
Abstract
Variation in traits affecting seed dispersal in plants has been attributed to selection exerted by dispersal agents. The potential for such selection was investigated in Virola calophylla (Myristicaceae) in Manú National Park, Peru, through identification of seed dispersal agents and of tree and fruit traits significantly affecting the quantity of seeds dispersed. Seventeen bird and one primate species (the spider monkey, Ateles paniscus) dispersed its seeds. Spider monkeys dispersed the majority of seeds (a minimum of 83% of all dispersed seeds). Visitation by dispersal agents depended only on the quantity of ripe fruit available during a tree observation. In contrast, seed removal increased with both greater quantity of ripe fruit and aril: seed ratio. When analyzed separately, seed removal by birds increased only with greater aril: seed ratio, whereas seed removal by spider monkeys was affected by the quantity of ripe fruit and phenological stage. The finding that dispersal agents responded differently to some tree and fruit traits indicates not only that dispersal agents can exert selection on traits affecting seed dispersal, but also that the resulting selection pressures are likely to be inconsistent. This conclusion is supported by the result that the proportion of the seed crop that was dispersed from individual trees, which accounted for cumulative dispersal by all agents, was not influenced by any tree or fruit trait evaluated. Comparing these results with those from studies of V. sebifera and V. nobilis in Panama revealed that the disperser assemblages of these three Virola species were congruent in their similar taxonomic representation. In Panama the proportion of V. nobilis seed crop dispersed was related positively to aril: seed ratio and negatively to seed mass, a result not found for V. calophylla in Peru. The greater importance of dispersal by primates versus birds in V. calophylla, relative to V. nobilis, may explain this difference. Thus, variation in disperser assemblages at regional scales can be another factor contributing to inconsistency in disperser-mediated selection on plant traits.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Early investigations of the relationships between plants and the animals that disperse their seeds were guided by the hypothesis that plant reproductive traits affecting seed dispersal and the ecology and behavior of dispersal agents have been shaped through coevolution (Snow 1971; McKey 1975). With the accumulation of observations indicating that specialized, coevolved relationships between plants and dispersal agents were rare came the realization that many ecological and evolutionary factors intervene to make fine-tuned, reciprocal evolutionary change unlikely (Wheelwright and Orians 1982; Howe 1984; Herrera 1985). However, because seed dispersal is selected even if it is costly in terms of resource investment (Hamilton and May 1977; Comins et al. 1980), plants are expected to evolve mechanisms promoting dispersal. In doing so, plant traits affecting dispersal may change in response to selection by dispersal agents (e.g., Lord et al. 2002), provided that individual plants experience fitness consequences resulting from such selection. These fitness consequences are mediated by effects on the quantity and quality of seed dispersal, where seed dispersal quantity refers to the number of seeds dispersed and quality refers to the probability that a dispersed seed will produce a reproductive adult (sensu Schupp 1993). This study investigated the potential for disperser-mediated selection on plant traits affecting the quantity of seeds dispersed.
Visitation and seed removal can be influenced by many traits of fruiting plants, including local abundance of fruit resources or neighborhood effects (Manasse and Howe 1983; Sargent 1990), phenology (Thompson and Willson 1979; Stiles 1980; Eriksson and Ehrlen 1998), crop size (Howe and Vande Kerckhove 1979; Howe and Vande Kerckhove 1980; Howe and Vande Kerckhove 1981; Davidar and Morton 1986; Murray 1987; Foster 1990a; Gryj and Dominguez 1996; Korine et al. 2000), energetic and nutrient contents of fruits and their correlates (Howe and Vande Kerckhove 1980; Foster 1990a; Sallabanks 1993), and seed size (Howe and Vande Kerckhove 1980). Considerable variation, however, exists among dispersal agents in their responses to these factors, which can result in inconsistency in selection on reproductive traits influencing the quantity of seeds dispersed (Herrera 1985), thereby increasing the relative influence of non-adaptive processes, such as genetic drift, affecting trait variation. Inconsistent selection can also result if variation in these traits is constrained by processes unrelated to seed dispersal, such as germination (Howe and Richter 1982; Parciak 2002) and seed predation (Benkman 1995).
The hypothesis that dispersal agents exert selective pressure on reproductive traits affecting seed dispersal leads to two predictions. First, patterns of visitation and seed removal by dispersers should vary significantly with differences in reproductive traits among individual plants. This can be considered a behavioral response by dispersal agents. Second, a strong relationship should exist between reproductive traits and the proportion of all seeds dispersed from individual tree crowns during an entire fruiting season. This can be considered a measure of the cumulative behavioral responses of dispersal agents. These predictions represent only the prerequisites for selection, because a response to disperser-mediated selection on reproductive traits depends on both the fitness consequences of dispersal by particular agents and the heritability and genetic architecture of traits. I evaluated whether these predictions held in the dispersal system of Virola calophylla (Myristicaeae), a subcanopy nutmeg tree species of lowland forests in South America that produces vertebrate-dispersed, fleshy fruits.
