
Abstract
In a bimanual-bisequential version of the serial reaction time (SRT) task participants performed two uncorrelated key-press sequences simultaneously, one with fingers of the left hand and the other with fingers of the right hand. Participants responded to location-based imperative stimuli. When two such stimuli appeared in each trial, the results suggest independent learning of the two sequences and the occurrence of intermanual transfer. Following extended practice in Experiment 2, transfer of acquired sequence knowledge was not complete. Also in Experiment 2, when only one stimulus appeared in each trial specifying the responses for both hands so that there was no basis for separate stimulus–stimulus or separate response–effect learning, independent sequence learning was again evident, but there was no intermanual transfer at all. These findings suggest the existence of two mechanisms of sequence learning, one hand-related stimulus-based and the other motor-based, with only the former allowing for intermanual transfer.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Effector-related sequence learning in a bimanual-bisequential serial reaction time task
The ability to acquire knowledge about sequential regularities is one of the characteristic features of human cognition. Since its introduction by Nissen and Bullemer (1987) the serial reaction time (SRT) task has become the dominant tool for exploring the mechanisms underlying sequence learning (for reviews see, e.g., Clegg, DiGirolamo, & Keele, 1998; Hoffmann, 2001; Rhodes, Bullock, Verwey, Averbeck, & Page, 2004). On each trial in a typical SRT experiment a stimulus appears in one of several locations, and participants are instructed to press the key assigned to that location as quickly as possible. The critical manipulation entails the introduction of structural redundancies in the stimulus sequence, typically the repetition of a fixed sequence. Following sufficient practice, sequence learning is usually demonstrated by a decrease of performance when the fixed sequence is replaced with a random sequence.
Because of the nature of a typical SRT task a fixed sequence of stimuli (S1–S2–S3–S4...) is confounded with a fixed sequence of responses (R1–R2–R3–R4...) as well as with regularities between responses and subsequent stimuli (R i –Si+1). Therefore, performance benefits for structured sequences can result both because participants learn to anticipate forthcoming stimuli (S–S learning; R–S learning) and because they learn to prepare forthcoming responses in advance (R–R learning). Indeed, there is evidence that each of these regularities contributes to sequence learning (S–S learning: e.g., Howard, Mutter, & Howard, 1992; Koch & Hoffmann, 2000; Remillard, 2003; R–R learning: e.g., Hoffmann & Koch, 1997, 1998; Hoffmann, Martin, & Schilling, 2003; Hoffmann & Sebald, 1996; Nattkemper & Prinz, 1997; Rüsseler & Rösler, 2000; R–S learning: e.g., Hazeltine, 2002; Hoffmann, Sebald, & Stöcker, 2001; Stöcker & Hoffmann, 2004; Stöcker, Sebald, & Hoffmann, 2003; Ziessler, 1998; Ziessler & Nattkemper, 2001). The question, then, arises to what degree the different regularities are learned independently of one another or in an integrated fashion (Deroost & Soetens, 2006; Frensch & Miner, 1995; Mayr, 1996; Riedel & Burton, 2006; Rüsseler, Münte, & Rösler, 2002; Schmidtke & Heuer, 1997).
The interplay between independent and integrated sequence learning is what Keele, Ivry, Mayr, Hazeltine, and Heuer (2003) recently aimed to capture in a model. The authors propose two subsystems: A unidimensional, automatic system encompassing independent learning modules which operate on input from single dimensions regardless of whether or not the respective stimuli are attended to. Furthermore, an attentional, multidimensional system is assumed which associates task-relevant stimuli from different dimensions provided that they are correlated and attended to. Next, we discuss briefly to what extent the assumed automatic and independent learning of stimulus sequences on different dimensions has been substantiated.
Most of the experiments on independent sequence learning presented participants with a response-correlated stimulus sequence (S&R) accompanied by a sequence of stimuli or stimulus attributes (S) which were irrelevant for responding so that they could remain unattended (Deroost & Soetens, 2006; Mayr, 1996; Riedel & Burton, 2006; Rüsseler et al., 2002). All theses studies provided reliable evidence for learning of the S&R sequence. However, none of these studies—Mayr (1996) being the only exception—yielded evidence for independent learning of the response irrelevant S-sequence. These findings suggest that pure stimulus sequences are only learned if the respective stimuli are attended to (cf. Hoffmann & Sebald, 2005; Jiménez & Méndez, 1999), which is at odds with the assumption of unidimensional learning modules, which automatically register any redundancy in the order of stimuli. However, there are two studies which suggest independent learning at least of attended sequences (Frensch & Miner, 1995; Schmidtke & Heuer, 1997).
Frensch and Miner (1995, Experiment 2) alternately presented one of two letters and one of two graphical symbols. Both letters and symbols appeared in a fixed 5-element sequence creating a compound sequence which was repeated every ten trials. The assignment of responses to stimuli, however, was randomly changed from trial to trial so that letters and symbols were both response-relevant and hence attended to but not correlated with the order of responses. After some practice either the letter sequence or the symbol sequence was randomized. In the letter change group only RTs for letters and in the symbol change group only RTs for symbols increased, clearly indicating independent learning of the two sequences.
Schmidtke and Heuer (1997) presented in between each element of a sequence of visual stimuli either a high-pitched or a low-pitched tone. Participants were asked to execute manual key presses in response to the visual stimuli and to press a foot pedal whenever a high-pitched tone appeared. In one of the conditions the order of the visual stimuli as well as the order of the tones followed a fixed 6-element sequence. After an acquisition phase the two sequences were shifted relative to each other so that the regularities between them were altered whereas the within-sequence regularities remained unaffected (‘shift probe’). Nevertheless, all responses were somewhat delayed indicating that participants had learned an integrated sequence of visual stimuli and tones. In additional test blocks either one of the sequences was replaced with a random sequence. In these ‘random probes’ RTs even increased for that task for which the stimulus sequence remained intact. This finding also indicated integrated learning. However, as RTs increased more in the ‘random probes’ than in the ‘shift probes’ Keele et al. (2003; p. 323) concluded that besides integrated learning of the compound sequence the two sequences were also at least partly learned independently of each other.
Taken together, both studies provide evidence in favor of independent learning of two response-relevant and hence attended stimulus sequences thus supporting the notion of independent albeit not automatic sequence learning modules (Keele et al., 2003). Frensch and Miner (1995) explicitly speculated that the letter and the symbol sequence they have used might be acquired in different compartments of working memory (Baddeley & Hitch, 1974): the letter sequence in the phonological loop and the symbol sequence in the visuo-spatial sketch pad. In case of Schmidtke and Heuer’s (1997) experiments, independent modules for visual and acoustic stimuli as well as for hand and foot movements are conceivable.
