
Abstract
One major characteristic of invasive alien species is their occurrence at high abundances in their new habitat. Flowering invasive plant species that are visited by native insects and overlap with native plant species in their pollinators may facilitate or disrupt native flower visitation and fertilisation by forming large, dense populations with high numbers of flowers and copious rewards. We investigated the direction of such a proposed effect for the alien invasive Rhododendron ponticum in Irish habitats. Flower visitation, conspecific and alien pollen deposition, fruit and seed set were measured in a self-compatible native focal plant, Digitalis purpurea, and compared between field sites that contained different relative abundances of R. ponticum. Flower visitation was significantly lower at higher alien relative plant abundances than at lower abundances or in the absence of the alien. Native flowers experienced a significant decrease in conspecific pollen deposition with increasing alien abundance. Heterospecific pollen transfer was very low in all field sites but increased significantly with increasing relative R. ponticum abundance. However, lower flower visitation and lower conspecific pollen transfer did not alter reproductive success of D. purpurea. Our study shows that indirect interactions between alien and native plants for pollination can be modified by population characteristics (such as relative abundance) in a similar way as interactions among native plant species. In D. purpurea, only certain aspects of pollination and reproduction were affected by high alien abundances which is probably a result of high resilience due to a self-compatible breeding system. Native species that are more susceptible to pollen limitation are more likely to experience fitness disadvantages in habitats with high relative alien plant abundances.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Exotic plant species can be found in almost all continents and biomes (Vitousek et al. 1997). Their impact is known to be related to their distribution range and abundance across that range (Parker et al. 1999). Some alien invasive plants have the ability to change the characteristics of their new habitats substantially at a large scale and transform conditions for co-occurring native plants by, for example, competition for physical resources and/or modification of ecosystem processes (Richardson et al. 2000; Weidenhamer and Callaway 2010). As well as abiotic resources such as water, nutrients, space and/or light which are crucial for plant survival and reproduction, an important biotic resource that many flowering plant species depend on are pollinators for sexual reproduction (Palmer et al. 2003). Thus, it is not surprising that pollination services have been suggested to shape the relationship between native and invasive alien flowering plant species (Traveset and Richardson 2006).
The impact of invasive aliens on native pollination services can be of a direct or an indirect nature. If the alien species uses the same flower visitors for pollen transfer as native co-flowering plants, it might act as a direct competitor or facilitator (Lopezaraiza-Mikel et al. 2007; Bartomeus et al. 2008b). Even if there is no overlap in pollinator species or if the invasive plant species is pollinated by abiotic vectors, invasion by an alien can cause displacement or replacement of native plants or a change in their colonisation patterns (Parker et al. 1999; Yurkonis and Meiners 2004) which might indirectly alter native pollination processes. Since the absolute and relative abundance of each plant species within a community structures the complex network of mutualistic interactions (Stang et al. 2006), it is not only the presence of an alien invader but also its abundance which is likely to affect pollination services of the co-occurring plant species (Muñoz and Cavieres 2008). Few observational and experimental studies have addressed the effect of abundance on pollination and reproductive success in multi-species assemblages. In native communities, indirect interactions for pollination among plant species are sensitive to morphological and physiological characteristics and the relative abundance of each species (Rathcke 1983). Individuals of more abundant species can either facilitate pollinator visitation of heterospecific individuals via pollinator sharing (magnet species effect; Thomson 1978; Moeller 2004), or they can monopolise flower constant pollinators due to the relatively more abundant rewards they offer (Chittka et al. 1997). Within the same species assemblage, facilitation of pollinator visitation can even shift to competition and vice versa as flower abundances of the species change along a continuum (Ghazoul 2006). In addition to this quantitative measure of pollinator visitation, the quality of visitation, e.g. the amount of heterospecific pollen deposited and conspecific pollen deposition and wastage, may also change according to the abundance and density of the plant species (Feinsinger et al. 1986; Caruso 2002; but see Waites and Ågren 2004). Changes in quantity and quality can translate into changes in reproductive success (Feinsinger et al. 1991). Indeed, there have also been examples where the presence or increased abundance of a native plant species did not affect conspecific pollen deposition or fitness of individuals of another native co-flowering target species (Feinsinger et al. 1986; Feldman 2008).
Interactions between alien and native plant species can be characterised in the same way as interactions among native species. They have competitive, facilitative or neutral effects on pollination and are also expected to respond to species abundance (Brown et al. 2002; Ghazoul 2004; Bjerknes et al. 2007; Morales and Traveset 2009). The rapidity of range expansion which successful invaders often exhibit in their new habitats can lead to high relative abundances of the invasive species in comparison to native species (Sakai et al. 2001). This might not only have implications for native plant populations but also for pollinator communities (Bjerknes et al. 2007). Therefore, invaded communities offer excellent opportunities to investigate basic population processes like pollination interactions (Sax et al. 2007). Understanding these processes is essential if we want to assess and manage impacts of alien invasions (Levine et al. 2003).
One of the most severe invaders in Britain and Ireland that requires sustainable management is Rhododendron ponticum L. (Ericaceae), an evergreen, perennial shrub that has invaded Irish heaths, bogs and woodlands over the last two centuries (Cross 1975). Its pink-purple, zygomorphic flowers are held in large inflorescences of 9–21 flowers which can form impressive floral displays; each flower produces large volumes of sugar-rich nectar which attracts a range of native flower visitors (Stout et al. 2006). In its invaded habitat, R. ponticum co-flowers with a variety of 20 and more native plant species (Cross 1975). Depending on the habitat and environmental conditions, R. ponticum occurs as scattered single individuals but can also form large clumps or pure stands (Cross 1975); among other factors, such differences in relative plant abundance are likely to characterise the impact an invasive alien species has on its invaded community (Morales and Traveset 2009).