Understanding the origins of plant-disperser interactions requires using a comparative approach that accounts for not only the ecological patterns across multiple closely related species, but, ideally, the evolutionary trends in those patterns, as well (Thompson 1999). Through such comparisons, the lability of plant-disperser relationships and the consistency of trends in these relationships replicated in different locations can be explored. Therefore, a second objective was to compare qualitatively the results from Peru to those for V. sebifera and V. nobilis on Barro Colorado Island (BCI) in Panama (Howe 1981; Howe and Vande Kerckhove 1981) using a similar field methodology. Species of Virola in general, and these three species in particular, share similar fruit morphology and adult size (Rodrigues 1980; van Roosmalen 1985a; Ribeiro et al. 1999), thus facilitating comparisons. The goal of the comparison was to evaluate the degree of similarity among these three Virola species in the composition of their disperser assemblages and in the relationships between tree and fruit traits and the quantity of seeds dispersed.
Materials and methods
Study site
This study was conducted from August 1999 through December 2000 at Cocha Cashu Biological Station (CCBS) in Manú National Park, Peru (11°54′S, 71°18′W, elevation ca. 400 m). The average annual rainfall is 2,000 mm, with most precipitation falling between October and April (Terborgh 1983). CCBS is situated in the floodplain of the Manú River and consists of a mosaic of mature and successional floodplain forest stands. Community-wide, fruit production has two phenological peaks annually, one near the beginning (November-December) and one near the middle (February) of the rainy season (Terborgh 1983). Manú National Park has experienced little impact from human development and exploitation, so that population densities of dispersal agents and plants are minimally influenced by human activities. This site has been described in detail in previous publications (Terborgh 1983; Foster 1990b; Gentry 1990; Terborgh et al. 1990).
Study species
Virola calophylla (Myristicaceae) is a common canopy tree in lowland moist forests of South America (Rodrigues 1980). At CCBS, V.calophylla begins to ripen fruit during the transition from the dry to wet seasons (mid-September) and continues through mid-December. Fruit of V. calophylla is similar to other Virola species, which are characterized by a bivalved, dark green capsule which opens upon ripening to expose a single arillate seed, the dispersible unit (diaspore; Rodrigues 1980; van Roosmalen 1985a; Ribeiro et al. 1999). The aril is bright red, lanceolate, and oily. Other species of Virola have been found to have high concentrations of lipids and secondary compounds (Howe and Vande Kerckhove 1981; Lemus and Castro 1989; Harrowven et al. 1998; Galetti et al. 2000). The seed comprises most of the volume of the diaspore.
Focal trees and frugivore observations
Fruiting V. calophylla individuals were located within the study area in mature floodplain and late successional forest habitats. Focal trees were selected to represent the range of crop sizes observed at the study site. Following the methodology of Howe (1981) and Howe and Vande Kerckhove (1981), observations of frugivores were conducted at focal trees during the hours from dawn to 11:00 a.m. and from 2:00 p.m. to dusk, and individual observation periods lasted 2–4 h. Observations sessions at each tree were distributed in a stratified random fashion to ensure that observations sampled the entire fruiting period of each individual tree. On some occasions, in order to follow spider monkeys (Ateles paniscus) for another study, observation sessions were terminated early when spider monkeys visited the tree (see below in Statistical analyses). Each focal tree (n=17) was observed for at least 30 h, for a total of 647 observation hours. Nine and ten trees were observed in 1999 and 2000, respectively, with two focal trees being observed in both years.
The following data were collected during each observation session. First, the quantity of available ripe fruit at the initiation of the observation session was estimated by counting the number of clusters of fruit that contained at least one ripe fruit (open capsule). Individual ripe fruits were easily viewed using binoculars. If visibility was too poor for ripe fruits to be seen at the beginning of the observation session, then a count was made when visibility had sufficiently improved. If a dispersal agent visited before a count was made, then the quantity of ripe fruit available was treated as missing data for that observation session in data analysis. There were 15 such cases among a total of 255 observation sessions. Second, for each individual frugivore that entered the focal tree, I recorded: (1) species, (2) times of arrival and departure, (3) the number of arillate seeds ingested, and (4) the number of seeds regurgitated or defecated. Spider monkeys commonly visited focal trees in groups. When more than one individual arrived and fed at a focal tree at the same time, it was not possible to collect data accurately on all of them. Therefore, all data were collected on one focal individual, and only arrival and departure times were collected for feeding, non-focal individuals.
