
Abstract
Host specialisation in parasites can be due to either limited exposure or limited adaptation to different host types. When the first barrier is lifted experimentally, the degree of adaptive specialisation can be studied. The tree-hole tick Ixodes arboricola is an endophilic parasite with a narrow host range, found in nest boxes used mainly by great and blue tits (Parus major, Cyanistes caeruleus) and to a lesser extent by pied flycatchers (Ficedula hypoleuca) and nuthatches (Sitta europaea). In the current study, we exposed two nestlings per nestbox of pied flycatchers (N = 14), blue tits (N = 18), great tits (N = 14), and nuthatches (N = 16) to ten I. arboricola nymphs each. We found no differences in attachment success 2 days after infestation (56 ± 4 % across species) nor were there any differences in tick engorgement weight (1.95 ± 0.03 mg across species), and moulting success was >90 % for ticks from all bird species. Hence, our data suggest that all bird species investigated here are suitable host species. This may enhance the ticks’ chances for persistence in cavities and dispersal among cavities inhabited by multiple host species, and supports the hypothesis that host use by ticks is limited by host ecology rather than by host specialisation.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Host specificity is a common feature of many parasite taxa as selection tends to favour specialisation of parasites to their local environment (Klompen et al. 1996; Giorgi et al. 2004; Poulin 2007). Host specificity may be the result of adaptive constraints because adaptations that increase performance on one host may hinder survival or fecundity on others (Joshi and Thompson 1995; Kassen 2002; Dietrich et al. 2013). Alternatively, observed patterns of host specificity may simply occur because parasites do not come into contact with other host species (Timms and Read 1999). For phytophagous insects, numerous studies have investigated if observed patterns of host use are attributable to variation in host competence or to limited parasite dispersal capabilities (Drès and Mallet 2002). For ectoparasites, however, such investigations are mostly lacking (but see, e.g. Tripet and Richner 1997; Sears et al. 2012; Gómez-Díaz et al. 2012; Dietrich et al. 2013).
Host use in ectoparasites is a complex of many traits, of which many are involved in the exploitation of the host (Poulin 2007). The performance of traits allowing parasites to complete a blood meal depends on the suitability of a host. First, parasites need to attach to a host and penetrate its skin, and attachment success is lower on unsuitable hosts (Dietrich et al. 2013). Second, once the skin is penetrated, parasites need to evade host defences, and it has been shown that blood meal duration is longer (Mccoy and Boulinier 2002; Labruna et al. 2009), engorgement success lower (Labruna et al. 2002; Labruna et al. 2009; Dietrich et al. 2013) and the blood meal smaller on unsuitable hosts (Olegário et al. 2011). Finally, moulting (Labruna et al. 2002; Mccoy and Boulinier 2002; Labruna et al. 2009) and hatching success (Olegário et al. 2011) are lower and moulting time is longer on unsuitable hosts (Labruna et al. 2009; Olegário et al. 2011). Thus, a plethora of traits is available to study the degree of host suitability of ectoparasites.
The family of hard ticks (Ixodidae) was formerly considered to consist mainly of host specialists, i.e. parasites infesting one or only a limited number of host species (Hoogstraal and Aeschlimann 1982). Ixodid ticks take a blood meal lasting several days, and during this period, they need to maintain blood flow and evade immunological and behavioural responses of the host (Sonenshine 1991; McCoy et al. 2013). The evasion of host defences can be a complex process and should select for host specificity (Magalhães et al. 2007). Even generalist ticks appear to be genetically differentiated among different types of hosts, as has been shown for Ixodes ricinus, a tick infesting mammals, birds and reptiles (Kempf et al. 2011), as well as Ixodes uriae, a tick infesting a wide range of seabird species (McCoy et al. 2001). However, it has been shown that host specificity of many ixodid ticks may have been overestimated because of incomplete sampling (Klompen et al. 1996; Nava and Guglielmone 2013). These studies also suggested that ticks are not limited by host use but rather by biogeography: abiotic conditions during the long off-host period of the life cycle and host ecological similarities define host use (Klompen et al. 1996; Nava and Guglielmone 2013). In addition, most studies consider host specificity from a quantitative perspective (i.e. number of host species used), whereas a qualitative framework (i.e. differential performance on different hosts), which may provide a more realistic picture of natural interactions, is largely lacking (Poulin 2007; McCoy et al. 2013).