In this study, we investigated the role of relative R. ponticum abundance on the pollination of a native target species Digitalis purpurea. Our objectives were to determine the effect of relative R. ponticum abundance on the number of insect visits to individual D. purpurea plants and flowers, conspecific and heterospecific (alien) pollen deposition on D. purpurea stigmata and seed set of D. purpurea.
Materials and methods
Native study species
Digitalis purpurea L. (Scrophulariaceae) is a facultative biennial herb that can be commonly found on earth banks and in transitional woodland habitats around Ireland (Fossitt 2000). Like R. ponticum, it produces pink-purple, zygomorphic flowers which secrete copious nectar, but in D. purpurea these are arranged in one or a few inflorescences per individual plant (Percival and Morgan 1965). Although D. purpurea is a self-compatible plant species, it is incapable of spontaneous autogamy and relies on biotic vectors for pollen transfer (Nazir et al. 2008). In Ireland, D. purpurea frequently co-occurs with R. ponticum, and the two species overlap in flowering time (Percival and Morgan 1965; Cross 1975) and are pollinated by the same functional group of pollinators (bumblebees; Percival and Morgan 1965; Stout 2007).
Given that R. ponticum not only resembles D. purpurea in phenology and pollinators but also in flower symmetry and colour, the alien is likely to impact pollination of D. purpurea, because phenotypic similarity between alien and native plants has been found to affect the outcome of alien–native plant interactions (Morales and Traveset 2009). Therefore, D. purpurea is a suitable native focal species for studying effects of R. ponticum and relative R. ponticum abundance on native pollination processes.
Study sites
Field observations were conducted during 2006 and 2007 at nine sites in the Wicklow Mountains area, Co. Wicklow, Ireland, in similar habitats over a range of altitudes (Table 1). We selected six invaded sites and three control sites. Control sites did not contain any R. ponticum individuals within a radius of at least 1 km. This distance minimised the impact of R. ponticum growing nearby on pollinator visitation to native plant species at these sites because most pollinators forage at a range that is on average smaller than this distance (Knight et al. 2005; Wolf and Moritz 2008). Invaded sites with the native target species D. purpurea were selected according to the abundance of the alien R. ponticum (Table 1). Relative abundance (% cover of plants) of R. ponticum was estimated in two (in one case three) randomly placed 20 m × 20 m quadrats which were surveyed on the ground and using aerial photographs. Since floral abundance per m2 R. ponticum shrub surface (measured by counting the number of open flowers in four randomly assigned 1 m × 1 m quadrats per invaded site) did not differ between sites of different relative R. ponticum abundance (ANOVA, F 1,4 = 0.19; P = 0.69), alien plant abundance is considered to be an estimate for alien flower abundance.
In 2006, we also estimated native flowering species cover and floral abundance by counting plant individuals, number of inflorescences and number of open flowers as well as estimating percentage cover of each species in ten 1 m × 1 m quadrats which were placed every 10 m along a 100-m transect. Thereby, R. ponticum thickets were not sampled. Vegetation data were used to obtain plant abundance estimates for the native target species D. purpurea as well as to calculate flower diversity indices for each site.
Insect visitation and nectar production
Both invaded and control field sites were visited once in each of 2006 and 2007 during peak R. ponticum flowering (June). Observations were conducted in fine weather conditions only (no rain and wind force <4 according to Beaufort Scale). During each visit, four flowering plant patches (of an average size of 1.0 ± 0.38 m2) of D. purpurea were investigated. In invaded sites, we chose D. purpurea patches in the vicinity (<10 m) of flowering R. ponticum individuals. Within each patch, the number of inflorescences and the number of open flowers on each inflorescence were counted, and patches were observed for 15 min between 0800 and 2000 hours. The identity of flower visitors, the number of inflorescences and number of flowers visited on each inflorescence were recorded. For each observed patch, we calculated visitation rate (i.e. the number of pollinator visits per 15 min/total number of observed flowers) and the number of visitors per inflorescences (i.e. the number of pollinators at the patch per 15 min/total number of inflorescences). Both response variables were included to allow us to explore subtle differences in pollination service.
After each observation session, we randomly picked ten openly accessible D. purpurea flowers and measured their nectar volume (nectar standing crop) with microcapillary glass pipettes (1 and 5 μl; Hirschmann Laborgeräte, Germany) and callipers.
Pollen desposition
In 2007, we collected receptive stigmata from one flower from each of 9 ± 1.0 SE (total 69) randomly chosen D. purpurea individuals at each site except for one site (Table 1) and stored them in vials filled with 70% ethanol. Stigmata were transferred to the laboratory, stained with a Safranin solution (0.5% Safranin in 50% ethanol) and individually placed on microscopic slides. Stigmata were gently squashed to allow inspection of the entire stigmatic surface. Both native and alien pollen on the stigmatic surface was counted using the tenfold magnification of a microscope (Leica, UK).
Seed set
In 2007, we selected 7–9 D. purpurea inflorescences at each of the sites (except for one site; Table 1) and counted and marked all their open flowers. Some of the marked inflorescences were vandalised or destroyed due to bad weather conditions, and so we sampled on average 6 ± 0.6 SE inflorescences at each site. Inflorescences were monitored and fruit capsules were harvested when mature. We measured their maximum length and maximum diameter with digital callipers (precision 0.01 mm; Moore & Wright, UK). From the total of 211 collected capsules, we randomly picked a subset of 55 fruits and counted the enclosed seeds. Seed numbers were most closely correlated to fruit diameter (Pearson’s correlation, r = 0.896, t 52 = 14.4, P < 0.001; two outliers were excluded from analysis), and so we estimated seed numbers of all marked fruits as 202 × fruit diameter − 666 (linear regression, adjusted R 2 = 0.799). Fruit/flower ratios (relative fruit set) and the average number of seeds per fruit in each inflorescence were then calculated whereby each capsule with at least one seed was considered to be a fruit.