Ripe fruits that were fresh and in good condition were collected from the ground below each focal tree or from fruit traps to obtain measurements of arillate seeds. Wet mass of the arillate seed and the seed alone were measured to the nearest 0.01 g using an electronic scale. The wet mass of aril was obtained by subtraction. The aril: seed ratio was calculated as aril divided by seed wet masses. At least five arillate seeds from each focal tree were measured. For two trees, I was not able to find five arillate seeds in adequate condition for measurement, and the sample size was two and three for these two trees.
At the end of the fruiting season in each year, each observation session was categorized phenologically according to the stage of fruit ripening of the focal tree on the day of the observation session. Stages of phenology that an observation session could fall into were the first, middle, or last third of the focal tree's fruiting period, based on the total number of days a tree had ripe fruit. These stages corresponded roughly to the beginning, peak, and ending of fruit ripening for each individual focal tree. Similarly, each focal tree was also categorized phenologically according to its timing of fruiting relative to the fruiting period of the V. calophylla population at CCBS (94 and 90 days in 1999 and 2000, respectively). Stages of phenology that a focal tree could fall into corresponded to the beginning, peak, and ending of fruit ripening of the V. calophylla population.
Fruit traps
Following the methods of Howe and Vande Kerckhove (1981), fruit traps were used to estimate crop size and proportion of seeds dispersed from individual trees. Fruit traps were made of fiberglass window screen with 1-mm mesh size and were 1 m2 in area. Traps were hung with twine from secure vegetation at a height of 1–1.5 m from the ground to prevent trampling from peccaries (Tayassu pecari and T. tajacu). Field tests showed that seeds and capsules did not bounce, fall, or wash out of traps. Traps were randomly placed underneath the crowns of all focal trees except for one (n=18) and sampled from 7.6–22.5% of the area under the crown (15.8±3.6%, x±SD), with a minimum of three traps placed under each tree. Traps were emptied every 4–8 days during the entire fruiting period of each focal tree, until no capsules or capsule valves were observed on the tree. The following data were collected from traps: (A) the total number of capsules without seeds, (B) the total number of capsules containing seeds, and (C) the total number of seeds. Crop size (the total number of seeds produced by a focal tree during its entire fruiting season) was estimated as the quantity (A+B) for all traps under a focal tree, divided by the proportion of the crown area sampled by traps. The number of seeds dispersed from a focal tree was estimated as the difference between the crop size and the number of seeds falling below the tree, which was estimated as the quantity (B+C) for all traps under a focal tree, divided by the proportion of the crown area sampled by traps. The proportion of seeds dispersed was calculated as the number of seeds dispersed divided by the crop size of the focal tree.
Statistical analyses
Data were analyzed with SAS (2000). The objective of statistical analyses was to identify tree and fruit traits that influenced the quantity of seeds dispersed in terms of the activity of individual dispersal agents and in terms of the proportion of all seeds dispersed cumulatively by all agents. I tested whether the quantity of seeds dispersed was affected by the following five traits, which have been demonstrated to affect seed dispersal (see Introduction): (1) crop size, (2) seed mass, (3) aril: seed ratio, (4) phenological stage, and (5) the quantity of ripe fruit available on a tree at the time of the observation session. These traits were independent variables in the statistical analyses described below. Weak or non-significant correlations among the variables (tested using Spearman rank and Pearson correlation tests) indicated that multicollinearity was unlikely. For example, the quantity of ripe fruit available during an observation and crop size are only weakly correlated in part because Virola species ripen their fruits over an extended time period. Hence, even a tree with a large crop size may display few or many ripe fruit at any given time.
Three linear models were used. The first and second addressed the quantity of seeds dispersed with respect to the behavioral responses of individual dispersal agents as (1) disperser visitation, or the probability of a focal tree receiving a feeding visit during an observation session (i.e., a session in which at least one seed was ingested) and (2) seed removal, or the number of seeds that a dispersal agent removed during a visit. Because observation sessions were not of equal length, only data based on the first visit to occur during an observation session were used. The first model was a generalized linear model (the genmod procedure in SAS) with a binomial distribution and logit link function, which was used to test whether the probability of visitation to a focal tree depended on the five tree and fruit traits. The dependent variable, visitation, could take values of zero (no visit during the observation session) or one (at least one visit during the observation session). The second model was a mixed model (the mixed procedure in SAS), which was used to test whether seed removal (the number of seeds removed by a disperser during a feeding visit) at a focal tree depended on the five tree and fruit traits. Patterns in disperser visitation and seed removal were analyzed using (1) all data combined, (2) data for all avian dispersal agents combined and (3) data for all primate dispersal agents combined.
The third model addressed proportion of seeds dispersed by all agents cumulatively during the entire fruiting period of each focal tree. The dependent variable was the proportion of the seed crop dispersed as estimated from fruit traps (hereafter, proportion of seeds dispersed). The mixed procedure in SAS was used to test whether the proportion of seeds dispersed was significantly influenced by (1) crop size, (2) seed mass, (3) aril: seed ratio, and (4) phenological stage of each focal tree.