Ixodes arboricola Schulze and Schlottke 1930 is an endophilic tick, i.e. it remains hidden inside its hosts’ nest and attaches when the host arrives (Salman and Tarrés-Call 2012). Endophilic ticks typically have a smaller host range than exophilic ticks, which are not associated with nests and encounter many different host species (Hoogstraal and Aeschlimann 1982; Salman and Tarrés-Call 2012). Indeed, I. arboricola chiefly infests cavity-nesting birds, with great and blue tits (Parus major, Cyanistes caeruleus) as its principal host (Walter et al. 1979; Hudde and Walter 1988; Heylen et al. 2014). It is unknown to what extent feeding performance differs among different cavity-nesting birds, thus to what extent I. arboricola can be considered a specialist or generalist parasite. This knowledge may provide more insight in the ecology of bird-tick interactions and more generally in the evolution of host specialisation (Poulin 2007; McCoy et al. 2013). In addition, I. arboricola is a carrier of the pathogens Rickettsia sp. and Borrelia burgdorferi s.l. (Špitalská et al. 2011; Heylen et al. 2013), and although I. arboricola does not feed on humans and livestock, it may share hosts with the generalist tick I. ricinus, which is the main European vector of tick-borne diseases (Hillyard 1996; Heylen et al. 2013). The degree of host specificity of I. arboricola will dictate tick dispersal and encounter rates with I. ricinus and may therefore increase pathogen circulation.
Here, we study the host specificity of I. arboricola by experimentally evaluating the tick’s infestation performance on nestlings of four cavity-nesting bird species that are known hosts of I. arboricola and use very similar nest sites: great and blue tits, nuthatches (Sitta europaea) and pied flycatchers (Ficedula hypoleuca). It was predicted that I. arboricola is able to infest nestlings of all four species of birds (Hudde and Walter 1988). However, we expected tick feeding performance (e.g. attachment to the host, survival and moulting time) to differ among species. Specifically, because parasites are frequently more infective on the most common host (Legros and Koella 2010), feeding performance was expected to be higher on the common great and blue tits than on the less abundant pied flycatchers and nuthatches.
Materials and methods
Study system
I. arboricola is widely distributed in the Palearctic Region, from central, western and northern Europe, eastward to Latvia and the European parts of Russia (Hudde and Walter 1988; Liebisch 1996). Apart from great and blue tits, the most frequently infested hosts of I. arboricola are nuthatches, treecreepers (Certhia brachydactyla) and pied flycatchers (Walter et al. 1979). Anecdotal reports are available from other hosts, e.g. Eurasian eagle-owl (Bubo bubo), marsh tit (Parus palustris), great spotted woodpecker (Dendrocopos major), common starling (Sturnus vulgaris) and Eurasian tree sparrow (Passer montanus) (Hudde and Walter 1988). Because I. arboricola infests cavity-nesting birds, its entire life cycle is restricted to natural and man-made cavities (Walter et al. 1979; Heylen et al. 2014), where it detaches after feeding (Heylen and Matthysen 2010; White et al. 2012; Heylen et al. 2014). There, immature ticks (larvae and nymphs) moult to the next developmental stage (nymph or adult, respectively), and adult female ticks lay eggs after their meal and die. With the exception of adult male ticks, which do not feed, every instar (larva, nymph, adult female) takes a single blood meal (Sonenshine 1991).
Great and blue tits (family Paridae) are the most common cavity-nesting birds in Western Europe and occur nearly everywhere in Flanders, Belgium (Gosler 1993; Vermeersch et al. 2004). In suitable deciduous forests, population densities may reach three to four pairs per hectare, particularly if nestboxes are provided (Gosler 1993; Dhondt 2010). Great and blue tits use nest boxes and natural cavities in the breeding season (April to June) to raise young and in winter (late September to early March) for roosting (Mainwaring 2011). When nest boxes are abundant, these are strongly preferred for breeding (Gosler 1993). Their nests consist of a foundation of moss, topped off with hair and, in the case of blue tits, feathers (Gosler 1993). They lay 4 to 12 eggs (blue tits up to 14) which require 15 to 20 days of incubation, and nestlings fledge at a length of 13–15 cm (great tits) or 11–12 cm (blue tits) and a weight of 14–22 g (great tits) or 7.5–14 g (blue tits) after 18 to 21 days (Gosler 1993; Fargallo and Johnston 1997). The natural prevalence of ticks infesting adult great and blue tits has been estimated at 7.5 % in the pre-breeding season (early March–early April) but much lower throughout the rest of the year (Heylen et al. 2014).
