
Abstract
Tropomyosin (TM) is a major allergen in shellfish, known to cross-react with mite, cockroach and/or some roundworm (nematode) TM. In this study, we aimed to express and purify TM from the parasitic nematode Anisakis pegreffii and also to characterise its cross-reactivity with TM from shellfish. A. pegreffii was isolated from the flathead tiger fish (Neoplatycephalus richardsoni) and characterised using single-strand conformation polymorphism (SSCP)-based sequencing of the first and second internal transcribed spacers (ITS-1 and ITS-2) of nuclear ribosomal DNA. The recombinant tropomyosin (rTM) of A. pegreffii was expressed, purified and confirmed by immunohistochemistry, sequencing and LC-MS/MS analyses. Immunohistochemistry showed the muscle and the base layer of the third-stage larvae (L3) of A. pegreffii as the location of TM in A. pegreffii. The molecular relationship of TM of A. pegreffii with homologs from other nematodes and crustaceans was inferred from phylogenetic analysis. Immunogenicity of TM from A. pegreffii was tested by immunoblotting, which showed that rTM from A. pegreffii binds to IgE from sera of patients with allergy to crustaceans. Immunoblotting also showed that the anti-TM monoclonal antibody (MAb) did not recognise rTM from A. pegreffii. The rTM from A. pegreffii was, however, recognised by anti-TM polyclonal antibodies (PAbs) as well as anti-crustacean polyclonal antibodies (PAbs). The detection of specific serum IgE antibody against parasite TM has been proposed as a useful approach for the diagnosis of parasite-induced allergy. The findings of this study merit further exploration of the cross-reactive allergenic proteins of Anisakis for improved, future diagnosis of allergenic diseases.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The third-stage larvae of the nematode worm Anisakis present a major economic problem for the fish-trade industry (Mattiucci et al. 1997; Mattiucci and Nascetti, 2008). They are also identified as potential threats to human health causing anisakiasis and as potential allergenic food-borne pathogens (Bao et al. 2017; Fumarola et al. 2009; Gregori et al. 2015; Kirstein et al. 2010; Umehara et al. 2007). Anisakis simplex third-stage larva (L3) is classified as Anisakis type I and has been regarded as the primary cause of anisakiasis, mostly acquired through the ingestion of infected, raw fish (Daschner et al. 2002). However, molecular studies have revealed that A. simplex represents at least three sibling species, namely A. simplex sensu stricto (s.s.) (Nascetti et al. 1986), A. pegreffii (Campana-Rouget and Biocca, 1955) and A. simplex C (Mattiucci et al. 2002; Mattiucci and Nascetti, 2007; Nascetti et al. 1986). It is now established that there is a wide distribution of A. pegreffii and A. simplex s.s. across the globe. They are characterised on the basis of their genetic and morphological features as well as their ecological traits, such as preference for the definitive host. These parasitic nematodes do have geographical distributions; they, however, do not have a preference for their distribution. Results of various researches worldwide have also shown A. simplex s.s. and A. pegreffii as the principal etiologic agents of anisakiasis (Abollo et al. 2003; Mattiucci et al. 1997, Mattiucci and Nascetti, 2006; 2008; Mattiucci et al. 2009).
Parasite hosts are constantly being challenged by the large numbers of somatic and excretory/secretory antigens produced by parasites, which in some cases would induce strong allergic reactions in hosts to these antigens (Acevedo and Caraballo 2011). Allergic reactions to Anisakis have been reported, and these allergens from Anisakis are considered to be of importance (Albrecht et al. 2008; Anibarro et al. 1998; Kamath et al. 2013; Kim et al. 2005).
Tropomyosin (TM), a major allergen in shellfish, is known to be an isoallergen and panallergen, which is cross-reactive with mite, cockroach and some nematode homologs (Lopata and Lehrer, 2009; Lopata et al. 2010). In many species of crustaceans, TM has been identified and found to be implicated in ~ 80% of cases associated with allergic reactions (Faeste et al. 2003). Functionally, shellfish TM shares various highly conserved IgE-binding B cell epitopes. These epitopes are known to induce immune responses and, in addition, also have a high cross-reactivity with TM of invertebrates of diverse species. In A. simplex, TM has been described as a major somatic allergen present in third- and fourth-stage larvae (O’Donnell et al. 1989). The proteins of A. simplex have been shown to display notable cross-reactivity to proteins of other invertebrates, such as house dust mites and crustaceans as well as related nematodes (Nieuwenhuizen and Lopata, 2013).
This study therefore aimed to characterise the TM of A. pegreffii and to determine whether TM of A. pegreffii shares epitopes with TM of A. simplex, and other organisms, particularly crustaceans. Results from this work will be used to infer the allergenicity of TM from A. pegreffii.
Material and methods
Isolation of anisakid larvae and morphological identification
Live larvae (L3) of A. pegreffii were recovered from the uppermost layer of the internal cavity and organs such as gut, pyloric caecum, stomach, gonads and liver of parasitised Neoplatycephalus richardsoni (flathead fish). The fish were purchased from a local seafood market in Melbourne and transported on ice to the laboratory for immediate processing. Encapsulated larvae were isolated from the fish, and larvae were freed from the host tissue as described by Werner et al. (2011). Briefly, capsules were opened and removed to liberate the larvae. Larvae were thereafter exhaustively washed in phosphate-buffered saline (PBS; pH 7.4) and then fixed in ethanol for subsequent examination. The anterior and posterior parts of each nematode were cut out with a sterile scalpel and preserved in lactophenol for morphological identification. The mid-section (3 mm) of the body was used for subsequent molecular studies. Using the light optical microscope, nematodes were examined morphologically and assigned to a larval ‘type’ (Cannon, 1977; Shamsi et al., 2011). Briefly, the presence or absence of mucron at the tail end as well as the length of the ventriculus was assessed to assign the larval type.