Statistical analysis
Plant community and floral display of study sites
We calculated the mean cover and the mean number of inflorescences across all ten 1-m2 quadrats for each entomophilous native species; this gave a relative plant abundance estimate for the area of each site not covered by R. ponticum. We then incorporated the R. ponticum floral abundance data to calculate the Shannon index [H′ = −Σp i × loge(p i )] of overall floral diversity (\( H^{\prime}_{\text{flower}} \)) using the number of inflorescences of each species as importance values (p i ) and to calculate the Shannon index of entomophilous plant species diversity (\( H^{\prime}_{\text{species}} \)) using plant species cover data as importance values. Spearman rank correlation was employed to correlate these diversity values and overall floral abundance (number of inflorescences) with R. ponticum abundance. Additionally, we correlated R. ponticum abundance with D. purpurea estimates of percentage cover, number of inflorescences and number of open flowers using Spearman rank correlations in MINITAB 13 (Minitab, 2000).
Insect visitation
We used Cochran’s and Anderson–Darling tests to test for homogeneity of variances and normal distribution of the data (respectively) and subsequently square root transformed data of both variables (flower visitation rate and number of insect visitors per inflorescences). One extreme outlier (outside the range of mean by 3.3 standard deviations) of one observed patch at one site was identified for both variables and was therefore omitted from the data analysis. Visitation data were missing for one site in 2007 (Table 1) due to unfavourable weather conditions, and so we used Generalised Estimation Equations (GEEs) in order to do justice to the unbalanced data and the nested sampling design (Quinn and Keough 2002). We analysed transformed data within an independent working correlation matrix due to its best fit to the model (i.e. lowest Quasi-likelihood under Independence Model Criterion QIC, an extension of the Akaike information criteria) in SPSS 15.0 (SPSS, 2006). A normal distribution model with an identity link function was used (Agresti 1996). In the models, ‘year’ was incorporated as a factor, ‘abundance’ as a covariate and individual field sites as subjects (i.e. units from which we repeatedly took replicated samples). We also included the interaction term (year × abundance) and the intercept in the models. The best-fit model according to the lowest QIC was “error + year + abundance” for both variables.
To obtain an estimate of pollinator sharing between the native and the invasive species, we calculated the niche overlap index (NOI; Pianka 1973) for the field season 2006. The range of the index from 0 (no overlap) to 1 (absolute overlap) mirrors both visitor identities and visit frequencies of the pollinator guild. Further, the number of D. purpurea visitor taxa at invaded sites was correlated with relative R. ponticum abundance using Spearman rank correlation.
Nectar standing crop
Nectar standing crop data were log + 1 transformed to homogenise variances and allow for a normal distribution of data. We then analysed the relation between mean flower visitation rates and mean transformed nectar standing crop using Pearson product-moment correlation. MINITAB 13 was used for analysis.
Pollen deposition and seed set
The linear dependence of pollen deposition and seed set on R. ponticum abundance was modelled with Generalised Estimation Equations (GEEs) in order to take into account the unbalanced data and to incorporate the nested sampling design. R. ponticum abundance was used as a covariate and individual field sites as subjects, and the intercept was included in the models. We analysed conspecific pollen counts assuming a Poisson distribution and log link function. Seed set per fruit was analysed assuming a normal distribution with an identity link function. Proportions of alien pollen number/total pollen number and of fruit number/flower number (relative fruit set) were analysed using binomial distribution and logit link function (Agresti 1996). We used an independent working correlation structure and performed analyses with SPSS 15.0. A post hoc power analysis was conducted according to Quinn and Keough (2002).
Results are presented in the following section as mean ± standard errors unless indicated differently.
Results
Plant community and floral display of study sites
Percentage cover (6.7 ± 2.32), number of inflorescences (2.0 ± 0.84) and number of open flowers (4.5 ± 1.84) of D. purpurea in the 1-m2 quadrats decreased but were not significantly correlated with increasing R. ponticum abundance (Spearman r cover = −0.16, r infloresc = −0.43, r flowers = −0.27, n = 8, P > 0.28 for all), i.e. abundance of D. purpurea in patches next to R. ponticum did not significantly differ from D. purpurea abundances at control sites.
Although overall floral abundance (11.8 ± 3.31 inflorescences per m2) did not show any significant correlation with abundance of R. ponticum (Spearman r = 0.15, P = 0.73), overall floral diversity (0.13 ≤ \( H^{\prime}_{\text{flower}} \) ≥ 1.39) and diversity of entomophilous plant species (0.05 ≤ \( H^{\prime}_{\text{species}} \) ≥ 1.11) decreased with increasing R. ponticum abundance, i.e. at sites where the invasive became more dominant (Spearman r flower = −0.83, P = 0.011 and r species = −0.85, P = 0.008).
Visitation rates
Flower visitation rate to D. purpurea did not significantly differ between years (Wald χ 2 = 0.41, df = 1, n total = 67, n units = 9, P = 0.521), but was negatively influenced by an increase in R. ponticum abundance (Wald χ 2 = 3.97, df = 1, n total = 67, n units = 9, P = 0.046; βabundance = −0.002 ± 0.0010 SE; Fig. 1). Similarly, the numbers of insect visitors per D. purpurea inflorescence did not significantly vary between years (Wald χ 2 = 0.08, df = 1, n total = 67, n units = 9, P = 0.777), but inflorescences received significantly fewer flower visitors in sites where R. ponticum was more abundant (Wald χ 2 = 5.03, df = 1, n total = 67, n units = 9, P = 0.025; βabundance = −0.003 ± 0.0013 SE).