Nuthatches (family Sittidae) also are resident cavity-nesting birds in most of Europe. Densities are typically much lower than those of great and blue tits with maxima up to 0.8 pairs per hectare but more typically around 0.2 to 0.5 pairs per hectare (Matthysen 1998; Vermeersch et al. 2004). Like tits, nuthatches use nest boxes and natural cavities in the breeding season (April to June) to raise young and in winter (late September to early March) for roosting (Mainwaring 2011). Nuthatches build nests with a foundation of rotten wood and bark and a top layer of loose lining material, typically bark flakes of Scots pine or other trees (Matthysen 1998). They lay 5 to 12 eggs which require 13 to 18 days of incubation. Nuthatch nestlings develop relatively slowly compared with similar-sized passerines and fledge at a length of 14 cm and weight of 17–28 g after 18 to 25 days (Matthysen 1998). The natural prevalence of ticks infesting adult nuthatches has been estimated at <5 % year-round (unpublished data).
Pied flycatchers (family Muscicapidae) are migratory birds that only come to Europe to breed and spend the remainder of the year at wintering grounds in west Africa (Lundberg and Alatalo 1992). They are less widespread than great tits and nuthatches, and within Flanders they are largely restricted to forests on sandy soils in the northeast (Vermeersch et al. 2004). Their distribution is more patchy than tits and nuthatches because pied flycatchers defend rather small territories, and local densities may be up to ten pairs per hectare (Vermeersch et al. 2004). Nest boxes and natural cavities are used only in the breeding season (April to June). Pied flycatcher nests consist of two layers: an outer layer usually of bark, twigs and dead leaves and an inner layer of dry grass, animal hair, root fibres and shafts of moss (Lundberg and Alatalo 1992). Pied flycatchers lay five to seven eggs that hatch after 13 to 15 days of incubation, and fledging occurs at a length of 13 cm and weight of 9.7–16.5 g after approximately 15 days (Lundberg and Alatalo 1992). No data concerning the natural prevalence of I. arboricola are available, but pied flycatchers are known hosts of I. arboricola (Walter et al. 1979; Hudde and Walter 1988; Liebisch 1996), and in our study population, we have found I. arboricola ticks in nest boxes that were used by pied flycatchers (unpublished data).
Experimental protocol
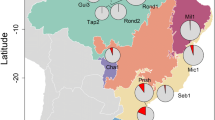
The study took place in the breeding season of 2013 in four experimental plots in northern Flanders, Belgium: Sterbos, Wuustwezel (SB); Wildertse Duintjes, Wildert (WD); Peerdsbos, Brasschaat (PB) and Zevenbergen, Lier (ZB) (Fig. 1). These plots are close together (pairwise distance <35 km) and consist of similar habitat, climate and species composition. Among the plots, there were two types of nest boxes: in SB and WD, nest boxes have a thin metal roof that slides in between the side walls and clamps that are screwed to the side walls (“slide” type). In PB and ZB, nest boxes have a wooden lid that rests on top of the walls and can be removed by pivoting hinges attached to the side (“pivot” type). Different plots had to be used to acquire a sufficient number of nuthatch nests because nuthatches are relatively rare. In order to minimise habitat differences between nests of the different species, we tried as much as possible to select all nests of great and blue tits in the vicinity of nuthatch nests. This was not always possible for pied flycatchers because these birds tend to breed in concentrated breeding groups (Lundberg and Alatalo 1992; Vermeersch et al. 2004). In total, we used seven nests of pied flycatchers (one in PB, six in SB), nine nests of blue tits (one in PB, five in SB, two in WD and one in ZB), seven nests of great tits (one in PB, four in SB and two in WD) and eight nests of nuthatches (one in PB, three in SB, two in WD and two in ZB).

Location of the experimental plots. Plots are abbreviated as follows: Sterbos, Wuustwezel (SB); Wildertse Duintjes, Wildert (WD); Peerdsbos, Brasschaat (PB) and Zevenbergen, Lier (ZB). The agglomeration of Antwerp is marked in grey
In every nest, we selected four nestlings with weights closest to the median weight of the brood. Two of these nestlings were infested with ticks by placing each nestling individually in a cotton bag (20 × 30 cm) that contained ten I. arboricola nymphs (experimental treatment; N = 620 ticks; N = 14 pied flycatchers, 18 blue tits, 16 great tits and 16 nuthatches). The other two nestlings were each placed in a separate bag without ticks (control treatment; N = 14 pied flycatchers, 18 blue tits, 14 great tits and 15 nuthatches). The bags were placed in the nest box for 1 h, after which the nestlings were placed back into the nest and the bags were inspected for unattached ticks. The nestlings were not inspected for ticks at this moment since unfed ticks are difficult to detect without prolonged manipulation which may disturb the attachment process (Heylen and Matthysen 2011). Nestlings were infested with ticks when the first flight feathers appeared. For great and blue tits, this is the eighth or ninth day after hatching, when the secondaries and secondary coverts develop (Gosler 1993); for nuthatches, this is the 11th day after hatching, when the primaries and primary coverts appear (Matthysen 1998); for pied flycatchers, this is the eighth day after hatching, when the primaries and secondaries appear (Lundberg and Alatalo 1992).
