
Abstract
A genetic basis of congenital isolated hypogonadotropic hypogonadism (CHH) can be defined in almost 50% of cases, albeit not necessarily the complete genetic basis. Next-generation sequencing (NGS) techniques have led to the discovery of a great number of loci, each of which has illuminated our understanding of human gonadotropin-releasing hormone (GnRH) neurons, either in respect of their embryonic development or their neuroendocrine regulation as the “pilot light” of human reproduction. However, because each new gene linked to CHH only seems to underpin another small percentage of total patient cases, we are still far from achieving a comprehensive understanding of the genetic basis of CHH. Patients have generally not benefited from advances in genetics in respect of novel therapies. In most cases, even genetic counselling is limited by issues of apparent variability in expressivity and penetrance that are likely underpinned by oligogenicity in respect of known and unknown genes. Robust genotype–phenotype relationships can generally only be established for individuals who are homozygous, hemizygous or compound heterozygotes for the same gene of variant alleles that are predicted to be deleterious. While certain genes are purely associated with normosmic CHH (nCHH) some purely with the anosmic form (Kallmann syndrome—KS), other genes can be associated with both nCHH and KS—sometimes even within the same kindred. Even though the anticipated genetic overlap between CHH and constitutional delay in growth and puberty (CDGP) has not materialised, previously unanticipated genetic relationships have emerged, comprising conditions of combined (or multiple) pituitary hormone deficiency (CPHD), hypothalamic amenorrhea (HA) and CHARGE syndrome. In this review, we report the current evidence in relation to phenotype and genetic peculiarities regarding 60 genes whose loss-of-function variants can disrupt the central regulation of reproduction at many levels: impairing GnRH neurons migration, differentiation or activation; disrupting neuroendocrine control of GnRH secretion; preventing GnRH neuron migration or function and/or gonadotropin secretion and action.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Congenital hypogonadotropic hypogonadism (CHH) is characterized by deficient secretion or action of gonadotropin-releasing hormone (GnRH)—in the absence of any other pituitary hormone deficiencies, or structural parasellar lesions—resulting in failure of gonadal hormone secretion and gametogenesis. By contrast, the majority of acquired diseases affecting the pituitary or hypothalamus—whether invasive, infiltrative, ischemic, traumatic, irradiational, inflammatory or metabolic—result in multiple hormone deficits. Once defined as “idiopathic”, a genetic cause is now apparent in almost 50% of CHH cases (Boehm et al. 2015; Young et al. 2019).
In respect of hypogonadotropic hypogonadism (HH) occurring as part of congenital combined pituitary hormone deficiency (CPHD), with or without non-endocrine anomalies, the aetiology is much less well understood and our knowledge is evolving more slowly, with over 90% of cases having no known genetic basis. However, alongside the classical CPHD genes encoding transcription factors central to embryonic pituitary cell fate specification, several CHH-associated genes have recently been found to underpin CPHD-associated HH (Raivio et al. 2012; Jayakody et al. 2012; McCabe et al. 2013; Izumi et al. 2014; McCormack et al. 2017).
Loss-of-function variants in CHH-associated genes can disrupt the central neuroendocrine regulation of reproduction at key points of vulnerability, comprising disrupted migration; differentiation or activation of GnRH neurons; disrupted neuroendocrine control of GnRH secretion; resistance to GnRH action; and primary impairment of gonadotropin secretion or action. In this review, we will examine the genetic basis of CHH and the lessons contained therein relating to human reproductive physiology and developmental biology.
Epidemiology
Although widely considered to be a rare disease, ascertainment of its true prevalence is constrained by the scarcity of published literature. An historic study of Frenchmen called up for military service gave a male CHH prevalence of 1 in 4415 (Fromantin et al. 1973), but more recently, a Finnish retrospective study of hospital records gave a lower prevalence (Laitinen et al. 2011). While considered to be a male-predominant condition, with a reported male: female gender ratio of 3:6 in a study based on patients attending specialist centres (Dzemaili et al. 2017), this is not supported by our current understanding of the genetics implicated, wherein only 3.5–10% (according to the population) of unrelated CHH males harbour ANOS1 mutations characteristic of X-linked inheritance (Sykiotis et al. 2010; Laitinen et al. 2011; Hanchate et al. 2012; Basaran et al. 2013; Tommiska et al. 2014; Stamou et al. 2019). The apparent male excess may reflect ascertainment bias, due to a greater likelihood of females being managed empirically by office- or community-based gynaecologists or primary care physicians. Indeed, closer examination of CHH kindreds (excluding those with ANOS1 mutations) has found a sex ratio among affected individuals within kindreds that approaches parity (Francou et al. 2016; Maione et al. 2018).
Notably, significant phenotypic and genetic overlap between CHH and combined pituitary hormone deficiency (CPHD—resulting from dysregulated morphogenesis of pituitary gland) has been observed. CPHD has an estimated prevalence of around 1 in 8000 (Fang et al. 2016b), although the full spectrum of pituitary hormonal deficiencies may evolve over a variable duration of up to decades, not necessarily being able to manifest in its entirety at birth or in early childhood.
Phenotypes
CHH diagnosis and onset
CHH is defined clinically by the failure to initiate or complete normal puberty, with a variable defect in the maturation of secondary sexual characteristics, external genitalia, gametogenesis (Young et al. 2019) Table 1. Issues of sexual function and psychosocial adjustment are common in later life, possibly reflecting the consequences of delayed diagnosis and treatment (Dwyer et al. 2019b). The diagnosis of CHH is necessarily one of exclusion, since it requires acquired, functional or structural conditions to be ruled out, principally constitutional delay of growth and puberty (CDGP), parasellar lesions, pituitary iron overload, hyperprolactinemia, energy-deficit, metabolic-associated or drug-induced HH (Young et al. 2019). Having done so, CHH is clinically and biochemically manifested through low (or inappropriately normal) levels of gonadotropins, associated with amenorrhea and low oestradiol in females and low levels of total testosterone (TTe) in males (Boehm et al. 2015).
Nevertheless, in an otherwise fit and healthy young adult with preserved linear growth, there is little else other than CHH to account for the profile of gonadotropin-deficient absent puberty, particularly if clinical red flags for CHH are present, such as characteristic non-reproductive defects (i.e., anosmia, clefting, deafness), or features of absent neonatal minipuberty (history of cryptorchidism or micropenis) (Swee and Quinton 2019a). Although it might have been anticipated that CHH and CDGP would share elements of a common genetic basis, this does not in fact appear to be the case (Cassatella et al. 2018).
TTe levels at diagnosis in males are typically very low (< 3.5 nmol/L) (Pitteloud et al. 2002; Miraoui et al. 2013; Boehm et al. 2015), although somewhat higher levels have been reported in CHH males harbouring functionally validated homozygous mutations such as GNRHR Gln106Arg (de Roux et al. 1997; Costa et al. 2001; Pitteloud et al. 2001; Cangiano et al. 2019). According to whether puberty is arrested after a normal initiation, with some growth of testicular volume and activation of the pituitary–gonadal axis, or never started, it is possible to distinguish partial and complete forms of CHH (Fig. 1).

(Adaptation from (Palmert and Boepple 2001); Copyright permission granted from Oxford University Press; License n. 477821043851)
GnRH function during lifespan and different presentations of GnRH deficiency in congenital hypogonadotropic hypogonadism
This distinction is important because partial activation of the HPG axis at puberty suggests a milder underlying defect, with the likelihood that gonadotropin and gonadal steroid secretion during perinatal minipuberty (lasting from 2 months antepartum to 3–6 months after birth) was at least partly conserved. CHH males with partial or arrested puberty at presentation (e.g., testis volume > 4 mL) exhibit low-amplitude, low-frequency, or sleep-entrained-only LH pulses and have significantly better fertility outcomes with gonadotropin treatment. However, the majority (around 2/3) of CHH patients present with absent puberty (e.g., testis volume ≤ 4 mL) and show a flat LH profile on frequent sampling (Pitteloud et al. 2002) (Fig. 1).
In 1997 Nachtigall et al. described a form of idiopathic acquired isolated HH in adulthood (AHH), with similar hypotestosteronaemia (TTe < 4–4.5 nmol/L), with all possible secondary causes rigorously excluded (Nachtigall et al. 1997; Dwyer et al. 2010). Manifestations of AHH include infertility, sexual dysfunction, asthenia and reduced body hair, but with no clinical features to suggest prior incomplete pubertal maturation, with the potential for transmission of genes to the offspring prior to the onset of infertility. This could explain the existence of CHH families (Nachtigall et al. 1997; Dwyer et al. 2010) (Table 1).
While the genetic basis of AHH is not well characterised (Stamou and Georgopoulos 2018), there is an enrichment of rare CHH genetic variants compared to the general population, especially in AHH subjects with TTe level below 6 nmol/L and onset before 41 years of age (Cangiano et al. 2019). AHH has also been associated with minor pubertal delay in earlier life, suggesting the possibility of an underlying pre-existing mild impairment of hypothalamic–pituitary–gonadal (HPG) axis (Gianetti et al. 2010; Dwyer et al. 2010; Zhu et al. 2015). Crucially, some AHH patients also harbour non-reproductive defects commonly associated with classical CHH of pre-pubertal onset, including anomalous olfactory bulbs and sulci on MRI (Bonomi et al. 2018), thus further supporting the idea of a shared genetic basis.
Reversal
Conversely, CHH patients can revert—permanently or transiently—to a normal autonomous function of the reproductive axis (Quinton et al. 1999; Raivio et al. 2007; Duan et al. 2014) (Fig. 1). This phenomenon, known as reversal, is not uncommon, with a prevalence among the patients ranging from 5–10% (Quinton et al. 1999; Raivio et al. 2007; Duan et al. 2014) to 20% (Sidhoum et al. 2014). Even though mutations in TAC3 and TACR3 genes may particularly predispose to reversal (Gianetti et al. 2010; Root 2010), there is no reliable genotype–phenotype correlation and reversal has been reported also in patients harbouring deleterious variants in PROKR2 (Sinisi et al. 2008); FGFR1 (Raivio et al. 2007; Costa-Barbosa et al. 2013), GNRHR (Raivio et al. 2007), HS6ST1 (Costa-Barbosa et al. 2013), SOX10 (Maione et al. 2016) and even ANOS1 (Ribeiro et al. 2007).
Whilst some groups have found larger testicular volume and higher stimulated LH levels at diagnosis correlate with a higher possibility of reversal (Duan et al. 2014), others have not (Raivio et al. 2007); nonetheless an enlargement of testicular size during testosterone treatment or unexpected pregnancy in female partner should prompt brief discontinuation of the treatment followed by biochemical re-evaluation, to assess a potential recovery of an autonomous gonadal function. Crucially, reversal may not be sustained, indicating a persistent fragility or vulnerability of the HPG axis (Santhakumar et al. 2014), so that lifelong monitoring is recommended (Sidhoum et al. 2014).
No convincing cases of reversal have been documented in females, possibly because this is clinically so difficult to unravel from the scenario of energy-deficit hypothalamic amenorrhea, which is far more common and wherein periods of remission and relapse over time are characteristic.
Olfactory defect
Although a distinct population arises from the neural crest, the majority of hypothalamic GnRH neurons originate in the olfactory placode around the 5th gestational week and migrate alongside the olfactory, vomeronasal and terminal nerves until they reach their final destination in the mediobasal hypothalamus, infundibulum and periventricular region (Chung and Tsai 2010; Casoni et al. 2016; Haines et al. 2018). This shared origin is the reason why CHH is associated with olfactory impairment in around 50% of cases, thereby defining Kallmann syndrome (Boehm et al. 2015; Young et al. 2019).
Once considered as entirely separate diseases, KS and nCHH may also constitute separate manifestations of the same genetic disease, since they often coexist in the same kindreds (Trarbach et al. 2007; Dodé and Hardelin 2009; Pitteloud et al. 2010; Brioude et al. 2010). Indeed, they partly share the same genetic milieu.
With the probable exception of ANOS1 mutations in hemizygosity, which are almost invariably associated to complete loss of olfaction, mutations of CHH genes involved in GnRH neuron migration/axon guidance and GnRH neuron fate specification or differentiation (neurodevelopmental genes), have been associated with a variable degree of olfactory defect, ranging from normosmia, hyposmia to complete anosmia, i.e., with either KS or nCHH phenotypes. However, mutations of CHH genes associated with impairment of GnRH neuron activation, GnRH secretion or GnRH action at the gonadotroph level only cause nCHH (Hudson et al. 1994; Lewkowitz-Shpuntoff et al. 2012).
Genotype–phenotype correlations in relation to non-reproductive developmental abnormalities
In addition to olfactory deficit and neonatal reproductive features of impaired third trimester androgenization from absent minipuberty (micropenis and cryptorchidism), there are other developmental defects that disproportionately affect CHH patients, including hearing impairment, renal abnormalities, mirror movements (bimanual synkinesia), craniofacial clefting and abnormalities of eye movements, features that are significantly more prevalent among KS patients than nCHH (Quinton et al. 2001; Bhagavath et al. 2006; Bonomi et al. 2018) Table 2. Moreover, even though precise genotype–phenotype correlations may be lacking in published studies, both synkinesia and renal agenesis are almost entirely restricted to males (Bonomi et al. 2018) and are most commonly associated with ANOS1 mutations (Quinton et al. 2001), whereas dental agenesis and midline facial defects (as well as some distinctive digital bone abnormalities such as polydactyly, syndactyly and camptodactyly) are more prevalent among subjects harbouring deleterious variants of FGF8 and FGFR1 (Costa-Barbosa et al. 2013).
Cryptorchidism and micropenis are significantly more common in men with complete—rather than partial CHH (Pitteloud et al. 2002; Bonomi et al. 2018) and, independent of this, are also more frequent in KS patients than nCHH (Quinton et al. 2001; Bhagavath et al. 2006; Bonomi et al. 2012, 2018).
The prevalence of hearing loss (ranges from 5 to 15% of total CHH patients) is higher in KS (Quinton et al. 2001; Bonomi et al. 2018) and is usually associated to mutations in CHD7, SOX10, IL17RD, ANOS1, SOX10 (Costa-Barbosa et al. 2013; Pingault et al. 2013; Takagi et al. 2014; Vaaralahti et al. 2014; Wang et al. 2018) and SOX2 (Kelberman et al. 2006; Steevens et al. 2017). Sensorineural hearing defects also form part of complex syndromes associated with neuroendocrine genes such as LHX3 and DMXL2 (Bonfig et al. 2011; Pozza et al. 2012) Table 3.
It should be recognized that there may not always be a clearly dividing line—rather, a progressive fade-in—between the classification of “CHH-with-non-reproductive-defects” and “CHH as part of a complex syndrome”. This is particularly the case in respect of CHH versus CHARGE syndrome with CHD7 variants, although genetics may be helpful here, with inherited point mutation associated with CHH and large de novo deletions with CHARGE (Balasubramanian et al. 2014). Nevertheless, a pragmatic approach would be to consider whether CHH defines the major burden of disease experienced by the patient or whether the clinical burden of non-reproductive anomalies predominates in daily life.
Genetics
Since the discovery of ANOS1 (formerly KAL1) in the pathogenesis of X-linked KS, many more genes have been evaluated in relation to CHH. The clinical relevance of variants in ANOS1, FGFR1 and GNRH/GNRHR and PROK2/PROKR2 has been confirmed in many studies, with more than fifty different variants in each gene, or pair of genes, described as “disease causing” in the Human Gene Mutation Database, HGMD (Stenson et al. 2014). However, for genes described more recently, there is wide variation in the strength, number and consistency of reported evidence, in the number of variants accepted as pathogenic, and in the percentage of total CHH cases estimated to be underpinned by disease-causing variants.
KISS1/KISS1R and TAC3/TAC3R are consolidated candidate loci, with proof of pathogenicity arising from different studies and having over 25 different verified disease-causing variants in relation to nCHH.
Other genes such as SEMA3A, FGF8, SOX2, SOX10 are accumulating more evidence, having more than ten different disease-causing variants for CHH. CHD7 is strongly associated both with CHARGE syndrome (in which CHH is embedded within a particularly severe combination of non-reproductive phenotypes) and also a significant proportion of nCHH and KS cases.
Much less common, but equally strong in terms of causation, are LEP/LEPR in the pathogenesis of CHH associated with childhood-onset hyperphagia and obesity, along with genes such as DMXL2 and SMCHD1 that cause specific syndromes with CHH as a part of a complex phenotype, and genes such as OTUD4, RNF216, PNPLA6, STUB1, POLR3A and POLR3B that cause neurologic syndromes combined with CHH.
Genes such as NDNF, AMH/AMHR2, NTN1/DCC, FEZF1, HS6ST1 and NMSF1 have only been functionally validated in very few or even single studies and so have not yet passed the test of reproducibility to confirm their role. Moreover, humans’ variants in DUSP6, FLRT3, SPRY4, CCDC141, SEMA7A, TBX3 and GLCE entirely lack functionally validation. Finally, the CPHD-associated genes account only for a minimal proportion of isolated CHH.
Another approach to understanding the physiological importance of each locus, and hence its role in the pathogenesis of CHH, is to examine the prevalence of rare genetic variants among patient cohorts compared to control populations, with significant enrichment of rare variants in these loci in CHH indicating a major disease-causing role (see Table 4). Hitherto, this approach has performed well whenever it has been possible to test against conventional gene validation techniques.
Herein, we report all genes linked with CHH—listed according to their prevalent pathogenic mechanism—to provide a quick and useful reference for clinicians and scientists seeking information on one or more specific genes. For each gene, we present available knowledge about pathophysiology, inheritance, associated phenotypes and the types of variants reported in literature. Many of the genes implicated in CHH are G-protein-coupled receptors (GPCRs) and their cognate ligands, with mutations of the receptor usually being an order of magnitude more frequent. Thus, where the phenotype is clearly similar, we have described the effects of receptor and ligand allelic variants in the same section.
With the possible exception of the most deleterious FGFR1, TUBB3, SMCHD1 and CHD7 mutations, it may be that none of the variants described below is able, of itself, to cause a CHH disease phenotype in the heterozygous state. Where a sufficient number of cases have been described having homozygosity (or compound heterozygosity) for a given gene, then a plausible genotype–phenotype correlation can be outlined with some certainty. However, in many cases, this is not possible and, in particular, the relative impacts of other known or unknown genetic variants may be hard to disentangle.