Molecular identification of anisakid larvae
Sodium dodecyl–sulphate/proteinase K treatment was used to extract genomic DNA from the mid-section of the larvae. This was purified using a mini-column according to the manufacturer’s protocol (Wizard DNA Clean-Up System, Promega, Fitchburg, WI, USA). Amplification of ITS-1 and ITS-2 nuclear rDNA, from 10 to 50 ng of genomic DNA, was performed in a PCR using the primer sets SS1/NC13R and SS2/NC2 respectively (Shamsi et al. 2008; Zhang et al., 2007). PCR conditions were as previously described (Jabbar et al. 2012). Known positive controls and negatives (no-DNA) were included for each set of PCRs. Amplicon (5 μl), following PCR, was examined on an ethidium bromide-stained 1.5% w/v agarose gel, and thereafter visualised using ultraviolet light.
Single-strand conformation polymorphism (SSCP) and sequencing
In order to determine sequence variation within and among amplicons, ITS-1 and ITS-2 amplified products were subjected to SSCP analysis (Gasser et al. 2006; protocol B). Amplicon representatives for unique SSCP profile for each locus were selected and treated with exonuclease I and shrimp alkaline phosphatase (Fermentas Inc., Glen Burnie, MD, USA). These were subsequently sequenced, using the same PCR primers, in a bi-directional, automated sequencing (Big-Dye terminator v.3.1, Applied Biosystems, Foster City, CA, USA). Each sequence quality was assessed by evaluating its electropherogram with the Bio-Edit program (Hall, 1999). Polymorphic sites were assigned using International Union of Pure and Applied Chemistry (IUPAC) codes. Reference sequences, AB277823 (representing A. pegreffii L3s from Scomber japonicus (Pacific chub mackerel)) (Umehara et al. 2008) and JQ900763 from Engraulis spp. (anchovy) (Mattiucci et al. 2013), from the GenBank database were compared with selected sequences.
Preparation of protein extracts
An extract was made from A. pegreffii by snap-freezing the posterior and anterior portions of 30 L3s (25 mg) in liquid nitrogen, homogenising the extract in 2.5 ml of a solution of 0.1 M Tris (hydroxymethy1) aminomethane and 0.5 M glycine buffer (pH 8.7). This extract was then sonicated using a probe sonicator (Branson digital sonifier, 35% amplitude; pulse on 15 s, pulse off 30 s; three cycles), and thereafter centrifuged for 5 min at 10,000×g to remove debris. The supernatant was filtered using a 0.45-μm membrane filter (Millipore, Billerica, MA, USA) and supernatant protein concentration was performed using Millipore concentration tubes (5 kDa cutoff; Amicon Ultra, USA). The total protein concentration of the extract was estimated using the Quick Start Bradford Assay kit (BioRad, USA) and bovine serum albumin (BSA) as a standard. The filtered and concentrated supernatant of the Anisakis extract was stored in 1-ml aliquots at − 80 °C prior to use.
Cloning of the tropomyosin coding sequence
Total RNA was extracted from A. pegreffii larvae (n = 35) using RNA Mini Kit (Bioline) and cDNA synthesised using a conventional reverse transcription protocol (Bioline) following the manufacturer’s instructions. The coding region of TM was PCR-amplified from this cDNA using the oligonucleotide primer pair A. pegreffii-TM forward (5′-CGGGATCCATAGACGCAATCAAGAAGAAGATG-3′)/A. pegreffii-TM reverse (5′-CGGAGCTCTATTAATATCCAGACAGCTCTTG-3′) under the following conditions: an initial denaturation at 94 °C for 2 min; the subsequent denaturation cycle was at 95 °C for 20 s, annealing at 55 °C for 15 s, and elongation at 72 °C for 50 s for 35 cycles, with a final elongation step at 72 °C for 7 min. The amplicon (855 bp) was purified using the Qiagen PCR Purification Kit following the manufacturer’s instructions and cloned into the sequencing vector, pCR 2.1, using a TOPO TA cloning kit and cloning reaction protocol (Invitrogen, Carlsbad, CA, USA). Subsequently, the coding region for TM was cloned into the expression vector pRSET A into the BamH1 and XhoI sites using T4 DNA ligase (Invitrogen, Carlsbad, CA, USA). Escherichia coli (BL21 strain) was then transformed with this construct by electroporation and incubated in SOC medium (Hanahan, 1983) at 37 °C for 4 h. Transformed E. coli was grown overnight on Luria Bertani (LB) agar with 100 μg/ml of ampicillin at 37 °C. Colonies were screened by PCR using the gene-specific oligonucleotide primers for the presence of inserts. Plasmid DNA was purified from overnight cultures using the QIAprep SPIN reaction kit (Qiagen, Germany) and sequenced to verify its integrity. Cells were stored in glycerol stock at −80 °C.
Recombinant TM was expressed at 37 °C in LB medium (containing 100 μg/ml of ampicillin) and grown until the culture reached middle to late log phase, after which the culture was induced with 0.1 mM IPTG. Recombinant TM was extracted from E. coli using lysis buffer (25 mM Tris–HCl, 300 mM NaCl, 1 mM imidazole, 2 mg/ml of lysozyme, pH 8) and purified employing a nickel-charged metal chelate affinity chromatography and ion exchange chromatography columns (GE Healthcare, USA). Purified protein was then stored in 1-ml aliquots at −80 °C prior to use.
