesis also received no support. We predicted that ringtails would be less sensitive to the fecal odors of the native lace monitor outside the predator’s geographical range than where the two species co-occur. However, ringtails fled quickly when presented with olfactory cues of the monitor in all sites, despite the allopatric occurrence of populations at Point Lonsdale and Phillip Island. This result can be interpreted in several ways: (1) ringtails show aversive behavior generally to predator odors; (2) aversive behavior is maintained by the presence of similar predator archetypes; (3) monitors and ringtails have co-occurred at all our study sites until so recently that ringtails still retain specific aversive behavior to monitors. We discuss these possibilities below and suggest that the latter two interpretations are probably most applicable.
Although ringtails showed similar avoidance of the fecal odors of fox and lace monitor, these odors are quite different to the human nose; given the well-developed powers of olfactory discrimination of P. peregrinus (Walker and Croft 1990) and the ability of animals in our study to distinguish between predator and strong control odors, it is plausible that ringtails should also be able to distinguish the two predator odors. It is more likely that monitor fecal odor is similar to that of other ringtail predators that occur in our study sites. These could include forest owls, such as the powerful owl (Ninox strenua), which hunts ringtails as a major prey species in southern Victoria (Cooke et al. 2002). If so, this could represent an example of the multi-predator hypothesis, with ringtails showing aversive behavior towards monitor odor because of persistent pressure from olfactorily similar predator types (Blumstein 2006). Future work could resolve the generality of ringtail responses to predator odor by running tests of animals’ responses to smaller and non-threatening sympatric predators, such as phascogales (Phascogale tapoatafa) or larger allopatric predators.
The past distribution of the lace monitor is not known, but spatial modeling suggests that this species can occupy diverse habitats in Victoria and would likely have occurred more broadly before disturbance and fragmentation of its preferred woodland habitats (Poodat et al. 2011). We do not know whether it once occurred at or near Point Lonsdale or the Bellarine Peninsula, but this possibility seems likely. In addition, gene flow between ringtails in previously continuous forest (Lancaster et al. 2011) may have been sufficient to maintain some level of recognition of lace monitors even in sites where monitors did not occur. Behavioral responses towards ancestral predators have been retained similarly in the Californian ground squirrel (Otospermophilus beecheyi), which recognizes snakes as potential predators after 70,000–300,000 years of isolation (Coss 1999), and in marine iguanas (Amblyrhynchus cristatus) over even longer periods (Berger et al. 2007). Likewise, deer mice (Peromyscus maniculatus) on islands previously inhabited by island foxes (Urocyon littoralis) still display aversion to fox odor, whereas rodents on historically fox-free islands do not (Orrock 2010). In many of these situations, the costs of maintaining anti-predator behaviors may be small and offset further if they reduce impacts from other predators. However, such responses are not uniform. Island primates (Simias concolor) exposed to ancestral predator cues may respond to the novel stimulus but not with specific anti-predator behaviors (Yorzinski and Ziegler 2007). Geckos also respond to sympatric predators rather than allopatric ones, with this response driven by the coevolution of prey behavior and predator foraging tactics rather than the loss of ancestral traits (Downes and Shine 1998).
One limitation of our study may have been in using only olfactory cues. Martin et al. (2010) found that roach (Rutilus rutilus) avoidance behavior changed between olfactory and visual cues, suggesting a difference between perceiving versus locating a threat. Evolutionary and ecological naiveté in prey can prevent predator recognition and the mounting of appropriate anti-predator behaviors. If specific anti-predator responses differ between perception and location, olfactory studies may be important in establishing a species’ capability in recognizing a predator, but insufficient to establish specific anti-predator behaviors. It is difficult to gauge if ringtails are employing appropriate behaviors to specific predators; if they are not, this may explain the lack of differentiation in discrete behavioral responses to predator cues (Table 2). Nevertheless, we have established that predator recognition occurs in this marsupial to both novel invasive and native predators regardless of their evolutionary history or geographical range.