In all statistical analyses, the assumption of homoscedasticity was met, and transformations were used when necessary to achieve normality of residuals. The full model with main effects and interactions was tested using Type III hypothesis tests (Shaw and Mitchell-Olds 1993), and insignificant terms were pooled into error. Two trees were sampled in both years. Therefore, for analyses involving the mixed procedure in SAS, "year" was included in the model as a random effect. For analyses using the genmod procedure in SAS, "tree" was considered nested within "year" to account for this sampling structure.
Results
Trees differed significantly in seed mass, aril: seed ratio, seed length, and seed width in both years, and most variation in these traits was attributable to among-tree, rather than within-tree variation (Table 1). Diameter at breast height was weakly correlated with crop size (Pearson r=0.37, P=0.0354, n=33).
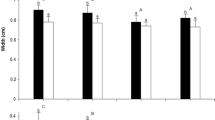
Seventeen bird species and one primate species were observed to disperse seeds of Virola calophylla in Peru (Table 2), relative to seven bird species for V. sebifera (Howe 1981) and seven bird species and one primate species for V. nobilis (Howe and Vande Kerckhove 1981; Fig. 1) in Panama. In Peru the spider monkey (Ateles paniscus) was the most frequent visitor and also removed the greatest number of seeds per visit (Table 2, Fig. 1a). Summed over all observations, spider monkeys removed 83% of all seeds observed to be removed (Table 2, Fig. 1c). This estimate is a minimum value because it only accounts for the seeds dispersed by the one focal individual out of a group of feeding spider monkeys that could be observed per visit. If the rate of seed ingestion by the focal individual is applied to all feeding members of the group, then approximately 92% (2,136 seeds) of dispersed seeds were dispersed by spider monkeys. In contrast, spider monkeys (A. geoffroyi) in Panama were not observed to disperse V. sebifera (Fig. 1b, d; Howe 1981). Although spider monkeys in Panama visited and removed seeds of V. nobilis (Fig. 1b, d; Howe and Vande Kerckhove 1981), they did so to a much lesser extent relative to spider monkeys at V. calophylla in Peru (Fig. 1a, c).

Visitation (a, b) and seed removal (c, d) by seed dispersers of Virola calophylla at Cocha Cashu Biological Station, Peru (a, c), and of V. sebifera and V. nobilis at Barro Colorado Island, Panama (b, d). Visitation includes non-feeding and feeding visits. For Panama black and grey shading in bars depict data for V. sebifera and V. nobilis, respectively. Data for V. sebifera and V. nobilis are from Howe (1981) and Howe and VandeKerckhove (1981), respectively. Bars in c and d, SD. Note changes in scales
In terms of individual responses of dispersal agents, the only trait that significantly affected the probability of visitation at V. calophylla was the quantity of ripe fruit available on the focal tree at the time of the observation session (χ2=5.98, df=1, P=0.0145). The probability of visitation was higher when more ripe fruits were available. Seed removal at V. calophylla was significantly influenced by both the availability of ripe fruit [F(1, 78)=11.08, P=0.0013] and the aril: seed ratio [F(1, 17)=9.05, P=0.0079]. Greater numbers of seeds were removed when there was more ripe fruit available and at trees with a higher ratio of aril: seed mass.
The visitation and seed removal data for V. calophylla were analyzed in separate models for all bird species combined versus for spider monkeys alone in order to investigate whether these dispersal agents respond differently to tree and fruit traits. Both spider monkeys and birds tended to have a higher probability of visitation with increasing availability of ripe fruits, but this effect was statistically significant only for birds (spider monkeys, χ 2=2.61, df=1, P=0.1062; birds, χ 2=6.77, df=1, P=0.0093). Birds removed more seeds per visit at trees with a higher aril: seed ratio [F(1, 15)=5.73, P=0.0303]. The number of seeds removed by birds per visit increased with increasing availability of ripe fruit, but only during a tree's early fruit-ripening period [significant interaction of phenology and ripe fruit availability; F(3, 54)=4.86, P=0.0369; slope of seed removal versus ripe fruit availability for early period, b=0.09, P=0.0004]. In contrast, spider monkeys removed more seeds when there was greater ripe fruit availability [F(1, 17)=6.86, P=0.0180], but they did not remove more seeds at trees with higher aril: seed ratio. Spider monkeys, however, did respond to phenological stage of the focal tree [F(2, 17)=4.84, P=0.0217]. They removed significantly more seeds per visit during the middle third of a tree's fruiting period than during the first third (differences of least squares means, t=−3.07, P=0.0070).