SDS-PAGE analysis
Electrophoresis was performed to analyse the protein repertoire in the parasite extract (Abdel Rahman et al. 2010; Abdel Rahman et al. 2013). The extract (12 μg) was briefly heated in Laemmli buffer with dithiothreitol, and ~ 10 μl of protein was loaded onto a 12% bis-acrylamide gel. The protein components were resolved at 170 V until the tracking dye reached the base. The separated proteins were detected by staining with Coomassie brilliant blue R250 (BioRad, Hercules, CA, USA).
Immunoblotting with monoclonal, polyclonal anti-tropomyosin and anti-crustacean antibody
To confirm the presence of native (n-) and recombinant (r-) TM of A. pegreffii, poly- and mono-clonal antibodies were used. Four micrograms of A. pegreffii protein extract and purified proteins were resolved by SDS-PAGE. The Semi-dry TransBlot Apparatus (BioRad, Hercules, CA, USA) was used to transfer protein onto activated polyvinylidene fluoride (PVDF) membrane. The membranes were then blocked in 5% (w/v) skim milk powder (SMP) in PBS containing 0.05% Tween-20 (PBS-T). The rTM membrane was exposed to (1) monoclonal anti-crustacean TM antibody, MAC 141 (ab50567; Abcam, Cambridge, MA, USA) diluted 1:6000 in 1% SMP in PBS-T and subsequently to rabbit anti-mouse IgG antibody conjugated to HRP (Sigma, St. Louis, MO, USA) diluted 1:8000. (2) An in-house-generated rabbit polyclonal anti-crustacean tropomyosin antibody (Kamath et al., 2014.) diluted 1:10,000 and then to goat anti-rabbit IgG antibody conjugated with HRP (Promega, USA) diluted 1:40,000. The n-TM was exposed to the in-house-generated rabbit polyclonal anti-crustacean tropomyosin antibody as described for rTPM. After the necessary washes (3×) using PBS-T, antibody binding was observed on the membrane using two methods: (a) enhanced chemiluminescent technique, as previously reported (Abdel Rahman et al., 2010, 2011). Briefly, blots were either incubated with chemiluminescent substrate (Sigma, St. Louis, MO, USA) and thereafter exposed to photographic film (GE Healthcare Biosciences, USA.) or (b) membrane was immersed in TMB (3,3′, 5,5′-tetramethylbenzidine) chromogenic HRP-substrate for 5 min, rinsed in distilled water and antibody-binding protein bands detected thereafter. The rTMs of shellfish were used as positive controls (obtained from Molecular Allergy Laboratory of Andreas Lopata).
Immunoblotting with IgE in human patient sera
To determine the allergenicity of rTM of A. pegreffii, IgE antibody reactivity was assessed by immunoblotting using a pool of sera from patients with confirmed shellfish allergy. Serum samples were obtained from patients with a history of allergic reactions to shellfish and/or positive by SPT and/or ImmunoCAP to prawn, crab, or lobster (mean age 32 ± 10.5 years). A serum from a healthy donor was the negative control used. Informed, written consent was obtained from all study subjects, and experimental protocols were approved by The Alfred Hospital Research Ethics Committee (Project number 192/07) and the Monash University Human Ethics Committee (MUHREC CF08/0225).
IgE immunoblotting was conducted as previously described (Abdel Rahman et al. 2013). Briefly, the proteins on the membrane were first incubated with the serum pool (1:10 in 1% SMP, PBS-T) and, thereafter, with rabbit anti-human IgE polyclonal antibody (DAKO Corporation, Carpinteria, CA, USA) and goat anti-rabbit IgG labelled with HRP (Promega, USA). IgE reactivity was then assessed.
Immunofluorescence staining and microscopy
L3s of A. pegreffii were placed in a small plastic cassette and embedded in optimal cutting temperature (OCT) compound medium. The OCT-solidified sample was frozen and cut into serial sections (7–10 μm thick) using a microtome cryostat machine. Sections were melted onto super frost slides and labelled. Drying of sections was by placing slides in blocks and leaving at 4 °C overnight. Sections were thereafter fixed by placing slides into cold acetone (100%) for 10 min and allowed to dry. To dissolve the OCT medium, the slides were put into a staining chamber containing PBS for 10 min. For the immunofluorescent labelling step, 200 μl of rabbit polyclonal anti-TM antibody solution (primary antibody) was added directly to slides to cover the whole sections and incubated at 4 °C overnight, followed by a PBS wash for 10 min. Subsequently, 200 μl of HRP mouse anti-rabbit IgG secondary antibodies conjugated to fluorescein isothiocyanate (FITC) was added directly to the slides and incubated at 24 °C for 30 min, followed by a PBS wash for 10 min. Detection was carried out using 20 μl of DAB substrate solution for 10 min, and slides were washed in PBS for 10 min to remove the excess of this solution. The slides were kept in a chamber in PBS overnight and then dried. Glycerol solution (50%; 5–10 μl) was pipetted onto the section and cover-slipped and allowed to dry. Images were then taken using a fluorescent microscope (Nikon) and studied.