The direct effects of invasive predators in Australia have contributed to the extinctions of several species of native vertebrates (Saunders et al. 2010), and the consumptive effects of these predators on extant species remain stronger that those on prey in other parts of the world (Salo et al. 2007). While prey naiveté provides a partial explanation for this, our results suggest that ringtails are no longer naïve and have rapidly developed anti-predator behavior which is similar to that shown towards a key native predator. Strauss et al. (2006) predicted that common and widespread native species would be more likely than rare or localized species to survive the initial impact of an invasive predator and then would be subject to strong selection to avoid it. Our results are consistent with the prediction of a widespread species undergoing selection to resist an invasive predator, and would further benefit from comparison with the responses shown by more localized species. From a conservation perspective, understanding the rate of behavioral adaptation by prey to novel predators is of great importance, and one that requires further research across a broad range of prey species.
References
Abbott I (2011) The importation, release, establishment, spread and early impact of prey animals of the red fox Vulpes vulpes in Victoria and adjoining parts of south-eastern Australia. Aust Zool 35:463–533
Apfelbach R, Blanchard CD, Blanchard RJ, Hayes RA, McGregor IS (2005) The effects of predator odors in mammalian prey species: a review of field and laboratory studies. Neurosci Biobehav Rev 29:1123–1144
Arenz CL, Leger DW (1997) Artificial visual obstruction, antipredator vigilance, and predator detection in the thirteen-lined ground squirrel (Spermophilus tridecemlineatus). Behaviour 134:1101–1114
Arnould C, Malosse C, Signoret JP, Descoins C (1998) Which chemical constituents from dog feces are involved in its food repellent effect in sheep? J Chem Ecol 24:559–576
Banks PB (1998) Responses of Australian bush rats, Rattus fuscipes, to the odor of introduced Vulpes vulpes. J Mammal 79:1260–1264
Banks PB, Dickman CR (2007) Alien predation and the effects of multiple levels of prey naivete. Trends Ecol Evol 22:229–230
Banks PB, Newsome AE, Dickman CR (2000) Predation by red foxes limits recruitment in populations of eastern grey kangaroos. Aust Ecol 25:283–291
Banks PB, Hughes NK, Rose TA (2003) Do native Australian small mammals avoid faeces of domestic dogs? Responses of Rattus fuscipes and Antechinus stuartii. Aust Zool 32:406–409
Berger J, Swenson JE, Persson IL (2001) Recolonizing carnivores and naive prey: conservation lessons from Pleistocene extinctions. Science 291:1036–1039
Berger S, Wikelski M, Romero LM, Kalko EK, Roedl T (2007) Behavioral and physiological adjustments to new predators in an endemic island species, the Galapagos marine iguana. Horm Behav 52:653–663
Berry O, Kirkwood R (2010) Measuring recruitment in an invasive species to determine eradication potential. J Wildl Manag 74:1661–1670
Blumstein DT (2006) The multipredator hypothesis and the evolutionary persistence of antipredator behavior. Ethology 112:209–217
Blumstein DT, Daniel JC (2005) The loss of anti-predator behaviour following isolation on islands. Proc Royal Soc B Biol Sci 272:1663–1668
Blumstein DT, Daniel JC (2007) Quantifying behavior the JWatcher way. Sinauer Associates, Sunderland
Blumstein DT, Daniel JC, Springett BP (2004) A test of the multi-predator hypothesis: rapid loss of antipredator behavior after 130 years of isolation. Ethology 110:919–934
Blumstein DT, Ferando E, Stankowich T (2009) A test of the multipredator hypothesis: yellow-bellied marmots respond fearfully to the sight of novel and extinct predators. Anim Behav 78:873–878
Brown GW, Triggs BE (1990) Diets of wild canids and foxes in east Gippsland Australia 1983–1987 using predator scat analysis. Aust Mammal 13:209–214
Cohen J (1988) Statistical power analysis for the behavioral sciences, 2nd edn. Lawrence Erlbaum Associates, Hillsdale
Cooke R, Wallis R, White J (2002) Use of vegetative structure by powerful owls in outer urban Melbourne, Victoria, Australia—implications for management. J Raptor Res 36:294–299