The study of Schmidtke and Heuer (1997) raises the question whether besides different stimuli also different effectors may constitute independent sequence learning modules. Several studies appear to rule out this possibility. These experiments indicated perfect or near to perfect transfer of acquired sequence knowledge from an effector used during practice to another effector in a subsequent transfer phase, which is at odds with an effector-specific representation of the acquired sequence knowledge (e.g., Cohen, Ivry, & Keele, 1990; Grafton, Hazeltine, & Ivry, 1998, 2002; Keele, Jennings, Jones, Caulton, & Cohen, 1995; Willingham, Wells, Farrell, & Stemwedel, 2000). However, Verwey and Clegg (2005) recently demonstrated an effector-specific, non-transferable component of sequence learning (see also Park & Shea, 2005; Verwey & Wright, 2004). Verwey and colleagues’ as well as Park and Shea’s experiments differ from previous investigations mainly in that participants completed a considerably higher number of sequence repetitions, suggesting that effector-specific sequence knowledge might be a result of extensive motor training.
In sum, the issue of independent sequence learning has been addressed repeatedly but is far from being settled. There is some evidence in favor of independent unidimensional learning modules provided that the sequential items are attended to. However, it still remains unclear what the dimensions are by which independent learning modules are to be separated. In the present study we were particularly interested in exploring to what extent different effectors may constitute independent learning modules. Except for the study of Schmidtke and Heuer (1997) this issue has not yet been addressed and is largely unresolved. This state of affairs is surprising as there are many actions which require the concerted use of different effectors. For example, typing or playing the piano involves finger movements of both hands in well ordered intertwined sequences. Likewise, everyday actions like opening a bottle of wine or brewing coffee require well coordinated and rather constant movement sequences involving both hands. It might well be that for such highly practiced coordinated movement sequences at least part of the sequence knowledge is acquired and stored in an effector-specific manner. For example, Daniel Barenboim’s playing of a piano sonata might rely at least in part on sequence knowledge separately stored ‘in the left and the right hand’.
The objectives of the present experiments were threefold. At the most general level, the experiments were designed to investigate the issue of concurrent learning of two uncorrelated S&R sequences. More specifically, we examined to what extent and under which conditions two simultaneously performed S&R sequences are learned independently of one another. Finally, we set out to determine to what extent any acquired independent sequence knowledge is based on representations relying on stimuli, that is, on sequence knowledge of perceptual origin which is transferable between effectors, or on representations of effector-specific movements, that is, on non-transferable knowledge about response sequences.
Experiment 1
In the present experiments participants practiced a repeating sequence of bimanual keypresses. On each trial two imperative stimuli appeared simultaneously, one for each hand, and participants were instructed to respond as simultaneously as possible with the appropriate fingers (for a similar procedure, see van der Graaf, de Jong, Maguire, Meiners, & Leenders, 2004). There was a fixed repeating sequence for the fingers of the left hand and another uncorrelated repeating sequence for the fingers of the right hand. Together these two hand-related sequences established a complex repeating compound sequence. This setting imitates requirements of coordinated hand movements in response to different environmental aspects.
Following extensive practice, three different types of test blocks were introduced: First, only one of the two hand-related sequences was replaced with a pseudo-random sequence. Second, both hand-related sequences were abolished, that is, both hands responded to a random sequence. Finally, transfer blocks were implemented in which the sequence practiced with the left hand was transferred to the right hand and vice versa.
In order to assess the amount of independent and integrated sequence learning, errors and RTs were evaluated. Errors were calculated for each hand separately whereas mean RTs were calculated for both hands together. As participants were instructed to execute the two responses simultaneously a delay in responding with one hand would also delay the response with the other hand so that a separate analysis of hand-related RTs does not make sense.
Assuming that on each trial the two required responses are selected more or less sequentially, the following data pattern would indicate independent learning of the two S&R sequences: Abolishing only one of the two sequences should result in a selective increase of errors in the respective hand and an increase of mean RTs, whereas abolishing both sequences should cause an increase of errors in both hands and a more pronounced increase of mean RTs. In contrast, learning of the compound sequence would be indicated by an equal increase of errors and RTs in both hands irrespective of whether the sequence of either of the hands or of both hands are abolished.
Furthermore, in case of independent learning the amount of intermanual transfer can be assessed by the performance in the transfer blocks. Better performance in the transfer blocks compared to performance in the test blocks in which both of the sequences are abolished would show that responding to a random sequence with one hand and to a transferred sequence with the other hand (i.e., a practiced sequence carried out with the ‘unpracticed’ hand) is easier than responding to two random sequences. Such an advantage would suggest that at least part of the sequence knowledge has been transferred from the practiced to the ‘unpracticed’ hand. If however, performance in transfer blocks equals performance with two random sequences, hand-related but non-transferable sequence knowledge is implicated.
Method
Participants
Twenty-four individuals (14 women; mean age 22.5 years) volunteered to participate in Experiment 1 either in partial fulfillment of course requirements or for a payment of €15. Twenty participants reported to be predominantly right-handed, the remaining four asserted to be predominantly left-handed.
Task and design
The presence or absence of the fixed sequences constituted the within-subjects factor. In particular, the following types of test blocks were implemented (see also Table 1): First, the fixed sequence participants had practiced with their left hand was replaced with a pseudo-random sequence while the right-hand sequence was retained (test block L rand). Second, the sequence participants had practiced with their right hand was replaced with a pseudo-random sequence while the left-hand sequence was retained (R rand). Third, both fixed sequences were replaced with pseudo-random sequences in the same test block (LR rand). In addition to that, there were test blocks in which, fourth, the sequence practiced with the left hand was transferred to the right hand while a pseudo-random sequence was presented for the left hand (L trans), and, fifth, the sequence practiced with the right hand was transferred to the left hand while a pseudo-random sequence was presented for the right hand (R trans). The sequences were transferred from the practiced to the unpracticed hand in such a way that the left-to-right ordering of stimulus (and key) locations was maintained, that is, the sequences were not mirrored.
Apparatus and materials
Stimulus presentation and response registration was controlled by the E-Prime software package (Schneider, Eschman, & Zuccolotto, 2002). Participants used a standard QWERTZ keyboard for responding. Stimuli were presented to participants on a 17-inch computer monitor. Responses and reaction times were recorded separately for the left and the right hand.