Phylogenetic analyses and amino acid alignment
Representative sequences obtained from Genbank for ITS-1 and ITS-2 respectively (Anisakis simplex C-AY826722.1; A. simplex-EU624342; Anisakis spp.-JN005759.1; Anisakis ziphidarum-AY826725.1; Anisakis brevispiculata-AB592794.1; Anisakis typica-JX848663.1, JX848680.1; Anisakis paggiae-AB592797.1,GU295975.1; Anisakis physeteris-EU718475.1, JX523714.1; Pseudoterranova decipiens-AJ413968.1, AJ413967.1 and Contracaecum rudolphii A-DQ316967.1, DQ316969.1) and the TM sequences obtained from the National Centre for Biotechnology Information (NCBI) for A. simplex, Ascaris lumbricoides, Onchocerca volvulus, Caenorhabditis elegans, Drosophila melanogaster, Penaeus monodon, Habronattus americanus, Periplaneta americana, Hediste diversicolor, Paralithodes camtschaticus, Homo sapiens and Gallus gallus (Uniprot accession IDs, JF902160, FJ655903, L41633.1, CELTMY1B, NM_169633, AY827100, AF034954.1, Y14854.1, AF216518.1, AB270632.1, J00910.1 and M19267.1, respectively) were aligned with the inferred amino acid ITS-1, ITS-2 and TM of A. pegreffii respectively, employing ClustalW alignment Thompson et al. (1994) in MEGA version. 6.0. Phylogenetic analyses of sequence data for the above-named species were performed using the maximum likelihood statistical method. The following default parameters were used for the phylogenetic tree construction: Poisson model, bootstrap (10,000 replicates; random seed) and complete deletion of gaps (Tamura et al. 2013).
In-gel tryptic digestion and LC-MS/MS analysis
Tryptic digestion was performed by excising a band corresponding to 37 kDa in SDS-PAGE protein analysis of A. pegreffii crude extract. The gel band was washed and destained twice in 50% acetonitrile with 50 mM triethylammonium bicarbonate (TEAB) and dehydrated with acetonitrile until the gel plug turned opaque. The gel plug was then rehydrated and incubated in reduction buffer, consisting of 50 mM tris (2-carboxyethyl) phosphine (TCEP) in 50 mM TEAB at 60 °C. Incubation was for 20 min with 20–50 μl of reduction buffer to cover the gel plug. This was followed by alkylation using 100 mM iodoacetamide in 50 mM TEAB at 24 °C in the dark. The sample was washed in 50 mM TEAB twice and dehydrated with acetonitrile. Digestion of the gel plug was with 1 μg of molecular-grade trypsin (Sigma-Aldrich) at 37 °C overnight. Tryptic peptides were extracted from the gel, acidified with formic acid and then analysed. Digested peptides were injected into the column, separated using a gradient of acetonitrile in water and analysed by liquid chromatography/mass spectroscopy (LC-MS/MS), using a Q-Exactive mass spectrometer (Thermo Scientific). The derived peak list was searched using the Mascot search engine (version 2.4; Matrix Science, London, UK) against the UniProt database. The TM from A. pegreffii was identified by peptide sequence identity to TM in this database in two separate experiments.
Results
Verification of the identity of L3s as A. pegreffii
The anisakid larvae from N. richardsoni (flathead fish) used in this study were individually identified as Anisakis larval type 1 (Cannon, 1977; Shamsi et al. 2011), which is consistent with A. pegreffii. The sequencing of ITS-1 and ITS-2 from such larvae supported this identification, with these sequences being 99% identical to two reference sequences (Fig. 1a, b). Phylogenetic analysis showed the concatenated ITS-1 and ITS-2 sequences representing A. pegreffii (those determined herein) grouped together to the exclusion of other members of the Anisakidae family (Fig. 2a, b).


The phylogenetic analysis of a ITS-1 and b ITS-2 sequences of Anisakis pegreffii and some members of the Anisakidae family
A. pegreffii tropomyosin (TM)—characterisation, phylogenetic relationship and localisation
The cDNA encoding TM of A. pegreffii is 855 bp in length and is 99% similar to that of A. simplex. A. pegreffii TM is 284 amino acids in length and has an estimated molecular mass of 33 kDa (Fig. 3a, b). The signature patterns for TM are L-K-E-A-E-x-R-A-E, indicated in red (underlined), conserved in the C-terminal region of A. pegreffii TM, and the highly conserved N-terminal motif DAIKKK, which binds actin (underlined in yellow).

a The cDNA and b the amino acid sequences encoding tropomyosin of Anisakis pegreffii. The signature patterns for TM are L-K-E-A-E-x-R-A-E, indicated in red (underlined), conserved in the C-terminal region of A. pegreffii TM, and the highly conserved N-terminal motif DAIKKK, which binds actin (underlined in yellow)
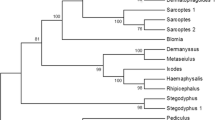
Phylogenetic analysis of TM homologues from A. pegreffii and other organisms, including A. simplex, A. lumbricoides, O. volvulus, C. elegans, D. melanogaster, P. monodon, H. americanus, P. americana, H. diversicolor, P. camtschaticus, humans and chickens, is shown in Fig. 4. Uniprot Accession IDs JF902160, FJ655903, L41633.1, CELTMY1B, NM_169633, AY827100, AF034954.1, Y14854.1, AF216518.1, AB270632.1, J00910.1 and M19267.1, respectively, show that Anisakis TM form a unique clade, clustering closely with A. lumbricoides. Interestingly, the TM representing shellfish is more closely related to the TM of Anisakis spp. than to other nematodes such as C. elegans and O. volvulus, which may explain their cross-reactivity.

Phylogenetic analysis of tropomyosin homologues of Anisakis pegreffii and other organisms, including Anisakis simplex, Ascaris lumbricoides, Onchocerca volvulus, Caenorhabditis elegans, Drosophila melanogaster, Penaeus monodon (giant tiger prawn), Homarus americanus (American lobster), Periplaneta americana (American cockroach), Haliotis diversicolor (coloured abalone), Paralithodes camtschaticus (red king crab), Homo sapiens (human) and Gallus gallus domesticus (chicken)
An independent analysis of the somatic extract of A. pegreffii by LC-MS/MS combined with a database search confirmed the presence of A. pegreffii TM with a match of 14 peptides and sequence coverage of 52% to A. simplex TM Ani s 3 (accession number: Q9NAS5) and a match of two peptides and sequence coverage of 15% to TM of O. volvulus (Ov-tmy-1; accession number: Q25632; Fig. 5 ).