Genes involved in GnRH neuron migration/axon guidance
ANOS1 (formerly KAL1)
ANOS1 encodes for an N-glycosylated protein called Anosmin-1, which is both expressed on the cell surface (Soussi-Yanicostas et al. 1996) and secreted (Rugarli et al. 1996). In 1991, it was the very first causal gene identified for CHH, discovered through positional cloning of DNA from patients having contiguous genes syndrome at Xp22.3 (Franco et al. 1991; Legouis et al. 1991). It is also the only gene in which a gene-deleted human foetus has been studied scientifically, showing failure of olfactory bulb development, with olfactory, terminal and vomeronasal axons ending in neurofibrillary tangles at the cribriform plate and GnRH neurons accumulating in the upper nasal septum and cribriform plate areas (Schwanzel-Fukuda et al. 1989).
The mode of inheritance is characteristically X-linked recessive, from the maternal healthy carrier to hemizygous affected male offspring. More than sixty mutations have been described until now, most of them nonsense mutations or large exon deletions. Nonetheless, a small number of missense variants have also been described in reported KS cases, principally affecting the whey acidic protein-like or the fibronectin-like type 3 domains and, hence, responsible for an impairment of disulphide bond formation and heparan binding, respectively (Kim et al. 2008b).
ANOS1 mutations have a high penetrance and expressivity and they are usually associated with a severe olfactory and reproductive phenotype (Oliveira et al. 2001; Salenave et al. 2008; Cangiano et al. 2019), with only one case of nCHH hitherto reported (Sato et al. 2004). Mutations in ANOS1 are also associated with bimanual synkinesia, midline defects, hearing loss and renal agenesis, which might relate to the known expression of ANOS1 in the developing kidney and other relevant sites (Duke et al. 1995; Costa-Barbosa et al. 2013).
The cited prevalence of ANOS1 mutations among affected individuals varies according to whether the study cohort comprised both sexes versus just males, total CHH versus just KS individuals, according to the proportion of sporadic cases versus familial cases and whether only unrelated individuals were included. These variables are not always clear from the literature. However, the prevalence of ANOS1 mutations among unrelated sporadic KS males may vary between 3.5 and 10% (Sykiotis et al. 2010; Laitinen et al. 2011; Hanchate et al. 2012; Basaran et al. 2013; Tommiska et al. 2014; Stamou et al. 2019).
Anosmin-1, the encoded glycoprotein, belongs to the superfamily of adhesion molecule that are important for neuronal adhesion and axonal extension and guidance. Indeed, Anosmin-1 is fundamental to olfactory axon guidance since—once the soluble form is incorporated to the extracellular matrix of the olfactory bulb—it promotes cell migration and the final targeting of olfactory axons, acting as a chemoattractant as well as a branch-promoting factor.
Immortalized migratory GnRH neurons exhibit a cell-specific chemotactic reaction when exposed to Anosmin-1-enriched media (Cariboni et al. 2004); anti-Anosmin-1 antibodies block the correct development of rats’ olfactory bulb branches in vitro (Soussi-Yanicostas et al. 2002) and, in C. elegans, an heterozygous mutation of the homologous of ANOS1 caused a highly penetrant axon-misrouting phenotype (Bülow et al. 2002). Moreover, Anosmin-1 co-localizes with FGFR1 in the olfactory structures during development (Ayari and Soussi-Yanicostas 2007) and it has been shown that ANOS1 could enhance FGF signalling interacting with FGFR–FGF–heparan sulphate proteoglycan complex on the cell surface (Gonzalez-Martinez 2004). However, functional studies in animal models have been severely circumscribed by the absence of a murine orthologue (Vezzoli et al. 2016).
HS6ST1
HS6ST1 encodes an enzyme that non-randomly introduces a sulphate in the 6-O-position of heparan sulphate—an extracellular matrix element important for guiding cell-to-cell communications during neural development and migration. Both in vivo and in vitro functional studies have shown that heparan sulphate proteoglycan chains are involved in the interaction and binding of Anosmin-1 to the cell membranes (Soussi-Yanicostas et al. 1996). Studies in C. elegans demonstrated that alteration of heparan 6-O-sulfotransferase worsened the ANOS1-related axonal defects (Bülow et al. 2002), suggesting that Anosmin-1 binds through this heparan sulphate proteoglycan to its cognate receptor (or other extracellular cues) to induce axonal branching and—if the system is altered—it can lead to axon misrouting. Moreover, heparan sulphate 6-O-sulfotransferase may also be involved in activation of the FGFR1 pathway through its synergy with Anosmin-1 (Tornberg et al. 2011).
In humans, missense allelic variants affecting highly conserved residues of HS6ST1 were first described in a cohort of 338 CHH patients (Tornberg et al. 2011) and a subsequent report found an association of HS6ST1 variants with both reversal and oligogenicity (Costa-Barbosa et al. 2013). Along with the great variance in the expressivity of the gene variants described, these observations suggest that alterations of HS6ST1 alone might not be sufficient to produce a CHH phenotype. Indeed, a recent report demonstrated the possible involvement of an HS6ST1 variant in case of self-limited delayed puberty (Howard et al. 2018) in contrast with other GnRH deficiency gene (Cassatella et al. 2018).
PROK2-PROKR2
PROK2, and its GPCR, PROKR2, are primarily expressed in the arcuate nucleus, the olfactory tract and the suprachiasmatic nucleus and are fundamental for migration and development of both GnRH and olfactory neurons (Ng 2005; Prosser et al. 2007). They were first described to be associated with CHH by Dodé et al. (2006), with Prok2/Prokr2 mutations causing agenesis or hypoplasia of the olfactory bulbs and abnormal migration of GnRH neurons in mice (Matsumoto et al. 2006; Pitteloud et al. 2007b). Associated phenotypes in humans include fibrous dysplasia, synkinesia and epilepsy (Cole et al. 2008).
Since then, many variants have been found in human CHH patients, both with and without anosmia, and in vitro studies have demonstrated a deleterious effect on downstream signalling (Dodé et al. 2006; Pitteloud et al. 2007b; Leroy et al. 2008; Cole et al. 2008; Monnier et al. 2009; Abreu et al. 2012; Libri et al. 2014; Cox et al. 2018). More recently, it was demonstrated that PROKR2 variants might also be responsible for a “biased signalling” effect on the receptor; indicating the need to test all potential couplings of PROKR2 when a likely causal allelic variant protein is pharmacologically tested (Libri et al. 2014; Sbai et al. 2014).
The majority of PROK2 and PROKR2 mutations are missense variants and patients exhibit an inconsistent phenotype due to a variable expressivity and penetrance. Whilst homozygous mutations are associated with a severe phenotype of high penetrance (Dodé et al. 2006; Abreu et al. 2008; Sarfati et al. 2010; Libri et al. 2014), PROKR2 heterozygous variants showed no genotype–phenotype correlation at all, in the pedigrees reported (Dodé et al. 2006; Falardeau et al. 2008; Monnier et al. 2009; Caronia et al. 2011; Raivio et al. 2012; Abreu et al. 2012; Sbai et al. 2014). Indeed, using criteria of the American College of Medical Genetics and Genomics, a recent reanalysis of published PROKR2 allelic variants—hitherto considered to be pathogenic—reassigned several of them to the category of uncertain significant or even benign (Cox et al. 2018). However, a similar reanalysis of (much rarer) published PROK2 variants is clearly overdue.
Moreover, certain functionally validated variants that were initially considered to have an autosomal dominant (AD) transmission with full expressivity and penetrance (Dodé et al. 2006), were subsequently also found in AHH (Libri et al. 2014; Cangiano et al. 2019), in cases of reversal (Sinisi et al. 2008) and in unaffected subjects (Pitteloud et al. 2007b; Abreu et al. 2008). Therefore, as the heterozygous phenotype is often expressed in associations with rare variants in other genes, oligogenicity may be an obligate requirement for heterozygous variants of PROK2/PROKR2 to cause disease (Leroy et al. 2008; Sykiotis et al. 2010).
Due to the discordant results of functional studies, it remains uncertain whether certain variants can exert a dominant-negative effect. For instance, in evaluating the in vitro co-expression of a mutated and a wild-type allele, (Monnier et al. 2009) found both no impairment in the signal transmission or other evidence supporting a dominant-negative effect, allowing autosomal dominant transmission, whereas (Abreu et al. 2012) and (Cox et al. 2018) did so.
Finally, heterozygous PROKR2 mutations—validated in vitro—are also associated with CPHD and septo-optic dysplasia (SOD), suggesting a role of this gene in pituitary development, which is supported by expression of Prokr2 in the pars nervosa of the pituitary in the murine animal model (Raivio et al. 2012; McCormack et al. 2017).
Semaphorin signalling (SEMA3A, SEMA3E, SEMA7A, PLXNA1)
Semaphorins belong to a family of proteins that—besides being crucial in the control of immune and vascular systems and tumorigenesis—play a pivotal role in neural circuit development, principally by controlling axon guidance processes. This guidance action is fundamental also for development of the olfactory system and the migration of GnRH neurons and many genes in this family have been linked to CHH (Messina and Giacobini 2013; Oleari et al. 2019b).
SEMA3A encodes a class 3 (i.e., interacting with neuropilins as co-receptor for the ligand) semaphorin that is expressed in the olfactory system as a secreted protein—which is essential for guidance of vomeronasal (Cariboni et al. 2011) and olfactory (Pasterkamp et al. 1998) axons—and was found to underpin human CHH in 2012 (Young et al. 2012; Hanchate et al. 2012). Data from a murine model demonstrated that Sema3a deletions, or missense mutations in the binding domain of NRP1 (encoding SEMA3A’s co-receptor, Neuropilin-1), impair GnRH neuron migration and normal development of the olfactory system (Cariboni et al. 2011; Hanchate et al. 2012).
SEMA3A variants in CHH patients are usually missense mutations found in an heterozygous state; once believed to be a gene with an autosomal dominant transmission, the complex pattern of inheritance together with accumulating evidence of associated variants in other CHH genes, now more convincingly signposts oligogenicity (Young et al. 2012; Hanchate et al. 2012; Känsäkoski et al. 2014).
Plexin-A1, encoded by PLXNA1, is the receptor activated by Semaphorin-3A and Neuropilin-1 and is important for neuronal development during embryonic growth. It is expressed in the olfactory system and the vomeronasal organ and nerve (Messina and Giacobini 2013; Marcos et al. 2017). Studies in the mouse have demonstrated that homozygous gene deletions produce a KS-like phenotype, albeit with incomplete penetrance (Marcos et al. 2017). Very recently the synergistic action of another member of the PLXNA family has been demonstrated. Thus, double Plxna1 and Plxna3 knockout mice phenocopy the olfactory and GnRH neuron defects observed in Sema3a-null mice (Oleari et al. 2019a).
A missense heterozygous mutation in SEMA3E, in association with a heterozygous CHD7 variant, affecting two brothers with Kallmann’s Syndrome was recently reported using sequencing techniques and computational modelling (Cariboni et al. 2015). Functional studies demonstrated that the Semaphorin 3E protein product is necessary for GnRH neuron survival upon entering the brain (Cariboni et al. 2015). Further, a de novo SEMA3E missense variant was reported in a patient with CHARGE syndrome, thereby reinforcing the relationship between semaphorins and CHD7 (Lalani et al. 2004). An enrichment in rare heterozygous SEMA3E variants in AHH is also consistent with variable expressivity (Cangiano et al. 2019).
Besides SEMA3A and SEMA3E, other components of the semaphorin family are being studied for their roles in neuron migration and as potential candidates for the pathogenesis of human CHH. Among them, the Sema7a ortholog was found to be involved in GnRH neuron migration in the mouse and in vitro, being widely expressed both in the nasal placode and along the olfactory/vomeronasal axonal scaffold and having an important role in the regulation of cell motility (Messina et al. 2011), the protein binds to Plexin-C1 to decrease integrin-mediated cell attachment and spreading (Messina and Giacobini 2013). A recent study found several SEMA7A heterozygous missense variants in CHH patients, albeit also in association with other candidate genes (Känsäkoski et al. 2014). Given the complex roles of semaphorins in axon guidance, neuronal migration, as well as in neuronal plasticity, it will not be no surprise if additional members of the semaphoring–plexin signalling pathways are found to be implicated in CHH (Oleari et al. 2019b).
NSMF
NMDA receptor, synaptonuclear signalling and neuronal migration factor (NSMF), also called “nasal embryonic LHRH factor” (NELF), is expressed during murine olfactory and GnRH neuron development (Kramer and Wray 2000, 2001), and the encoded Jacob protein was believed to be required for guidance of olfactory axonal projections and consequent GnRH migration (Xu et al. 2010). NMSF variants were subsequently reported in human CHH (Miura et al. 2004; Tornberg et al. 2011). However, recent in vivo findings in murine Nsmf knockouts do not support a major role for Jacob protein in the migration of GnRH-positive neurons during early development (Spilker et al. 2016). This is consistent with the allelic variants reported in humans being heterozygous, associated with other verified causative gene mutations and not fully segregating with the clinical phenotype (Pitteloud et al. 2007a; Xu et al. 2011). Thus, the extreme rarity of this gene variants together with these reported observations question the role of NSMF in the pathogenesis of CHH and, at the very least, make monogenic causation of CHH by loss-of-function mutations very unlikely.
WDR11
Synergistic with EMX1 (a transcription factor encoded by Empty Spiracles Homeobox 1—EMX1) in the development of murine olfactory neurons, WD Repeat Domain 11 shuttles between nucleus and cytoplasm to activate transcription (Kim et al. 2010). Initially expressed widely during central nervous system (CNS) development—but particularly in the regions involved in the development of hypothalamic GnRH neurons (Kim et al. 2010)—it later localizes just to olfactory structures, cerebellum and hippocampus. In 2010, Kim et al. described several missense heterozygous variants in WDR11 domains important for protein–protein interaction, in patients with both KS and nCHH (Kim et al. 2010), alone or associated with mutations in other known genes (Kim and Layman 2011), suggesting the possibility of oligogenic or AD inheritance. A WDR11 variant has also been reported in association with a PROKR2 variant in patient with CPHD and absent pituitary stalk (McCormack et al. 2017).
FEZF1
FEZF1 is a zinc finger protein acting as a repressor of transcription during the embryogenesis of olfactory structures; enabling olfactory and GnRH neurons to access the brain. Loss-of-function missense (affecting the central C2H2 motif necessary for protein stability) or nonsense autosomal recessive (AR) allelic variants of this gene are associated with severe and highly penetrant KS phenotypes (Kotan et al. 2014). Furthermore, KO mice replicate the human phenotype, with hypoplasic olfactory structures and CHH (Hirata et al. 2006; Watanabe et al. 2009). Nevertheless, the lack of new reported human variants since then suggests either the extreme rarity of FEZF1 variants in CHH/KS pathogenesis or that other mutational events might explain the reported phenotypes in the two independent consanguineous index families.
CCDC141
CCDC141 is a cytoskeletal scaffolding protein, with a role in cellular motility. It is expressed in GnRH neurons and its missense allelic variants have been reported in association with nCHH, incomplete penetrance and possible reversal (Turan et al. 2017), complex inheritance patterns observed in families described seeming to support oligogenicity. In an animal model, CCDC141 was shown to be important for GnRH neuron migration along olfactory axon fascicles, but without affecting development of the fascicles themselves, consistent with the postulated human phenotype (Hutchins et al. 2016).
Netrin signalling (DCC and NTN1)
Netrin-1 (NTN1) is an axon guidance secreted protein, containing an Anosmin1-like fibronectin type-III (FN3) domain. DCC encodes its receptor, with the animal model exhibiting a KS-like phenotype (Schwarting et al. 2004; Lakhina et al. 2012). In a study of 133 CHH patients, six patients harboured heterozygous DCC missense variants and two harboured heterozygous NTN1 missense variants. Five of six subjects had olfactory impairment and all had severe GnRH deficiency (Bouilly et al. 2018). Although inheritance was compatible with AD transmission, some patients also harboured mutations in other CHH genes and two of them had variants in both DCC and NTN1, thus also supporting oligogenic inheritance. In vitro studies of these variants revealed altered intracellular signalling associated with alterations in cell morphology due to defective binding (Bouilly et al. 2018). Interestingly, alterations in DCC and NTN1 signalling had already been associated with congenital mirror movements and corpus callosum agenesis (Srour et al. 2010; Marsh et al. 2017),
SOX10 and TUBB3
SOX10 is a transcription factor involved in the early development of neural crest cells, which are a population of multipotent precursor cells arising from the neural tube that differentiate into various specific cell types. SOX10 also has critical influence on auditory function through its expression in the melanocytic intermediate cells of the cochlear stria vascularis during early development of the inner ear (Breuskin et al. 2009). Mutations are classically associated with Waardenburg syndrome, a rare disorder characterized by sensorineural congenital hearing loss and abnormal pigmentation of the hair, skin and eye (Pingault et al. 2010). However, SOX10 loss-of-function mutations have also been found in nearly 40% of KS patients having with hearing impairment (Pingault et al. 2013).
Given that the prevalence of deafness in KS individuals is estimated to be only 5% and that SOX10 mutations are rare in KS individuals with normal hearing, this relationship appears highly significant. SOX10 mutations were shown to affect olfactory ensheathing cells during early embryonic development of the peripheral olfactory system in mouse models, leading to the disrupted development of the olfactory neurons as well as impaired migration of GnRH neurons, thus underpinning the KS phenotype (Pingault et al. 2013).
Similar to SOX10, genetic alterations in TUBB3 have been associated with disrupted migration of neural crest cells. The TUBB3 protein is a member of tubulin family and its alterations are responsible for a multiple neurological complex syndrome (Whitman et al. 2016; Ceylan et al. 2017; Huang et al. 2018). A single missense mutation (p.E410K) of TUBB3 was also associated with KS and severe peripheral neuropathy, having with an AD inheritance pattern with strong genotype–phenotype correlation (Chew et al. 2013; Balasubramanian et al. 2015; Nakamura et al. 2018).