At the day of infestation, weight and tarsus length were recorded and nestlings received a metal ring for identification. Weight and tarsus length were recorded again when the nestlings were fully developed (14 days for pied flycatchers, 15 days for great and blue tits and 19 days for nuthatches). All I. arboricola nymphs that were used in the current study fed on adult great tits as larvae in early 2013 and were progeny of adult ticks that fed on great tit nestlings in the breeding season of 2012. The adult ticks came from a laboratory stock that has been established in 2007 with ticks from nest boxes used by great and blue tits in woodland areas near Antwerp (mainly PB) and kept at outside temperature and 85 % relative humidity in the dark. The laboratory stock has been maintained by allowing ticks to infest great tits (and only very few individuals on blue tits in the start-up phase of the laboratory stock) in several studies (Heylen and Matthysen 2011).
Tick preference and success
Nestlings were inspected for ticks 2 days after infestation and again when the nestlings were fully developed by holding the nestling firmly and brushing the feathers apart with tweezers. Ticks were counted but not removed. No ticks were found at the second inspections.
The upper surfaces of the nest boxes were checked regularly from the day of the first nestling inspection until a week after the nestlings had fledged for detached, engorged ticks, which usually crawl to the top of the nest box (Heylen and Matthysen 2011). Ticks were collected and weighed to the nearest 0.1 mg in clusters of one to five individuals (average, 2.24 ± 0.16 individuals) to minimise measurement error in the low regions of the balance. After weighing, ticks were rinsed to prevent fungal infections by placing them in a small tube with distilled water and shaking gently for a few seconds. Subsequently, they were kept in the dark at 25 °C and 85 % relative humidity until they moulted. Moulting condition was monitored every 3 days for a period of 30 days.
Statistical analyses
All data analyses were done in R v 3.0.1 (R Core Team 2013). We used linear mixed-effects models from package LME4 Version 1.0 (Bates et al. 2013) to evaluate whether attachment success at day 0 (i.e. the proportion of nymphs attached to the nestlings 1 h after infestation), attachment success at day 2 (i.e. the proportion of nymphs attached to the nestlings 2 days after infestation relative to the proportion of initially attached nymphs), nymph feeding success (i.e. the proportion of nymphs collected from a nest box relative to the proportion of initially attached nymphs), engorgement weight of the nymphs and moulting success were significantly different across bird species, and the effect of tick infestation intensity (i.e. the number of ticks counted 2 days after infestation) with changes in nestling body weight between the moment of infestation and full development and, in a second model, the association of infestation intensity with changes in nestling tarsus length between the moment of infestation and full development.
Shapiro-Wilk tests were used to evaluate whether all data were normally distributed and transformations were carried out where necessary. In all models, bird species was used as a fixed factor. In the models for attachment success at day 0 and day 2, engorgement weight, moulting success and changes in body weight and tarsus length but not in the model for feeding success, we used bird nest as a random effect. In the model for feeding success, we used the type of nest box (“slide” or “pivot”) as an additional fixed factor and tested the interaction with host species. In the models for changes in body weight and tarsus length, we used nestlings from both the experimental and control treatment, and in the other models we used only nestlings from the experimental treatment. Due to the limited number of nests per plot, we did not use the experimental plot as a factor. For engorgement weight, the average weight of each cluster of nymphs was considered as the weight of individual nymphs from that cluster. Moulting time of nymphs was analysed with a frailty survival model from package parfm Version 2.5.3 (Rotolo and Munda 2013) with bird nest as a random effect. All surviving nymphs moulted within 30 days. Individuals that died during this period (5.3 %) were handled as right-censored data. Differences among bird species were tested in pairwise comparisons corrected with Benjamini-Hochberg procedure for false discovery rate.
Results
All data were normally distributed; hence, no transformations were performed. Attachment success at day 0 was significantly different among bird species (F (3, 58) = 3.45, p = 0.03; Fig. 2a, Table 1). Nuthatches were accepted as hosts by 0.78 ± 0.04 of I. arboricola nymphs, which is significantly higher than attractiveness of blue tits (estimated difference, 0.22 ± 0.08; p = 0.01) and great tits (estimated difference, 0.21 ± 0.08; p = 0.02). Attachment success at day 2 was 0.56 ± 0.04 across all bird species. No significant differences in attachment success at day 2 were observed among bird species (F (3, 58) = 2.55, p = 0.07; Fig. 2b, Table 1).