Peptides of tropomyosin protein identified by LC-MS/MS from somatic extracts of A. pegreffii. Yellow, peptides matching Anisakis simplex tropomyosin protein; green, peptides matching Onchocerca volvulus tropomyosin protein
To localise TM in A. pegreffii, L3 larvae were subjected to immunofluorescent staining with an anti-TM polyclonal antibody. This staining revealed preferential binding to and intense labelling of native TM in the cuticle of A. pegreffii L3 larvae (Fig. 6).

Localisation of tropomyosin protein in the cuticle of Anisakis pegreffii L3 larvae by immunofluorescent staining. Treated samples = sections incubated with anti-tropomyosin antibody
SDS-PAGE and immunoblot analyses of A. pegreffii and/or shellfish proteins
The analysis of native extracts from shellfish and A. pegreffii showed a variety of proteins ranging from 10 to 250 kDa (Fig. 7a, b). Whole protein extracts from shellfish and A. pegreffii showed many intense bands significantly in the size between 30 and 50 kDa on a typical SDS-PAGE gel. Distinct protein bands at 37–39 kDa were observed for the shellfish extracts. The rTM protein of A. pegreffii runs slightly slower than the predicted molecular weight (Fig. 7b, c).

a SDS-PAGE gel of the recombinant tropomyosin protein of Anisakis pegreffii. Lane 1 = molecular marker, lane 2 = crude extract of Anisakis pegreffii, lane 3 = recombinant tropomyosin of Anisakis pegreffii. b visualisation of the sizes of recombinant tropomyosin proteins of a crustacean and Anisakis pegreffii using SDS-PAGE. c Immunoblot analysis using anti-tropomyosin polyclonal antibody. Lane 1 = molecular marker, lane 2 = positive control (recombinant tropomyosin of black tiger 37 kDa), lane 3 = fish extract protein (negative control), lane 4 = crude extract of Anisakis pegreffii (41 kDa), lane 5 = recombinant tropomyosin protein of Anisakis pegreffii (44 kDa)
In the immunoblot analysis, a distinct band of 37 kDa was detected in crustacean extract using the monoclonal anti-TM antibody, which represented a positive control (Fig. 8a). However, no band was detected in A. pegreffii extract, by this antibody, even using a prolonged exposure time. Using anti-TM crustacean polyclonal antibody, a distinct band of 37 kDa was observed in the crustacean TM (Fig. 8a) and a single band of the same size was detected in the crude extract of A. pegreffii (lane 6; Fig. 8b). The anti-TM crustacean polyclonal antibody, in addition, also recognised a band of ~ 37 kDa in A. pegreffii extract (lane 6; Fig. 8c). A subsequent immunoblot analysis of rTM of A. pegreffii probed with sera from patients with shellfish allergy revealed IgE antibody binding (Fig. 9). Most of the sera (60%) showed some or faint reactivity.

Immunoblot analysis with (a) Monoclonal (b) Polyclonal anti–tropomyosin antibodies and (c) Polyclonal anti-crustacean antibody against Anisakis pegreffii crude extract. L = Lane; L1 = Molecular marker, L2 = Black Tiger (positive control), L3 = King Prawn (positive control), L4 = Green prawn (positive control), L5 = Banana prawn (positive control), L6 = Anisakis pegreffii, L7 = Phosphate buffered saline (PBS; negative control)

Immunoblot analysis of recombinant tropomyosin of Anisakis pegreffii probed with sera from shellfish-allergic patients. Keys: L = lane, M = molecular marker, L1–10 = patients’ sera (DM32, GC10, AN01, CF 31, JP47, CF31, DG46, KF41 and EV09, respectively), L11 = fish extract, L12 = negative control. This figure shows Anisakis pegreffii recombinant protein tropomyosin allergen at 44 kDa in L1, 2, 5, 6, 7, 8 and 9. The positive bands are shown in the red box
Discussion
Consumption of raw or undercooked seafood contaminated with Anisakis L3s can result in anisakiasis and/or allergic reaction in humans (Daschner et al. 2002). Such allergic responses is characterised by high-level production of IgE antibodies to both somatic and/or excretory/secretory proteins of Anisakis L3s. Diagnostic testing is by detection of specific IgE antibody binding to crude extract of parasite or the recombinant proteins derived from such allergen genes.
TM is the major allergen protein of invertebrates, which has epitopes, featuring high cross-reactivity among invertebrate TMs from other species and which is suspected to elicit hypersensitivity reactions in atopic individuals. Due to its wide distribution, it is considered a pan-allergen (Reese et al. 1999; Acevedo and Caraballo, 2011).