Coss RG (1999) Effects of relaxed natural selection on the evolution of behavior. In: Foster SA, Endler JA (eds) Geographic variation in behavior: perspectives on evolutionary mechanisms. Oxford University Press, Oxford, pp 180–208
Coss RG, Ramakrishnan U (2000) Perceptual aspects of leopard recognition by wild bonnet macaques (Macaca radiata). Behaviour 137:315–335
Devineau O, Shenk TM, White GC, Doherty PF, Lukacs PM, Kahn RH (2010) Evaluating the Canada lynx reintroduction programme in Colorado: patterns in mortality. J Appl Ecol 47:524–531
Dickman CR (2012) Fences or ferals? Benefits and costs of conservation fencing in Australia. In: Somers MJ, Hayward MW (eds) Fencing for conservation: restriction of evolutionary potential or a riposte to threatening processes?. Springer, New York, pp 43–63
Dickman CR, Doncaster CP (1984) Responses of small mammals to red fox (Vulpes vulpes) odour. J Zool 204:521–531
Diment AN (2010) Monitoring the ecological impacts of invasive predator control. PhD thesis. University of Sydney, Sydney
Downes S, Shine R (1998) Sedentary snakes and gullible geckos: predator–prey coevolution in nocturnal rock-dwelling reptiles. Anim Behav 55:1373–1385
Endler JA, Houde AE (1995) Geographic-variation in female preferences for male traits in Poecilia reticulata. Evolution 49:456–468
Fendt M, Endres T, Lowry CA, Apfelbach R, McGregor IS (2005) TMT-induced autonomic and behavioral changes and the neural basis of its processing. Neurosci Biobehav Rev 29:1145–1156
Glen AS, Dickman CR (2005) Complex interactions among mammalian carnivores in Australia, and their implications for wildlife management. Biol Rev 80:387–401
Glen AS, Dickman CR (2008) Niche overlap between marsupial and eutherian carnivores: does competition threaten the endangered spotted-tailed quoll? J Appl Ecol 45:700–707
Glen AS, Fay AR, Dickman CR (2006) Diets of sympatric red foxes (Vulpes vulpes) and wild dogs (Canis lupus) in the Northern Rivers Region, New South Wales. Aust Mammal 28:101–104
Glen AS, de Tores PJ, Sutherland DR, Morris KD (2009) Interactions between chuditch (Dasyurus geoffroii) and introduced predators: a review. Aust J Zool 57:347–356
Glen AS, Pennay M, Dickman CR, Wintle BA, Firestone KB (2011) Diets of sympatric native and introduced carnivores in the Barrington Tops, eastern Australia. Aust Ecol 36:290–296
Gliddon JW (1968) Phillip Island; in picture and story. Committee of Trust “Warley”, Cowes Bush Nursing Hospital, Cowes
Gresser S (1996) Anti-predator behaviour of the common brushtail possum (Trichosurus vulpecula) at Burrendong Dam. PhD thesis. University of Sydney, Sydney
Griffin AS, Blumstein DT, Evans C (2000) Training captive-bred or translocated animals to avoid predators. Conserv Biol 14:1317–1326
Griffin AS, Evans CS, Blumstein DT (2001) Learning specificity in acquired predator recognition. Anim Behav 62:577–589
Guarino F (2001) Diet of a large carnivorous lizard, Varanus varius. Wildl Res 28:627–630
Hayes RA, Nahrung HF, Wilson JC (2006) The response of native Australian rodents to predator odours varies seasonally: a by-product of life history variation? Anim Behav 71:1307–1314
Hollen LI, Manser MB (2007) Persistence of alarm-call behaviour in the absence of predators: a comparison between wild and captive-born meerkats (Suricata suricatta). Ethology 113:1038–1047
How RA, Barnett JL, J BA, Humphries WF, Martin R (1984) The population biology of Pseudocheirus peregrinus in a Leptospermum laevigatum thicket. In: Smith AP, Hume ID (eds) Possums and gliders. Surrey Beatty and Sons/Australian Mammal Society, Sydney, pp 261–268
Hudgens BR, Garcelon DK (2011) Induced changes in island fox (Urocyon littoralis) activity do not mitigate the extinction threat posed by a novel predator. Oecologia 165:699–705
Jessop TS, Urlus J, Lockwood T, Gillespie G (2010) Preying possum: assessment of the diet of lace monitors (Varanus varius) from coastal forests in southeastern Victoria. Biawak 4:59
Jones ME, Smith GC, Jones SM (2004) Is anti-predator behaviour in Tasmanian eastern quolls (Dasyurus viverrinus) effective against introduced predators? Anim Conserv 7:155–160