The imperative stimuli were asterisks 5 mm in diameter presented in black on a white background. Asterisks could appear in any one of six horizontally aligned locations on the screen, each of which was marked by a horizontal line 8 mm in length also appearing in black. The asterisks were presented centered above these lines. The lines (locations) were arranged in two groups of three with a distance of 45 mm between the groups. Within each group the lines were 6 mm apart.
The six keys S, D, F, J, K, and L on the keyboard served as response keys and were assigned from left to right to the six lines (locations) on the screen. The response keys in turn were assigned from left to right to the ring, middle, and index finger of the left hand and the index, middle, and ring finger of the right hand.
Two imperative stimuli appeared simultaneously on every trial: one stimulus in one of the locations on the left-hand side (referred to here as 1, 2, 3, from left to right), and another stimulus in one of the locations on the right-hand side (also referred to here as 1, 2, 3, from left to right). During training, the left-hand stimuli followed a repeating sequence independently of the right-hand stimuli, which followed another repeating sequence.
A 5-element sequence (32121) and a 6-element sequence (121323) were used. The 6-element sequence is a second-order conditional sequence, that is, at least two preceding elements are required to predict the next one in the sequence. The 5-element sequence contains two first-order conditional transitions (3–2 and 2–1) and two-third-order conditional transition (321−2 and 121−3). Because the two sequences are of different length, they are uncorrelated and establish a common dual-stimulus sequence which repeats every 30 trials.
Pseudo-random sequences were 90 elements long so that each matched the length of an entire test block. Furthermore, they were constructed to resemble the fixed sequences which they replaced in that stimuli appeared with the same frequency and did not repeat on consecutive trials. From a large set of such pseudo-random sequences as many were selected as there were test blocks in the experiment under the constraint that the selected pseudo-random sequences shared as few triples as possible with the to-be-replaced fixed sequence. Specifically, out of the total of 88 triples (3-tuples) contained in each of those pseudo-random sequences selected to replace the 5-element sequence (triples wrapping around from the last to the first elements in a 90-element pseudo-random sequence were not counted because no such wrap-around occurred in the test blocks), either 45 or 46 triples matched one of the 5 triples contained in the 5-element sequence. Similarly, each of those pseudo-random sequences replacing the 6-element sequence contained between 29 and 32 triples (out of a total of 88) matching one of the 6 triples in the 6-element sequence. The same pseudo-random sequences were used for each participant.
Procedure
Participants were tested individually. Half of the participants practiced the 5-element sequence with the left hand and the 6-element sequence with the right hand, while the assignment was reversed for the remaining participants.
The experiment was conducted in three sessions scheduled for different days with a maximum of 1 day between any two consecutive sessions. Session 1 started with a warm-up block in which pseudo-random sequences were presented for both hands, followed by ten fixed-sequence blocks. Session 2 comprised 15 blocks, the first seven of which were fixed-sequence blocks. Beginning with the eighth block (i.e., after 408 repetitions of the 5-element sequence and 340 repetitions of the 6-element sequence), four test blocks alternated with four fixed-sequence blocks. The four test blocks were: one L rand, one R rand, and two LR rand. The order of these test blocks was counterbalanced across participants with the pair of LR rand test blocks being treated as one entity. Session 3 started with five fixed-sequence blocks, and beginning with the sixth block (i.e., after a total of 624 repetitions of the 5-element sequence and 520 repetitions of the 6-element sequence, not counting sequence repetitions in session 2 L rand and R rand test blocks), four test blocks alternated with four fixed-sequence blocks as described for session 2. The order of presentation of these test blocks was also counterbalanced across participants with the additional constraint that no participant received the same ordering of test blocks as in session 2. Finally, blocks 14 and 15 were test blocks of the L trans and R trans type. The order in which these transfer blocks appeared was counterbalanced across participants independently of the counterbalancing of the first four test blocks. The session concluded with a final fixed-sequence block.
Each fixed-sequence block comprised 120 trials, the warm-up block and all of the test blocks comprised 90 trials each. Each fixed-sequence block began at a different position in the compound 30-element sequence established by the two hand-related sequences. Each trial began with the simultaneous presentation of two imperative stimuli. As soon as the participant had executed two responses the next stimuli were presented. A response–stimulus interval (RSI) of 0 ms was primarily chosen in order to optimize conditions for the acquisition of effector-specific sequence knowledge. It has been suggested that the absence of an RSI may be advantageous for effector-specific learning (Verwey & Wright, 2004), although it certainly is not a necessary condition as demonstrated by Verwey and Clegg’s (2005) finding of effector-specific learning at an RSI of 200 ms. Additionally, the absence of an RSI may have resulted in limited explicit learning (cf. Destrebecqz & Cleeremans, 2001).Footnote 1
When one or both of the participant’s responses were incorrect the German word for error (‘Fehler’) flashed briefly (for 27 ms; 2 refresh cycles of the monitor) in red color below the row of location lines.
Prior to session 1 participants received written instructions presented on the screen. Participants were informed about the assignment of locations on the screen to keys on the keyboard and to fingers of the two hands as described above. Participants were told that two asterisks would appear in two of the locations in every trial, indicating which pair of keys to press. No mention was made of sequences. Both speed and accuracy were stressed in the instructions. Furthermore, participants were instructed to perform the two responses in each trial as simultaneously as possible. Instructions were repeated prior to the start of sessions 2 and 3. In each session participants took self-terminated rest periods between blocks during which a text on the screen reminded the participants of the requirements regarding speed and accuracy as well as the synchrony of the responses. The participants initiated each block by pressing the space bar.
After completing the SRT task in session 3, participants were debriefed about the presence and the exact length of the two sequences and were asked to recall both of them in full length, beginning with the sequence they had practiced with the left hand. More specifically, participants were asked to write down either the sequence of key presses or the sequence of stimuli and they were encouraged to guess if they could not recall parts of a sequence. They were also told that they could use their hands during recall and start at any position in the sequence.
Results
In order to focus presentation of results, only data from session 3 will be presented in detail. Session 2 data yielded largely the same results. For each family of pairwise comparisons (paired-samples t tests) we subjected P-values to the Bonferroni adjustment.