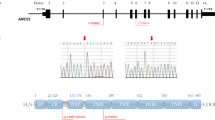
AMH and AMHR2
AMH belongs to the TGF-β family of proteins and its cognate receptor is AMHR2. Signalling in this pathway plays a pivotal role in sex differentiation, with homozygous loss-of-function mutations leading to persistence of Mullerian ducts in males. However, later in gestation, it also plays a role in the regulation of GnRH neuron migration towards the brain, through an autocrine mechanism intervening in axonal growth and pathfinding (Liu et al. 2004). In fact AMH is expressed in migratory GnRH neurons in both mouse and human foetuses, and Amhr2-deficient mice show abnormal development of the peripheral olfactory system and defective GnRH cells migration, with reduced fertility in adults (Cimino et al. 2016; Malone et al. 2019).
Only recently, heterozygous missense AMH allelic variants in the N-terminal pro-protein domain have been found in both KS and nCHH subjects and an in-frame 27-nucleotide deletion in the catalytic intracellular serine/threonine domain of the receptor AMHR2 found in a patient with nCHH. These variants are present in both male and female subjects with absent or partial puberty, one of whom also exhibited high-arched palate (Malone et al. 2019). These findings are consistent with AD inheritance with variable expressivity (variability in spontaneous puberty and even fertility since the same variants were found in parents), although oligogenicity obviously cannot be excluded. Therefore, only a wider genetic study on a large patient cohort will clarify the role of these genes in CHH/KS pathogenesis.
NDNF
NDNF (neuron-derived neurotrophic factor) is involved in neuron survival, migration and neurite outgrowth (Kuang et al. 2010). It belongs to the FN3 superfamily of genes, which encode proteins typically involved in protein–protein interactions related to cell adhesion, migration and embryonic development (Bencharit et al. 2007). Ndnf deficient mice showed anomalies in GnRH neuron migration to the hypothalamus and development of the olfactory axonal scaffold, findings that were further confirmed using a Zebrafish ndnf morpholino-based knockdown model (Messina et al. 2020).
This is the most recent gene to be associated with CHH, with four different heterozygous variants reported in different subjects having KS with severe GnRH deficiency. Pedigree analysis of the affected kindreds is consistent with AD inheritance with variable expressivity and incomplete penetrance, although oligogenicity cannot be excluded. Only one of the allelic variants was missense, two were nonsense mutations and one was a frameshift that introduced a premature stop codon. Only the latter three variants—associated with a truncated protein—resulted in a loss of function in vitro (Messina et al. 2020).
GnRH neuron and gonadotroph differentiation and fate specification
FGFR1 synexpression group
FGFR1 is a member of the tyrosine kinase receptor superfamily of proteins and gain-of-function mutations are associated with craniosynostosis. Fgfr1 plays a role in GnRH neuron proliferation and migration to the hypothalamus as well as directly promoting olfactory bulb development. Accordingly, loss-of-function FGFR1 mutations can lead to defective GnRH neuron migration through abnormal olfactory bulb morphogenesis, although some individuals express a “pure” neuroendocrine nCHH phenotype without non-reproductive anomalies, implying a putative role in the control of GnRH secretion that remains to be defined.
Indeed, FGFR1 was the first gene in which inactivating mutations were found in both normosmic CHH and KS. Loss-of-function mutations in FGFR1 have been identified in approximately 10% of CHH patients, with the majority of pathogenic variants being missense mutations (Dodé et al. 2003). It has a potentially AD inheritance, albeit associated in many kindreds with markedly incomplete penetrance, interfamily variability (Pitteloud et al. 2006) and oligogenicity (Pitteloud et al. 2007a). Phenotypically, affected individuals are enriched with skeletal manifestations, including craniofacial clefting, digit anomalies (syndactyly, oligodactyly and clinodactyly) and dental agenesis (Jarzabek et al. 2012; Costa-Barbosa et al. 2013).
Loss-of-function FGFR1 mutations were recently also implicated in Hartsfield syndrome, characterized by holoprosencephaly and split hand/foot malformation (absent or partial development of the central rays of the hands or feet) (Simonis et al. 2013), consistent with the observation that, among the rare subgroup of CHH patients with split hand/foot malformation, around 90% harbour FGFR mutations (Villanueva et al. 2015).
FGF8 functions as a potent ligand for FGFR1. FGF8 is expressed in the diencephalon and prospective hypothalamus during embryonic development (McCabe et al. 2011a). It is a critical morphogen for GnRH neuron fate specification and olfactory system development. Targeted transgenic mice lacking olfactory placode Fgf8 expression fail to develop GnRH neurons. In humans, FGF8 mutations account for no more than 1% of CHH cases, with an apparently AD mode of inheritance. Cleft lip or palate and other midline defects have been reported in subjects harbouring the mutation. FGF8 mutations have also been found to be associated with recessive holoprosencephaly, craniofacial defects and hypothalamic–pituitary dysfunction (McCabe et al. 2011a). Studies in mice revealed a decrease in 30–50% of total GnRH neurons in those harbouring monogenic heterozygous Fgf8 mutations, whereas mice with digenic Fgfr1/Fgf8 mutations suffered a greater reduction in GnRH neurons, suggesting a high degree of interaction between these FGF signalling factors in promoting tropic support for the emergence of GnRH neurons in the olfactory placode (Chung et al. 2010).
Fgf17 is co-expressed with Fgf8 in the olfactory placode and is thus considered a member of the Fgf8 synexpression group (Miraoui et al. 2013). Possessing a high degree of homology to FGF8, FGF17 shares similar signalling effect through isoform FGFR1c and is thus implicated in GnRH neuron embryonic morphogenesis (Olsen et al. 2006). Heterozygous FGF17 mutations have been identified in three patients with CHH (2 KS and 1 normosmic) (Miraoui et al. 2013).
IL17RD is a single transmembrane glycoprotein with sequence resembling the intracellular domain of the interleukin-17 receptor. It is an antagonist of the FGF signalling pathway, both at the level of the FGF receptors, including FGFR1, as well as the downstream components of the Ras–ERK1/2 pathway. Having close spatiotemporal interdependence with FGF8 in the olfactory placode suggests an equally critical role for the initial phase of GnRH neuron fate specification (Miraoui et al. 2013). Additionally, it likely exerts modulatory effect on embryonic optic development via the FGF8–FGFR1c pathway, consistent with the association of IL17RD mutations in KS with congenital hearing impairment. Phenotypic expression is likely only in the presence of biallelic IL17RD mutations or of oligogenicity, rather than from a single allelic defect (Miraoui et al. 2013).
SPRY4 encodes sprout homolog 4, a protein that exerts antagonistic effect on FGF signalling via inhibition of the receptor-transduced mitogen-activated protein kinase (MARK) signalling pathway. It is located upstream of RAS gene activation and disrupts the formation of active GTP-RAS (Hanafusa et al. 2002). In murine studies, SPRY4 is shown to regulate neurite outgrowth in PC12 cells, a widely utilized model of neuronal differentiation, and hippocampal neurons. Knockout mice exhibit craniofacial and limb defects (Taniguchi et al. 2007) (Hausott et al. 2012). Close to 4% of a CHH cohort in one study were found to harbour SPRY4 mutations, with two-thirds manifesting impaired olfaction.
DUSP6 encodes a member of the dual specificity protein phosphatase which inactivates members of the mitogen-activated protein (MAP) kinase superfamily. In mice studies, Dusp6 mutant alleles result in skeletal dwarfism, craniosynostosis and hearing loss (Li et al. 2007). Human DUSP6 mutations account for no more than 1% of CHH subjects and are found in both KS and normosmic CHH phenotypes, with other associated non-reproductive features being hearing impairment and abnormal dentition (Miraoui et al. 2013).
FLRT3 encodes a member of the fibronectin leucine-rich transmembrane (FLRT) family that interacts with FGFR during embryogenesis. The transmembrane cell adhesion protein is characterized by a cluster of leucine-rich repeats and one fibronectin type-III domain within the extracellular matrix. As an axon guidance-related factor and enhancer of the FGF network, it promotes the activation of FGF signalling via ERK phosphorylation and neurite outgrowth in rat neuronal cells (Böttcher et al. 2004; Robinson et al. 2004). FLRT3 variants were found in three unrelated KS individuals (two females and one male) manifesting CHH with partial or complete absence of puberty (Miraoui et al. 2013).
KLB encodes β-Klotho, a type I single-pass transmembrane protein that functions as the primary high-affinity receptor for FGF21, which in turn exerts its effect via signalling through the β-klotho/FGFR1c receptor complex (Ming et al. 2012). In mice, lack of Klb results in defective FGF21-mediated GnRH secretion by hypothalamic GnRH neurons, leading to pubertal delay, altered oestrous cyclicity and subfertility (Xu et al. 2017). In a cohort of CHH subjects, 4% were found to harbour heterozygous loss-of-function KLB mutations; of the seven heterozygous variants identified in the probands, six were missense variants and one was an in-frame deletion (Xu et al. 2017). As the FGF21/KLB/FGFR1 pathway also mediates metabolic processes, it may reinforce a linkage between metabolic and reproductive processes.
CHD7
CHD7 encodes chromodomain helicase DNA-binding protein 7, which is expressed in a variety of foetal tissues including the developing brain. It is one of two CHH genetic loci shared with CHARGE syndrome, a rare developmental disorder of autosomal dominant inheritances, characterized by iris coloboma, congenital heart disease, choanal atresia, mental and growth retardation, genital hypoplasia, and ear malformations or deafness (Jongmans et al. 2006).
CHH occurs in around 2/3 of patients carrying CHD7 mutations (Jongmans et al. 2006); conversely, only 6% of patients CHH (with or without anosmia) harbour CHD7 mutations (Kim et al. 2008a). This is not surprising given the overlapping features of olfactory impairment and CHH observed in these two groups of patients (Jongmans et al. 2009). Therefore, while multisystem involvement and large de novo deletions characterize classical CHARGE syndrome, CHD7-associated CHH appears to result from inherited point mutations (Kim et al. 2008a; Balasubramanian et al. 2014).
IGSF10
IGSF10 is a member of the immunoglobulin superfamily, which appears to form part of a complex system of chemostatic agents directing the early migration of GnRH neurons from the nasal region to the hypothalamus (Wray 2010); aberrant signalling due to pathogenic IGSF10 variants can lead to impaired GnRH neuron migration to the hypothalamus (reduced population or mis-timed arrival), as supported by an Igsf10 knockdown-zebrafish model, wherein disruption of immature GnRH neuron migration was demonstrated. The authors postulated that variable phenotypes may manifest according to the burden of other associated mutations (oligogenicity), ranging from self-limited delayed puberty to CHH (Howard et al. 2016).
SOX2
SOX2, SRY (Sex Determining Region Y)-box 2 gene, encodes a transcriptional regulator in pluripotent stem cells, required for maintaining their pluripotency, as well as directing their neural differentiation (Zhang and Cui 2014). Sox2 is important in the development of the anterior pituitary gland in the mouse. Indeed, although Sox2−/− null embryos die shortly after implantation (Avilion et al. 2003), heterozygous mutant mice showed a variable altered size and shape of the pituitary gland and a significant reduction in pituitary hormone content in affected mice (Kelberman et al. 2006).
Human SOX2 mutations are associated with bilateral anophthalmia or severe microphthalmia, with anterior pituitary hypoplasia, CHH and genital abnormalities in males (Fantes et al. 2003; Williamson et al. 2006; Kelberman et al. 2006; Jayakody et al. 2012). Additional forebrain defects include hypoplasia of the corpus callosum, hypothalamic hamartoma and hippocampal malformation (8, 10), frequently associated with additional abnormalities, including oesophageal atresia, sensorineural hearing loss and learning difficulties (Sisodiya et al. 2006; Kelberman et al. 2006).
NR0B1 (encoding DAX-1)
Encoded by NR0B1, DAX-1 (dosage sensitive sex reversal adrenal hypoplasia, critical region on the X-chromosome, gene 1) is an orphan nuclear receptor expressed in the adrenals, gonads, pituitary gonadotrophs and ventromedial hypothalamus, and therefore plays a key role in adrenal and reproductive development. Accordingly, NR0B1 mutations result in an X-linked form of primary adrenal hypoplasia congenital, associated with CHH and impaired spermatogenesis (Muscatelli et al. 1994). Most patients present first with neonatal adrenal insufficiency and, subsequently, in adolescence with pubertal delay due to CHH arising from a combined hypothalamic, pituitary and gonadal defect (Habiby et al. 1996). However, rare cases of late onset in adulthood have been reported (Kyriakakis et al. 2017).
GnRH neuron activation and networking
Kisspeptin and KISS1R
Kisspeptin (KISS1) and its receptor KISS1R (formerly GPR54) were identified as causal genes for CHH in 2003 (de Roux et al. 2003; Seminara et al. 2003) and 2012 (Topaloglu et al. 2012), respectively. The identification of the hypothalamic kisspeptin neuronal network has transformed our perception of the control and activation of GnRH secreting neurons at puberty. Indeed, kisspeptin (formerly known as metastin) is a strong activator of the hypothalamic–pituitary–gonadal axis in both human and animal models. KISS1 encodes four different products, whose binding to KISS1R is independent of their length (Vezzoli et al. 2016).
KISS1 is expressed in periventricular and arcuate nuclei (Gottsch et al. 2004) and is the main regulator of GnRH neuron activation and hormone secretion, as demonstrated by the precocious puberty phenotype associated with a gain-of-function mutation in KISS1R (Teles et al. 2008). Both Kiss1 and Kiss1r knockout mice phenocopy human CHH (Seminara et al. 2003; Funes et al. 2003; de Tassigny et al. 2007). Since these two genes play no role in GnRH neuron migration and development they produce a “pure neuroendocrine” nCHH phenotype with an AR mode of inheritance (de Roux et al. 2003; Seminara et al. 2003; Cerrato et al. 2006; Topaloglu et al. 2012). So far, there are reports of both nonsense mutations impairing receptor function (de Roux et al. 2003) and membrane targeting (Seminara et al. 2003), and missense mutations affecting domains essential for the activation of KISS1R (Seminara et al. 2003), its expression on the membrane, or its signalling (Semple et al. 2005; Tenenbaum-Rakover et al. 2007). Both a deletion and a missense variant found in the third intracellular loop decreased KISS1R signalling (de Roux et al. 2003; Semple et al. 2005).
TAC3 and TACR3
TAC3 encodes a tachykinin called Neurokinin B (NKB), and TACR3 its cognate receptor (a GPCR of the rhodopsin family). Both are expressed mainly in the arcuate nucleus of the hypothalamus (together with KISS1) in KNDy (kisspeptin–neurokinin–dynorphin) cells projecting to GnRH neurons (Goodman et al. 2007; Navarro et al. 2009; Wakabayashi et al. 2010), and are believed to play an important role in GnRH activation during puberty, foetal life and perinatal minipuberty (Gianetti et al. 2010). They were first associated with AR transmission of CHH in 2009, with in vitro validated allelic variants being missense (Topaloglu et al. 2009; Gianetti et al. 2010), nonsense (Gianetti et al. 2010), or splice-site variants (Young et al. 2010), impairing receptor signalling or post-translational modifications necessary for correct activation of the tachykinin motif.
Rat models initially gave contrasting results on the stimulatory effect of NKB, probably due to the necessity for physiological levels of sex steroids for its stimulatory effect to be expressed, which was subsequently confirmed in many species (Navarro 2013). Human mutations are associated with nCHH, often with cryptorchidism and micropenis, but potentially with greater propensity for reversal (Gianetti et al. 2010).
Direct secretion and action of GnRH
GNRHR and GNRH1
Gonadotropin-releasing hormone (encoded by GNRH1) and its receptor (encoded by GNRHR) are the primary actors in the regulation of human reproduction. The pulsatile secretion of GnRH in the median eminence and the interaction with its receptor on pituitary gonadotrophs modulate the release of gonadotropins (Seeburg and Adelman 1984). Thus far, GNRH1 is the only hypothalamic releasing hormone gene found mutated in humans. Although GnRH1 was the very first candidate gene for CHH (Weiss et al. 1991), mutations were only identified in humans 10 years ago due to their exceptional rarity (Bouligand et al. 2009; Chan et al. 2009). By contrast, GNRHR was among the first three genes to be associated with CHH (de Roux et al. 1997; Layman et al. 1998).
GnRH resistance mediated by deleterious variants of GNRHR has a heterogeneous phenotype and inconstant expressivity causing both CHH and AHH, with mixed AR and oligogenic inheritance (Cerrato et al. 2006), and with most AR cases exhibiting compound heterozygosity. It is a common cause of familial nCHH with more than 25 variants reported in the literature (Cioppi et al. 2019) and can also produce incomplete phenotypes with milder or partial disease (de Roux et al. 1999; Pitteloud et al. 2001; Cangiano et al. 2019; Cioppi et al. 2019), including possible reversal forms (Raivio et al. 2007). The majority of the variants found in humans are missense variants that variably impair GnRH signalling through abnormal intracellular trafficking, hormone binding, G-protein coupling or receptor expression (Bianco and Kaiser 2009).
Gnrh1 knockout mice show a similar phenotype to human CHH (Pask et al. 2005; Wu et al. 2010). In contrast, human GNRH1 gene allelic variants (nonsense variants with AR or oligogenic inheritance), are exceptionally rare and are all located in the secreted decapeptide itself (Bouligand et al. 2009; Chan et al. 2009; Mengen et al. 2016). In the mouse model, mutations of Gnrh1 produce infertility, sexual immaturity, and tooth and mineralization defects (Cattanach et al. 1977; Tiong et al. 2007). However, in humans, GNRHR and GNRH1 mutations are associated with nCHH without non-reproductive defects, since they have no known role in normal embryonic development (de Roux et al. 1997; Beranova et al. 2001; de Roux 2006).
Actual or functional energy-deficit CHH
LEP and LEPR
Leptin, encoded by LEP (formerly Ob), is a hormone secreted by white adipose tissue, in proportion to the total fat mass and inversely regulated by exercise. It acts through the leptin receptor, encoded by LEPR (formerly Db) and belonging to the class 1 cytokine receptor family. Rodents harbouring homozygous mutations in Lep and LepR exhibit early-onset morbid obesity, hyperphagia and decreased energy expenditure (Zhang et al. 1994; Lee et al. 1996). Parabiosis experiments performed in the pre-genetic era, wherein the blood circulations of Lep and LepR mice were linked, famously resulted in dramatic transformation of the Lep mouse from a hyperphagic obese phenotype into one of starvation with cessation of food intake, the LepR phenotype remaining necessarily unchanged.