Proportion (±SE) of I. arboricola nymphs that were attached to nestlings of pied flycatchers (PF), blue tits (BT), great tits (GT) and nuthatches (NH) on day 0 (a) and day 2 (b). Letter codes (a, b) refer to species that do not differ significantly
The proportion of nymphs collected from nestboxes differed significantly among bird species (F (3, 27) = 3.45, p = 0.03; Fig. 3, Table 1). Specifically, significantly more nymphs were collected from blue tit nests (0.47 ± 0.06) than from pied flycatcher nests (estimated difference, 0.33 ± 0.10; p = 0.02). There were no differences in the proportion of collected nymphs between different types of nest boxes (F (1, 27) = 1.65, p = 0.21) nor was there an interaction between host species and nest box type (F (3, 27) = 2.03, p = 0.14).

Proportion (±SE) of I. arboricola nymphs attached at day 0 collected from nest boxes of pied flycatchers (PF), blue tits (BT), great tits (GT) and nuthatches (NH). Letter codes (a, b) refer to species that do not differ significantly
The average engorgement weight of nymphs was 1.95 ± 0.03 mg across all bird species. No significant differences were found among bird species (F (3, 54) = 2.24, p = 0.12; Fig. 4, Table 1). Moulting success was 0.95 ± 0.02 across all bird species, and there were no significant differences among bird species (F (3, 129) = 2.68, p = 0.61; Table 1).

Engorgement weight (mg ± SE) of I. arboricola nymphs collected from nest boxes of pied flycatchers (PF), blue tits (BT), great tits (GT) and nuthatches (NH)
All nymphs that survived moulted within 30 days after the blood meal was initiated. Moulting time was significantly different among bird species (χ 2 (1, N = 126) = 10.51, p < 0.01; Fig. 5, Table 1). Specifically, moulting time of nymphs that fed on nuthatches (26.79 ± 0.79 days) was longer than that of nymphs that fed on pied flycatchers (23.79 ± 0.61 days; p = 0.04) and great tits (23.81 ± 0.48 days; p = 0.04).

Kaplan-Meier survival plot of moulting time of I. arboricola nymphs collected from pied flycatchers (PF), blue tits (BT), great tits (GT) and nuthatches (NH). Nymphs that fed on nuthatches took longer to moult than those that fed on pied flycatchers and great tit
There were no significant interactions between bird species and tick infestation intensity in the models analysing change in weight (F (3, 119) = 1.80, p = 0.15) or change in tarsus length (F (3, 119) = 0.24, p = 0.87) of the nestlings during the experiment. Tick infestation intensity was associated neither to changes in weight (F (1, 119) = 0.70, p = 0.41) nor to changes in tarsus length (F (1, 119) = 2.99, p = 0.09). There were significant differences in the change in weight (F (3, 119) = 12.54, p < 0.01; Table 1) and tarsus length (F (3, 119) = 5.19, p < 0.01; Table 1) among bird species during the experiment. Post hoc analyses were not conducted for the latter factors because this is outside the scope of the current study.
Discussion
The majority of studies that previously investigated host specificity in ticks have examined the number of host species used (for a review, see McCoy et al. 2013), whereas few studies have investigated feeding performance on different host species (but see, e.g. Labruna et al. 2002; Olegário et al. 2011; Martins et al. 2012). Host specialisation of endophilic ticks, of which host range and dispersal can be expected to be low in comparison to exophilic ticks (Salman and Tarrés-Call 2012), has only been investigated in the seabird tick Ixodes uriae (McCoy et al. 2013; Dietrich et al. 2013). The current study is the first qualitative test of host specificity of an endophilic tick in a terrestrial ecosystem.
The results indicate that I. arboricola ticks that originate from great tits and have fed on these birds exclusively for several generations feed readily and with success on a number of other, less abundant bird species. This is not surprising since I. arboricola has previously been found on these bird species (Walter et al. 1979). We cannot exclude the possibility that there are specifically adapted genotypes that perform better on these less abundant hosts, but such a scenario is very unlikely because parasites are, in general, adapted to the most commonly encountered host (Legros and Koella 2010). Nevertheless, we found that feeding success was not higher on the most abundant and principal hosts of I. arboricola, i.e. great and blue tits (Walter et al. 1979; Heylen et al. 2014), than on the much less abundant hosts, i.e. nuthatches and pied flycatchers. Hence, great and blue tits, nuthatches and pied flycatchers can all be considered suitable host species for I. arboricola. This finding supports the hypothesis that host use by ticks is by host ecology rather than host specialisation (Klompen et al. 1996; Nava and Guglielmone 2013).