Frugivores removed 24–90% of seeds in the crops of V. calophylla trees, a range consistent with removal at V. nobilis in Panama (Howe and Vande Kerckhove 1981). In terms of the proportion of all seeds dispersed, there was no significant relationship between any of the tree and fruit traits and the proportion of seeds dispersed from focal trees. In contrast, seed dispersal at V. nobilis in Panama had a significant positive relationship with aril: seed ratio and a significant negative relationship with seed size (Howe and Vande Kerckhove 1980, 1981).
Discussion
This investigation found that avian and primate dispersal agents responded differently to tree and fruit traits in selecting the fruits they ingested, and, hence, the seeds that they dispersed. Although both spider monkeys and birds removed more Virola calophylla seeds when there were more ripe fruits available on a tree, only avian dispersal agents removed more seeds from trees with a higher aril: seed ratio. Spider monkeys appeared to respond exclusively to the availability of ripe fruit, as they removed more seeds not only when there were more ripe fruits on a tree, but also during the peak of a tree's fruiting period, when more fruits were ripened per day on a tree. Thus, these results suggest that in V. calophylla in Peru, selection on some plant traits varied among different classes of dispersal agents (i.e., birds versus primates) and may result in substantially weaker selection on some reproductive traits affecting seed dispersal.
Inconsistent selection among different dispersal agents is one factor that may contribute to the observed lack of specialized plant-disperser relationships (Wheelwright and Orians 1982; Howe 1984; Herrera 1985). A multi-site study of avian fruit preferences in Puerto Rican forests found that for many plant species, the identity of the disperser removing the most seeds changed across sites, thereby reducing the overall selective impact of frugivores (Carlo et al. 2003). In V. calophylla, the result that the proportion of all seeds dispersed was not significantly related to any tree or fruit trait evaluated also supports the hypothesis that plant-disperser interactions are too diffuse to result in strong selection on any one plant trait affecting seed dispersal.
Geographic comparisons of Virola species
Detailed comparative investigations of the dispersal systems of closely related plant species in different geographic locations are few (but see Howe 1977; Howe and Vande Kerckhove 1979; Bleher and Bohning-Gaese 2001). Such comparative studies, however, are important if we are to understand the origins of plant-disperser relationships and the potential for coevolution to have shaped them (Jordano 1995). In particular, the species pool of potential dispersal agents, as well as the behavior of particular disperser species, may vary at multiple spatial scales (Gautier-Hion et al. 1993; Bleher and Bohning-Gaese 2001; Chapman and Chapman 2002), contributing to geographic variation in plant-disperser relationships.
Disperser assemblages
Consistent with their similarities in fruit morphology, the species compositions of the disperser assemblages of the three Virola species were broadly congruent. Among the three disperser assemblages, seven bird families and one primate family were represented. In general, in each family, species that are allopatric replacements of each other, and hence ecologically similar, were represented as dispersers of the three Virola species. The only two exceptions were the Motmotidae (motmots), which did not visit V. calophylla in Peru, and the Icteridae (caciques), which did not visit Virola species in Panama. The greater number of bird species in the disperser assemblage of V. calophylla reflects the overall greater avian species richness of Peruvian, relative to Panamanian, forests (Terborgh et al. 1990; Robinson et al. 2000).
Despite similar disperser assemblages, the Virola species in Panama and Peru differ greatly in the importance of dispersal by birds versus primates. Visitation by birds, especially Ramphastids (toucans and araçaris) and Trogonids (trogons), was more frequent in both Panamanian congeners, compared to V. calophylla, which received the most frequent visitation by A. paniscus. Seed removal per visit, as well as total seed removal at V. calophylla, was highest by A. paniscus. In contrast, seed removal by birds accounted for the majority of seed dispersal for V. sebifera and V. nobilis. A parallel pattern exists for the tree, Casearia corymbosa (Flacourtiaceae), in that disperser assemblages in Costa Rican dry and wet forests were similar at higher taxonomic levels, but differed in the identity of the seed disperser dispersing the majority of seeds (Howe 1977; Howe and Vande Kerckhove 1979).
The difference among these three Virola species in the relative importance of seed dispersal by birds versus primates is likely to be explained in part by the fact that population densities of A. geoffroyi on BCI have been depressed since before 1923 because of extirpation by hunting (Milton 1993). Following reintroduction to BCI in 1959–1961, there was one group of 12 spider monkeys on BCI in 1974 (approximate population density assuming use of the entire island, 0.8 individuals / km2; Milton 1993). In contrast, estimated population density of A. paniscus at CCBS in 1976–1977 was 25 individuals / km2 (Terborgh 1983). More recent investigations of A. geoffroyi on BCI have shown that population densities have increased and confirm the dietary importance of V. nobilis for these monkeys (Campbell 2000).