In humans, leptin signalling deficiency caused by loss-of-function LEP or LEPR mutations is an extremely rare cause of monogenic obesity and CHH. Notably, individuals with heterozygous LEP or LEPR mutations do not exhibit pubertal or reproductive disorders, as only one functional copy is required for normal reproductive function (Clément et al. 1998). In a hyper-selected cohort of 300 subjects with hyperphagia and childhood-onset obesity, the prevalence of pathogenic LEPR mutations was only 3% (Farooqi et al. 2007b), whereas the prevalence of homozygous leptin variants in general population is estimated to be 1 in 4.4 Mio. (Nunziata et al. 2017).
Affected individuals suffer from severe obesity as a consequence of relentless hyperphagia. Furthermore, as a clear indication of the critical link that leptin serves between nutritional status and hypothalamic–pituitary–gonadal function, they also manifest CHH and fail to undergo normal puberty. In leptin-deficient subjects, recombinant leptin therapy was effective in reversing HH and restoring gonadal and sexual function (Farooqi et al. 1999; Farooqi 2002). Moreover, recombinant leptin also restored menstrual cyclicity and reproductive function in women with hypothalamic amenorrhea (HA) and low fat mass due to energy deficit (Welt et al. 2004).
Although the effect of leptin on the gonadotropic axis is remarkable, GnRH neurons do not themselves express LEPR, their activity being instead modulated indirectly via forebrain neurons expressing LEPR that are afferent to GnRH (Quennell et al. 2009). Although LEPR is also expressed in a subset of kisspeptin neurons (Smith et al. 2006), the main site of leptin action to regulate reproduction is not directly on kisspeptin neurons, but rather through cells in the ventral premammillary nucleus (PMV) (Cravo et al. 2013; Cavalcante et al. 2014; Iacovazzo et al. 2016). Finally, it was recently demonstrated that PACAP (pituitary adenylate cyclase-activating polypeptide) secreted by leptin-responsive neurons in the hypothalamic ventral premammillary nucleus (PMV), also serves as a mediator for leptin to exert its reproductive role (Ross et al. 2018).
HA is a reasonably common condition exhibited by women with energy deficit due to overexercising or restriction of food intake, short of frank anorexia nervosa that requires nutritional rehabilitation. The observation that only a proportion of women facing similar degrees of energy deficit develop HA was recently explained by the observation that women with HA are enriched with deleterious heterozygous variants of CHH genes compared with normally menstruating controls (Caronia et al. 2011).
By contrast, energy-deficit functional CHH seems to be much rarer in males (who are paradoxically more predisposed to develop functional CHH in relation to obesity). Moreover, no convincing enrichment with deleterious CHH alleles has been demonstrated in these men (Dwyer et al. 2019a).
Defects of gonadotropin secretion or action
PCSK1
PCSK1 encodes neuroendocrine convertase, whose deficiency results in impaired prohormone processing; in particular this enzyme is responsible for processing pro-opiomelanocortin in the corticotroph to produce adrenocorticotropic hormone and lipotropin. Rare cases of compound heterozygosity and homozygosity for mutation in the PCSK1 gene have been reported in subjects with associated features of early-onset severe obesity and hyperphagia with CHH (Jackson et al. 1997; Farooqi et al. 2007a).
LHB and FSHB
Biallelic inactivating mutations in the specific β-subunits of LH and FSH have been described as rare causes of men and women presenting with reproductive disorders and are characterized by an unusual hormonal profile, not typical of CHH, wherein levels of only one of the two gonadotropins are low, whilst levels of the other are normal or even elevated. Homozygosity and compound heterozygosity with AR mode of transmission have been reported (Lofrano-Porto et al. 2007; Basciani et al. 2012).
Men with pathogenic LHB variants express normal sexual differentiation, implying in utero responsiveness to chorionic gonadotropin (hCG), but exhibit pubertal failure, Leydig cell hypoplasia and spermatogenic arrest from the lack of LH-stimulated testosterone secretion (Basciani et al. 2012). Accordingly, restoration of testicular growth and spermatogenesis with hCG therapy was described in a man with homozygous missense LHB mutation (Valdes-Socin et al. 2009). By contrast, affected women may undergo normal puberty, but then manifest secondary amenorrhoea and anovulatory infertility (Lofrano-Porto et al. 2007).
In human and mouse studies, FSH is essential for qualitatively and quantitatively normal sperm production in males as well as for normal female pubertal development and fertility via the regulation of ovarian follicular development. Deleterious FSHB mutations are autosomal recessive, with men harbouring biallelic FSHB mutations undergoing normal puberty, but remaining azoospermic, whereas women present with abnormal pubertal development and primary amenorrhoea (Layman et al. 1997, 2002).
Gonadotroph differentiation and CPHD
An entire subset of genes act through impairment in the development, differentiation, or fate specification of pituitary cells, typically generating a complex phenotype with CPHD. However, many genes previously associated with CHH (such as PROKR2, FGF8, FGFR1, SOX2, WDR11) have recently also been associated with CPHD (including septo-optic dysplasia—SOD) and vice versa (McCabe et al. 2011b, 2013, 2015; Raivio et al. 2012; Jayakody et al. 2012; Izumi et al. 2014), sometimes even interacting in an oligogenic manner (McCormack et al. 2017).
LHX4
LHX4, encoding Lim Homeobox gene 4, is highly conserved in evolution and plays an important role as a transcriptional regulator of embryonic development. Human LHX4 encodes a 390-amino acid protein that has a tandem pair of LIM domains and one homeodomain, and is highly homologous to the human LHX3 protein, except for the N-terminal region (Kawamata et al. 2002). As demonstrated in null mice models, both Lhx3 and Lhx4 regulate proliferation, differentiation and fate specification of pituitary-specific cell lineages, dictating pituitary gland identity and controlling developmental decisions at multiple steps of organogenesis (Sheng et al. 1997).
In 2001 Machinis et al. described LHX4 heterozygous mutations in four affected members of a French family with a syndrome of autosomal dominant short stature, pituitary and cerebellar defects and abnormalities of the sella turcica (Machinis et al. 2001). Following this first report, other authors reported allelic variants in LHX4 gene in different domains of the protein—all associated with hypopituitarism, including CHH. Nevertheless, LHX4 mutations remain a relatively rare cause of CPHD (Pfaeffle et al. 2008).
HESX1
HESX1 is another homeobox gene and an important transcription repressor involved in the morphogenesis of pituitary gland, forebrain, hypothalamus and optic nerve (Dattani et al. 1998). It has also been shown to be a transcriptional target of OTX2. Variable inheritance patterns and penetrance are described, leading to a range of phenotypic manifestations including KS, isolated GHD and CPHD, with or without associated forebrain and ocular defects (McNay et al. 2007; Newbern et al. 2013; Fang et al. 2016a). HESX1 mutations have been identified in approximately 1% of CHH and septo-optic dysplasia patients. Both HESX1 recessive and dominant variants may underlie CPHD (Romero et al. 2011).
PITX2 and GATA2
PITX2 is another homeobox gene. The PITX2 protein plays a critical role in pituitary organogenesis, including the growth of Rathke’s pouch and maintaining expression of the transcription factors PROP1 and HESX1, and later in the fate specification and expansion of gonadotrophs and PUO1F1 (PIT1) lineages within the ventral and caudomedial anterior pituitary (Dattani and Robinson 2000; Suh et al. 2002). AD inheritance of PITX2 produces Axenfeld–Rieger syndrome, with alterations of the anterior segment of the eyes, dental abnormalities and hypopituitarism (Seifi and Walter 2018), including potentially CHH. In mouse models, diminished expression of gonadotrope markers Lhb, Fshb and Gnrhr in Pitx2 hypomorphs point to the importance of Pitx2 in cell fate specification (Suh et al. 2002). Moreover, Pitx2 is necessary to drive the cascade of transcription factors (starting with Gata2) in cell lineage fate specification and proliferation (Suh et al. 2002).
GATA2 is expressed in the developing pituitary as well as adult gonadotropes and thyrotropes. Gata2 is genetically downstream of transcription factor Pitx2 (Suh et al. 2002). In pituitary-specific knockout mice, gonadotropin secretion was reduced both basally and in response to castration challenge, albeit fertility was preserved (Charles et al. 2006). Consistent with this finding, GATA2 was demonstrated to participate in the modulation of LHβ gene promoter and Gnrhr promotor expression by other investigators (Lo et al. 2011; Schang et al. 2013).
PROP1
PROP1 is the most frequently mutated gene implicated in non-syndromic CPHD, accounting for up to 50% of familial cases. It acts both as a repressor in down-regulation of HESX1 and as an activator of POU1F1, possibly via Wnt/β-catenin signalling, in the differentiation of anterior pituitary cells including gonadotropes (Dasen and Rosenfeld 2001; Olson et al. 2006). AR mode of inheritance of loss-of-function PROP1 variants is observed in kindreds. Compared to growth hormone and thyrotropin deficiencies, which typically manifest early in life, gonadotropin deficiency may only evolve to present later as pubertal failure during adolescence (Turton et al. 2005; Kelberman et al. 2009).
LHX3
LHX3 belongs to the Lin11, Isl-1 and Mec-3 (LIM) homeodomain protein family of transcription factors. It is expressed in the neural tube and Rathke’s pouch and has an essential role in regulating pituitary development and the differentiation of gonadotropes, thyrotropes, somatotropes and lactotrophs, as well as in the organisation of spinal cord neurons (Tsuchida et al. 1994; Bach et al. 1995). Mutations of LHX3 are AR, with majority found to be missense or nonsense mutations. Besides anterior pituitary hormone deficiencies, affected individuals may present with short neck with limited neck rotation and vertebral abnormalities (Bhangoo et al. 2006; Pfaeffle et al. 2007). As LHX3 variants disrupt development of inner ear, affected individuals may also manifest a varying degree of hearing loss (Bonfig et al. 2011; Pozza et al. 2012).
SOX3
SOX3 belongs to the SRY-related HMG transcription factor family. It is expressed widely in the central nervous system with a critical role in hypothalamic–pituitary morphogenesis. Duplications and deletions in the polyalanine tract of SOX3 have been shown to result in varying severity of anterior pituitary deficiencies (Alatzoglou et al. 2011). In a recent Japanese genome-wide copy number analysis and systematic mutation screening study, polyalanine deletions in SOX3 were described for the first time as a genetic cause of normosmic CHH amongn a cohort of patients (Izumi et al. 2014).
OTX2
Orthodenticle Drosophila homolog 2 (OTX2) is a homeobox family transcription factor. Otx2 is expressed in anterior neuroectoderm during early embryogenesis development and participates critically in the formation of the rostrum, midbrain and eyes (Kurokawa et al. 2004; Henderson et al. 2009). Around 1/3 of subjects harbouring OTX2 mutations were found to have pituitary insufficiency, including CHH (Schilter et al. 2011). OTX2 mutations also account for 2–3% of cases of congenital anophthalmia/microphthalmia (Wyatt et al. 2008). The variable penetrance of OTX2 variants, even within the same family (Gorbenko Del Blanco et al. 2012), is likely determined by oligogenicity and environmental factors (Tajima et al. 2013). In mice, heterozygous Otx male embryos had a substantial reduction in the total GnRH neuron population due to impaired neuronal migration from the nasal placode, resulting in subfertility and compromised production of luteinising hormone in adulthood; by contrast, heterozygous female mutants did not survive to adulthood (Diaczok et al. 2011; Hoffmann et al. 2019).
GLI2
GLI2 is a zinc finger transcription factor involved in the regulation of the Sonic Hedgehog signal transduction, expressed in the ventral diencephalon and oral ectoderm, which induces BMP4 and FGF8 expression and pituitary progenitors, respectively. Mutant mice with inactivating Gli2 manifest hypoplasia of the anterior pituitary gland and anomalies of the midline central diencephalon. In humans, GLI2 inactivating mutations are associated with a phenotype at the milder end of the holoprosencephaly spectrum, including pituitary gland malformation and craniofacial defects as well as varying degree of hypopituitarism (isolated GHD through to CPHD) (Kevelam et al. 2012). The inheritance pattern follows that of autosomal dominant with incomplete penetrance and variable expressivity (Kevelam et al. 2012).
Syndromes of CHH with cerebellar ataxia
A group of genes discovered with next-generation techniques have been associated with neurological syndromes (especially ataxia) that are usually associated with CHH.
POLR3A and POLR3B encode catalytic subunits of RNA polymerase and AR inheritance of mutations results in abnormal central nervous system white matter: the hypomyelinating leukodystrophy (Daoud et al. 2013) with prominent cerebellar dysfunction, oligodontia and CHH (Bernard et al. 2011; Tétreault et al. 2011).
OTUD4, RNF216 and STUB1 encode proteins involved in protein quality control. AR or oligoallelic inheritance of variants in these genes leads to disordered ubiquitination, determining the Gordon Holmes syndrome (Seminara et al. 2002; Margolin et al. 2013; Shi et al. 2014), which is characterized by hypogonadotropic hypogonadism (both of hypothalamic and pituitary origin), spinocerebellar ataxia, spasticity and dementia. AR allelic variants in PNPLA6 can also produce either Gordon Holmes syndrome, or Boucher–Neuhäuser syndrome (lacking spasticity, but instead exhibiting visual impairment due to chorioretinal dystrophy). PNPLA6 encodes an enzyme necessary for synthesis of acetylcholine and mutations are usually located in the C-terminal phospholipid esterase domain, thus inactivating its catalytic function (Synofzik et al. 2014; Topaloglu et al. 2014).
Other syndromic forms
DMXL2
Encoding a synaptic scaffold protein for the regulators of the GTPase Rab3a expressed in exocytosis vesicles of GnRH neurons and gonadotrophs, DMXL2 is associated with a polyendocrine-polyneuropathy syndrome, with ataxia and dysarthria, characterized by mild CHH, central hypothyroidism, peripheral demyelinating neuropathy and severe hypoglycaemia that later degenerated into insulin-dependent diabetes mellitus (Tata et al. 2014). It has AD transmission and a mild reproductive phenotype with incomplete puberty and normal sense of smell. In heterozygous knockout mice it is associated with a reduction in the number of GnRH neurons, retardation of puberty and impaired fertility (Tata et al. 2014). Functional studies found DMXL2 to be involved in both constitutive and glucose-induced secretion of insulin by pancreatic beta cells.
TBX3
TBX3 is a member of the family of transcriptional factors sharing a DNA-binding domain known as T-box. As it is expressed in pituitary gland, gonads and genital tubercles, as well as mammary glands and limbs, TBX3 haploinsufficiency results in ulnar–mammary syndrome (UMS), a rare AD characterized by under-developed external genitalia, delayed puberty, ulnar ray defects and hypoplasia of nipples and apocrine glands (Bamshad et al. 1997). Heterozygous variants of TBX3 have been described in two unrelated families with nCHH and pituitary hypoplasia, which is in line with the high prevalence of pubertal delay reported in the UMS literature (Sasaki et al. 2002; Galazzi et al. 2018).
SMCHD1
SMCHD1 is an epigenetic repressor, intervening in the inactivation of the X-chromosome (Blewitt et al. 2008) and is especially expressed in immature olfactory neurons (Nickell et al. 2012). In 2017 Shaw et al. identified heterozygous missense SMCHD1 mutations in the GHKL-type ATPase domain in 84% of patients with Bosma arhinia microphthalmia syndrome (Shaw et al. 2017), associated with CHH in 97% of subjects. This AD syndrome produces a severe highly penetrant disease. The zebrafish model showed a reduction of the projection length of the terminal nerve (where GnRH3 neurons are located) along with ethmoid plate abnormalities (Shaw et al. 2017).
Inheritance, oligogenicity and variants of uncertain significance
All modes of transmission have been demonstrated in relation to the genetics of CHH involved, albeit with an ever-expanding role for oligogenicity since it was first reported (Pitteloud et al. 2007a, 2010; Sykiotis et al. 2010). Earlier publications in the field of CHH genetics (antecedent to the introduction of NGS techniques—NGS) necessarily lacked the capacity to identify potential deleterious variants in other loci, whether known or unknown. More recently, NGS is uncovering ever-increasing evidence of oligogenicity, at least partly explaining the apparent variable penetrance and expressivity of certain variants. Many heterozygous variants were previously believed to have a dominant (or incompletely penetrant) transmission, but we now know that only the most disrupting ones have a true AD inheritance and, in the vast majority of cases, CHH is more likely to arise from genetic hemizygosity (ANOS1), homozygosity, compound heterozygosity, or oligogenicity.
In this review we have reported more than sixty genes associated to CHH. However, no single study has ever simultaneously evaluated this wide assortment of loci in a large patient cohort so as to provide a better understanding in a larger number of CHH patients. Knowledge of the oligogenic basis of CHH makes genotype–phenotype characterization increasingly uncertain, but it also explains the great variability found within and between kindreds that apparently share the same pathogenic genetic variant. The overlap between KS, nCHH, CPHD, AHH and HA further complicates the scenario making it even more essential to deploy NGS analyses using ever-wider panels of candidate genes (Raivio et al. 2012; Jayakody et al. 2012).
However, these advances in genomics come at a cost, with NGS allowing simultaneous screening of a vast number of genes, great numbers of variants of unknown clinical significance (VUS) are necessarily thrown up. The challenge now is to more cheaply, rapidly and reliably identify those variants that are truly pathogenic. Recent studies investigating individuals from general population as controls with NGS techniques have found 10–20% of them (according to the number of genes screened and the filtering criteria used to define a rare variant) harbouring heterozygous missense VUS in a CHH candidate gene (Sykiotis et al. 2010; Cassatella et al. 2018; Cangiano et al. 2019), thereby reminding that not all the rare variants are able to disrupt the function of a protein. Therefore, many groups are implementing new methods to increase the specificity of genetic analyses so as to better distinguish pathogenic variants from among the VUS (Richards et al. 2015; Cox et al. 2018).
Genetic counselling
With appropriate gonadotropin therapy and assisted reproductive techniques, many CHH men and women now have improved prospect of achieving fertility. In these individuals, genetic counselling forms an important component of the fertility management because of the potential risk of transmission to offspring. However, with the increasingly complex genetic architecture of CHH/KS propelled by massively parallel next-generation sequencing, providing informed, reliable and relevant genetic counselling has become incredibly challenging (Maione et al. 2018).