Even though overall tick success was high across the four bird species, we found some variation across species that may be explained by particular host traits. The highest attachment success at day 0 was found for nuthatches, whereas no effect of host species was observed for tick attachment success at day 2, engorgement weight and tick survival. This contrasts with studies on host specificity in the seabird tick I. uriae, where host origin affected feeding success but not host attraction on different host species (Mccoy and Boulinier 2002; Dietrich et al. 2013). Given that nuthatches are relatively uncommon hosts, it seems unlikely that high attractiveness to ticks would be due to host preference. Rather, we suggest that it may have been a by-product of nuthatch nestlings being considerably larger and heavier than the other bird species. Since the same size of cotton bag was used for all birds, nuthatches may have been easier for ticks to detect.
Moulting time was longest for nymphs that fed on nuthatches. This suggests that these birds are the least suitable hosts for I. arboricola because moulting time of ixodid ticks is usually longer on unsuitable hosts (Labruna et al. 2009; Olegário et al. 2011). This is not in line with the prediction that parasites are better adapted to the most frequently encountered hosts because then the lowest success should have been found on pied flycatchers (Legros and Koella 2010). Since the difference in moulting time across species is rather small and we found no differences in engorgement weight and moulting success across species, we expect the difference in moulting time not to be due to host specificity. Rather, we expect that the immune system of nuthatches was better developed at the moment of infestation because nuthatch development takes longer and nestlings were older—albeit at a similar stage of morphological development—at the moment of infestation than the other birds. The current data do not allow further testing of this hypothesis.
The proportion of ticks collected from the nestboxes was lowest for pied flycatcher nests. Since we only collected nymphs from the top of the nestbox, this result can be due to lower survival of ticks and/or to a lower proportion of ticks that actually moved to the top of the box. Survival in pied flycatcher nests might be low because ticks are prone to desiccation (Sonenshine 1991). Pied flycatcher nests are considerably drier than nests of great and blue tits because of the nest material used and the lower total biomass of the nestlings (Lundberg and Alatalo 1992). At the same time, the outer layer of pied flycatcher nests, a woven structure of bark and twigs, may be more difficult for ticks to manoeuvre through than the dense cluster of fresh moss encountered in tit nests and loose lining material in nuthatch nests. Hence, fewer ticks may have been able to move to the top of the nest box where we collected them. An untested hypothesis is that pied flycatchers are better in grooming, i.e. removing ectoparasites, than other birds. If this would be true, however, one would expect differences in tick infestation intensity, and we found no such differences when nestlings were inspected 2 days after infestation.
We found no effect of I. arboricola on nestling growth for any of the bird species investigated here. Whereas many tick species have strong direct effects on their hosts (McCoy et al. 2013), virulence, i.e. the impact on the fitness of the host, of I. arboricola on great tits has previously been shown to be low (Heylen and Matthysen 2010; Heylen and Matthysen 2011). We previously argued that low virulence can be expected in an endophilic tick which is highly dependent on individual hosts that return to the same cavity and on transmission from adult birds to their offspring (Heylen and Matthysen 2011). The current study suggests that low virulence of I. arboricola also applies to other cavity-nesting birds, thus enhancing its chances for persistence in cavities and dispersal among cavities inhabited by multiple host species. This may in turn increase the circulation of pathogens such as Rickettsia sp. and Borrelia burgdorferi s.l., for which I. arboricola is a known carrier (Špitalská et al. 2011; Heylen et al. 2013).
In this study, we focused on juveniles of four small songbirds with similar ecology and a large overlap in use of nest cavities. There are also occasional reports on I. arboricola infestation of larger cavity-nesting birds such as tawny owls (Strix aluco), Western jackdaws (Corvus monedula) and common starlings (Sturnus vulgaris) (Hudde and Walter 1988). Transmission of ticks to these species may be more limited than among the small passerines in the current study because these birds do not breed or roost in the same type of cavity as the principal hosts of I. arboricola (Mainwaring 2011; Heylen et al. 2014). Thus, we cannot exclude the possibility that host specificity occurs in I. arboricola across a wider range of host types, even including the possibility of specialised host races, as is the case in I. uriae infesting seabird species with different nest sites within a mixed colony (McCoy et al. 2001). Experimental infestations of additional cavity-nesting bird species will be required to test these scenarios.
In conclusion, the current study showed that I. arboricola nymphs readily feed on nestlings of pied flycatchers, great and blue tits and nuthatches, with some minor differences in feeding performance which may be accredited to the physiological and ecological differences among these birds. The results of the current study support recent suggestions that host specificity in ticks is limited by ecological similarities among hosts rather than host use in strict sense.