Flower visitation rate (number of visits per flower per 15 min) to Digitalis purpurea in habitats of different relative Rhododendron ponticum abundance in two consecutive years. For visual display, data were jittered along the x- and y-dimension so that overlapping data points are visible. Line indicates best-fit across both years; flower visitation rate: y = −0.002x + 0.388, r 2 = 0.056, P = 0.046. Note data are square root transformed
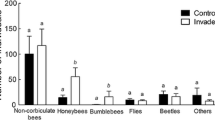
Digitalis purpurea at uninvaded control sites was only visited by bumblebees whereas D. purpurea at invaded sites received also visits from flies and syrphids (Table 2). Still, D. purpurea at control sites and D. purpurea at invaded sites overlapped largely in their pollinators (NOI = 0.881). Overlap in flower visitors between R. ponticum and D. purpurea increased by approx. 50% when D. purpurea co-occurred with R. ponticum at invaded sites (NOI = 0.402) in comparison to D. purpurea at control sites (NOI = 0.269; Table 2). The higher number of taxa visiting D. purpurea at invaded sites was not related to the relative abundance of R. ponticum at these sites (Spearman r = –0.34, n = 6, P > 0.51).
Nectar standing crop
Nectar standing crop of D. purpurea varied between 0 and 21.9 μl with less nectar on average in 2006 (1.0 μl ± 0.18) compared with 2007 (2.6 μl ± 0.60), and was negatively related to flower visitation rates of D. purpurea (Pearson correlation; r = −0.805; n = 16; P < 0.001; Fig. 2).

Correlation of flower visitation rate (mean ± SE) and nectar standing crop (mean ± geometric SE) of Digitalis purpurea. Note log scale used on y-axis
Pollen desposition
Deposition of conspecific pollen varied considerably from 24 to 3,034 conspecific pollen grains per individual D. purpurea stigma ranging from 1,238 ± 129 grains per stigma in control sites to 644 ± 123 SE in sites of high R. ponticum abundance. We observed a negative relationship between conspecific pollen deposition and the abundance of alien R. ponticum (Wald χ 2 = 5.46, df = 1, n total = 68, n units = 8, P = 0.019; βabundance = −0.006 ± 0.0025 SE; Fig. 3a).

a Number of Digitalis purpurea pollen grains and b percentage of alien Rhododendron ponticum pollen found on stigmata of D. purpurea as a function of relative R. ponticum abundance. For visual display, data were jittered along the x-dimension so that overlapping data points are visible. Lines indicate best-fit; conspecific pollen: y = −5.01x + 1,102, r2 = 0.076, P = 0.019; alien pollen: y = 0.0032x − 0.011, r2 = 0.27, P < 0.001
In contrast, pollen deposition of alien pollen on native D. purpurea stigmata was very low and ranged from zero to nine pollen grains per stigma. We did not find any R. ponticum pollen on stigmata of D. purpurea in control sites. Alien pollen in relation to the total amount of pollen found on D. purpurea stigmata was positively related to R. ponticum abundance (Wald χ 2 = 62.25, df = 1, n total = 65, n units = 8, P < 0.001; βabundance = 0.057 ± 0.0072 SE; Fig. 3b).
Seed set
Digitalis purpurea fruit set was high in all sites (control, 96.5% ± 2.64; low R. ponticum abundance, 92.5% ± 2.54; high R. ponticum abundance, 100% ± 0). Individual fruits contained between 1 and 1,455 (estimated) seeds with an average of 739 ± 21.2 seeds per fruit. Neither fruit set nor the number of seeds per fruit were related to R. ponticum abundance (fruits per flower, Wald χ2 = 0.47, df = 1, ntotal = 48, nunits = 8, P = 0.494; Fig. 4; seeds per fruit, Wald χ2 = 3.66, df = 1, ntotal = 48, nunits = 8, P = 0.056).

Fruit set (geometric mean ± SE) per site in relation to relative abundance of Rhododendron ponticum. Note log scale used on y-axis
We detected low power (α = 0.094) for the analysis on seed set per fruit. However, even an increase in the number of sites by a factor of five (overall n = 45) would not have greatly increased the power of the experiments (α = 0.426).
Discussion
Rhododendron ponticum occupies various habitats in Ireland and often occurs in high abundance (Usher 1986). Here, we demonstrated that high abundance is related to differences in both the diversity of entomophilous plant communities as well as the pollination of a self-compatible native plant species, but that they ultimately did not influence its reproductive success.
Pollinator attraction is one of the first steps of several stages that a plant has to overcome in order to successfully produce offspring. Flower visitors to D. purpurea were more similar to R. ponticum visitors in the presence of the invader than in its absence, suggesting that the presence of R. ponticum modified the identity of D. purpurea flower visitors. Although the species similarity of flower visitors between R. ponticum and D. purpurea was smaller than has been observed for alien–native species pairs in other systems (Muñoz and Cavieres 2008), both species overlapped considerably in their visitors, which could foster direct interspecific interactions between the two plant species. If their generalist pollinators foraged randomly and did not distinguish between flowers of different species, individual visitation rates would not change across various relative densities of a species (Levin and Anderson 1970). In reality, pollinators assess and distinguish between individual rewards of each species (Hill et al. 2001), incorporating information on species’ flower complexity (Stout et al. 1998) as well as the relative abundance of each species (Feinsinger et al. 1991; Stout et al. 1998) and may refuse or prefer to visit a species beyond what is expected by its relative frequency. Therefore, it is not surprising that we observed lower visitation rates to D. purpurea in more invaded field sites. Studies on other invasive species revealed similar disruptive effects on native visitation at high alien abundance (Ghazoul 2004; Muñoz and Cavieres 2008; but see Grabas and Laverty 1999).
The decrease in visitation cannot be fully attributed to an interaction between relative frequency and reward levels or flower complexity of the two species because both R. ponticum and D. purpurea have relatively easily accessible flowers that secrete a similar amount of sugar (Percival and Morgan 1965; Mejías et al. 2002). Lower visitation rates to D. purpurea are more likely to be explained by flower preference and higher constancy of pollinators towards the relatively more abundant R. ponticum at sites of high abundance. Such a monopolisation would suggest the occurrence of interspecific competition and has been observed in other invasive species before (Chittka and Schürkens 2001; Ghazoul 2004).