One of the significant challenges in predicting the risk of transmission of disease features is the difficulty in obtaining complete information on the phenotypes in all family members (ideally across three generations). It is further complicated by phenotypic expression that could be modified by different underlying genetic background (oligogenicity) as well as environmental exposure; variation in phenotype is frequently observed within the same family due to incomplete penetrance. Therefore, even for mutations with well-established classical Mendelian mode of transmission, variable expressivity and disease severity are likely.
Moreover, while oligogenicity has emerged to be an important genetic cause in a significant proportion of CHH subjects (Miraoui et al. 2013), distinguishing true oligogenicity from rare potentially deleterious variants that have yet to be equivocally linked to CHH remains a key challenge. Adding to the uncertainty is the different impact of individual mutations on disease manifestations by variants in other loci. It may also be impossible to pinpoint the likelihood of oligogenic transmission to offspring of affected parents in such cases because of missing information in respect of many of these putative genes.
On the other hand, knowledge of the specific genetic mutation could have important therapeutic implications. For instance, individuals with X-linked NR0B1 pathogenic variants tend to exhibit dismal response to gonadotropin therapy in the induction of spermatogenesis, with subsequent microsurgical sperm retrieval being required to have any hope of biological parenthood (Mantovani et al. 2006). These men should be counselled specifically, with alternative options such as donor sperm or adoption discussed early on.
Practically, for individuals harbouring mutations with classical Mendelian inheritance mode of transmission (AD, AR, or X-linked transmission), genetic counselling can be provided accordingly, along with other considerations as mentioned above. In cases of oligogenicity, a general estimate of 5–10% transmission risk may be counselled, while engaging them in continual discussion as novel information may arise from the constantly evolving genetic landscape from time to time (Sykiotis et al. 2010; Swee and Quinton 2019b). Importantly, recognizing that biochemical screening of offspring for minipuberty can be performed during the first 2–3 months of life, irrespective of whether the parental genotype is known, is a crucial early strategy in facilitating the exclusion of significant GnRH deficiency; it should be prudent to discuss this with potential future parents during the counselling process.
References
Abreu AP, Trarbach EB, de Castro M et al (2008) Loss-of-function mutations in the genes encoding prokineticin-2 or prokineticin receptor-2 cause autosomal recessive Kallmann syndrome. J Clin Endocrinol Metab 93:4113–4118. https://doi.org/10.1210/jc.2008-0958
Abreu AP, Noel SD, Xu S et al (2012) Evidence of the importance of the first intracellular loop of prokineticin receptor 2 in receptor function. Mol Endocrinol 26:1417–1427. https://doi.org/10.1210/me.2012-1102
Alatzoglou KS, Kelberman D, Cowell CT et al (2011) Increased transactivation associated with SOX3 polyalanine tract deletion in a patient with hypopituitarism. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2010-1239
Avilion AA, Nicolis SK, Pevny LH et al (2003) Multipotent cell lineages in early mouse development depend on SOX2 function. Genes Dev 17:126–140. https://doi.org/10.1101/gad.224503
Ayari B, Soussi-Yanicostas N (2007) FGFR1 and anosmin-1 underlying genetically distinct forms of Kallmann syndrome are co-expressed and interact in olfactory bulbs. Dev Genes Evol 217:169–175. https://doi.org/10.1007/s00427-006-0125-0
Bach I, Rhodes SJ, Pearse RV et al (1995) P-Lim, a LIM homeodomain factor, is expressed during pituitary organ and cell commitment and synergizes with Pit-1. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.92.7.2720
Balasubramanian R, Choi JH, Francescatto L et al (2014) Functionally compromised CHD7 alleles in patients with isolated GnRH deficiency. Proc Natl Acad Sci USA 111:17953–17958. https://doi.org/10.1073/pnas.1417438111
Balasubramanian R, Chew S, MacKinnon SE et al (2015) Expanding the phenotypic spectrum and variability of endocrine abnormalities associated with TUBB3 E410K syndrome. J Clin Endocrinol Metab 100:E473–E477. https://doi.org/10.1210/jc.2014-4107
Bamshad M, Lin RC, Law DJ et al (1997) Mutations in human TBX3 alter limb, apocrine and genital development in ulnar-mammary syndrome. Nat Genet 16:311–315. https://doi.org/10.1038/ng0797-311
Basaran Y, Bolu E, Unal HU et al (2013) Multiplex ligation dependent probe amplification analysis of KAL1, GNRH1, GNRHR, PROK2 and PROKR2 in male patients with idiopathic hypogonadotropic hypogonadism. Endokrynol Pol 64:285–292. https://doi.org/10.5603/EP.2013.0007
Basciani S, Watanabe M, Mariani S et al (2012) Hypogonadism in a patient with two novel mutations of the luteinizing hormone β-subunit gene expressed in a compound heterozygous form. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2012-1986
Bencharit S, Bin CC, Siddiqui A et al (2007) Structural insights into fibronectin type III domain-mediated signaling. J Mol Biol 367:303–309. https://doi.org/10.1016/j.jmb.2006.10.017
Beranova M, Oliveira LM, Bédécarrats GY et al (2001) Prevalence, phenotypic spectrum, and modes of inheritance of gonadotropin-releasing hormone receptor mutations in idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab 86:1580–1588. https://doi.org/10.1210/jcem.86.4.7395
Bernard G, Chouery E, Putorti ML et al (2011) Mutations of POLR3A encoding a catalytic subunit of RNA polymerase Pol III cause a recessive hypomyelinating leukodystrophy. Am J Hum Genet 89:415–423. https://doi.org/10.1016/j.ajhg.2011.07.014
Bhagavath B, Podolsky RH, Ozata M et al (2006) Clinical and molecular characterization of a large sample of patients with hypogonadotropic hypogonadism. Fertil Steril 85:706–713. https://doi.org/10.1016/j.fertnstert.2005.08.044
Bhangoo APS, Hunter CS, Savage JJ et al (2006) A novel LHX3 mutation presenting as combined pituitary hormonal deficiency. J Clin Endocrinol Metab 91(3):747–753. https://doi.org/10.1210/jc.2005-2360
Bianco SDC, Kaiser UB (2009) The genetic and molecular basis of idiopathic hypogonadotropic hypogonadism. Nat Rev Endocrinol 5:569–576. https://doi.org/10.1038/nrendo.2009.177
Blewitt ME, Gendrel A-V, Pang Z et al (2008) SmcHD1, containing a structural-maintenance-of-chromosomes hinge domain, has a critical role in X inactivation. Nat Genet 40:663–669. https://doi.org/10.1038/ng.142
Boehm U, Bouloux P-M, Dattani MT et al (2015) European Consensus Statement on congenital hypogonadotropic hypogonadism—pathogenesis, diagnosis and treatment. Nat Rev Endocrinol 11:547–564. https://doi.org/10.1038/nrendo.2015.112
Bonfig W, Krude H, Schmidt H (2011) A novel mutation of LHX3 is associated with combined pituitary hormone deficiency including ACTH deficiency, sensorineural hearing loss, and short neck—a case report and review of the literature. Eur J Pediatr. https://doi.org/10.1007/s00431-011-1393-x
Bonomi M, Libri DV, Guizzardi F et al (2012) New understandings of the genetic basis of isolated idiopathic central hypogonadism. Asian J Androl 14:49–56. https://doi.org/10.1038/aja.2011.68
Bonomi M, Vezzoli V, Krausz C et al (2018) Characteristics of a nationwide cohort of patients presenting with isolated hypogonadotropic hypogonadism (IHH). Eur J Endocrinol 178:23–32. https://doi.org/10.1530/EJE-17-0065
Böttcher RT, Pollet N, Delius H, Niehrs C (2004) The transmembrane protein XFLRT3 forms a complex with FGF receptors and promotes FGF signalling. Nat Cell Biol. https://doi.org/10.1038/ncb1082
Bouilly J, Messina A, Papadakis G et al (2018) DCC/NTN1 complex mutations in patients with congenital hypogonadotropic hypogonadism impair GnRH neuron development. Hum Mol Genet 27:359–372. https://doi.org/10.1093/hmg/ddx408
Bouligand J, Ghervan C, Tello JA et al (2009) Isolated familial hypogonadotropic hypogonadism and a GNRH1 mutation. N Engl J Med 360:2742–2748. https://doi.org/10.1056/NEJMoa0900136
Breuskin I, Bodson M, Thelen N et al (2009) Sox10 promotes the survival of cochlear progenitors during the establishment of the organ of Corti. Dev Biol. https://doi.org/10.1016/j.ydbio.2009.09.007
Brioude F, Bouligand J, Trabado S et al (2010) Non-syndromic congenital hypogonadotropic hypogonadism: clinical presentation and genotype-phenotype relationships. Eur J Endocrinol 162:835–851. https://doi.org/10.1530/EJE-10-0083
Bülow HE, Berry KL, Topper LH et al (2002) Heparan sulfate proteoglycan-dependent induction of axon branching and axon misrouting by the Kallmann syndrome gene kal-1. Proc Natl Acad Sci USA 99:6346–6351. https://doi.org/10.1073/pnas.092128099
Cangiano B, Duminuco P, Vezzoli V et al (2019) Evidence for a common genetic origin of classic and milder adult-onset forms of isolated hypogonadotropic hypogonadism. J Clin Med. https://doi.org/10.3390/jcm8010126
Cariboni A, Pimpinelli F, Colamarino S et al (2004) The product of X-linked Kallmann’s syndrome gene (KAL1) affects the migratory activity of gonadotropin-releasing hormone (GnRH)-producing neurons. Hum Mol Genet 13:2781–2791. https://doi.org/10.1093/hmg/ddh309
Cariboni A, Davidson K, Rakic S et al (2011) Defective gonadotropin-releasing hormone neuron migration in mice lacking SEMA3A signalling through NRP1 and NRP2: implications for the aetiology of hypogonadotropic hypogonadism. Hum Mol Genet 20:336–344. https://doi.org/10.1093/hmg/ddq468
Cariboni A, André V, Chauvet S et al (2015) Dysfunctional SEMA3E signaling underlies gonadotropin-releasing hormone neuron deficiency in Kallmann syndrome. J Clin Invest 125:2413–2428. https://doi.org/10.1172/JCI78448
Caronia LM, Martin C, Welt CK et al (2011) A genetic basis for functional hypothalamic amenorrhea. N Engl J Med 364:215–225. https://doi.org/10.1056/NEJMoa0911064
Casoni F, Malone SA, Belle M et al (2016) Development of the neurons controlling fertility in humans: new insights from 3D imaging and transparent fetal brains. Development 143:3969–3981. https://doi.org/10.1242/dev.139444
Cassatella D, Howard SR, Acierno JS et al (2018) Congenital hypogonadotropic hypogonadism and constitutional delay of growth and puberty have distinct genetic architectures. Eur J Endocrinol 178:377–388. https://doi.org/10.1530/EJE-17-0568
Cattanach BM, Iddon CA, Charlton HM et al (1977) Gonadotrophin-releasing hormone deficiency in a mutant mouse with hypogonadism. Nature 269:338–340. https://doi.org/10.1038/269338a0
Cavalcante JC, Bittencourt JC, Elias CF (2014) Distribution of the neuronal inputs to the ventral premammillary nucleus of male and female rats. Brain Res 1582:77–90. https://doi.org/10.1016/j.brainres.2014.07.034
Cerrato F, Shagoury J, Kralickova M et al (2006) Coding sequence analysis of GNRHR and GPR54 in patients with congenital and adult-onset forms of hypogonadotropic hypogonadism. Eur J Endocrinol 155(Suppl):S3–S10. https://doi.org/10.1530/eje.1.02235
Ceylan AC, Gursoy H, Yildirim N et al (2017) Clinical heterogeneity associated with TUBB3 gene mutation in a Turkish family with congenital fibrosis of the extraocular muscles. Ophthalmic Genet 38:288–290. https://doi.org/10.1080/13816810.2016.1193881
Chan Y-M, de Guillebon A, Lang-Muritano M et al (2009) GNRH1 mutations in patients with idiopathic hypogonadotropic hypogonadism. Proc Natl Acad Sci USA 106:11703–11708. https://doi.org/10.1073/pnas.0903449106
Charles MA, Saunders TL, Wood WM et al (2006) Pituitary-specific Gata2 knockout: effects on gonadotrope and thyrotrope function. Mol Endocrinol. https://doi.org/10.1210/me.2005-0378
Chew S, Balasubramanian R, Chan W-M et al (2013) A novel syndrome caused by the E410K amino acid substitution in the neuronal β-tubulin isotype 3. Brain 136:522–535. https://doi.org/10.1093/brain/aws345
Chung WCJ, Tsai P-S (2010) Role of fibroblast growth factor signaling in gonadotropin-releasing hormone neuronal system development. Kallmann syndrome and hypogonadotropic hypogonadism. KARGER, Basel, pp 37–50
Chung WCJ, Matthews TA, Tata BK, Tsai PS (2010) Compound deficiencies in multiple fibroblast growth factor signalling components differentially impact the murine gonadotrophin-releasing hormone system. J Neuroendocrinol. https://doi.org/10.1111/j.1365-2826.2010.02024.x
Cimino I, Casoni F, Liu X et al (2016) Novel role for anti-Müllerian hormone in the regulation of GnRH neuron excitability and hormone secretion. Nat Commun 7:10055. https://doi.org/10.1038/ncomms10055
Cioppi F, Riera-Escamilla A, Manilall A et al (2019) Genetics of ncHH: from a peculiar inheritance of a novel GNRHR mutation to a comprehensive review of the literature. Andrology 7:88–101. https://doi.org/10.1111/andr.12563
Clément K, Vaisse C, Lahlou N et al (1998) A mutation in the human leptin receptor gene causes obesity and pituitary dysfunction. Nature. https://doi.org/10.1038/32911
Cole LW, Sidis Y, Zhang C et al (2008) Mutations in prokineticin 2 and prokineticin receptor 2 genes in human gonadotrophin-releasing hormone deficiency: molecular genetics and clinical spectrum. J Clin Endocrinol Metab 93:3551–3559. https://doi.org/10.1210/jc.2007-2654
Costa EMF, Bedecarrats GY, Mendonca BB et al (2001) Two novel mutations in the gonadotropin-releasing hormone receptor gene in Brazilian patients with hypogonadotropic hypogonadism and normal olfaction1. J Clin Endocrinol Metab 86:2680–2686. https://doi.org/10.1210/jcem.86.6.7551
Costa-Barbosa FA, Balasubramanian R, Keefe KW et al (2013) Prioritizing genetic testing in patients with Kallmann syndrome using clinical phenotypes. J Clin Endocrinol Metab 98:E943–E953. https://doi.org/10.1210/jc.2012-4116
Cox KH, Oliveira LMB, Plummer L et al (2018) Modeling mutant/wild-type interactions to ascertain pathogenicity of PROKR2 missense variants in patients with isolated GnRH deficiency. Hum Mol Genet 27:338–350. https://doi.org/10.1093/hmg/ddx404
Cravo RM, Frazao R, Perello M et al (2013) Leptin signaling in Kiss1 neurons arises after pubertal development. PLoS ONE 8:e58698. https://doi.org/10.1371/journal.pone.0058698
Dai W, Wu J, Zhao Y et al (2019) Functional analysis of SOX10 mutations identified in Chinese patients with Kallmann syndrome. Gene 702:99–106. https://doi.org/10.1016/j.gene.2019.03.039
Daoud H, Tétreault M, Gibson W et al (2013) Mutations in POLR3A and POLR3B are a major cause of hypomyelinating leukodystrophies with or without dental abnormalities and/or hypogonadotropic hypogonadism. J Med Genet 50:194–197. https://doi.org/10.1136/jmedgenet-2012-101357
Dasen JS, Rosenfeld MG (2001) Signaling and transcriptional mechanisms in pituitary development. Annu Rev Neurosci 24:327–355. https://doi.org/10.1146/annurev.neuro.24.1.327
Dattani MT, Robinson IC (2000) The molecular basis for developmental disorders of the pituitary gland in man. Clin Genet 57(5):337–346. https://doi.org/10.1034/j.1399-0004.2000.570503.x
Dattani MT, Martinez-Barbera JP, Thomas PQ et al (1998) Mutations in the homeobox gene HESX1/Hesx1 associated with septo-optic dysplasia in human and mouse. Nat Genet. https://doi.org/10.1038/477
Diaczok D, Divall S, Matsuo I et al (2011) Deletion of Otx2 in GnRH neurons results in a mouse model of hypogonadotropic hypogonadism. Mol Endocrinol. https://doi.org/10.1210/me.2010-0271
Dodé C, Hardelin J-P (2009) Kallmann syndrome. Eur J Hum Genet 17:139–146. https://doi.org/10.1038/ejhg.2008.206
Dodé C, Levilliers J, Dupont JM et al (2003) Loss-of-function mutations in FGFR1 cause autosomal dominant Kallmann syndrome. Nat Genet. https://doi.org/10.1038/ng1122
Dodé C, Teixeira L, Levilliers J et al (2006) Kallmann syndrome: mutations in the genes encoding prokineticin-2 and prokineticin receptor-2. PLoS Genet 2:e175. https://doi.org/10.1371/journal.pgen.0020175
Duan J, Chen R-R, Li L et al (2014) Reversal of idiopathic hypogonadotropic hypogonadism: a cohort study in Chinese patients. Asian J Androl. https://doi.org/10.4103/1008-682X.145072