References
Bates D, Maechler M, Bolker B (2013) lme4: linear mixed-effects models using Eigen and S4
Dhondt AA (2010) Effects of competition on great and blue tit reproduction: intensity and importance in relation to habitat quality. J Anim Ecol 79:257–265. doi:10.1111/j.1365-2656.2009.01624.x
Dietrich M, Lobato E, Boulinier T, McCoy KD (2013) An experimental test of host specialization in a ubiquitous polar ectoparasite: a role for adaptation? J Anim Ecol. doi:10.1111/1365-2656.12170
Drès M, Mallet J (2002) Host races in plant–feeding insects and their importance in sympatric speciation. Philos Trans R Soc B 357:471–492. doi:10.1098/rstb.2002.1059
Fargallo JA, Johnston RD (1997) Breeding biology of the blue Tit Parus caeruleus in a montane Mediterranean deciduous forest: the interaction of latitude and altitude. J Fur Ornithol 138:83–92. doi:10.1007/bf01651654
Giorgi MS, Arlettaz R, Guillaume F et al (2004) Causal mechanisms underlying host specificity in bat ectoparasites. Oecologia 138:648–654. doi:10.1007/s00442-003-1475-1
Gómez-Díaz E, Morris-Pocock JA, González-Solís J, McCoy KD (2012) Trans-oceanic host dispersal explains high seabird tick diversity on Cape Verde islands. Biol Lett 8:616–619. doi:10.1098/rsbl.2012.0179
Gosler A (1993) The great tit. Hamlyn, London
Heylen DJA, Matthysen E (2010) Contrasting detachment strategies in two congeneric ticks (Ixodidae) parasitizing the same songbird. Parasitology 137:661–667. doi:10.1017/s0031182009991582
Heylen DJA, Matthysen E (2011) Differential virulence in two congeneric ticks infesting songbird nestlings. Parasitology 138:1011–1021. doi:10.1017/S0031182011000618
Heylen DJA, Tijsse E, Fonville M et al (2013) Transmission dynamics of Borrelia burgdorferi s.l. in a bird tick community. Environ Microbiol 29:663–673. doi:10.1111/1462-2920.12059
Heylen DJA, Van Oosten AR, Devriendt N et al (2014) Seasonal feeding activity of the tree-hole tick, Ixodes arboricola. Parasitology. doi:10.1017/S0031182014000225
Hillyard PD (1996) Ticks of north-west Europe: keys and notes for identification of the species. Published for the Linnean Society of London and the Estuarine and Coastal Sciences Association by Field Studies Council
Hoogstraal H, Aeschlimann A (1982) Tick-host specificity. Bull la Soc Entomol Suisse 55:5–32
Hudde H, Walter G (1988) Verbreitung und Wirtswahl der Vogelzecke Ixodes arboricola. Vogelwarte 34:201–207
Joshi A, Thompson JN (1995) Trade-offs and the evolution of host specialization. Evol Ecol 9:82–92. doi:10.1007/bf01237699
Kassen R (2002) The experimental evolution of specialists, generalists, and the maintenance of diversity. J Evol Biol 15:173–190. doi:10.1046/j.1420-9101.2002.00377.x
Kempf F, De Meeus T, Vaumourin E et al (2011) Host races in Ixodes ricinus, the European vector of Lyme borreliosis. Infect Genet Evol 11:2043–2048. doi:10.1016/j.meegid.2011.09.016
Klompen JSH, Black WC IV, Keirans JE, Oliver JH Jr (1996) Evolution of ticks. Annu Rev Entomol 41:141–161. doi:10.1146/annurev.en.41.010196.001041
Labruna MB, Souza SLP, Menezes AC et al (2002) Life-cycle and host specificity of Amblyomma tigrinum (Acari: Ixodidae) under laboratory conditions. Exp Appl Acarol 26:115–125. doi:10.1023/A:1020957122256
Labruna MB, Cabrera RR, Pinter A (2009) Life cycle of Ixodes luciae (Acari: Ixodidae) in the laboratory. Parasitol Res 105:1749–1753. doi:10.1007/s00436-009-1621-8
Legros M, Koella JC (2010) Experimental evolution of specialization by a microsporidian parasite. BMC Evol Biol 10:1471–2148. doi:10.1186/1471-2148-10-159
Liebisch G (1996) Biology and life cycle of Ixodes (Pholeoixodes) arboricola Schulze and Schlottke, 1929 (Ixodidae). In: Mitchell R, Hom DJ, Glen RN, Welboum WC (eds) Acarol. IX Vol. 1 Proc. Ohio Biological Survey, Columbus, OH, USA, pp 453–455
Lundberg A, Alatalo RV (1992) The pied flycatcher. Academic, San Diego
Magalhães S, Forbes MR, Skoracka A et al (2007) Host race formation in the Acari. Exp Appl Acarol 42:225–238. doi:10.1007/s10493-007-9091-0