Alternatively, D. purpurea at more invaded sites might have attracted fewer insect pollinators to its flowers because of its reduced relative abundance in the site. Plants are likely to suffer from pollinator limitation at low relative abundances due to decreased attractiveness to pollinators (Aizen and Harder 2007). Lower intraspecific facilitation among D. purpurea individuals is therefore a possible explanation for the observed pattern.
A third proposition is that flower visitation by bees can be positively associated with the diversity of flowering communities (Petanidou and Ellis 1996; but see Hegland and Boeke 2006) implying that D. purpurea might have encountered lower interspecific facilitation by other native species due to the reduced native entomophilous plant diversity at sites where R. ponticum was highly abundant.
Lastly, the presence of an invasive species can also impact pollinator communities, e.g. increase pollinator densities like other mass-flowering plants (Westphal et al. 2003) or reduce pollinator diversity (Moroń et al. 2009). Not only the decrease in flower visitation but also the observed general change in D. purpurea’s visitor assamblage in the presence of the invasive alien may be related to such impacts. Abundance, diversity and the identity of pollinators influence flower visitation and successful pollen deposition (Gómez et al. 2010) but may be differently affected by an invasive plant species. Studies on the whole plant and pollinator network are needed to test for such relationships.
We unfortunately cannot distinguish between the four mechanisms, i.e. between higher interspecific alien competition, lower intraspecific facilitation, lower interspecific native facilitation and changes in pollinator community, because the abundance and diversity of native plants in our sites was greatly affected by the abundance of the alien. In natural field sites, relative abundances of co-occurring species are often negatively correlated so that manipulative experiments are required to identify underlying mechanisms (Kunin 1997). Despite the ambiguity about the precise mechanism, which reduced visitation to D. purpurea inflorescences and flowers, this study shows that heavy invasion by R. ponticum may disrupt pollinator visitation to native plants.
Clearly, pollen transfer is a function of flower visitation rate (Wilcock and Neiland 2002), and the disrupted visitation rate is likely to impact pollen deposition in D. purpurea. Frequency of pollen transfer events (i.e. the quantity of visitation) alone is not necessarily a strong correlative of successful pollination if the composition of pollen loads (i.e. the quality of visitation) is variable (Rathcke 1983).
Considering that the two plant species used, in almost half the cases, the same pollinator species, we expected interspecific flower movement to occur. As a consequence, heterospecific pollen may be transferred to and interact with native stigmata (Morales and Traveset 2008). Although heterospecific pollen ratios increased with increasing R. ponticum abundance, we still found very little foreign pollen on native D. purpurea stigmata. This is in line with other studies that found low alien pollen deposition on native stigmata in natural communities (Larson et al. 2006; Bartomeus et al. 2008a; Jakobsson et al. 2008; but see Ghazoul 2002). Flower constancy exhibited by bees and hoverflies (Goulson and Wright 1998) was not measured in this study, but it might plausibly explain our observations at high R. ponticum abundance, when alien pollen should be highly abundant in the pollen transport web (Lopezaraiza-Mikel et al. 2007). Even if pollinators moved between heterospecific flowers, alien pollen deposition might be low due to segregated pollen placement on the pollinators’ bodies (mechanical isolation; Grant 1994). Mechanical isolation is likely because D. purpurea flowers place and collect pollen very precisely on the dorsal surface of most visiting insects (personal observation). R. ponticum pollen is more diffusely placed on the visitors’ bodies because the long viscin threads that adhesively join the pollen grain tetrads can easily stick to and drape over the insects’ bodies (King and Buchmann 1995). Furthermore, pollinator behaviour (e.g. grooming; Harder and Wilson 1998) may restrict placement and transfer of heterospecific pollen. Bumblebees, the main pollinators in our study system, often do not touch the reproductive parts of R. ponticum due to size constraints (Stout 2007) and therefore might not pick up large amounts of alien pollen. Due to the low heterospecific pollen loads on D. purpurea stigmata, it is unlikely that alien pollen interfered negatively with the fertilisation in D. purpurea individuals.
Increased conspecific pollen loss in the presence of an invasive species is thought to be one of the mechanisms for native pollination disruption (Morales and Traveset 2008). However, the proposed higher pollinator flower constancy at high alien abundance makes D. purpurea pollen loss rather unlikely. The decrease in conspecific D. purpurea pollen deposition with increasing R. ponticum abundance is probably directly related to lower visitation rates. Similarly, Ghazoul (2002) and Larson et al. (2006) observed a parallel decrease in native visitation rate and conspecific pollen deposition for some native species in different invaded communities (but see Grabas and Laverty 1999).
Nevertheless, conspecific pollen loads on D. purpurea stigmata were relatively high even in sites with lower visitation rates. Less frequently visited D. purpurea flowers held more nectar, and this surplus in rewards might have triggered a change in pollinator behaviour resulting in longer visits within each flower (Pappers et al. 1999), which can increase successful pollen transfer (Harder 1990). Muñoz and Cavieres (2008) showed that longer native visits at low local alien abundance were positively related to an increase in native seed set, implying higher native pollen deposition caused by longer visitation. Here, we did not measure visitation time at a flower level and so we cannot determine whether there was an alteration in pollinator behaviour. Native pollen deposition might also have been high because D. purpurea flowers offer large amounts of pollen (sixfold or more than other native species; Nienhuis 2009). This may increase pollen loads transferred and deposited by each pollinator so that even a single visit might be sufficient to fertilise the majority of ovules.
Visitation rates and pollen deposition are usually known to be good predictors for reproductive success (Waites and Ågren 2004; Kirchner et al. 2005). So why does D. purpurea fruit and seed set not mirror the differences in visitation and pollen deposition?