Kats LB, Dill LM (1998) The scent of death: chemosensory assessment of predation risk by prey animals. Ecoscience 5:361–394
King D, Green B (1999) The biology of Varanid lizards. University of NSW Press, Sydney
Kinnear JE, Onus ML, Sumner NR (1998) Fox control and rock-wallaby population dynamics–II. An update. Wildl Res 25:81–88
Kovacs EK, Crowther MS, Webb JK, Dickman CR (2012) Population and behavioural responses of native prey to alien predation. Oecologia 168:947–957
Kruuk H (1972) Surplus killing by carnivores. J Zool 166:233–244
Lancaster ML, Taylor AC, Cooper SJB, Carthew SM (2011) Limited ecological connectivity of an arboreal marsupial across a forest/plantation landscape despite apparent resilience to fragmentation. Mol Ecol 20:2258–2271
Lima SL (1998) Nonlethal effects in the ecology of predator–prey interactions—what are the ecological effects of anti-predator decision-making? Bioscience 48:25–34
Lindenmayer DB, MacGregor C, Welsh A, Donnelly CF, Brown D (2008) The use of hollows and dreys by the common ringtail possum (Pseudocheirus peregrinus) in different vegetation types. Aust J Zool 56:1–11
Macdonald DW (1980) Patterns of scent marking with urine and faeces amongst carnivore communities. Symp Zool Soc London 45:107–139
Martin CW, Fodrie FJ, Heck KL, Mattila J (2010) Differential habitat use and antipredator response of juvenile roach (Rutilus rutilus) to olfactory and visual cues from multiple predators. Oecologia 162:893–902
Massaro M, Starling-Windhof A, Briskie JV, Martin TE (2008) Introduced mammalian predators induce behavioural changes in parental care in an endemic New Zealand bird. Plos One 3(6):e2331
McEvoy J, Sinn DL, Wapstra E (2008) Know thy enemy: behavioural response of a native mammal (Rattus lutreolus velutinus) to predators of different coexistence histories. Aust Ecol 33:922–931
McLean IG, Lundie-Jenkins G, Jarman PJ (1996) Teaching an endangered mammal to recognise predators. Biol Conserv 75:51–62
Mella VSA, Cooper CE, Davies S (2010) Predator odour does not influence trappability of southern brown bandicoots (Isoodon obesulus) and common brushtail possums (Trichosurus vulpecula). Aust J Zool 58:267–272
Miller B, Ralls K, Reading RP, Scott JM, Estes J (1999) Biological and technical considerations of carnivore translocation: a review. Anim Conserv 2:59–68
Muller-Schwarze D (2006) Chemical ecology of vertebrates. Cambridge University Press, Cambridge
Murray AJ, Poore RN, Dexter N (2006) Project Deliverance—the response of ‘critical weight range’ mammals to effective fox control in mesic forest habitats in far East Gippsland, Victoria. Department of Sustainability and Environment, Melbourne
Olsson M, Wapstra E, Swan G, Snaith E, Clarke R, Madsen T (2005) Effects of long-term fox baiting on species composition and abundance in an Australian lizard community. Aust Ecol 30:907–913
Orrock JL (2010) When the ghost of predation has passed: do rodents from islands with and without fox predators exhibit aversion to fox cues? Ethology 116:338–345
Phillips BL, Shine R (2006) An invasive species induces rapid adaptive change in a native predator: cane toads and black snakes in Australia. Proc Royal Soc B Biol Sci 273:1545–1550
Poodat F, Arrowsmith C, Farmer E (2011) Modelling habitat networks using the concept of graph theory. In: Surveying and Spatial Sciences Biennial Conference, Wellington
Ritchie EG, Johnson CN (2009) Predator interactions, mesopredator release and biodiversity conservation. Ecol Lett 12:982–998
Russell BG, Banks PB (2007) Do Australian small mammals respond to native and introduced predator odours? Aust Ecol 32:277–286
Russell BG, Smith B, Augee ML (2003) Changes to a population of common ringtail possums (Pseudocheirus peregrinus) after bushfire. Wildl Res 30:389–396
Salo P, Korpimäki E, Banks PB, Nordstrom M, Dickman CR (2007) Alien predators are more dangerous than native predators to prey populations. Proc Royal Soc B Biol Sci 274:1237–1243
Salo P, Banks PB, Dickman CR, Korpimäki E (2010) Predator manipulation experiments: impacts on populations of terrestrial vertebrate prey. Ecol Monogr 80:531–546