Mainwaring MC (2011) The use of nestboxes by roosting birds during the non-breeding season: a review of the costs and benefits. Ardea 99:167–176. doi:10.5253/078.099.0206
Martins TF, Moura M, Labruna MB (2012) Life-cycle and host preference of Amblyomma ovale (Acari: Ixodidae) under laboratory conditions. Exp Appl Acarol 56:151–158. doi:10.1007/s10493-011-9506-9
Matthysen E (1998) The nuthatches. Academic, San Diego
Mccoy KD, Boulinier T (2002) Local adaptation of the ectoparasite Ixodes uriae to its seabird host. Evol Ecol Res 4:441–456
McCoy KD, Boulinier T, Tirard C, Michalakis Y (2001) Host specificity of a generalist parasite: genetic evidence of sympatric host races in the seabird tick Ixodes uriae. J Evol Biol 14:395–405. doi:10.1046/j.1420-9101.2001.00290.x
McCoy KD, Léger E, Dietrich M (2013) Host specialization in ticks and transmission of tick-borne diseases: a review. Front Cell Infect Microbiol. doi:10.3389/fcimb.2013.00057
Nava S, Guglielmone AA (2013) A meta-analysis of host specificity in Neotropical hard ticks (Acari: Ixodidae). Bull Entomol Res 103:216–224. doi:10.1017/S0007485312000557
Olegário MMM, Gerardi M, Tsuruta SA, Szabó MPJ (2011) Life cycle of the tick Amblyomma parvum Aragão, 1908 (Acari: Ixodidae) and suitability of domestic hosts under laboratory conditions. Vet Parasitol 179:203–208. doi:10.1016/j.vetpar.2011.01.056
Poulin R (2007) Evolutionary ecology of parasites. Princeton University Press, Princeton
R Core Team (2013) R: a language and environment for statistical computing.
Rotolo F, Munda M (2013) Parfm: parametric frailty models in R. J Stat Softw 51:1–20
Salman MD, Tarrés-Call J (2012) Ticks and tick-borne diseases: geographical distribution and control strategies in the Euro-Asia Region. CAB International, Wallingford
Sears BF, Schlunk AD, Rohr JR (2012) Do parasitic trematode cercariae demonstrate a preference for susceptible host species? PLoS One 7:e51012. doi:10.1371/journal.pone.0051012
Sonenshine DE (1991) Biology of ticks. Oxford University Press, Oxford
Špitalská E, Literák I, Kocianová E, Taragel’ová V (2011) The importance of Ixodes arboricola in transmission of Rickettsia spp., Anaplasma phagocytophilum, and Borrelia burgdorferi sensu lato in the Czech Republic, central Europe. Vector-Borne Zoonotic Dis 11:1235–1241. doi:10.1089/vbz.2010.0210
Timms R, Read AF (1999) What makes a specialist special? Trends Ecol Evol 14:333–334. doi:10.1016/s0169-5347(99)01697-3
Tripet F, Richner H (1997) The coevolutionary potential of a “generalist” parasite, the hen flea Ceratophyllus gallinae. Parasitology 115:419–427. doi:10.1017/s0031182097001467
Vermeersch G, Anselin A, Devos K, et al. (2004) Atlas van de Vlaamse broedvogels 2000–2002. Mededelingen van het Instituut voor Natuurbehoud 23, Brussel
Walter G, Liebisch A, Streichert J (1979) Untersuchungen zur Biologie und Verbreitung von Zecken (Ixodoidea, Ixodidae) in Norddeutschland. Angew Ornithol 5:65–73
White J, Heylen DJA, Matthysen E (2012) Adaptive timing of detachment in a tick parasitizing hole-nesting birds. Parasitology 139:264–270. doi:10.1017/s0031182011001806
Acknowledgments
We would like to thank F. J. B. Brosens, J. Elst and H. Pauwels for field assistance and the owners of Sterbos, Wuustwezel, for the permission to use this study area. This research was supported by FWO grant G.0049.10 to EM and an FWO postdoctoral fellowship to DJAH. The experimental protocol was approved by the Ethical Committee of the University of Antwerp and experiments on birds were licensed by the Agentschap voor Natuur en Bos.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Van Oosten, A.R., Heylen, D.J.A. & Matthysen, E. Host specificity of a bird-specialised endophilic ectoparasite, the tree-hole tick Ixodes arboricola . Parasitol Res 113, 4397–4405 (2014). https://doi.org/10.1007/s00436-014-4116-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-014-4116-1