The power of our seed set analysis was very low so that we might have failed to detect a positive or negative effect of alien abundance on native seed set. In the majority of studies conducted to date, negative impacts of alien species on native seed set have been found (Chittka and Schürkens 2001; Brown et al. 2002). However, these studies have focused on self-incompatible plant species. In self-compatible plants, interspecific effects of abundance on reproductive success may be absent or weaker (Feinsinger et al. 1991; Caruso 1999) possibly because of the reproductive assurance these species experience through delayed spontaneous autogamy (Kalisz and Vogler 2003) or facilitated selfing. In our case, although D. purpurea is incapable of spontaneous autogamy, it is self-compatible (Nazir et al. 2008) and could have achieved within-flower or within-plant selfing (geitonogamy) by pollinators conducting longer visits per flower and/or visiting more flowers per inflorescence in less frequently visited plants. D. purpurea is mainly pollinated by bumblebees, i.e. pollinators that are able to fly long distances when foraging (Osborne et al. 2008). Thus, negative effects of low local conspecific abundance, e.g. pollen limitation or self-pollination, might be outweighed by the high mobility of pollen vectors that may import cross-pollen from more distant populations (Somanathan et al. 2004). Therefore, both increased selfing and high-quality cross-pollen transfer may explain the lack of impacts on seed set in our study system.
A high conspecific pollen/seed number ratio suggests that D. purpurea was not pollen limited. Still, average seed set was not only lower than the average seed set estimated by a previous study but was also half their value for maximum seed set (Buxton and Darlington 1932). Our results may therefore indicate limitation by factors other than pollen (e.g. water, nutrients or hosts) that do not correlate with the level of R. ponticum invasion. Such resource limitations can overshadow the abundant availability of pollen and so plants may not capitalise from higher pollen loads (Parker and Haubensak 2002; Totland et al. 2006).
Insect visitation has been described as a function of floral density at a habitat scale (100 m2 to 1 ha; Johnson et al. 2003) but fitness can also be related to abundance at a local scale (2–10 m radii; Kirchner et al. 2005). Hence, choosing a habitat scale might have concealed alien abundance effects on native reproduction that only operate within close neighbourhood (as in Muñoz and Cavieres 2008). R. ponticum invasion manifests at a habitat scale with single individuals covering on average 4 m2 (Erfmeier and Bruelheide 2004) and forming large clumps. In this field study, we were, therefore, more interested in alien impacts at a larger scale. However, manipulations of the abundance of R. ponticum and D. purpurea at a local scale also did not reveal any response of D. purpurea’s reproductive success to R. ponticum abundance (unpublished data).
Conclusions
Individuals of native species can display substantial plasticity that allows the integration of an alien species in the existing plant–pollinator web (Vilà et al. 2009). Although we did observe a disruptive effect of alien abundance on native flower visitation and conspecific pollen deposition, our study indicates that R. ponticum does not alter reproductive success of populations of the self-compatible D. purpurea. However, individuals of species with self-incompatible breeding systems or low pollen production might respond to decreased visitation rates and pollen deposition differently than D. purpurea (Ashman et al. 2004). This highlights the need for investigating not only pollinator visitation but also pollen deposition and seed set. Other factors such as pollen quality and quantity seem to be of equal importance for understanding the direct and indirect interactions between alien and native plant species pollination. These factors as well as native and alien species’ abundance should be integrated into further studies on impacts of invasion if we want to draw generalised conclusions.
References
Agresti A (1996) An introduction to categorical data analysis. Wiley, New York
Aizen MA, Harder LD (2007) Expanding the limits of the pollen-limitation concept: effects of pollen quantity and quality. Ecology 88:271–281
Ashman T-L, Knight TM, Steets JA, Amarasekare P, Burd M, Campbell DR, Dudash MR, Johnston MO, Mazer SJ, Mitchell RJ, Morgan MT, Wilson WG (2004) Pollen limitation of plant reproduction: ecological and evolutionary causes and consequences. Ecology 85:2408–2421
Bartomeus I, Bosch J, Vilà M (2008a) High invasive pollen transfer, yet low deposition on native stigmas in a Carpobrotus-invaded community. Ann Bot 102:417–424
Bartomeus I, Vilà M, Santamaría L (2008b) Contrasting effects of invasive plants in plant–pollinator networks. Oecologia 155:761–770
Bjerknes A-L, Totland Ø, Hegland SJ, Nielsen A (2007) Do alien plant invasions really affect pollination success in native plant species? Biol Conserv 138:1–12
Brown BJ, Mitchell RJ, Graham SA (2002) Competition for pollination between an invasive species (purple loosestrife) and a native congener. Ecology 83:2328–2336
Buxton BH, Darlington CD (1932) Crosses between Digitalis purpurea and Digitalis ambigua. New Phytol 31:225–240
Caruso CM (1999) Pollination of Ipomopsis aggregata (Polemoniaceae): effects of intra- vs. interspecific competition. Am J Bot 86:663–668
Caruso CM (2002) Influence of plant abundance on pollination and selection on floral traits of Ipomopsis aggregata. Ecology 83:241–254
Chittka L, Schürkens S (2001) Successful invasion of a floral market—an exotic Asian plant has moved in on Europe’s river-banks by bribing pollinators. Nature 411:653