RTs from error trials (at least one incorrect response) were excluded from analysis (6.7%), as were outlier RTs (2.5 SD above or below the z-transformed mean RT as determined separately for every participant, every block, and every hand; 2.8%). Furthermore, we excluded RTs from those trials in which RTs for the left and right hand differed by more than 100 ms (2.9%). For the remaining trials the RTs of the left and the right hand were averaged. From these mean bimanual RTs for each trial the median RT was computed for every block. The means of the individual median RTs are shown in Fig. 1. For each block error rates were computed separately for both hands. The two test blocks of the type LR rand were treated as a single test block. RT costs as an index of sequence learning were computed as the difference between the median RT in a test block and the mean of the median RTs in baseline blocks. For test blocks L rand, R rand, and LR rand the baseline blocks were defined as the regular block preceding the first of these test blocks and the regular block following the last of these test blocks as well as those regular blocks in between (i.e., blocks 31, 33, 35, 37, and 39 in session 3). For test blocks L trans and R trans the baseline blocks were the regular blocks adjacent to these test blocks (i.e., blocks 39 and 42 in session 3). Error costs were computed in a manner analogous to RT costs. Error costs were obviously very small proportions, which raises the issue of whether parametric statistical analyses are appropriate for error cost data. Yet, none of the error costs variables differed significantly from a normal distribution, all Kolmogorov–Smirnov Zs between 0.345 and 1.171, all P > 0.128. Therefore, we decided to analyze the data with the more powerful parametric procedures instead of non-parametric statistical tests. Whenever necessary, the degrees of freedom in repeated-measures analyses of variance were adjusted with the Greenhouse–Geisser epsilon (ε GG) in order to correct for any significant violations (Mauchly test) of the sphericity assumption. The unadjusted degrees of freedom are reported together with the respective ε GG, if a correction has been carried out. The corresponding reported P-values reflect the adjusted degrees of freedom.

Means of individual median RTs in Experiment 1. For presentation purposes, test blocks are ordered in a consistent manner; in reality, the order of test blocks was counterbalanced across participants. Also, both for session 2 and session 3 the two LR rand test blocks were combined into a single test block for presentation purposes. This is why there is no block 25 and no block 38 in this figure
There were significant RT costs in every test block, all t(23) > 7.35, all P ≤ 0.001. For relevant RT cost means and error cost means see Fig. 2a.

Mean RT and error costs (error bars represent standard errors of the means) in each of the test blocks in session 3 of Experiment 1 separately for the entire sample (a) and for the sub-sample of participants who did not possess above-chance explicit knowledge about the sequences (b)
Independent learning of hand-related sequences: RT costs
An ANOVA on session 3 RT costs with repeated measures on the factor test block (L rand, R rand, LR rand) revealed a significant main effect, F(2, 46) = 16.15, P ≤ 0.001, reflecting that RT costs (a) did not differ significantly between test blocks in which only one sequence was randomized while the other was retained (L rand and R rand), t(23) = 0.66, but (b) were smaller in those test blocks than when both sequences were randomized (test block LR rand), both t(23) > 4.90, both P ≤ 0.001. There was no negative correlation between the individual RT costs in test blocks L rand and R rand, r(24) = 0.21, P ≤ 0.33, indicating that there was no trade-off between learning of the left-hand sequence and learning of the right-hand sequence.
Independent learning of hand-related sequences: error costs
An ANOVA on session 3 error costs with repeated measures on the factors Hand (left, right) and Test Block (L rand, R rand, LR rand) revealed a significant main effect test block, F(2, 46) = 5.89, P ≤ 0.01, indicating generally higher error costs in test block LR rand (2.9%) than in test blocks L rand (1.6%) and R rand (1.8%), both t(23) > 3.02, both P ≤ 0.05, between which error costs did not differ significantly, t(23) = 0.52. The main effect Hand was not significant, F(1, 23) = 2.68, P ≤ 0.115, but the critical interaction between the factors hand and test block was significant, F(2, 46) = 20.34, P ≤ 0.001, ε GG = 0.743.
In order to unpack this interaction, separate ANOVAs on error costs with repeated measures on the factor test block were computed for the left and the right hand. Both of these analyses revealed a significant main effect test block, both F(2, 46) > 15.37, both P ≤ 0.001, both ε GG < 0.824. Significant right-hand error costs were evident in test blocks R rand and LR rand, both t(23) > 4.98, both P ≤ 0.001, between which error costs did not differ significantly, t(23) = 0.54. Right-hand error costs were, however, significantly higher in those test blocks than in test block L rand, both t(23) > 4.09, both P ≤ 0.001, in which right-hand error costs were not significant, t(23) = 0.15. Significant left-hand error costs were evident in test blocks L rand and LR rand, both t(23) > 4.40, both P ≤ 0.001, between which error costs did not differ significantly, t(23) = 1.63. Left-hand error costs were, however, significantly higher in those test blocks than in test block R rand, both t(23) > 4.62, both P ≤ 0.001, in which left-hand error costs were not significant, t(23) = −0.58. In other words, performance suffered only for that hand which lost its sequence while performance with the other hand, which retained its sequence, was unaffected.
Intermanual transfer: RT costs
An ANOVA on session 3 RT costs with repeated measures on the factor transfer block (LR rand, L trans, R trans) revealed a significant main effect, F(2, 46) = 21.86, P ≤ 0.001, indicating increased RT costs in LR rand test blocks compared to both the L trans and the R trans test block, both t(23) > 5.48, both P ≤ 0.001, between which RT costs did not differ significantly, t(23) = 0.03. Transfer blocks were somewhat more regular than the LR rand block because on any given trial there are only two possible subsequent stimulus combinations compared to four possible subsequent stimulus combinations in test block LR rand. Consequently, the reduced costs in the transfer blocks might be due to within-block learning. In order to explore this possibility we ran an analysis including block half (first vs. second) as an additional within-subjects factor. However, the critical transfer block × block half interaction was not significant, F(2, 46) = 3.04, P ≤ 0.058. Moreover, higher RT costs in test block LR rand than in transfer blocks L trans and R trans, both t(23) > 3.19, both P ≤ 0.05, were already evident in the first block half, F(2, 46) = 9.61, P ≤ 0.001. In sum, performance suffered less when executing the transferred sequence together with a pseudo-random sequence than when executing two pseudo-random sequences indicating intermanual transfer.
Intermanual transfer: error costs
An ANOVA on session 3 error costs with repeated measures on the factors hand (left, right) and transfer block (LR rand, L trans, R trans) revealed a significant main effect transfer block, F(2, 46) = 8.11, P ≤ 0.005, ε GG = 0.789, indicating generally higher error costs in test block LR rand (2.9%) than in test blocks L trans (1.1%) and R trans (1.1%), both t(23) > 3.48, both P ≤ 0.01, between which error costs did not differ significantly, t(23) < 0.01. The main effect Hand was not significant, F(1, 23) = 2.63, P ≤ 0.118, but the critical interaction between the factors hand and transfer block was significant, F(2, 46) = 11.04, P ≤ 0.001.