In this study, immunoblotting results revealed that the recombinant A. pegreffii TM protein was not recognised by an anti-TM monoclonal antibody. Thus, we demonstrated that the specific epitope, to which the crustacean anti-TM monoclonal antibody reacts, is distinct in sequence from Anisakis TM, which is consistent with the findings from the phylogenetic analysis (Fig. 4). By contrast, immunoblotting using anti-TM and anti-crustacean polyclonal antibodies showed reactivity with A. pegreffii TM. Polyclonal antibodies usually recognise multiple epitopes of a protein. Hence, overall reactivity, with the use of a polyclonal antibody, may not be greatly affected by small changes in the structure of single or a small number of epitopes (Lipman et al. 2005). In addition, to confirm that the rTM of A. pegreffii retained the allergenic characteristics of natural A. pegreffii TM, IgE in sera from shellfish allergic patients reacted with rTM of A. pegreffii. This result agrees strongly with other studies that have reported sera reaction from allergic patients with A. simplex TM (Acevedo and Caraballo, 2011). Although TM of A. pegreffii did not react with all patient sera, this result validates cross-reactivity between TM of A. pegreffii and shellfish allergic patient sera. The close relatedness of some of these species can be observed in the phylogenetic tree (Fig. 4). Phylogenetic analysis showed that TM of A. pegreffii has closer relationship to TM from other nematodes and crustaceans but revealed less similarity to TM from vertebrates (chickens and humans). These results are in agreement with the proposal that TM is a pan-allergen.
We localised TM in A. pegreffii for the first time using an immunohistochemistrical approach. TM was detected in the muscles and under the basal layer of the cuticle of A. pegreffii L3 larvae using anti-TM polyclonal antibodies. This finding agrees with several studies that have reported the detection of TM in the muscles of parasites (Jenkins et al. 1998; Macgregor and Shore 1990). rTM protein was observed to be larger than the native one and migrated slower on SDS-PAGE than its predicted mass of 37 kDa. The slower run is a consequence of the higher mass of the recombinant protein due to the N-terminal extension encoded by the expression vector.
Sequence similarity searches showed that A. pegreffii TM has the highest identity to the alpha-TM isoform from L3s of A. simplex (98%) and A. lumbricoides (84%), respectively. The deduced amino acid sequence of A. pegreffii L3 TM has revealed that the TM signature, L-K-E-A-E-x-R-A-E, is conserved in the C-terminal region at position 231–239, as is the highly conserved N-terminal motif D-A-I-K-K-K, which is significant for actin binding. Findings from this study agree with several studies reporting similar results in other organisms (Asturias et al. 2000; Santos et al. 1999; Santos et al. 2008). Furthermore, a potential N-linked glycosylation site N-R-S in TM of A. pegreffii was identified from the data obtained from this study.
The protein sequence of TM from A. pegreffii L3 showed a sequence of seven heptad-repeating amino acids residues usually labelled abcdefg. Results similar to this, were obtained by Asturias et al. (2000) who investigated the stage 3 larvae of A. simplex rTM and showed that in the sequence ‘a’ and ‘d’ amino acids, are the hydrophobic positions of each chain. Interactions of these amino acids with each other result in the parallel α-helical coiled coil chain for TM (Mason and Arndt, 2004). This report presents evidence of high indications of α-helical coiled-coil structures of A. pegreffi’s rTM. This study confirms cross-reactivity between A. pegreffii and shellfish TM. It also demonstrates that the specific epitope to which the crustacean anti-TM monoclonal antibody reacts is not conserved in Anisakis; however, the nematode TM tested share one or more epitopes with crustacean TM, as polyclonal antibodies can recognise TM from both species.
In conclusion, we have confirmed the presence of TM protein in A. pegreffi with anti-TM crustacean polyclonal antibodies. In contrast, A. pegreffii TM was not recognised by immunoblotting with an anti- crustacean TM monoclonal antibody (MAb). This information suggests that A. pegreffi TM share one or more epitopes with crustacean TM, as the PAbs is able to recognise multiple epitopes on any one antigen, while they lack specific antigenic determinant, defined by the anti-TM monoclonal antibody present in crustacean TM. The result of this study showed that the size of A. pegreffii -TM is similar with previously studied A. simplex and A. lumbricoides TM, as may be expected by the close relationship between these species. Furthermore, results obtained from this study indicate that the rTM of A. pegreffii binds to IgE from different sera samples of crustacean allergic patients confirming cross reactivity between nematodes and different classes of shellfish. Previous studies had shown the function of TM protein to be associated with motility. In the current study, we substantiated the evidence of the association of TM proteins with motility role by determining its localisation in A. pegreffii by immunohistochemistry. This showed A. pegreffii -TM to be located in both the muscles and under the basal layer of the cuticle in the A. pegreffii L3 anatomy. Future studies should explore the cross-reactivity of the allergenic protein repertoire of Anisakis in more detail in order to work toward improving diagnosis of allergenic diseases in humans.