The comparisons among V. calophylla, V. sebifera, and V. nobilis demonstrated that one important difference, the relative importance of primates versus birds, in their disperser assemblages contributed to variation in the potential for selection on plant traits by dispersers. These results suggest that disperser-mediated selection on traits of congeneric plants can vary among locations. The difference between Peru and Panama in the importance of seed mass and aril: seed ratio on the proportion of all seeds dispersed is likely to be related to the difference in the importance of dispersal by primates versus birds in Peru versus Panama. In Peru, the result that removal of V. calophylla seeds by birds was significantly higher with greater aril: seed ratio is consistent with the findings for V. nobilis in Panama, whereas no such relationship existed for A. paniscus. Unlike V. nobilis in Panama, seed mass did not significantly influence visitation or seed removal in V. calophylla in Peru, suggesting that spider monkeys may be less constrained than birds while foraging to minimize the quantity of ballast ingested. In addition, because seed mass of V. nobilis is approximately three times greater than that of V. calophylla, avian dispersal agents may be even more biased towards selecting individuals producing smaller seeds in V. nobilis than in V. calophylla.
Evolutionary perspectives
At a larger geographic scale, the importance of spider monkeys to dispersal of V. calophylla in Peru is consistent with studies in other South American sites in Surinam (A. paniscus feeding on V. melionii; van Roosmalen 1985b), French Guiana (A. paniscus and V. sp. nov.; Forget and Sabatier 1997), Ecuador (A. belzebuth and V. sp.; Dew 2001), French Guiana (A. paniscus and V. michelli; Guillotin et al. 1994), and Colombia (A. belzebuth and V. flexuosa; Stevenson et al. 2000). Toucans were also reported to disperse Virola species elsewhere in South America (V. oleifera and V. garneri in Brazil; Galetti et al. 2000 and V. surinamensis in Guyana; Bourne 1974). The similarities between the species compositions of the disperser assemblages of the three Virola species in Panama and Peru, supported by these corroborating observations for other Virola species, suggest that the similar traits of Virola trees and fruits attract similar disperser assemblages, even across large geographic distances.
One simple explanation for such similar disperser assemblages is that these relationships characterized ancestral species of Virola and have been retained throughout the diversification of the group. An alternative explanation is that the diversification of Virola predated the appearance of the bird and primate species that now disperse its seeds (sensu Janzen and Martin 1982). Similar responses to Virola tree and fruit traits on the part of dispersal agents in different geographic locations may therefore explain the similar disperser assemblages presently observed. Further investigations using phylogenetic comparative methods to examine the rates of character evolution in traits relevant for seed dispersal in relation to disperser assemblages of Virola species, as well as the sister clade to Virola, are needed to resolve the origins of the similar disperser assemblages that appear to characterize the extant Virola species studied to date.
This study has focused on the ability of dispersal agents to exert selection on plant traits by affecting the quantity of seeds dispersed. However, a complete understanding of a plant's response to selection requires additional information. First, knowledge of the heritability and genetic architecture of the trait in question, as well as the genetic characteristics of the population, are necessary (Howe 1984; Falconer 1991). Data are lacking on heritability of reproductive traits in tropical trees, but evidence from agricultural and silvicultural species suggests that some traits affecting seed dispersal, such as those examined here, have moderate to high heritability (Giannini and Bellari 1995; Matziris 1998; Yao and Mehlenbacher 2000; Hardner et al. 2001). Finally, understanding the quality of seed dispersal (Schupp 1993) in terms of per capita seed survival and seedling establishment, is critical to evaluating the fitness consequences for seeds dispersed by particular dispersal agents, and hence the potential outcome of that agent's activity in terms of selection on reproductive traits.
References
Benkman CW (1995) The impact of tree squirrels (Tamiasciurus) on limber pine seed dispersal adaptations. Evolution 49:585–592
Bleher B, Bohning-Gaese K (2001) Consequences of frugivore diversity for seed dispersal, seedling establishment, and the spatial pattern of seedlings and trees. Oecologia 129:385–394
Bourne GR (1974) The red-billed toucan in Guyana. The Living Bird 13:99–126
Campbell CJ (2000) The reproductive biology of black-handed spider monkeys (Ateles geoffroyi): Integrating behavior and endocrinology. Ph.D. thesis, University of California, Berkeley
Carlo TA, Collazo JA, Groom MJ (2003) Avian fruit preferences across a Puerto Rican forested landscape: pattern consistency and implications for seed removal. Oecologia 134:119–131
Chapman CA, Chapman LJ (2002) Plant-animal coevolution: is it thwarted by spatial and temporal variation in animal foraging? In: Levey DJ, Silva WR, Galetti M (eds) Seed dispersal and frugivory: ecology, evolution, and conservation. CABI, New York, pp 275–290
Comins HN, Hamilton WD, May RM (1980) Evolutionarily stable dispersal strategies. J Theor Biol 82:205–230
Davidar P, Morton ES (1986) The relationship between fruit crop sizes and fruit removal rates by birds. Ecology 67:262–265
Dew JL (2001) Synecology and seed dispersal in woolly monkeys (Lagothrix lagotricha poeppigii) and spider monkeys (Ateles belzebuth belzebuth) in Parque Nacional Yasuní, Ecuador. Ph.D. thesis, University of California, Davis
Eriksson O, Ehrlen J (1998) Phenological adaptations in fleshy vertebrate-dispersed fruits of temperate plants. Oikos 82:617–621.