Duke VM, Winyard PJD, Thorogood P et al (1995) KAL, a gene mutated in Kallmann’s syndrome, is expressed in the first trimester of human development. Mol Cell Endocrinol 110:73–79. https://doi.org/10.1016/0303-7207(95)03518-C
Dwyer AA, Hayes FJ, Plummer L et al (2010) The long-term clinical follow-up and natural history of men with adult-onset idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab 95:4235–4243. https://doi.org/10.1210/jc.2010-0245
Dwyer AA, Chavan NR, Lewkowitz-Shpuntoff H et al (2019a) Functional hypogonadotropic hypogonadism in men: underlying neuroendocrine mechanisms and natural history. J Clin Endocrinol Metab 104:3403–3414. https://doi.org/10.1210/jc.2018-02697
Dwyer AA, Smith N, Quinton R (2019b) Psychological aspects of congenital hypogonadotropic hypogonadism. Front Endocrinol (Lausanne) 10:353. https://doi.org/10.3389/fendo.2019.00353
Dzemaili S, Tiemensma J, Quinton R et al (2017) Beyond hormone replacement: quality of life in women with congenital hypogonadotropic hypogonadism. Endocr Connect 6:404–412. https://doi.org/10.1530/EC-17-0095
Falardeau J, Chung WCJ, Beenken A et al (2008) Decreased FGF8 signaling causes deficiency of gonadotropin-releasing hormone in humans and mice. J Clin Invest 118:2822–2831. https://doi.org/10.1172/JCI34538
Fang Q, Benedetti AFF, Ma Q et al (2016a) HESX1 mutations in patients with congenital hypopituitarism: variable phenotypes with the same genotype. Clin Endocrinol (Oxf). https://doi.org/10.1111/cen.13067
Fang Q, George AS, Brinkmeier ML et al (2016) Genetics of combined pituitary hormone deficiency: roadmap into the genome era. Endocr Rev 37(6):636–675. https://doi.org/10.1210/er.2016-1101
Fantes J, Ragge NK, Lynch S-A et al (2003) Mutations in SOX2 cause anophthalmia. Nat Genet 33:461–463. https://doi.org/10.1038/ng1120
Farooqi IS (2002) Leptin and the onset of puberty: insights from rodent and human genetics. Semin Reprod Med. https://doi.org/10.1055/s-2002-32505
Farooqi IS, Jebb SA, Langmack G et al (1999) Effects of recombinant leptin therapy in a child with congenital leptin deficiency. N Engl J Med. https://doi.org/10.1056/NEJM199909163411204
Farooqi IS, Volders K, Stanhope R et al (2007a) Hyperphagia and early-onset obesity due to a novel homozygous missense mutation in prohormone convertase 1/3. J Clin Endocrinol Metab 92(9):3369–3373. https://doi.org/10.1210/jc.2007-0687
Farooqi IS, Wangensteen T, Collins S et al (2007b) Clinical and molecular genetic spectrum of congenital deficiency of the leptin receptor. N Engl J Med. https://doi.org/10.1056/NEJMoa063988
Franco B, Guioli S, Pragliola A et al (1991) A gene deleted in Kallmann’s syndrome shares homology with neural cell adhesion and axonal path-finding molecules. Nature 353:529–536. https://doi.org/10.1038/353529a0
Francou B, Bouligand J, Voican A et al (2011) Normosmic congenital hypogonadotropic hypogonadism due to TAC3/TACR3 mutations: characterization of neuroendocrine phenotypes and novel mutations. PLoS ONE 6:e25614. https://doi.org/10.1371/journal.pone.0025614
Francou B, Paul C, Amazit L et al (2016) Prevalence of KISS1 Receptor mutations in a series of 603 patients with normosmic congenital hypogonadotrophic hypogonadism and characterization of novel mutations: a single-centre study. Hum Reprod 31:1363–1374. https://doi.org/10.1093/humrep/dew073
Fromantin M, Gineste J, Didier A, Rouvier J (1973) Impuberism and hypogonadism at induction into military service. Statistical study. Probl Actuels Endocrinol Nutr 16:179–199
Funes S, Hedrick JA, Vassileva G et al (2003) The KiSS-1 receptor GPR54 is essential for the development of the murine reproductive system. Biochem Biophys Res Commun 312:1357–1363. https://doi.org/10.1016/j.bbrc.2003.11.066
Galazzi E, Duminuco P, Moro M et al (2018) Hypogonadotropic hypogonadism and pituitary hypoplasia as recurrent features in Ulnar-Mammary syndrome. Endocr Connect 7:1432–1441. https://doi.org/10.1530/EC-18-0486
Gianetti E, Tusset C, Noel SD et al (2010) TAC3/TACR3 mutations reveal preferential activation of gonadotropin-releasing hormone release by neurokinin B in neonatal life followed by reversal in adulthood. J Clin Endocrinol Metab 95:2857–2867. https://doi.org/10.1210/jc.2009-2320
Gonçalves CI, Aragüés JM, Bastos M et al (2017) GNRHR biallelic and digenic mutations in patients with normosmic congenital hypogonadotropic hypogonadism. Endocr Connect 6:360–366. https://doi.org/10.1530/EC-17-0104
Gonçalves CI, Patriarca FM, Aragüés JM et al (2019) High frequency of CHD7 mutations in congenital hypogonadotropic hypogonadism. Sci Rep 9:1597. https://doi.org/10.1038/s41598-018-38178-y
Gonzalez-Martinez D (2004) Anosmin-1 modulates fibroblast growth factor receptor 1 signaling in human gonadotropin-releasing hormone olfactory neuroblasts through a heparan sulfate-dependent mechanism. J Neurosci 24:10384–10392. https://doi.org/10.1523/JNEUROSCI.3400-04.2004
Goodman RL, Lehman MN, Smith JT et al (2007) Kisspeptin neurons in the arcuate nucleus of the ewe express both dynorphin A and neurokinin B. Endocrinology 148:5752–5760. https://doi.org/10.1210/en.2007-0961
Gorbenko Del Blanco D, Romero CJ, Diaczok D et al (2012) A novel OTX2 mutation in a patient with combined pituitary hormone deficiency, pituitary malformation, and an underdeveloped left optic nerve. Eur J Endocrinol. https://doi.org/10.1530/EJE-12-0333
Gottsch ML, Cunningham MJ, Smith JT et al (2004) A role for kisspeptins in the regulation of gonadotropin secretion in the mouse. Endocrinology 145:4073–4077. https://doi.org/10.1210/en.2004-0431
Habiby RL, Boepple P, Nachtigall L et al (1996) Adrenal hypoplasia congenita with hypogonadotropic hypogonadism: evidence that DAX-1 mutations lead to combined hypothalamic and pituitary defects in gonadotropin production. J Clin Invest 98:1055–1062. https://doi.org/10.1172/JCI118866
Haines DE, Mihailoff GA, Parent AD, Perkins E (2018) The hypothalamus. Fundam Neurosci Basic Clin Appl 442–456:e1. https://doi.org/10.1016/B978-0-323-39632-5.00030-X
Hanafusa H, Torii S, Yasunaga T, Nishida E (2002) Sprouty1 and Sprouty2 provide a control mechanism for the Ras/MAPK signalling pathway. Nat Cell Biol 4:850–858. https://doi.org/10.1038/ncb867
Hanchate NK, Giacobini P, Lhuillier P et al (2012) SEMA3A, a gene involved in axonal pathfinding, is mutated in patients with kallmann syndrome. PLoS Genet 8:e1002896. https://doi.org/10.1371/journal.pgen.1002896
Hausott B, Vallant N, Schlick B et al (2012) Sprouty2 and -4 regulate axon outgrowth by hippocampal neurons. Hippocampus. https://doi.org/10.1002/hipo.20910
Henderson RH, Williamson KA, Kennedy JS et al (2009) A rare de novo nonsense mutation in OTX2 causes early onset retinal dystrophy and pituitary dysfunction. Mol Vis 15:2442–2447
Hirata T, Nakazawa M, Yoshihara S et al (2006) Zinc-finger gene Fez in the olfactory sensory neurons regulates development of the olfactory bulb non-cell-autonomously. Development 133:1433–1443. https://doi.org/10.1242/dev.02329
Hoffmann HM, Pandolfi EC, Larder R, Mellon PL (2019) Haploinsufficiency of homeodomain proteins Six3, Vax1, and Otx2 causes subfertility in mice via distinct mechanisms. Neuroendocrinology. https://doi.org/10.1159/000494086
Howard SR, Guasti L, Ruiz-Babot G et al (2016) IGSF 10 mutations dysregulate gonadotropin-releasing hormone neuronal migration resulting in delayed puberty. EMBO Mol Med 8:626–642. https://doi.org/10.15252/emmm.201606250
Howard SR, Oleari R, Poliandri A et al (2018) HS6ST1 insufficiency causes self-limited delayed puberty in contrast with other GnRH deficiency genes. J Clin Endocrinol Metab 103:3420–3429. https://doi.org/10.1210/jc.2018-00646
Huang H, Yang T, Shao Q et al (2018) Human TUBB3 mutations disrupt netrin attractive signaling. Neuroscience 374:155–171. https://doi.org/10.1016/j.neuroscience.2018.01.046
Hudson R, Laska M, Berger T et al (1994) Olfactory function in patients with hypogonadotropic hypogonadism: an all-or-none phenomenon? Chem Senses 19:57–69. https://doi.org/10.1093/chemse/19.1.57
Hutchins BI, Kotan LD, Taylor-Burds C et al (2016) CCDC141 mutation identified in anosmic hypogonadotropic hypogonadism (Kallmann Syndrome) alters GnRH neuronal migration. Endocrinology 157:1956–1966. https://doi.org/10.1210/en.2015-1846
Iacovazzo D, Carlsen E, Lugli F et al (2016) Factors predicting pasireotide responsiveness in somatotroph pituitary adenomas resistant to first-generation somatostatin analogues: an immunohistochemical study. Eur J Endocrinol 174:241–250. https://doi.org/10.1530/EJE-15-0832
Izumi Y, Suzuki E, Kanzaki S et al (2014) Genome-wide copy number analysis and systematic mutation screening in 58 patients with hypogonadotropic hypogonadism. Fertil Steril. https://doi.org/10.1016/j.fertnstert.2014.06.017
Jackson RS, Creemers JWM, Ohagi S et al (1997) Obesity and impaired prohormone processing associated with mutations in the human prohormone convertase 1 gene. Nat Genet. https://doi.org/10.1038/ng0797-303
Jarzabek K, Wolczynski S, Lesniewicz R et al (2012) Evidence that FGFR1 loss-of-function mutations may cause variable skeletal malformations in patients with Kallmann syndrome. Adv Med Sci. https://doi.org/10.2478/v10039-012-0036-4
Jayakody SA, Andoniadou CL, Gaston-Massuet C et al (2012) SOX2 regulates the hypothalamic-pituitary axis at multiple levels. J Clin Invest 122:3635–3646. https://doi.org/10.1172/JCI64311
Jongmans MCJ, Admiraal RJ, Van Der Donk KP et al (2006) CHARGE syndrome: the phenotypic spectrum of mutations in the CHD7 gene. J Med Genet. https://doi.org/10.1136/jmg.2005.036061
Jongmans MCJ, van Ravenswaaij-Arts CMA, Pitteloud N et al (2009) CHD7 mutations in patients initially diagnosed with Kallmann syndrome—the clinical overlap with CHARGE syndrome. Clin Genet. https://doi.org/10.1111/j.1399-0004.2008.01107.x
Känsäkoski J, Fagerholm R, Laitinen EM et al (2014) Mutation screening of SEMA3A and SEMA7A in patients with congenital hypogonadotropic hypogonadism. Pediatr Res 75:641–644. https://doi.org/10.1038/pr.2014.23
Kawamata N, Sakajiri S, Sugimoto K-J et al (2002) A novel chromosomal translocation t(1;14)(q25;q32) in pre-B acute lymphoblastic leukemia involves the LIM homeodomain protein gene, Lhx4. Oncogene 21:4983–4991. https://doi.org/10.1038/sj.onc.1205628
Kelberman D, Rizzoti K, Avilion A et al (2006) Mutations within Sox2/SOX2 are associated with abnormalities in the hypothalamo-pituitary-gonadal axis in mice and humans.e. J Clin Invest 116:2442–2455. https://doi.org/10.1172/JCI28658
Kelberman D, Turton JPG, Woods KS et al (2009) Molecular analysis of novel PROP1 mutations associated with combined pituitary hormone deficiency (CPHD). Clin Endocrinol (Oxf). https://doi.org/10.1111/j.1365-2265.2008.03326.x
Kevelam SHG, Van Harssel JJT, Van Der Zwaag B et al (2012) A patient with a mild holoprosencephaly spectrum phenotype and heterotaxy and a 1.3 Mb deletion encompassing GLI2. Am J Med Genet Part A. https://doi.org/10.1002/ajmg.a.34350
Kim H-G, Layman LC (2011) The role of CHD7 and the newly identified WDR11 gene in patients with idiopathic hypogonadotropic hypogonadism and Kallmann syndrome. Mol Cell Endocrinol 346:74–83. https://doi.org/10.1016/j.mce.2011.07.013
Kim HG, Kurth I, Lan F et al (2008a) Mutations in CHD7, encoding a chromatin-remodeling protein, cause idiopathic hypogonadotropic hypogonadism and Kallmann syndrome. Am J Hum Genet. https://doi.org/10.1016/j.ajhg.2008.09.005
Kim S-H, Hu Y, Cadman S, Bouloux P (2008b) Diversity in fibroblast growth factor receptor 1 regulation: learning from the investigation of Kallmann syndrome. J Neuroendocrinol 20:141–163. https://doi.org/10.1111/j.1365-2826.2007.01627.x
Kim H-G, Ahn J-W, Kurth I et al (2010) WDR11, a WD protein that interacts with transcription factor EMX1, is mutated in idiopathic hypogonadotropic hypogonadism and Kallmann syndrome. Am J Hum Genet 87:465–479. https://doi.org/10.1016/j.ajhg.2010.08.018
Kotan LD, Hutchins BI, Ozkan Y et al (2014) Mutations in FEZF1 cause Kallmann syndrome. Am J Hum Genet 95:326–331. https://doi.org/10.1016/j.ajhg.2014.08.006
Kramer PR, Wray S (2000) Novel gene expressed in nasal region influences outgrowth of olfactory axons and migration of luteinizing hormone-releasing hormone (LHRH) neurons. Genes Dev 14:1824–1834
Kramer PR, Wray S (2001) Nasal embryonic LHRH factor (NELF) expression within the CNS and PNS of the rodent. Brain Res Gene Expr Patterns 1:23–26
Kuang X-L, Zhao X-M, Xu H-F et al (2010) Spatio-temporal expression of a novel neuron-derived neurotrophic factor (NDNF) in mouse brains during development. BMC Neurosci 11:137. https://doi.org/10.1186/1471-2202-11-137
Kurokawa D, Kiyonari H, Nakayama R et al (2004) Regulation of Otx2 expression and its functions in mouse forebrain and midbrain. Development. https://doi.org/10.1242/dev.01220
Kyriakakis N, Shonibare T, Kyaw-Tun J et al (2017) Late-onset X-linked adrenal hypoplasia (DAX-1, NR0B1): two new adult-onset cases from a single center. Pituitary. https://doi.org/10.1007/s11102-017-0822-x
Laitinen E-M, Vaaralahti K, Tommiska J et al (2011) Incidence, phenotypic features and molecular genetics of Kallmann syndrome in Finland. Orphanet J Rare Dis 6:41. https://doi.org/10.1186/1750-1172-6-41
Lakhina V, Marcaccio CL, Shao X et al (2012) Netrin/DCC signaling guides olfactory sensory axons to their correct location in the olfactory bulb. J Neurosci 32:4440–4456. https://doi.org/10.1523/JNEUROSCI.4442-11.2012
Lalani SR, Safiullah AM, Molinari LM et al (2004) SEMA3E mutation in a patient with CHARGE syndrome. J Med Genet 41:e94. https://doi.org/10.1136/jmg.2003.017640
Layman LC, Lee EJ, Peak DB et al (1997) Delayed puberty and hypogonadism caused by mutations in the follicle-stimulating hormone β-subunit gene. N Engl J Med. https://doi.org/10.1056/NEJM199708283370905
Layman LC, Cohen DP, Jin M et al (1998) Mutations in gonadotropin-releasing hormone receptor gene cause hypogonadotropic hypogonadism. Nat Genet 18:14–15. https://doi.org/10.1038/ng0198-14
Layman LC, Porto ALA, Xie J et al (2002) FSHβ gene mutations in a female with partial breast development and a male sibling with normal puberty and azoospermia. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.87.8.3702
Lee GH, Proenca R, Montez JM et al (1996) Abnormal splicing of the leptin receptor in diabetic mice. Nature. https://doi.org/10.1038/379632a0
Legouis R, Hardelin JP, Levilliers J et al (1991) The candidate gene for the X-linked Kallmann syndrome encodes a protein related to adhesion molecules. Cell 67:423–435. https://doi.org/10.1016/0092-8674(91)90193-3
Leroy C, Fouveaut C, Leclercq S et al (2008) Biallelic mutations in the prokineticin-2 gene in two sporadic cases of Kallmann syndrome. Eur J Hum Genet 16:865–868. https://doi.org/10.1038/ejhg.2008.15
Lewkowitz-Shpuntoff HM, Hughes VA, Plummer L et al (2012) Olfactory phenotypic spectrum in idiopathic hypogonadotropic hypogonadism: pathophysiological and genetic implications. J Clin Endocrinol Metab 97:E136–E144. https://doi.org/10.1210/jc.2011-2041
Li C, Scott DA, Hatch E et al (2007) Dusp6 (Mkp3) is a negative feedback regulator of FgF-stimulated ERK signaling during mouse development. Development. https://doi.org/10.1242/dev.02701
Libri DV, Kleinau G, Vezzoli V et al (2014) Germline prokineticin receptor 2 (PROKR2) variants associated with central hypogonadism cause differental modulation of distinct intracellular pathways. J Clin Endocrinol Metab 99:E458–E463. https://doi.org/10.1210/jc.2013-2431
Liu G, Beggs H, Jürgensen C et al (2004) Netrin requires focal adhesion kinase and Src family kinases for axon outgrowth and attraction. Nat Neurosci 7:1222–1232. https://doi.org/10.1038/nn1331
Lo A, Zheng W, Gong Y et al (2011) GATA transcription factors regulate LHβ gene expression. J Mol Endocrinol 47:45–58. https://doi.org/10.1530/JME-10-0137
Lofrano-Porto A, Barra GB, Giacomini LA et al (2007) Luteinizing hormone beta mutation and hypogonadism in men and women. N Engl J Med. https://doi.org/10.1056/NEJMoa071999