References
Abdel Rahman AMA, Kamath S, Lopata AL, Helleur RJ (2010) Analysis of the allergenic proteins in black tiger prawn (Penaeus monodon) and characterization of the major allergen tropomyosin using mass spectrometry. Rapid Commun Mass Spectrom 24:2462–2470
Abdel Rahman AM, Kamath S, Lopata AL, Helleur RJ (2011) Biomolecular characterization of allergenic proteins in snow crab (Chionoecetes opilio) and de novo sequencing of the second allergen arginine kinase using tandem mass spectrometry. Proteomics 74:231–241
Abdel Rahman AM, Kamath SD, Gagne S, Lopata AL, Helleur R (2013) Comprehensive proteomics approach in characterizing and quantifying allergenic proteins from northern shrimp: toward better occupational asthma prevention. J Proteome Res 12:647–656
Abollo E, Paggi L, Pascual S, D'Amelio S (2003) Occurrence of recombinant genotypes of Anisakis simplex ss and Anisakis pegreffii (Nematoda: Anisakidae) in an area of sympatry. Infect Genet Evol 3:175–181
Acevedo N, Caraballo L (2011) IgE cross-reactivity between Ascaris lumbricoides and mite allergens: possible influences on allergic sensitization and asthma. Parasite Immunology 33:309–321
Albrecht M, Alessandri S, Conti A, Reuter A, Lauer I, Vieths S, Reese G (2008) High-level expression purification and physico- and immunochemical characterisation of recombinant Pen a 1: a major allergen of shrimp. Mol Nutr food res 52(Suppl 2):186–195
Anibarro B, Seoane FJ (1998) Occupational conjunctivitis caused by sensitization to Anisakis simplex. J Allergy Clin Immunol 102:331–332
Asturias JA, Eraso E, Martinez A (2000) Cloning and high level expression in Escherichia coli of an Anisakis simplex tropomyosin isoform. Mol Biochem Parasitol 108:263–266
Bao M, Pierce GJ, Pascual S, Gonzalez-Munoz M, Mattiucci S, Mladineo I, Cipriani P, Buselic I, Strachan NJ (2017) Assessing the risk of an emerging zoonosis of worldwide concern: anisakiasis. Sci Rep 7:43699
Campana-Rouget Y, Biocca E (1955) A new species of Anisakis in a Mediterranean seal. Ann Parasitol Hum Comp 30:477–480
Cannon LRG (1977) Some ecological relationships of larval ascaridoids from south-eastern Queensland marine fishes. Int J Parasitol 7:227–232
Daschner A, Cuéllar C, Sánchez-Pastor S, Pascual CY, Martín-Esteban M (2002) Gastro-allergic anisakiasis as a consequence of simultaneous primary and secondary immune response. Parasite Immunol 24:243–251
Faeste CK, Wiker HG, Løvik M, Egaas E (2003) Hidden shellfish allergen in a fish cake. Allergy 58:1204–1205
Fumarola L, Monno R, Ierardi E, Rizzo G, Giannelli G, Lalle M, Pozio E (2009) Anisakis pegreffi etiological agent of gastric infections in two Italian women. Foodborne Pathog Dis 6:1157–1159
Gasser RB, Hu M, Chilton NB, Campbell BE, Jex AJ, Otranto D, Cafarchia C, Beveridge I, Zhu XQ (2006) Single-strand conformation polymorphism (SSCP) for the analysis of genetic variation. Nat Protoc 1:3121–3128
Gregori M, Roura Á, Abollo E, González AF, Pascual S (2015) Anisakis simplex complex (Nematoda: Anisakidae) in zooplankton communities from temperate NE Atlantic waters. J Nat Hist 49(13–14):755–773. https://doi.org/10.1080/00222933.2014.979260
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Hanahan D (1983) Studies on transformation of Escherichia coli with plasmids. Mol Biol 166:557–580
Jabbar A, Khoon AT, Hui TX, Schaeffner BC, Jex AR, Nolan MJ, Lopata AL, Gasser RB, Beveridge I (2012) Mutation scanning-based analysis of anisakid larvae from Sillago flindersi from Bass Strait Australia. Electrophoresis 33:499–505
Jenkins RE, Taylor MJ, Gilvary NJ, Bianco AE (1998) Tropomyosin implicated in host protective responses to microfilariae in onchocerciasis. Proc Natl Acad Sci USA 95:7550–7555
Kamath SD, Abdel Rahman AM, Komoda T, Lopata AL (2013) Impact of heat processing on the detection of the major shellfish allergen tropomyosin in crustaceans and molluscs using monoclonal antibodies. Food Chem 141(4):4031–4039
Kamath SD, Thomassen MR, Saptarshi SR, Nguyen HM, Aasmoe L, Bang BE, Lopata AL (2014) Molecular and immunological approaches in quantifying the air-borne food allergen tropomyosin in crab processing facilities. Int J Hyg Environ Health 217(7):740–750
Kim JS, Kim KH, Cho S, Park HY, Cho SW, Kim YT, Joo KH, Lee JS (2005) Immunochemical and biological analysis of allergenicity with excretory-secretory products of Anisakis simplex third stage larva. Int Arch Allergy Immunol 136(4):320–328
Kirstein F, Horsnell WG, Nieuwenhuizen N, Ryffel B, Lopata AL, Brombacher F (2010) Anisakis pegreffii-induced airway hyperresponsiveness is mediated by gamma interferon in the absence of interleukin-4 receptor alpha responsiveness. Infect Immun 78:4077–4086
Lipman NS, Jackson LR, Trudel LJ, Weis-Garcia F (2005) Monoclonal versus polyclonal antibodies: distinguishing characteristics applications and information resources. J ILAR 46:258–268
Lopata AL, Lehrer SB (2009) New insights into seafood allergy. Curr Opin Allergy Clin Immunol 9:270–277
Lopata AL, O'Hehir RE, Lehrer SB (2010) Shellfish allergy. Clin Exp Allergy 40:850–858
Macgregor AN, Shore SJ (1990) Immunocytochemistry of cytoskeletal proteins in adult Schistosoma mansoni. Int J Parasitol 20:279–284
Mason JM, Arndt KM (2004) Coiled coil domains: stability specificity and biological implications. Chembiochem 5:170–176
Mattiucci S, Nascetti G (2006) Molecular systematics phylogeny and ecology of anisakid nematodes of the genus Anisakis Dujardin 1845: an update. Parasite 13:99–113