Falconer DS (1991) Quantitative genetics. Longman, New York
Forget P, Sabatier D (1997) Dynamics of the seedling shadow of a frugivore-dispersed tree species in French Guiana. J Trop Ecol 13:767–773
Foster MS (1990a) Factors influencing bird foraging preferences among conspecific fruit trees. Condor 92:844–854
Foster RB (1990b) The floristic composition of the Rio Manú Floodplain Forest. In: Gentry AH (ed) Four neotropical forests. Yale University Press, New Haven, pp 99–111
Galetti M, Laps R, Pizo MA (2000) Frugivory by toucans (Ramphastidae) at two altitudes in the Atlantic forest of Brazil. Biotropica 32:842–850
Gautier-Hion A, Gautier J-P, Maisels F (1993) Seed dispersal versus seed predation: an intersite comparison of two related African monkeys. Vegetatio 107/108:237–244
Gentry AH (1990) Four neotropical forests. Yale University Press, New Haven
Giannini R, Bellari C (1995) Heritability estimate of seed germination parameters in Pinus leucodermis Antoine. Seed Sci Technol 23:385–392
Gryj EO, Dominguez GA (1996) Fruit removal and postdispersal survivorship in the tropical dry forest shrub Erythroxylum havanense: ecological and evolutionary implications. Oecologia 108:368–374
Guillotin M, Dubost G, Sabatier D (1994) Food choice and food competition among the three major primate species of French Guiana. J Zool 233:551–579
Hamilton WD, May RM (1977) Dispersal in stable habitats. Nature 269:578–581
Hardner C, Winks C, Russ S, Gallagher E (2001) Genetic parameters for nut and kernel traits in macadamia. Euphytica 117:151–161
Harrowven DC, Newman NA, Knight CA (1998) On the identity of a neo-lignan from the fruits of Virola sebifera. Tetrahedron Lett 39:6757–6760
Herrera CM (1985) Determinants of plant-animal coevolution: the case of mutualistic dispersal of seeds by vertebrates. Oikos 44:132–141
Howe HF (1977) Bird activity and seed dispersal of a tropical wet forest tree. Ecology 58:539–550
Howe HF (1981) Dispersal of a neotropical nutmeg (Virola sebifera) by birds. Auk 98:88–98
Howe HF (1984) Constraints on the evolution of mutualisms. Am Nat 123:764–777
Howe HF, Richter WM (1982) Effects of seed size on seedling size in Virola surinamensis; a within and between tree analysis. Oecologia 53:347–351
Howe HF, Vande Kerckhove GA (1979) Fecundity and seed dispersal of a tropical tree. Ecology 60:180–189
Howe HF, Vande Kerckhove GA (1980) Nutmeg dispersal by tropical birds. Science 210:925–927
Howe HF, Vande Kerckhove GA (1981) Removal of wild nutmeg (Virola surinamensis) crops by birds. Ecology 62:1093–1106
Janzen DH, Martin PS (1982) Neotropical anachronisms: The fruits the gomphotheres ate. Science 215:19–27
Jordano P (1995) Angiosperm fleshy fruits and seed dispersers: a comparative of adaptation and constraints in plant-animal interactions. Am Nat 145:163–191
Korine C, Kalko EKV, Herre EA (2000) Fruit characteristics and factors affecting fruit removal in a Panamanian community of strangler figs. Oecologia 123:560–568
Lemus SM, Castro O (1989) Potential antifungal neolignans from Virola koschnyi fruits. Spectrosc Int J 7:353–358
Lord JM, Markey AS, Marshall J (2002) Have frugivores influenced the evolution of fruit traits in New Zealand? In: Levey DJ, Silva WR, Galetti M (eds) Seed dispersal and frugivory: ecology, evolution, and conservation. CABI, New York, pp 55–68
Manasse RS, Howe HF (1983) Competition for dispersal agents among tropical trees: influences of neighbors. Oecologia 59:185–190
Matziris D (1998) Genetic variation in cone and seed characteristics in a clonal seed orchard of Aleppo pine grown in Greece. Silvae Genet 47:37–41
McKey D (1975) The ecology of coevolved seed dispersal systems. In: Gilbert LE, Raven PH (eds) Coevolution of animals and plants. University of Texas, Austin, pp 159–191
Milton K (1993) Diet and social organization of a free-ranging spider monkey population: the development of species-typical behavior in the absence of adults. In: Pererira ME, Fairbanks LA (eds) Juvenile primates: life history, development, and behavior. Oxford University Press, Oxford, pp 136–144