In order to unpack this interaction, separate ANOVAs on error costs with repeated measures on the factor transfer block were computed for the left and the right hand. Both of these ANOVAs yielded a significant main effect transfer block, both F(2, 46) > 7.12, both P ≤ 0.01. Significant right-hand error costs were evident in test blocks LR rand and R trans, both t(23) > 3.27, both P ≤ 0.005, between which error costs did not differ significantly, t(23) = 1.68. Right-hand error costs were, however, significantly higher in these test blocks than in test block L trans, both t(23) > 3.18, both P ≤ 0.05, in which right-hand error costs were not significant, t(23) < 1. Significant left-hand error costs were present in test blocks LR rand and L trans, both t(23) > 3.43, both P ≤ 0.01, between which error costs did not differ significantly, t(23) = 0.42. Left-hand error costs were, however, significantly higher in these test blocks than in test block R trans, both t(23) > 2.61, both P ≤ 0.05, in which left-hand error costs were not significant, |t(23)| < 1. Consistent with RT results, the transferred sequence did not have as detrimental an effect on performance as a pseudo-random sequence. Instead, performance suffered only for that hand with which a random sequence was executed while no performance decrements were evident for the other hand with which the transferred sequence was executed.
Free recall
Participants’ performance in the post-experimental recall task was scored by determining the number of recalled triples which indeed were part of the respective repeating hand-sequence. Six participants recalled the complete 6-element sequence, four participants recalled the complete 5-element sequence, and only two participants recalled both sequences completely. These twelve participants were considered as having explicit sequence knowledge. The remaining twelve participants recalled a mean number of 2.25 triples (out of 6; SD = 0.75; min = 1; max = 3) from the 6-element sequence and a mean number of 1.42 triples (out of 5; SD = 1.31; min = 0; max = 3) from the 5-element sequence. These participants were considered as having only fragmentary sequence knowledge.
Explicit versus implicit sequence learning
In order to assess whether our results depend on the acquisition of explicit sequence knowledge, we repeated all analyses for the sub-sample of 12 participants who displayed only fragmentary explicit sequence knowledge. We obtained the same pattern of results as for the entire sample (see Fig. 2b).
Discussion
In a bimanual version of the SRT task, participants executed two uncorrelated S&R sequences simultaneously, one with the left hand and the other with the right hand. After extensive practice, either one of the two sequences or both were replaced with a pseudo-random sequence. The resulting RT costs were significantly larger when both sequences were randomized than when only one of the sequences was randomized. As participants suffered similarly from losing either of the two sequences and individual RT costs were not negatively correlated the data suggest independent learning of the two sequences. This conclusion is further supported by hand-related error costs, that is, left hand errors increased only when the sequence of the left hand was abolished and right hand errors increased only when the right-hand sequence was abolished.
Additionally, RT costs in transfer blocks in which the sequence practiced with the one hand was transferred (parallel-shifted, not mirrored) to the other hand (while the hand with which the transferred sequence had been practiced executed a pseudo-random sequence) were significantly smaller than when both sequences were abolished. This suggests that the acquired sequence knowledge allows for transfer between hands. Intermanual transfer was additionally confirmed by the fact that there were no significant error costs for that hand which executed a transferred sequence.
Thus, the results of Experiment 1 are consistent with independent learning of two uncorrelated, hand-related S&R sequences. As the same pattern of results was obtained for a sub-sample of participants who acquired only fragmentary if any explicit sequence knowledge, the underlying learning mechanisms presumably do not require the sequence structures to be recognized. Furthermore, the acquired sequence knowledge appeared to be effector-independent inasmuch as it was available for intermanual transfer. The question remains, then, what the acquired sequence knowledge may be based on. At least two possibilities are to be considered.
First, and probably most obvious, participants may have benefited from facilitation of the forthcoming stimulus locations for each of the two hands. Second, participants may have benefited from facilitation of the forthcoming responses for each hand, either by facilitation of the locations of the to-be-pressed keys (Willingham et al., 2000) or by facilitation of the to-be-moved fingers (R–R learning).
Experiment 2 was designed in order to better assess the different impact that stimulus and response facilitation may have on the observed sequence learning. For this purpose a new condition was introduced in which the responses of both hands were specified by only one stimulus so that there were no longer two different stimulus sequences available for separate learning. Secondly, we used longer and more complex sequences in Experiment 2 in order to make integrated learning of the compound sequence more unlikely. This should also render the acquisition of explicit sequence knowledge more difficult. Finally, the training was prolonged in order to increase the chances of obtaining a manifestation of non-transferable, hand-specific sequence knowledge.
Experiment 2
In Experiment 2 participants performed a bimanual SRT task essentially similar to the one described for Experiment 1 except for an additional variation of stimulus presentation: the dual-stimulus condition was a replication of Experiment 1, with two stimuli, one for each hand, appearing simultaneously on every trial. In contrast, in the single-stimulus condition the responses for the left and the right hand were specified by only one imperative stimulus. In both conditions participants practiced the same uncorrelated sequences, one with fingers of the left hand, the other with fingers of the right hand.
In the dual-stimulus condition we expected to find independent learning of the two sequences resulting in transferable sequence knowledge just like in Experiment 1. In the single-stimulus condition, however, which hardly allows for hand-related stimulus–stimulus (S–S) or response–stimulus (R–S) learning, weaker indications of (a) independent sequence learning and (b) intermanual transfer are to be expected, if and only if stimulus facilitation plays the crucial role for independent sequence learning.
Method
Because Experiment 2 was identical to Experiment 1 in most respects only the differences are described here.
Participants
Twenty-four individuals (13 women; mean age 23 years) volunteered to participate in Experiment 2 either in partial fulfillment of course requirements or for a payment of €25. Twenty participants reported to be predominantly right-handed, three asserted to be ambidextrous, and one participant was predominantly left-handed.
Apparatus and materials
In the dual-stimulus condition, asterisks again appeared in any one of six horizontally aligned locations. Unlike in Experiment 1, the six locations were not arranged in two groups of three. Instead, there was a distance of 6 mm between any two adjacent locations. Consequently, the visual separation of the two stimulus sequences was less obvious.