Mattiucci S, Nascetti G (2007) Genetic diversity and infection levels of anisakid nematodes parasitic in fish and marine mammals from Boreal and Austral hemispheres. Vet Parasitol 148:43–57
Mattiucci S, Nascetti G (2008) Advances and trends in the molecular systematics of anisakid nematodes with implications for their evolutionary ecology and host-parasite co-evolutionary processes. Adv Parasitol 66:47–148
Mattiucci S, Nascetti G, Cianchi R, Paggi L, Arduino P, Margolis L, Brattey J, Webb S, Damelio S, Orecchia P, Bullini L (1997) Genetic and ecological data on the Anisakis simplex complex with evidence for a new species (Nematoda Ascaridoidea Anisakidae). J Parasitol 83:401–416
Mattiucci S, Paggi L, Nascetti G, Portes Santos C, Costa G, Di Beneditto AP, Ramos R, Argyrou M, Cianchi R, Bullini L (2002) Genetic markers in the study of Anisakis typica (Diesing 1860): larval identification and genetic relationships with other species of Anisakis Dujardin 1845 (Nematoda: Anisakidae). Syst Parasitol 51:159–170
Mattiucci S, Paoletti M, Webb SC (2009) Anisakis nascettii n sp (Nematoda: Anisakidae) from beaked whales of the southern hemisphere: morphological description genetic relationships between congeners and ecological data. Syst Parasitol 74:199–217
Mattiucci S, Fazii P, De Rosa A, Paoletti M, Megna AS, Glielmo A, De Angelis M, Costa A, Meucci C, Calvaruso V, Sorrentini I, Palma G, Bruschi F, Nascetti G (2013) Anisakiasis and gastroallergic reactions associated with Anisakis pegreffii infection. Italy Emerg Infect Dis 19:496–499
Nascetti G, Paggi L, Orecchia P, Smith JW, Mattiucci S, Bullini L (1986) Electrophoretic studies on the Anisakis simplex complex (Ascaridida:Anisakidae) from the Mediterranean and North-East Atlantic. Int J Parasitol 16:633–640
Nieuwenhuizen NE, Lopata AL (2013) Anisakis—a food-borne parasite that triggers allergic host defences. Int J Parasitol 43:1047–1057
O’Donnell IJ, Dineen JK, Wagland BM, Letho S, Dopheide TAA, Grant WN, Ward CW (1989) Characterization of the major immunogen in the excretory secretory products of exsheathed 3rd-stage larvae of Trichostrongylus-colubriformis. Int J Parasitol 19:793–802
Reese G, Ayuso R, Lehrer SB (1999) Tropomyosin: an invertebrate pan-allergen. Int Arch Allergy Immunol 119:247–258
Santos AB, Chapman MD, Aalberse RC, Vailes LD, Ferriani VP, Oliver C, Rizzo MC, Naspitz CK, Arruda LK (1999) Cockroach allergens and asthma in Brazil: identification of tropomyosin as a major allergen with potential cross-reactivity with mite and shrimp allergens. J allergy Clin Immunol 104(2 Pt 1):329–337
Santos AB, Rocha GM, Oliver C, Ferriani VP, Lima RC, Palma MS, Sales VS, Aalberse RC, Chapman MD, Arruda LK (2008) cross-reactive IgE antibody responses to tropomyosins from Ascaris lumbricoides and cockroach. J Allergy Clin Immunol 121:1040–1046
Shamsi S, Gasser R, Beveridge I, Shabani AA (2008) Contracaecum pyripapillatum n sp (Nematoda: Anisakidae) and a description of C multipapillatum (von Drasche 1882) from the Australian pelican Pelecanus conspicillatus. Parasitol Res 103:1031–1039
Shamsi S, Eisenbarth A, Saptarshi S, Beveridge I, Gasser RB, Lopata AL (2011) Occurrence and abundance of anisakid nematode larvae in five species of fish from southern Australian waters. Parasitol Res 108:927–934
Tamura K, Stecher G, Peterson D, Filipski A, Kumar S (2013) Mega 6: molecular evolutionary genetics analysis version 6.0 Mol Biol Evol 30:2725–2729
Thompson JD, Higgins DG, Gibson TJ (1994) ClustalW: improving the sensitivity of progressive multiple sequence alignment through sequence weighting position specific gap penalties and weight matrix choice. Nucleic Acids Res 22:4673–4680
Umehara A, Kawakami Y, Araki J, Uchida A (2007) Molecular identification of the etiological agent of the human anisakiasis in Japan. Parasitol Int 56:211–215
Umehara A, Kawakami Y, Araki J, Uchida A (2008) Multiplex PCR for the identification of Anisakis simplex sensu stricto Anisakis pegreffii and the other anisakid nematodes. Parasitol Int 57:49–53
Werner MT, Fæste CK, Levsen A, Egaas E (2011) A quantitative sandwich ELISA for the detection of Anisakis simplex protein in seafood. Eur Food Res Technol 232:157
Zhang L, Hu M, Shamsi S, Beveridge I, Li H, Xu Z, Li L, Cantacessi C, Gasser RB (2007) The specific identification of anisakid larvae from fishes from the Yellow Sea China using mutation scanning-coupled sequence analysis of nuclear ribosomal DNA. Mol Cell Probes 21:386–390
Acknowledgements
We thank Dr. Sandip Kamath for assisting with the immunoblots and Prof Robyn O’Hehir for donating serum samples.
Funding
Andreas Lopata received financial support via an ARC Future Fellowship for this work. Robin Gasser received support from the Australian Research Council (ARC), the National Health and Medical Research Council (NHMRC) of Australia, Yourgene Bioscience (Taiwan), Melbourne Water Corporation and The University of Melbourne.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Informed, written consent was obtained from all study subjects, and experimental protocols were approved by The Alfred Hospital Research Ethics Committee (Project number 192/07) and the Monash University Human Ethics Committee (MUHREC CF08/0225).
Rights and permissions
About this article

Cite this article
Asnoussi, A., Aibinu, I.E., Gasser, R.B. et al. Molecular and immunological characterisation of tropomyosin from Anisakis pegreffii . Parasitol Res 116, 3291–3301 (2017). https://doi.org/10.1007/s00436-017-5642-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-017-5642-4