Murray KG (1987) Selection for optimal fruit crop size in bird-dispersed plants. Am Nat 129:18–31
Parciak W (2002) Seed size, number, and habitat of a fleshy-fruited plant: consequences for seedling establishment. Ecology 83:794–808
Ribeiro JELS, Hopkins MJG, Vicentini A, Sothers CA, Costa MAS, de Brito JM, de Souza MAD, et al (1999) Flora da Reserva Ducke: Guia de intentificação das plantas vasculares de uma floresta de terra-firme na Amazônia Central. INPA-DFID, Manaus
Robinson WD, Brawn JD, Robinson SK (2000) Forest bird community structure in central Panama: Influence of spatial scale and biogeography. Ecol Monogr 70:209–235
Rodrigues WA (1980) Revisao taxonomica das especies de Virola Aublet (Myristicaceae) do Brasil. Acta Amazon 10:1-127
Sallabanks R (1993) Hierarchical mechanisms of fruit selection by an avian frugivore. Ecology 74:1326–1336
Sargent S (1990) Neighborhood effects on fruit removal by birds: a field experiment with Viburnum dentatum (Caprifoliaceae). Ecology 71:1289–1298
SAS (2000) The SAS System Release 8.1. SAS Institute, Cary, N.C.
Schupp EW (1993) Quantity, quality, and the effectiveness of seed dispersal by animals. Vegetatio 107/108:15–29
Shaw RG, T Mitchell-Olds (1993) ANOVA for unbalanced data: an overview. Ecology 74:1638–1645
Snow DW (1971) Evolutionary aspects of fruit-eating by birds. Ibis 113:194–202
Stevenson PR, Quinones MJ, Ahumada JA (2000) Influence of fruit availability on ecological overlap among four neotropical primates at Tinigua National Park, Colombia. Biotropica 32:533–544
Stiles EW (1980) Patterns of fruit presentation and seed dispersal in bird-disseminated woody plants in the eastern deciduous forest. Am Nat 116:670–688
Terborgh J (1983) Five new world primates. Princeton University Press, Princeton, N.J.
Terborgh J, Robinson SK, Parker TA III, Munn C, Pierpont N (1990) Structure and organization of an Amazonian forest bird community. Ecol Monogr 60:213–238
Thompson JN (1999) Specific hypotheses on the geographic mosaic of coevolution. Am Nat 153:S1-S14
Thompson JN, Willson MF (1979) Evolution of temperate fruit/bird interactions: phenological strategies. Evolution 33:973–982
van Roosmalen MGM (1985a) Fruits of the Guianan Flora. Institute of Systematic Botany, Utrecht University, and Silvicultural Department of Wageningen Agricultural University, Wageningen
van Roosmalen MGV (1985b) Habitat preferences, diet, feeding strategy, and social organization of the black spider monkey (Ateles paniscus paniscus Linnaeus 1758) in Surinam. Acta Amazon 15:1-238
Wheelwright NT, Orians GH (1982) Seed dispersal by animals: contrasts with pollen dispersal, problems of terminology, and constraints on coevolution. Am Nat 119:402–413
Yao Q, Mehlenbacher SA (2000) Heritability, variance components and correlation of morphological and phenological traits in hazelnut. Plant Breeding 119:369–381
Acknowledgements
This research was supported by a Pre-doctoral Fellowship from the National Science Foundation, a Research Award from the American Ornithologists' Union, and a Clark Research Grant and a Dissertation Travel Grant from the University of Illinois. The Instituto Nacionál de Recursos Naturales in Peru graciously granted permission to conduct this research. I thank my graduate committee, Carol K. Augspurger, Scott K. Robinson, George O. Batzli, Jeffrey D. Brawn, and Stephen Portnoy for their mentorship. Jeffrey D. Brawn, Mercedes S. Foster, Henry F. Howe, and Thomas J. Near provided stimulating discussions of this research. I thank Mark Gardiner, Luis Alza Leon, Timothy Paine, and Edwin Salazar Zapata for tireless assistance in the field. Susanne Aref provided important help with statistical analyses. I thank Thomas J. Near for his support and encouragement.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Russo, S.E. Responses of dispersal agents to tree and fruit traits in Virola calophylla (Myristicaceae): implications for selection. Oecologia 136, 80–87 (2003). https://doi.org/10.1007/s00442-003-1239-y
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00442-003-1239-y