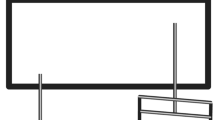
In the single-stimulus condition, the imperative stimuli (asterisks) could appear in any one of nine cells in a 3 × 3 matrix displayed centered on the screen, standing on one of its vertices (see Fig. 3). The length of each side of the cells was 15 mm. The rows and columns of the matrix were marked by straight lines 9 mm in length appearing at the lower left side and lower right side of the matrix, respectively.

Stimulus–response assignment in the two presentation modes in Experiment 2. The keys which participants were instructed to press simultaneously in response to the depicted stimulus display are colored dark gray
The left-hand keys S, D, and F were assigned compatibly from left to right to the rows of the matrix (numbered 1, 2, 3, from left to right), while the right-hand keys J, K, and L were assigned compatibly from left to right to the columns of the matrix (also numbered 1, 2, 3, from left to right). In this way, a single stimulus specified both a response for the left hand and a response for the right hand.
In both conditions, the sequences of the left and the right hand were determined by fixed sequences in regular blocks. An 8-element sequence (32131312) and a 9-element sequence (212132313) were used. In both sequences, prediction of the respective next element required the knowledge of up to four preceding elements, so that the sequences were not only more complex but also better balanced than in Experiment 1. When presented simultaneously, these two uncorrelated sequences establish a compound sequence which repeats every 72 trials.
The pseudo-random sequences were 144 elements long so that each matched the length of an entire test block. They were constructed to conform to properties of the fixed sequences they replaced as described for Experiment 1. Again, pseudo-random sequences were selected that shared as few quadruples (4-tuples) as possible with the to-be-replaced fixed sequence. Specifically, out of the total of 141 quadruples contained in each of the pseudo-random sequences selected to replace the 8-element sequence either 46 or 48 quadruples matched one of the 8 quadruples contained in the 8-element sequence. Similarly, each of the pseudo-random sequences replacing the 9-element sequence contained either 29 or 31 quadruples (out of a total of 141) matching one of the 9 quadruples in the 9-element sequence. The same pseudo-random sequences were used for each participant.
Procedure
Half of the participants were assigned to the dual-stimulus and the single-stimulus condition each. Within each condition, half of the participants practiced the 8-element sequence with the left hand and the 9-element sequence with the right hand while the assignment was reversed for the other participants.
Session 1 started with a warm-up block in which pseudo-random sequences were presented for both hands, followed by 15 fixed-sequence blocks. Session 2 comprised 20 fixed-sequence blocks. Session 3 started with five fixed-sequence blocks, and beginning with the sixth block (i.e., after 720 repetitions of the 8-element sequence and 640 repetitions of the 9-element sequence), five test blocks alternated with five fixed-sequence blocks. These test blocks were: L rand, R rand, LR rand, L trans, and R trans. The order of the first three and the last two of these test blocks was counterbalanced across participants independently of each other.
Each fixed-sequence block comprised 144 trials, as did all of the test blocks. The warm-up block contained only 72 trials. Each fixed-sequence block began at a randomly determined position in the 72-element compound sequence established by the two hand-related sequences.
Prior to session 1, participants received written instructions presented on the screen. For the dual-stimulus condition these were exactly the same as in Experiment 1. Participants in the single-stimulus condition were additionally informed about the assignment of rows and columns in the matrix to keys on the keyboard and to fingers of the two hands as described above. After completing the SRT task in session 3, participants completed a sequence recall task as described for Experiment 1.
Results
Again, we subjected P-values to the Bonferroni adjustment for each family of pairwise comparisons (paired-samples t tests).
RT and error data were prepared as described for Experiment 1. In the dual-stimulus condition RTs from 8.7% of trials were excluded (4.4% error trials, 2.6% outlier RTs, 1.6% trials with asynchronous responses) and in the single-stimulus condition RTs from 10% of trials were excluded (5.8% error, 2.9% outlier, 1.3% asynchronous). The means of the individual median RTs in each block are shown in Fig. 4. As described for Experiment 1, RT costs for each test block were computed as the difference between the median RT in that test block and the mean of the median RTs in the baseline regular blocks. For test blocks L rand, R rand, and LR rand, all adjacent blocks provided the data for the baseline (i.e., blocks 41, 43, 45, and 47). Accordingly, for the test blocks L trans and R trans, the adjacent blocks 47, 49, and 51 constituted the baseline. Error costs were computed in a manner analogous to RT costs. Again, none of the error cost variables differed significantly from a normal distribution, all Kolmogorov–Smirnov Zs between 0.481 and 1.152, all P > 0.140. Therefore, we decided to stick to parametric analysis with the degrees of freedom in repeated-measures ANOVAs adjusted for violations of the sphericity assumption as for Experiment 1.

Means of individual median RTs in Experiment 2 separately for the two presentation modes. For presentation purposes, test blocks are ordered in a consistent manner; in reality, the order of test blocks was counterbalanced across participants. The ordinate’s scale has been adjusted for data from session 3 (blocks 37–51) to aid comparison with data from Experiment 1
There were significant RT costs in every test block when including data from both presentation modes (the dual-stimulus and the single-stimulus condition), all t(23) > 6.65, all P ≤ 0.001, as well as when analyzing RT costs separately for each presentation mode, all t(11) > 4, all P ≤ 0.01. For relevant RT cost means and error cost means see Figs. 5 and 6a.

Mean RT and error costs (error bars represent standard errors of the means) in each of the test blocks in Experiment 2 separately for the dual-stimulus (a) and the single-stimulus (b) presentation mode

Mean RT and error costs (error bars represent standard errors of the means) in each of the test blocks in Experiment 2 separately for the entire sample (a) and for the sub-samples of participants who did not possess above-chance explicit knowledge about the sequences (b)
Independent learning of hand-related sequences: RT Costs
A mixed-factors ANOVA on RT costs with repeated measures on the factor test block (L rand, R rand, LR rand) and with presentation mode (dual-stimulus condition, single-stimulus condition) as the between-subjects factor revealed a significant main effect test block, F(2, 44) = 26.10, P ≤ 0.001, indicating higher RT costs in test block LR rand than in test block L rand and R rand, both t(23) > 5.58, both P ≤ 0.001, between which RT costs did not differ significantly, t(23) = 0.03. The main effect presentation mode was not significant, F(1, 22) = 0.16, but the interaction was, F(2, 44) = 3.45, P ≤ 0.05.