Machinis K, Pantel J, Netchine I et al (2001) Syndromic short stature in patients with a germline mutation in the LIM homeobox LHX4. Am J Hum Genet 69:961–968. https://doi.org/10.1086/323764
Maione L, Brailly-Tabard S, Nevoux J et al (2016) Reversal of congenital hypogonadotropic hypogonadism in a man with Kallmann syndrome due to SOX10 mutation. Clin Endocrinol (Oxf) 85:988–989. https://doi.org/10.1111/cen.13231
Maione L, Dwyer AA, Francou B et al (2018) Genetic counseling for congenital hypogonadotropic hypogonadism and Kallmann syndrome: new challenges in the era of oligogenism and next-generation sequencing. Eur J Endocrinol 178(3):R55–R80 https://doi.org/10.1530/EJE-17-0749
Malone SA, Papadakis GE, Messina A et al (2019) Defective AMH signaling disrupts GnRH neuron development and function and contributes to hypogonadotropic hypogonadism. Elife. https://doi.org/10.7554/eLife.47198
Mantovani G, De Menis E, Borretta G et al (2006) DAX1 and X-linked adrenal hypoplasia congenita: clinical and molecular analysis in five patients. Eur J Endocrinol. https://doi.org/10.1530/eje.1.02132
Marcos S, Monnier C, Rovira X et al (2017) Defective signaling through plexin-A1 compromises the development of the peripheral olfactory system and neuroendocrine reproductive axis in mice. Hum Mol Genet 26:2006–2017. https://doi.org/10.1093/hmg/ddx080
Margolin DH, Kousi M, Chan Y-M et al (2013) Ataxia, dementia, and hypogonadotropism caused by disordered ubiquitination. N Engl J Med 368:1992–2003. https://doi.org/10.1056/NEJMoa1215993
Marsh APL, Heron D, Edwards TJ et al (2017) Mutations in DCC cause isolated agenesis of the corpus callosum with incomplete penetrance. Nat Genet 49:511–514. https://doi.org/10.1038/ng.3794
Matsumoto S-I, Yamazaki C, Masumoto K-H et al (2006) Abnormal development of the olfactory bulb and reproductive system in mice lacking prokineticin receptor PKR2. Proc Natl Acad Sci USA 103:4140–4145. https://doi.org/10.1073/pnas.0508881103
McCabe MJ, Gaston-Massuet C, Tziaferi V et al (2011a) Novel FGF8 mutations associated with recessive holoprosencephaly, craniofacial defects, and hypothalamo-pituitary dysfunction. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2011-0454
McCabe MJ, Gaston-Massuet C, Gregory LC et al (2013) Variations in PROKR2, but not PROK2, are associated with hypopituitarism and septo-optic dysplasia. J Clin Endocrinol Metab 98:E547–E557. https://doi.org/10.1210/jc.2012-3067
McCabe MJ, Hu Y, Gregory LC et al (2015) Novel application of luciferase assay for the in vitro functional assessment of KAL1 variants in three females with septo-optic dysplasia (SOD). Mol Cell Endocrinol 417:63–72. https://doi.org/10.1016/j.mce.2015.09.010
McCormack SE, Li D, Kim YJ et al (2017) Digenic inheritance of PROKR2 and WDR11 mutations in pituitary stalk interruption syndrome. J Clin Endocrinol Metab 102:2501–2507. https://doi.org/10.1210/jc.2017-00332
McNay DEG, Turton JP, Kelberman D et al (2007) HESX1 mutations are an uncommon cause of septooptic dysplasia and hypopituitarism. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2006-1609
Men M, Wu J, Zhao Y et al (2019) Genotypic and phenotypic spectra of FGFR1, FGF8, and FGF17 mutations in a Chinese cohort with idiopathic hypogonadotropic hypogonadism. Fertil Steril. https://doi.org/10.1016/j.fertnstert.2019.08.069
Mengen E, Tunc S, Kotan LD et al (2016) Complete idiopathic hypogonadotropic hypogonadism due to homozygous GNRH1 mutations in the mutational hot spots in the region encoding the decapeptide. Horm Res Paediatr 85:107–111. https://doi.org/10.1159/000441977
Messina A, Giacobini P (2013) Semaphorin signaling in the development and function of the gonadotropin hormone-releasing hormone system. Front Endocrinol (Lausanne) 4:133. https://doi.org/10.3389/fendo.2013.00133
Messina A, Ferraris N, Wray S et al (2011) Dysregulation of Semaphorin7A/β1-integrin signaling leads to defective GnRH-1 cell migration, abnormal gonadal development and altered fertility. Hum Mol Genet 20:4759–4774. https://doi.org/10.1093/hmg/ddr403
Messina A, Pulli K, Santini S et al (2020) Neuron-derived neurotrophic factor is mutated in congenital hypogonadotropic hypogonadism. Am J Hum Genet 106:58–70. https://doi.org/10.1016/j.ajhg.2019.12.003
Ming AYK, Yoo E, Vorontsov EN et al (2012) Dynamics and distribution of Klothoβ (KLB) and fibroblast growth factor receptor-1 (FGFR1) in living cells reveal the fibroblast growth factor-21 (FGF21)-induced receptor complex. J Biol Chem. https://doi.org/10.1074/jbc.M111.325670
Miraoui H, Dwyer AA, Sykiotis GP et al (2013) Mutations in FGF17, IL17RD, DUSP6, SPRY4, and FLRT3 are identified in individuals with congenital hypogonadotropic hypogonadism. Am J Hum Genet 92:725–743. https://doi.org/10.1016/j.ajhg.2013.04.008
Miura K, Acierno JS, Seminara SB (2004) Characterization of the human nasal embryonic LHRH factor gene, NELF, and a mutation screening among 65 patients with idiopathic hypogonadotropic hypogonadism (IHH). J Hum Genet 49:265–268. https://doi.org/10.1007/s10038-004-0137-4
Monnier C, Dodé C, Fabre L et al (2009) PROKR2 missense mutations associated with Kallmann syndrome impair receptor signalling activity. Hum Mol Genet 18:75–81. https://doi.org/10.1093/hmg/ddn318
Muscatelli F, Strom TM, Walker AP et al (1994) Mutations in the DAX-1 gene give rise to both X-linked adrenal hypoplasia congenita and hypogonadotropic hypogonadism. Nature. https://doi.org/10.1038/372672a0
Nachtigall LB, Boepple PA, Pralong FP, Crowley WF (1997) Adult-onset idiopathic hypogonadotropic hypogonadism—a treatable form of male infertility. N Engl J Med 336:410–415. https://doi.org/10.1056/NEJM199702063360604
Nakamura Y, Matsumoto H, Zaha K et al (2018) TUBB3 E410K syndrome with osteoporosis and cough syncope in a patient previously diagnosed with atypical Moebius syndrome. Brain Dev 40:233–237. https://doi.org/10.1016/j.braindev.2017.12.006
Navarro VM (2013) Interactions between kisspeptins and neurokinin B. Adv Exp Med Biol 784:325–347. https://doi.org/10.1007/978-1-4614-6199-9_15
Navarro VM, Gottsch ML, Chavkin C et al (2009) Regulation of gonadotropin-releasing hormone secretion by kisspeptin/dynorphin/neurokinin B neurons in the arcuate nucleus of the mouse. J Neurosci 29:11859–11866. https://doi.org/10.1523/JNEUROSCI.1569-09.2009
Newbern K, Natrajan N, Kim H-G et al (2013) Identification of HESX1 mutations in Kallmann syndrome. Fertil Steril 99:1831–1837. https://doi.org/10.1016/j.fertnstert.2013.01.149
Ng KL (2005) Dependence of olfactory bulb neurogenesis on Prokineticin 2 signaling. Science 308:1923–1927. https://doi.org/10.1126/science.1112103
Nickell MD, Breheny P, Stromberg AJ, McClintock TS (2012) Genomics of mature and immature olfactory sensory neurons. J Comp Neurol 520:2608–2629. https://doi.org/10.1002/cne.23052
Nunziata A, Borck G, Funcke J-B et al (2017) Estimated prevalence of potentially damaging variants in the leptin gene. Mol Cell Pediatr. https://doi.org/10.1186/s40348-017-0074-x
Oleari R, Caramello A, Campinoti S et al (2019a) PLXNA1 and PLXNA3 cooperate to pattern the nasal axons that guide gonadotropin-releasing hormone neurons. Development. https://doi.org/10.1242/dev.176461
Oleari R, Lettieri A, Paganoni A et al (2019b) Semaphorin signaling in GnRH neurons: from development to disease. Neuroendocrinology 109:193–199. https://doi.org/10.1159/000495916
Oliveira LM, Seminara SB, Beranova M et al (2001) The importance of autosomal genes in Kallmann syndrome: genotype-phenotype correlations and neuroendocrine characteristics. J Clin Endocrinol Metab 86:1532–1538. https://doi.org/10.1210/jcem.86.4.7420
Olsen SK, Li JYH, Bromleigh C et al (2006) Structural basis by which alternative splicing modulates the organizer activity of FGF8 in the brain. Genes Dev. https://doi.org/10.1101/gad.1365406
Olson LE, Tollkuhn J, Scafoglio C et al (2006) Homeodomain-mediated β-catenin-dependent switching events dictate cell-lineage determination. Cell. https://doi.org/10.1016/j.cell.2006.02.046
Palmert MR, Boepple PA (2001) Commentary: variation in the timing of puberty: clinical spectrum and genetic investigation. J Clin Endocrinol Metab 86:2364–2368
Pask AJ, Kanasaki H, Kaiser UB et al (2005) A novel mouse model of hypogonadotrophic hypogonadism: N-ethyl-N-nitrosourea-induced gonadotropin-releasing hormone receptor gene mutation. Mol Endocrinol 19:972–981. https://doi.org/10.1210/me.2004-0192
Pasterkamp RJ, De Winter F, Holtmaat AJ, Verhaagen J (1998) Evidence for a role of the chemorepellent semaphorin III and its receptor neuropilin-1 in the regeneration of primary olfactory axons. J Neurosci 18:9962–9976
Pfaeffle RW, Savage JJ, Hunter CS et al (2007) Four novel mutations of the LHX3 gene cause combined pituitary hormone deficiencies with or without limited neck rotation. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2006-2177
Pfaeffle RW, Hunter CS, Savage JJ et al (2008) Three novel missense mutations within the LHX4 gene are associated with variable pituitary hormone deficiencies. J Clin Endocrinol Metab 93:1062–1071. https://doi.org/10.1210/jc.2007-1525
Pingault V, Ente D, Le Moal FD et al (2010) Review and update of mutations causing Waardenburg syndrome. Hum Mutat 31(4):391–406. https://doi.org/10.1002/humu.21211
Pingault V, Bodereau V, Baral V et al (2013) Loss-of-function mutations in SOX10 cause Kallmann syndrome with deafness. Am J Hum Genet 92:707–724. https://doi.org/10.1016/j.ajhg.2013.03.024
Pitteloud N, Boepple PA, DeCruz S et al (2001) The fertile eunuch variant of idiopathic hypogonadotropic hypogonadism: spontaneous reversal associated with a homozygous mutation in the gonadotropin-releasing hormone receptor1. J Clin Endocrinol Metab 86:2470–2475. https://doi.org/10.1210/jcem.86.6.7542
Pitteloud N, Hayes FJ, Boepple PA et al (2002) The role of prior pubertal development, biochemical markers of testicular maturation, and genetics in elucidating the phenotypic heterogeneity of idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab 87:152–160. https://doi.org/10.1210/jcem.87.1.8131
Pitteloud N, Acierno JS, Meysing A et al (2006) Mutations in fibroblast growth factor receptor 1 cause both Kallmann syndrome and normosmic idiopathic hypogonadotropic hypogonadism. Proc Natl Acad Sci USA. https://doi.org/10.1073/pnas.0600962103
Pitteloud N, Quinton R, Pearce S et al (2007a) Digenic mutations account for variable phenotypes in idiopathic hypogonadotropic hypogonadism. J Clin Invest 117:457–463. https://doi.org/10.1172/JCI29884
Pitteloud N, Zhang C, Pignatelli D et al (2007b) Loss-of-function mutation in the prokineticin 2 gene causes Kallmann syndrome and normosmic idiopathic hypogonadotropic hypogonadism. Proc Natl Acad Sci USA 104:17447–17452. https://doi.org/10.1073/pnas.0707173104
Pitteloud N, Durrani S, Raivio T, Sykiotis GP (2010) Complex genetics in idiopathic hypogonadotropic hypogonadism. Kallmann syndrome and hypogonadotropic hypogonadism. KARGER, Basel, pp 142–153
Pozza SBD, Hiedl S, Roeb J et al (2012) A recessive mutation resulting in a disabling amino acid substitution (T194R) in the LHX3 homeodomain causes combined pituitary hormone deficiency. Horm Res Paediatr 77(1):41–51. https://doi.org/10.1159/000335929
Prosser HM, Bradley A, Caldwell MA (2007) Olfactory bulb hypoplasia in Prokr2 null mice stems from defective neuronal progenitor migration and differentiation. Eur J Neurosci 26:3339–3344.https://doi.org/10.1111/j.1460-9568.2007.05958.x
Quennell JH, Mulligan AC, Tups A et al (2009) Leptin indirectly regulates gonadotropin-releasing hormone neuronal function. Endocrinology. https://doi.org/10.1210/en.2008-1693
Quinton R, Cheow HK, Tymms DJ et al (1999) Kallmann’s syndrome: is it always for life? Clin Endocrinol (Oxf) 50:481–485. https://doi.org/10.1046/j.1365-2265.1999.00708.x
Quinton R, Duke VM, Robertson A et al (2001) Idiopathic gonadotrophin deficiency: genetic questions addressed through phenotypic characterization. Clin Endocrinol (Oxf) 55:163–174. https://doi.org/10.1046/j.1365-2265.2001.01277.x
Raivio T, Falardeau J, Dwyer A et al (2007) Reversal of idiopathic hypogonadotropic hypogonadism. N Engl J Med 357:863–873. https://doi.org/10.1056/NEJMoa066494
Raivio T, Avbelj M, McCabe MJ et al (2012) Genetic overlap in kallmann syndrome, combined pituitary hormone deficiency, and septo-optic dysplasia. J Clin Endocrinol Metab 97:E694–E699. https://doi.org/10.1210/jc.2011-2938
Ribeiro RS, Vieira TC, Abucham J (2007) Reversible Kallmann syndrome: report of the first case with a KAL1 mutation and literature review. Eur J Endocrinol 156:285–290. https://doi.org/10.1530/eje.1.02342
Richards S, Aziz N, Bale S et al (2015) Standards and guidelines for the interpretation of sequence variants: a joint consensus recommendation of the American College of Medical Genetics and Genomics and the Association for Molecular Pathology. Genet Med 17:405–424. https://doi.org/10.1038/gim.2015.30
Robinson M, Parsons Perez MC, Tébar L et al (2004) FLRT3 is expressed in sensory neurons after peripheral nerve injury and regulates neurite outgrowth. Mol Cell Neurosci. https://doi.org/10.1016/j.mcn.2004.06.008
Romero CJ, Pine-Twaddell E, Radovick S (2011) Novel mutations associated with combined pituitary hormone deficiency. J Mol Endocrinol 46(3):R93–R102. https://doi.org/10.1530/JME-10-0133
Root AW (2010) Reversible isolated hypogonadotropic hypogonadism due to mutations in the neurokinin B regulation of gonadotropin-releasing hormone release. J Clin Endocrinol Metab 95:2625–2629. https://doi.org/10.1210/jc.2010-0733
Ross RA, Leon S, Madara JC et al (2018) PACAP neurons in the ventral premammillary nucleus regulate reproductive function in the female mouse. Elife. https://doi.org/10.7554/eLife.35960
de Roux N (2006) GnRH receptor and GPR54 inactivation in isolated gonadotropic deficiency. Best Pract Res Clin Endocrinol Metab 20:515–528. https://doi.org/10.1016/j.beem.2006.10.005
de Roux N, Young J, Misrahi M et al (1997) A family with hypogonadotropic hypogonadism and mutations in the gonadotropin-releasing hormone receptor. N Engl J Med 337:1597–1603. https://doi.org/10.1056/NEJM199711273372205
de Roux N, Young J, Brailly-Tabard S et al (1999) The same molecular defects of the gonadotropin-releasing hormone receptor determine a variable degree of hypogonadism in affected kindred1. J Clin Endocrinol Metab 84:567–572. https://doi.org/10.1210/jcem.84.2.5449
de Roux N, Genin E, Carel J-C et al (2003) Hypogonadotropic hypogonadism due to loss of function of the KiSS1-derived peptide receptor GPR54. Proc Natl Acad Sci 100:10972–10976. https://doi.org/10.1073/pnas.1834399100
de Tassigny XA, Fagg LA, Dixon JPC et al (2007) Hypogonadotropic hypogonadism in mice lacking a functional Kiss1 gene. Proc Natl Acad Sci 104:10714–10719. https://doi.org/10.1073/pnas.0704114104
Rugarli EI, Ghezzi C, Valsecchi V, Ballabio A (1996) The Kallmann syndrome gene product expressed in COS cells is cleaved on the cell surface to yield a diffusible component. Hum Mol Genet 5:1109–1115. https://doi.org/10.1093/hmg/5.8.1109
Salenave S, Chanson P, Bry H et al (2008) Kallmann’s syndrome: a comparison of the reproductive phenotypes in men carrying KAL1 and FGFR1/KAL2 mutations. J Clin Endocrinol Metab 93:758–763. https://doi.org/10.1210/jc.2007-1168
Santhakumar A, Balasubramanian R, Miller M, Quinton R (2014) Reversal of isolated hypogonadotropic hypogonadism: long-term integrity of hypothalamo-pituitary-testicular axis in two men is dependent on intermittent androgen exposure. Clin Endocrinol (Oxf) 81:473–476
Sarfati J, Dodé C, Young J (2010) Kallmann syndrome caused by mutations in the PROK2 and PROKR2 genes: pathophysiology and genotype-phenotype correlations. Front Horm Res 39:121–132. https://doi.org/10.1159/000312698
Sasaki G, Ogata T, Ishii T et al (2002) Novel mutation of TBX3 in a Japanese family with ulnar-mammary syndrome: implication for impaired sex development. Am J Med Genet 110:365–369. https://doi.org/10.1002/ajmg.10447