Saunders GR, Gentle MN, Dickman CR (2010) The impacts and management of foxes Vulpes vulpes in Australia. Mamm Rev 40:181–211
Short J, Kinnear JE, Robley A (2002) Surplus killing by introduced predators in Australia—evidence for ineffective anti-predator adaptations in native prey species? Biol Conserv 103:283–301
Sih A, Christensen B (2001) Optimal diet theory: when does it work, and when and why does it fail? Anim Behav 61:379–390
Sih A, Bolnick DI, Luttbeg B, Orrock JL, Peacor SD, Pintor LM, Preisser E, Rehage JS, Vonesh JR (2010) Predator–prey naivete, antipredator behavior, and the ecology of predator invasions. Oikos 119:610–621
Smith B, Augee M, Rose S (2003) Radio-tracking studies of common ringtail possums, Pseudocheirus peregrinus, in Manly Dam Reserve, Sydney. Proc Linnean Soc NSW 124:183–194
SPSS (2006) SPSS v. 15.0. SPSS Inc., Chicago
Stoddart DM (1982) Does trap odour influence estimation of population size of the short-tailed vole, Microtus agrestis? J Anim Ecol 51:375–386
Stokes VL, Pech RP, Banks PB, Arthur AD (2004) Foraging behaviour and habitat use by Antechinus flavipes and Sminthopsis murina (Marsupialia: Dasyuridae) in response to predation risk in eucalypt woodland. Biol Conserv 117:331–342
Strauss SY, Lau JA, Carroll SP (2006) Evolutionary responses of natives to introduced species: what do introductions tell us about natural communities? Ecol Lett 9:354–371
Sutherland DR, Predavec M (1999) The effects of moonlight on microhabitat use by Antechinus agilis (Marsupialia: Dasyuridae). Aust J Zool 47:1–17
Terborgh J. Lopez L, Nunez VP, Rao M, Shahabuddin G et al (2001) Ecological meltdown in predator-free forest fragments. Science 294:1923–1926
Triblehorn JD, Yager DD (2005) Acoustic interactions between insects and bats: a model of the interplay of neural and ecological specializations. In: Barbosa P, Castellanos I (eds) Ecology of predator–prey interactions. Oxford University Press, New York, pp 77–104
Triggs B, Brunner H, Cullen JM (1984) The food of fox, dog and cat in Croajingalong National Park, southeastern Victoria. Aust Wildl Res 11:491–499
Van Dyck S, Strahan R (2008) The mammals of Australia, 3rd edn. Reed New Holland, Sydney
Vernet-Maury E (1980) Trimethyl-thiazoline in fox feces: a natural alarming substance for the rat. Olfaction Taste 7:407
Walker LV, Croft DB (1990) Odour preferences and discrimination in captive ringtail possums (Pseudocheirus peregrinus). Int J Comp Psychol 3:215–234
Wayne AF, Cowling A, Rooney JF, Ward CG, Wheeler IB, Lindenmayer DB, Donnelly CF (2005a) Factors affecting the detection of possums by spotlighting in Western Australia. Wildl Res 32:689–700
Wayne AF, Cowling A, Ward CG, Rooney JF, Vellios CV, Lindenmayer DB, Donnelly CF (2005b) A comparison of survey methods for arboreal possums in jarrah forest, Western Australia. Wildl Res 32:701–714
Weavers BW (1988) Vital statistics of the lace monitor lizard (Varanus varius) in south-eastern Australia. Vic Nat 105:142–145
Weavers BW (1989) Diet of the lace monitor lizard Varanus varius in south-eastern Australia. Aust Zool 25:83–86
Yorzinski JL, Ziegler T (2007) Do naive primates recognize the vocalizations of felid predators? Ethology 113:1219–1227
Acknowledgments
The authors gratefully acknowledge the assistance of Tim Jessop. Kath Handasyde, Mark Elgar and Graeme Coulson provided invaluable advice in the initial planning stages. This work was conducted under Department of Sustainability Wildlife and National Parks Act (1975) research permit 10005037, and University of Melbourne ethics permit 0911328.2. The authors also acknowledge Michael Lindeman (Arthur Rylah Institute) and Mike Taylor (Zoos Victoria) for providing predator scats and all the volunteers who assisted with field work. This work was supported by Zoos Victoria and the Ecological Society of Australia.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Janne Sundell.
Rights and permissions
About this article
Cite this article
Anson, J.R., Dickman, C.R. Behavioral responses of native prey to disparate predators: naiveté and predator recognition. Oecologia 171, 367–377 (2013). https://doi.org/10.1007/s00442-012-2424-7
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-012-2424-7