In order to unpack this interaction, separate ANOVAs on RT costs with repeated measures on the factor test block were computed for both presentation modes. In the dual-stimulus condition the data pattern of Experiment 1 was repeated: the main effect test block was significant, F(2, 22) = 28.77, P ≤ 0.001, indicating that RT costs did not differ significantly between blocks L rand and R rand, t(11) < 1.1, P > 0.91, but were lower in these blocks than in block LR rand, both t(11) > 6.05, both P ≤ 0.001. In the single-stimulus condition the main effect test block was also significant, F(2, 22) = 5.45, P ≤ 0.05, and RT costs did not differ between blocks L rand and R rand, t(11) = 0.83. However, different from the dual-stimulus condition only in block L rand were RT costs lower than in block LR rand, t(11) = 3.12, P ≤ 0.05, whereas the difference between blocks R rand and LR rand was not significant, t(11) = 2.48, P ≤ 0.092. Furthermore, the mean difference between RT costs in blocks L rand and R rand on the one hand and block LR rand on the other hand was substantially smaller (M = 28.9 ms) than in the dual-stimulus condition (M = 55.6 ms), F(1, 22) = 5.15, P ≤ 0.05, although RT costs in LR rand blocks did not differ significantly between the two conditions, t(22) = 1.56, P ≤ 0.133.
There was no negative correlation between individual RT costs in test blocks L rand and R rand, r(24) = 0.371, P ≤ 0.074 (dual-stimulus condition: r(12) = 0.521, P ≤ 0.082; single-stimulus condition: r(12) = 0.305, P ≤ 0.335), indicating that there was no trade-off between learning of the left-hand sequence and learning of the right-hand sequence.
Independent learning of hand-related sequences: error costs
A mixed-factors ANOVA on error costs with repeated measures on the factors hand (left, right) and test block and with presentation mode as the between-subjects factor revealed a significant interaction between the factors hand and test block, F(2, 44) = 8.38, P ≤ 0.005, ε GG = 0.767. The main effect test block approached but did not reach significance, F(2, 44) = 2.66, P ≤ 0.081. No other main effects or interactions were significant, all F < 0.80.
In order to further analyze the significant interaction, separate ANOVAs with repeated measures on the factor test block were computed for left-hand and for right-hand error costs. Because there were no significant interactions involving presentation mode this factor was dropped.
The ANOVA for left-hand error costs revealed a significant main effect test block, F(2, 46) = 4.97, P ≤ 0.05, indicating significant costs in test blocks L rand and LR rand, both t(23) > 2.85, both P ≤ 0.01, but not in block R rand, t(23) = 0.79, P ≤ 0.437. Costs in block L rand but not in block LR rand were significantly higher than in block R rand, t(23) = 2.89, P ≤ 0.05, and t(23) = 2.26, P ≤ 0.10, respectively. Costs did not differ between blocks L rand and LR rand, t(23) = 1.12.
The ANOVA for right-hand error costs revealed a significant main effect test block, F(2, 46) = 8.42, P ≤ 0.001, indicating significant error costs in test blocks R rand and LR rand, both t(23) > 4.56, both P ≤ 0.001, which did not differ from each other, t(23) = 1.09, but which were significantly higher than those in test block L rand, both t(23) > 2.83, both P ≤ 0.05, for which there were no significant error costs, t(23) = 0.78, P ≤ 0.445. Thus, as in Experiment 1, performance suffered only for that hand which lost its sequence while performance with the other hand, which retained its sequence, was unaffected.
Intermanual transfer: RT costs
A mixed factors ANOVA on RT costs with repeated measures on the factor transfer block (LR rand, L trans, R trans) and with presentation mode (dual-stimulus condition, single-stimulus condition) as the between-subjects factor revealed a significant main effect transfer block, F(2, 44) = 4.06, P ≤ 0.05. The critical transfer block × presentation mode interaction was significant, F(2, 44) = 4.37, P ≤ 0.05. No other main effects or interactions were significant, all F < 1.
In order to unpack the significant transfer block × presentation mode interaction, separate ANOVAs on RT costs with repeated measures on the factors block half (first, second) and transfer block were computed for data from each presentation mode. The factor block half was included to evaluate the within-block learning account of intermanual transfer as discussed for Experiment 1.
For data from the dual-stimulus condition, the ANOVA revealed a significant main effect transfer block, F(2, 22) = 10.21, P ≤ 0.001, indicating that RT costs were significantly higher in test block LR rand than in test blocks L trans and R trans, both t(11) > 2.82, both P ≤ 0.05. RT costs did not differ significantly between test blocks L trans and R trans, t(11) = 1.47. That is, as in Experiment 1, performance suffered less when executing the transferred sequence together with a pseudo-random sequence than when executing two pseudo-random sequences, thus indicating intermanual transfer. In contrast, for data from the single-stimulus condition there was no evidence for intermanual transfer as the main effect transfer block was not significant, F(2, 22) = 0.02.
Both in the dual-stimulus and in the single-stimulus condition neither the main effect block half was significant, both F(1, 11) < 0.10, nor was the block half × transfer block interaction significant, F(2, 22) = 2.60, P ≤ 0.097, and F(2, 22) = 2.91, P ≤ 0.076, respectively. Thus, there were no reliable indications of within-block learning in transfer blocks.
Intermanual transfer: Error costs
A mixed factors ANOVA on error costs with repeated measures on the factors Hand (left, right), block half (first, second), and transfer block, and with presentation mode as the between-subjects factor was computed. The interaction between the factors transfer block and presentation mode approached but did not reach significance, F(2, 44) = 3.01, P ≤ 0.06. The other main effects and interactions were not significant either, all F < 2.78. Separate ANOVAs with repeated measures on the factors hand and test block for the dual-stimulus and the single-stimulus condition revealed no significant effects, all F < 3.18.
Free recall
None of the participants recalled either of the two sequences completely. In order to split the sample into two sub-samples as in Experiment 1, we considered participants as possessing at least noticeable explicit sequence knowledge if they recalled 3 or more quadruples from the 8-element or 4 or more quadruples from the 9-element sequence. The other participants were considered as possessing negligible explicit sequence knowledge.
In the dual-stimulus condition, six participants possessed noticeable explicit sequence knowledge. They recalled a mean number of 2.7 quadruples from the 8-element sequence (SD = 1.21; min = 1; max = 4) and a mean number 4.8 quadruples (SD = 0.75; min = 4; max = 6) from the 9-element sequence. The remaining six participants with negligible explicit sequence knowledge recalled a mean number of 1.2 quadruples (SD = 0.41; min = 1; max = 2) from the 8-element sequence and a mean number of 1.5 quadruples (SD = 1.05; min = 0; max = 3) from the 9-element sequence.