Chittka L, Gumbert A, Kunze J (1997) Foraging dynamics of bumble bees: correlates of movements within and between plant species. Behav Ecol 8:239–249
Cross JR (1975) Rhododendron ponticum L. J Ecol 63:345–364
Erfmeier A, Bruelheide B (2004) Comparsion of native and invasive Rhododendron ponticum populations: growth, reproduction and morphology under field conditions. Flora 199:120–133
Feinsinger P, Murray KG, Kinsman S, Busby WH (1986) Floral neighborhood and pollination success in four hummingbird-pollinated cloud forest plant species. Ecology 67:449–464
Feinsinger P, Tiebout HM III, Young BE (1991) Do tropical bird-pollinated plants exhibit density-dependent interactions? Field experiments. Ecology 72:1953–1963
Feldman TS (2008) The plot thickens: does low density affect visitation and reproductive success in a perennial herb, and are these effects altered in the presence of a co-flowering species? Oecologia 156:807–817
Fossitt JA (2000) A guide to habitats in Ireland, 1st edn. The Heritage Council, Dublin
Ghazoul J (2002) Flowers at the front line of invasion? Ecol Entomol 27:638–640
Ghazoul J (2004) Alien abduction: disruption of native plant–pollinator interactions by invasive species. Biotropica 36:156–164
Ghazoul J (2006) Floral diversity and the facilitation of pollination. J Ecol 94:295–304
Gómez JM, Abdelaziz M, Lorite J, Jesús Muñoz-Pajares A, Perfectti F (2010) Changes in pollinator fauna cause spatial variation in pollen limitation. J Ecol 98:1243–1252
Goulson D, Wright NP (1998) Flower constancy in the hoverflies Episyrphus balteatus (Degeer) and Syrphus ribesii (L.) (Syrphidae). Behav Ecol 9:213–219
Grabas GP, Laverty TM (1999) The effect of purple loosestrife (Lythrum salicaria L.; Lythraceae) on the pollination and reproductive success of sympatric co-flowering wetland plants. Ecoscience 6:230–242
Grant V (1994) Modes and origins of mechanical and ethological isolation in angiosperms. Proc Natl Acad Sci USA 91:3–10
Harder LD (1990) Pollen removal by bumble bees and its implications for pollen dispersal. Ecology 71:1110–1125
Harder LD, Wilson WG (1998) Theoretical consequences of heterogeneous transport conditions for pollen dispersal by animals. Ecology 79:2789–2807
Hegland SJ, Boeke L (2006) Relationships between the density and diversity of floral resources and flower visitor activity in a temperate grassland community. Ecol Entomol 31:532–538
Hill PSM, Hollis J, Wells H (2001) Foraging decisions in nectarivores: unexpected interactions between flower constancy and energetic rewards. Anim Behav 62:729–737
Jakobsson A, Padrón B, Traveset A (2008) Pollen transfer from invasive Carpobrotus spp. to natives—a study of pollinator behaviour and reproduction success. Biol Conserv 141:136–145
Johnson SD, Peter CI, Nilsson LA, Ågren J (2003) Pollination success in a deceptive orchid is enhanced by co-occurring rewarding magnet plants. Ecology 84:2919–2927
Kalisz S, Vogler DW (2003) Benefits of autonomous selfing under unpredictable pollinator environments. Ecology 84:2928–2942
King MJ, Buchmann SL (1995) Bumble bee-initiated vibration release mechanism of Rhododendron pollen. Am J Bot 82:1407–1411
Kirchner F, Luijten SH, Imbert E, Riba M, Mayol M, González-Martínez SC, Mignot A, Colas B (2005) Effects of local density on insect visitation and fertilization success in the narrow-endemic Centaurea corymbosa (Asteraceae). Oikos 111:130–142
Knight ME, Martin AP, Bishop S, Osborne JL, Hale RJ, Sanderson RA, Goulson D (2005) An interspecific comparison of foraging range and nest density of four bumblebee (Bombus) species. Mol Ecol 14:1811–1820
Kunin WE (1997) Population size and density effects in pollination: pollinator foraging and plant reproductive success in experimental arrays of Brassica kaber. J Ecol 85:225–234
Larson DL, Royer RA, Royer MR (2006) Insect visitation and pollen deposition in an invaded prairie plant community. Biol Conserv 130:148–159
Levin DA, Anderson WW (1970) Competition for pollinators between simultaneously flowering species. Am Nat 104:455–467
Levine JM, Vilà M, D’Antonio CM, Dukes JS, Grigulis K, Lavorel S (2003) Mechanisms underlying the impacts of exotic plant invasions. Proc R Soc Lond B 270:775–781
Lopezaraiza-Mikel ME, Hayes RB, Whalley MR, Memmott J (2007) The impact of an alien plant on a native plant–pollinator network: an experimental approach. Ecol Lett 10:539–550
Mejías JA, Arroyo J, Ojeda F (2002) Reproductive ecology of Rhododendron ponticum (Ericaceae) in relict Mediterranean populations. Bot J Linn Soc 140:297–311
Moeller DA (2004) Facilitative interactions among plants via shared pollinators. Ecology 85:3289–3301
Morales CL, Traveset A (2008) Interspecific pollen transfer: magnitude, prevalence and consequences for plant fitness. Crit Rev Plant Sci 27:221–238
Morales CL, Traveset A (2009) A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol Lett 12:716–728
Moroń D, Lenda M, Skórka P, Szentgyörgyi H, Settele J, Woyciechowski M (2009) Wild pollinator communities are negatively affected by invasion of alien goldenrods in grassland landscapes. Biol Conserv 142:1322–1332
Muñoz AA, Cavieres LA (2008) The presence of a showy invasive plant disrupts pollinator service and reproductive output in native alpine species only at high densities. J Ecol 96:459–467
Nazir R, Reshi Z, Wafai BA (2008) Reproductive ecology of medicinally important Kashmir Himalayan species of Digitalis L. Plant Species Biol 23:59–70
Nienhuis CM (2009) The interactions between native bees and alien plants. PhD dissertation, Trinity College Dublin, Ireland