Sato N, Katsumata N, Kagami M et al (2004) Clinical assessment and mutation analysis of Kallmann syndrome 1 (KAL1) and fibroblast growth factor receptor 1 (FGFR1, or KAL2) in five families and 18 sporadic patients. J Clin Endocrinol Metab 89:1079–1088. https://doi.org/10.1210/jc.2003-030476
Sbai O, Monnier C, Dodé C et al (2014) Biased signaling through G-protein-coupled PROKR2 receptors harboring missense mutations. FASEB J 28:3734–3744. https://doi.org/10.1096/fj.13-243402
Schang AL, Granger A, Quérat B et al (2013) GATA2-induced silencing and LIM-homeodomain protein-induced activation are mediated by a Bi-functional response element in the rat GnRH receptor gene. Mol Endocrinol. https://doi.org/10.1210/me.2012-1182
Schilter KF, Schneider A, Bardakjian T et al (2011) OTX2 microphthalmia syndrome: four novel mutations and delineation of a phenotype. Clin Genet. https://doi.org/10.1111/j.1399-0004.2010.01450.x
Schwanzel-Fukuda M, Bick D, Pfaff DW (1989) Luteinizing hormone-releasing hormone (LHRH)-expressing cells do not migrate normally in an inherited hypogonadal (Kallmann) syndrome. Mol Brain Res 6:311–326. https://doi.org/10.1016/0169-328X(89)90076-4
Schwarting GA, Raitcheva D, Bless EP et al (2004) Netrin 1-mediated chemoattraction regulates the migratory pathway of LHRH neurons. Eur J Neurosci 19:11–20. https://doi.org/10.1111/j.1460-9568.2004.03094.x
Seeburg PH, Adelman JP (1984) Characterization of cDNA for precursor of human luteinizing hormone releasing hormone. Nature 311:666–668. https://doi.org/10.1038/311666a0
Seifi M, Walter MA (2018) Axenfeld-Rieger syndrome. Clin Genet 93:1123–1130. https://doi.org/10.1111/cge.13148
Seminara SB, Acierno JS, Abdulwahid NA et al (2002) Hypogonadotropic hypogonadism and cerebellar ataxia: detailed phenotypic characterization of a large, extended kindred. J Clin Endocrinol Metab 87:1607–1612. https://doi.org/10.1210/jcem.87.4.8384
Seminara SB, Messager S, Chatzidaki EE et al (2003) The GPR54 gene as a regulator of puberty. N Engl J Med 349:1614–1627. https://doi.org/10.1056/NEJMoa035322
Semple RK, Achermann JC, Ellery J et al (2005) Two novel missense mutations in G protein-coupled receptor 54 in a patient with hypogonadotropic hypogonadism. J Clin Endocrinol Metab 90:1849–1855. https://doi.org/10.1210/jc.2004-1418
Shaw ND, Brand H, Kupchinsky ZA et al (2017) SMCHD1 mutations associated with a rare muscular dystrophy can also cause isolated arhinia and Bosma arhinia microphthalmia syndrome. Nat Genet 49:238–248. https://doi.org/10.1038/ng.3743
Sheng HZ, Moriyama K, Yamashita T et al (1997) Multistep control of pituitary organogenesis. Science 278:1809–1812. https://doi.org/10.1126/science.278.5344.1809
Shi C-H, Schisler JC, Rubel CE et al (2014) Ataxia and hypogonadism caused by the loss of ubiquitin ligase activity of the U box protein CHIP. Hum Mol Genet 23:1013–1024. https://doi.org/10.1093/hmg/ddt497
Sidhoum VF, Chan Y-M, Lippincott MF et al (2014) Reversal and relapse of hypogonadotropic hypogonadism: resilience and fragility of the reproductive neuroendocrine system. J Clin Endocrinol Metab 99:861–870. https://doi.org/10.1210/jc.2013-2809
Simonis N, Migeotte I, Lambert N et al (2013) FGFR1 mutations cause hartsfield syndrome, the unique association of holoprosencephaly and ectrodactyly. J Med Genet. https://doi.org/10.1136/jmedgenet-2013-101603
Sinisi AA, Asci R, Bellastella G et al (2008) Homozygous mutation in the prokineticin-receptor2 gene (Val274Asp) presenting as reversible Kallmann syndrome and persistent oligozoospermia: case report. Hum Reprod 23:2380–2384. https://doi.org/10.1093/humrep/den247
Sisodiya SM, Ragge NK, Cavalleri GL et al (2006) Role of SOX2 mutations in human hippocampal malformations and epilepsy. Epilepsia 47:534–542. https://doi.org/10.1111/j.1528-1167.2006.00464.x
Smith JT, Acohido BV, Clifton DK, Steiner RA (2006) KiSS-1 neurones are direct targets for leptin in the ob/ob mouse. J Neuroendocrinol 18:298–303. https://doi.org/10.1111/j.1365-2826.2006.01417.x
Soussi-Yanicostas N, Hardelin JP, Arroyo-Jimenez MM et al (1996) Initial characterization of anosmin-1, a putative extracellular matrix protein synthesized by definite neuronal cell populations in the central nervous system. J Cell Sci 109(Pt 7):1749–1757
Soussi-Yanicostas N, de Castro F, Julliard AK et al (2002) Anosmin-1, defective in the X-linked form of Kallmann syndrome, promotes axonal branch formation from olfactory bulb output neurons. Cell 109:217–228. https://doi.org/10.1016/s0092-8674(02)00713-4
Spilker C, Grochowska KM, Kreutz MR (2016) What do we learn from the murine Jacob/Nsmf gene knockout for human disease? Rare Dis (Austin, Tex) 4:e1241361. https://doi.org/10.1080/21675511.2016.1241361
Srour M, Riviere JB, Pham JMT et al (2010) Mutations in DCC cause congenital mirror movements. Science 328:592–592. https://doi.org/10.1126/science.1186463
Stamou MI, Georgopoulos NA (2018) Kallmann syndrome: phenotype and genotype of hypogonadotropic hypogonadism. Metabolism 86:124–134. https://doi.org/10.1016/j.metabol.2017.10.012
Stamou MI, Varnavas P, Plummer L et al (2019) Next-generation sequencing refines the genetic architecture of Greek GnRH-deficient patients. Endocr Connect 8:468–480. https://doi.org/10.1530/EC-19-0010
Steevens AR, Sookiasian DL, Glatzer JC, Kiernan AE (2017) SOX2 is required for inner ear neurogenesis. Sci Rep 7:4086. https://doi.org/10.1038/s41598-017-04315-2
Stenson PD, Mort M, Ball EV et al (2014) The human gene mutation database: building a comprehensive mutation repository for clinical and molecular genetics, diagnostic testing and personalized genomic medicine. Hum Genet 133:1–9
Suh H, Gage PJ, Drouin J, Camper SA (2002) Pitx2 is required at multiple stages of pituitary organogenesis: pituitary primordium formation and cell specification. Development 129(2):329–337
Swee DS, Quinton R (2019a) Congenital hypogonadotrophic hypogonadism: minipuberty and the case for neonatal diagnosis. Front Endocrinol (Lausanne) 10:97. https://doi.org/10.3389/fendo.2019.00097
Swee DS, Quinton R (2019b) Managing congenital hypogonadotrophic hypogonadism: a contemporary approach directed at optimizing fertility and long-term outcomes in males. Ther Adv Endocrinol Metab. https://doi.org/10.1177/2042018819826889
Sykiotis GP, Plummer L, Hughes VA et al (2010) Oligogenic basis of isolated gonadotropin-releasing hormone deficiency. Proc Natl Acad Sci 107:15140–15144. https://doi.org/10.1073/pnas.1009622107
Synofzik M, Gonzalez MA, Lourenco CM et al (2014) PNPLA6 mutations cause Boucher-Neuhauser and Gordon Holmes syndromes as part of a broad neurodegenerative spectrum. Brain 137:69–77. https://doi.org/10.1093/brain/awt326
Tajima T, Ishizu K, Nakamura A (2013) Molecular and clinical findings in patients with LHX4 and OTX2 mutations. Clin Pediatr Endocrinol 22(2):15–23. https://doi.org/10.1292/cpe.22.15
Takagi M, Narumi S, Hamada R et al (2014) A novel KAL1 mutation is associated with combined pituitary hormone deficiency. Hum Genome Var 1:14011. https://doi.org/10.1038/hgv.2014.11
Taniguchi K, Ayada T, Ichiyama K et al (2007) Sprouty2 and Sprouty4 are essential for embryonic morphogenesis and regulation of FGF signaling. Biochem Biophys Res Commun. https://doi.org/10.1016/j.bbrc.2006.11.107
Tata B, Huijbregts L, Jacquier S et al (2014) Haploinsufficiency of Dmxl2, encoding a synaptic protein, causes infertility associated with a loss of GnRH neurons in mouse. PLoS Biol 12:e1001952. https://doi.org/10.1371/journal.pbio.1001952
Teles MG, Bianco SDC, Brito VN et al (2008) A GPR54-activating mutation in a patient with central precocious puberty. N Engl J Med 358:709–715. https://doi.org/10.1056/NEJMoa073443
Tenenbaum-Rakover Y, Commenges-Ducos M, Iovane A et al (2007) Neuroendocrine phenotype analysis in five patients with isolated hypogonadotropic hypogonadism due to a L102P inactivating mutation of GPR54. J Clin Endocrinol Metab 92:1137–1144. https://doi.org/10.1210/jc.2006-2147
Tétreault M, Choquet K, Orcesi S et al (2011) Recessive mutations in POLR3B, encoding the second largest subunit of Pol III, cause a rare hypomyelinating leukodystrophy. Am J Hum Genet 89:652–655. https://doi.org/10.1016/j.ajhg.2011.10.006
Tiong J, Locastro T, Wray S (2007) Gonadotropin-releasing hormone-1 (GnRH-1) is involved in tooth maturation and biomineralization. Dev Dyn 236:2980–2992. https://doi.org/10.1002/dvdy.21332
Tommiska J, Känsäkoski J, Christiansen P et al (2014) Genetics of congenital hypogonadotropic hypogonadism in Denmark. Eur J Med Genet 57:345–348. https://doi.org/10.1016/j.ejmg.2014.04.002
Topaloglu AK, Reimann F, Guclu M et al (2009) TAC3 and TACR3 mutations in familial hypogonadotropic hypogonadism reveal a key role for Neurokinin B in the central control of reproduction. Nat Genet 41:354–358. https://doi.org/10.1038/ng.306
Topaloglu AK, Tello JA, Kotan LD et al (2012) Inactivating KISS1 mutation and hypogonadotropic hypogonadism. N Engl J Med 366:629–635. https://doi.org/10.1056/NEJMoa1111184
Topaloglu AK, Lomniczi A, Kretzschmar D et al (2014) Loss-of-function mutations in PNPLA6 encoding neuropathy target esterase underlie pubertal failure and neurological deficits in Gordon Holmes syndrome. J Clin Endocrinol Metab 99:E2067–E2075. https://doi.org/10.1210/jc.2014-1836
Tornberg J, Sykiotis GP, Keefe K et al (2011) Heparan sulfate 6-O-sulfotransferase 1, a gene involved in extracellular sugar modifications, is mutated in patients with idiopathic hypogonadotrophic hypogonadism. Proc Natl Acad Sci USA 108:11524–11529. https://doi.org/10.1073/pnas.1102284108
Trarbach EB, Silveira LG, Latronico AC (2007) Genetic insights into human isolated gonadotropin deficiency. Pituitary 10:381–391. https://doi.org/10.1007/s11102-007-0061-7
Tsuchida T, Ensini M, Morton SB et al (1994) Topographic organization of embryonic motor neurons defined by expression of LIM homeobox genes. Cell. https://doi.org/10.1016/0092-8674(94)90027-2
Turan I, Hutchins BI, Hacihamdioglu B et al (2017) CCDC141 mutations in idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab 102:1816–1825. https://doi.org/10.1210/jc.2016-3391
Turton JPG, Mehta A, Raza J et al (2005) Mutations within the transcription factor PROP1 are rare in a cohort of patients with sporadic combined pituitary hormone deficiency (CPHD). Clin Endocrinol (Oxf). https://doi.org/10.1111/j.1365-2265.2005.02291.x
Vaaralahti K, Tommiska J, Tillmann V et al (2014) De novo SOX10 nonsense mutation in a patient with Kallmann syndrome and hearing loss. Pediatr Res 76:115–116. https://doi.org/10.1038/pr.2014.60
Valdes-Socin H, Salvi R, Thiry A et al (2009) Testicular effects of isolated luteinizing hormone deficiency and reversal by long-term human chorionic gonadotropin treatment. J Clin Endocrinol Metab. https://doi.org/10.1210/jc.2008-1584
Vezzoli V, Duminuco P, Bassi I et al (2016) The complex genetic basis of congenital hypogonadotropic hypogonadism. Minerva Endocrinol 41:223–239
Villanueva C, Jacobson-Dickman E, Xu C et al (2015) Congenital hypogonadotropic hypogonadism with split hand/foot malformation: a clinical entity with a high frequency of FGFR1 mutations. Genet Med. https://doi.org/10.1038/gim.2014.166
Wakabayashi Y, Nakada T, Murata K et al (2010) Neurokinin B and dynorphin A in kisspeptin neurons of the arcuate nucleus participate in generation of periodic oscillation of neural activity driving pulsatile gonadotropin-releasing hormone secretion in the goat. J Neurosci 30:3124–3132. https://doi.org/10.1523/JNEUROSCI.5848-09.2010
Wang Y, Gong C, Qin M et al (2017) Clinical and genetic features of 64 young male paediatric patients with congenital hypogonadotropic hypogonadism. Clin Endocrinol (Oxf) 87:757–766. https://doi.org/10.1111/cen.13451
Wang F, Zhao S, Xie Y et al (2018) De novo SOX10 nonsense mutation in a patient with Kallmann syndrome, deafness, iris hypopigmentation, and hyperthyroidism. Ann Clin Lab Sci 48:248–252
Watanabe Y, Inoue K, Okuyama-Yamamoto A et al (2009) Fezf1 is required for penetration of the basal lamina by olfactory axons to promote olfactory development. J Comp Neurol 515:565–584. https://doi.org/10.1002/cne.22074
Weiss J, Adams E, Whitcomb RW et al (1991) Normal sequence of the gonadotropin-releasing hormone gene in patients with idiopathic hypogonadotropic hypogonadism. Biol Reprod 45:743–747. https://doi.org/10.1095/biolreprod45.5.743
Welt CK, Chan JL, Bullen J et al (2004) Recombinant human leptin in women with hypothalamic amenorrhea. N Engl J Med 351:987–997. https://doi.org/10.1056/NEJMoa040388
Whitman MC, Andrews C, Chan W-M et al (2016) Two unique TUBB3 mutations cause both CFEOM3 and malformations of cortical development. Am J Med Genet A 170A:297–305. https://doi.org/10.1002/ajmg.a.37362
Williamson KA, Hever AM, Rainger J et al (2006) Mutations in SOX2 cause anophthalmia-esophageal-genital (AEG) syndrome. Hum Mol Genet 15:1413–1422. https://doi.org/10.1093/hmg/ddl064
Wray S (2010) From nose to brain: development of gonadotrophin-releasing hormone -1 neurones. J Neuroendocrinol 22(7):743–753. https://doi.org/10.1111/j.1365-2826.2010.02034.x
Wu S, Wilson MD, Busby ER et al (2010) Disruption of the single copy gonadotropin-releasing hormone receptor in mice by gene trap: severe reduction of reproductive organs and functions in developing and adult mice. Endocrinology 151:1142–1152. https://doi.org/10.1210/en.2009-0598
Wyatt A, Bakrania P, Bunyan DJ et al (2008) Novel heterozygous OTX2 mutations and whole gene deletions in anophthalmia, microphthalmia and coloboma. Hum Mutat. https://doi.org/10.1002/humu.20869
Xu N, Bhagavath B, Kim H-G et al (2010) NELF is a nuclear protein involved in hypothalamic GnRH neuronal migration. Mol Cell Endocrinol 319:47–55. https://doi.org/10.1016/j.mce.2009.11.016
Xu N, Kim H-G, Bhagavath B et al (2011) Nasal embryonic LHRH factor (NELF) mutations in patients with normosmic hypogonadotropic hypogonadism and Kallmann syndrome. Fertil Steril 95(1613–20):e1–7. https://doi.org/10.1016/j.fertnstert.2011.01.010
Xu C, Messina A, Somm E et al (2017) KLB, encoding β-Klotho, is mutated in patients with congenital hypogonadotropic hypogonadism. EMBO Mol Med 9:1379–1397. https://doi.org/10.15252/emmm.201607376
Young J, Bouligand J, Francou B et al (2010) TAC3 and TACR3 defects cause hypothalamic congenital hypogonadotropic hypogonadism in humans. J Clin Endocrinol Metab 95:2287–2295. https://doi.org/10.1210/jc.2009-2600
Young J, Metay C, Bouligand J et al (2012) SEMA3A deletion in a family with Kallmann syndrome validates the role of semaphorin 3A in human puberty and olfactory system development. Hum Reprod 27:1460–1465. https://doi.org/10.1093/humrep/des022
Young J, Xu C, Papadakis GE et al (2019) Clinical management of congenital hypogonadotropic hypogonadism. Endocr Rev 40:669–710. https://doi.org/10.1210/er.2018-00116
Zhang S, Cui W (2014) Sox2, a key factor in the regulation of pluripotency and neural differentiation. World J Stem Cells 6:305–311. https://doi.org/10.4252/wjsc.v6.i3.305
Zhang Y, Proenca R, Maffei M et al (1994) Positional cloning of the mouse obese gene and its human homologue. Nature. https://doi.org/10.1038/372425a0
Zhu J, Choa RE-Y, Guo MH et al (2015) A shared genetic basis for self-limited delayed puberty and idiopathic hypogonadotropic hypogonadism. J Clin Endocrinol Metab 100:E646–E654. https://doi.org/10.1210/jc.2015-1080
Acknowledgements
Researches from Prof. M. Bonomi were partially supported by funds from IRCCS Istituto Auxologico Italiano (Ricerca Corrente funds: O5C202_2012); from Dept of Clinical Sciences and Community Health (Piano di Sostegno alla Ricerca—Linea 2: RV_RIC_AT18MBONO).
Author information
Authors and Affiliations
Corresponding authors
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Biagio Cangiano: ResID: K-1236-2018 and Marco Bonomi: ResID: T-6487-2019.
Rights and permissions
About this article

Cite this article
Cangiano, B., Swee, D.S., Quinton, R. et al. Genetics of congenital hypogonadotropic hypogonadism: peculiarities and phenotype of an oligogenic disease. Hum Genet 140, 77–111 (2021). https://doi.org/10.1007/s00439-020-02147-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00439-020-02147-1