Osborne JL, Martin AP, Carreck NL, Swain JL, Knight ME, Goulson D, Hale RJ, Sanderson RA (2008) Bumblebee flight distances in relation to the forage landscape. J Anim Ecol 77:406–415
Palmer TM, Stanton ML, Young TP (2003) Competition and coexistence: exploring mechanisms that restrict and maintain diversity within mutualist guilds. Am Nat 162:S63–S79
Pappers SM, de Jong TJ, Klinkhamer PGL, Meelis E (1999) Effects of nectar content on the number of bumblebee approaches and the length of visitation sequences in Echium vulgare (Boraginaceae). Oikos 87:580–586
Parker IM, Haubensak KA (2002) Comparative pollinator limitation of two non-native shrubs: do mutualisms influence invasions? Oecologia 130:250–258
Parker IM, Simberloff D, Lonsdale WM, Goodell K, Wonham M, Kareiva PM, Williamson MH, Von Holle B, Moyle PB, Byers JE, Goldwasser L (1999) Impact: toward a framework for understanding the ecological effects of invaders. Biol Invasions 1:3–19
Percival M, Morgan P (1965) Observations on the floral biology of Digitalis species. New Phytol 64:1–22
Petanidou T, Ellis WN (1996) Interdependence of native bee faunas and floras in changing Mediterranean communities. In: Matheson A, Buchmann SL, O’Toole C, Westrich P, Williams IH (eds) The conservation of bees. Academic, London, pp 210–226
Pianka ER (1973) The structure of lizard communities. Annu Rev Ecol Syst 4:53–74
Quinn GP, Keough MJ (2002) Experimental design and data analysis for biologists, 1st edn. Cambridge University Press, Cambridge
Rathcke BJ (1983) Competition and facilitation among plants for pollination. In: Real LA (ed) Pollination biology. Academic, Orlando, pp 305–329
Richardson DM, Pyšek P, Rejmánek M, Barbour MG, Panetta FD, West CJ (2000) Naturalization and invasion of alien plants: concepts and definitions. Divers Distrib 6:93–107
Sakai AK, Allendorf FW, Holt JS, Lodge DM, Molofsky J, With KA, Baughman S, Cabin RJ, Cohen JE, Ellstrand NC, McCauley DE, O’Neil P, Parker IM, Thompson JN, Weller SG (2001) The population biology of invasive species. Annu Rev Ecol Syst 32:305–332
Sax DF, Stachowicz JJ, Brown JH, Bruno JF, Dawson MN, Gaines SD, Grosberg RK, Hastings A, Holt RD, Mayfield MM, O’Connor MI, Rice WR (2007) Ecological and evolutionary insights from species invasions. Trends Ecol Evol 22:465–471
Somanathan H, Borges RM, Chakravarthy VS (2004) Does neighborhood floral display matter? Fruit set in carpenter bee-pollinated Heterophragma quadriloculare and beetle-pollinated Lasiosiphon eriocephalus. Biotropica 36:139–147
Stang M, Klinkhamer PGL, van der Meijden E (2006) Size constraints and flower abundance determine the number of interactions in a plant–flower visitor web. Oikos 112:111–121
Stout JC (2007) Pollination of invasive Rhododendron ponticum (Ericaceae) in Ireland. Apidologie 38:198–206
Stout JC, Allen JA, Goulson D (1998) The influence of relative plant density and floral morphological complexity on the behaviour of bumblebees. Oecologia 117:543–550
Stout JC, Parnell JAN, Arroyo J, Crowe TP (2006) Pollination ecology and seed production of Rhododendron ponticum in native and exotic habitats. Biodivers Conserv 15:755–777
Thomson JD (1978) Effect of stand composition on insect visitation in two-species mixtures of Hieracium. Am Midl Nat 100:431–440
Totland Ø, Nielsen A, Bjerknes A-L, Ohlson M (2006) Effects of an exotic plant and habitat disturbance on pollinator visitation and reproduction in a boreal forest herb. Am J Bot 93:868–873
Traveset A, Richardson DM (2006) Biological invasions as disruptors of plant reproductive mutualisms. Trends Ecol Evol 21:208–216
Usher MB (1986) Invasibility and wildlife conservation: invasive species on nature reserves. Philos Trans R Soc Lond B 314:695–710
Vilà M, Bartomeus I, Dietzsch AC, Petanidou T, Steffan-Dewenter I, Stout JC, Tscheulin T (2009) Invasive plant integration into native plant–pollinator networks across Europe. Proc R Soc Lond B 276:3887–3893
Vitousek PM, D’Antonio CM, Loope LL, Rejmánek M, Westbrooks R (1997) Introduced species: a significant component of human-caused global change. NZ J Ecol 21:1–16
Waites AR, Ågren J (2004) Pollinator visitation, stigmatic pollen loads and among-population variation in seed set in Lythrum salicaria. J Ecol 92:512–526
Weidenhamer JD, Callaway RM (2010) Direct and indirect effects of invasive plants on soil chemistry and ecosystem function. J Chem Ecol 36:59–69
Westphal C, Steffan-Dewenter I, Tscharntke T (2003) Mass flowering crops enhance pollinator densities at a landscape scale. Ecol Lett 6:961–965
Wilcock C, Neiland R (2002) Pollination failure in plants: why it happens and when it matters. Trends Plant Sci 7:270–277
Wolf S, Moritz RFA (2008) Foraging distance in Bombus terrestris L. (Hymenoptera: Apidae). Apidologie 39:419–427
Yurkonis KA, Meiners SJ (2004) Invasion impacts local species turnover in a successional system. Ecol Lett 7:764–769
Acknowledgments
Thanks to Karl Duffy for field assistance and to Steven Waldren and the members of staff of the Trinity Botanic Gardens for technical support. We thank Coillte Ireland for access to field sites and usage permission, and Bill Kunin for helpful comments on the manuscript. This study was funded by Science Foundation Ireland as part of a Basic Research Grant (04/BR/B0637) and a UREKA Phytotechnology Ireland Summer School Grant, both awarded to J.C.S. This research complies with current Irish laws.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by Miguel Franco.
Rights and permissions
About this article
Cite this article
Dietzsch, A.C., Stanley, D.A. & Stout, J.C. Relative abundance of an invasive alien plant affects native pollination processes. Oecologia 167, 469–479 (2011). https://doi.org/10.1007/s00442-011-1987-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-011-1987-z