In the single-stimulus condition seven participants were classified as possessing noticeable explicit sequence knowledge: they recalled a mean number of 3 quadruples from the 8-element sequence (SD = 1.29; min = 1; max = 4) and a mean number 5 quadruples (SD = 1.15; min = 3; max = 6) from the 9-element sequence. The remaining five participants recalled a mean number of 1.8 corresponding quadruples (SD = 0.45; min = 1; max = 2) from the 8-element sequence and a mean number of 1.8 corresponding quadruples (SD = 1.30; min = 0; max = 3) from the 9-element sequence.
Explicit versus implicit sequence learning
The sub-sample of participants with only negligible explicit sequence knowledge showed almost the same data pattern as the entire sample (see Fig. 6b).
Discussion
Participants performed the bimanual-bisequential SRT task with either two distinct stimuli or only one stimulus specifying the required simultaneous responses of the left and the right hand. Following extensive practice, independent learning of the two sequences was tested in a series of test blocks. RT costs were higher when both sequences were replaced with a pseudo-random sequence than when only one of the sequences was abolished. As individual RT costs in the test blocks with only one of the sequences abolished were not negatively correlated independent learning of the two sequences is indicated. The same pattern of results was obtained for a sub-sample with only negligible if any explicit sequence knowledge. Therefore, the learning of the hand-related sequences is presumably independent of the recognition of the sequences.
Concerning the two stimulus presentation modes, the critical increase of RT costs with both compared to only one of the sequences randomized turned out to be clearly more pronounced with two stimuli compared to one stimulus, although the pattern of hand-related error costs corroborates the conclusion that independent sequence learning had occurred.
Intermanual transfer also was significantly influenced by the stimulus presentation mode: transfer from the practiced to the unpracticed hand was evident in the dual-stimulus condition whereas in the single-stimulus condition, the data provided no evidence for intermanual transfer. This finding is in line with the notion that the hand-related but nevertheless transferable sequence knowledge we found in Experiment 1 and in the dual-stimulus condition of Experiment 2 most probably relies on the facilitation of forthcoming stimuli in the hand-related stimulus sequences. This can take the form of both S–S and R–S learning. If there is only one stimulus for both sequences, separate hand-related stimulus sequences no longer exist so that neither hand-related S–S nor R–S learning can take place. Nevertheless, the data, in particular the error costs, suggest that also in the single-stimulus condition hand-related sequence knowledge has been acquired. As such knowledge could hardly rely on stimulus facilitation (i.e., S–S or R–S learning) it most likely reflects response facilitation (R–R learning), that is, hand-specific motor sequence knowledge, which does not lend itself to intermanual transfer.
It deserves mention that intermanual transfer was weaker in the dual-stimulus condition of Experiment 2 than in Experiment 1. This becomes especially apparent if RT costs in transfer blocks L trans and R trans are compared with RT costs in test blocks R rand and L rand, respectively, that is, if one compares performance decrements resulting from executing with one hand a regular and with the other hand a random sequence depending on whether the regular sequence is performed by the hand which has previously practiced this sequence or by the other hand which has previously practiced another sequence. In Experiment 1 there is virtually no difference between these two conditions (see Fig. 2) suggesting that intermanual transfer is nearly complete. In contrast, in Experiment 2 (see Fig. 6a), transfer blocks (L trans, R trans) yielded higher RT costs than test blocks (R rand, L rand), F(1, 22) = 24.27, P ≤ 0.001.
We account for this incompleteness of intermanual transfer by the same reasoning as for the failure of transfer in the single-stimulus condition, that is, we argue that despite the presentation of separate stimuli, learning of hand-related stimulus sequences has been hampered compared to Experiment 1 for the following reasons: first, because the separation of the hand-related stimuli was less obvious; second, because the amount of training was increased which is known to make the occurrence of effector-specific (non-transferable) sequence knowledge more likely (e.g., Park & Shea, 2005; see also Bapi, Doya, & Harner, 2000); and third, because the amount of explicit sequence knowledge was substantially reduced, which has been suggested to make implicit effector-specific learning more prominent (cf. Hikosaka, Nakamura, Sakai, & Nakahara, 2002).Footnote 2
General discussion
The present experiments explored sequence learning in a situation in which participants simultaneously responded with the left and the right hand to uncorrelated repeating stimulus sequences, thus mimicking everyday actions requiring the coordinated use of both hands in a recurring canonical way. There were three objectives:
First, the experiments investigated to what extent the two hand-related stimulus-response sequences (S&R) are learned independently of each other. To the best of our knowledge the present experiments constitute the first direct investigation of the learning of two simultaneously executed uncorrelated S&R sequences. Second, we explored the extent to which any acquired independent sequence knowledge was bound to the respective hand or available for intermanual transfer. Finally, the experiments aimed to discover the representational nature of any independently acquired hand-related sequence knowledge. In particular, we assumed that transferable sequence knowledge primarily relies on stimulus facilitation resulting from either S–S or R–S learning, whereas non-transferable knowledge should primarily rely on response facilitation resulting from R–R learning. Accordingly, we examined whether the amount of transferable sequence knowledge is reduced in favor of non-transferable sequence knowledge when the two response sequences are triggered by a single stimulus instead of two stimuli, so that the acquisition of hand-related stimulus codes is impeded.
This manipulation of the stimulus presentation mode additionally allowed us to explore the extent to which different effectors might constitute separate unidimensional learning modules in the sense of Keele et al.’s (2003) theory of sequence learning. When there is only a single stimulus there is virtually no basis for the establishment of separate stimulus-related learning modules. Consequently, any independent learning of the two sequences would have to be ascribed to the operation of separate learning modules for the left and the right hand.
The method we employed required simultaneous responding with the left and the right hand to corresponding stimuli. In Experiment 1 and in the dual-stimulus condition of Experiment 2, each of two stimuli specified the response for one hand. In the single-stimulus condition of Experiment 2, however, only one stimulus appeared on each trial with its position in a row and a column of a matrix specifying the left and the right hand response, respectively. The order of stimuli and of responses for both hands was determined by two uncorrelated repeating sequences. After training, either one or both of the sequences were replaced with a random sequence or a sequence practiced with one hand was to be performed with the other hand. Increases of RTs and error rates in these test blocks in comparison to adjacent regular blocks (RT costs and error costs, respectively) were used as indicators of sequence learning.
RT costs in test blocks with both sequences abolished were generally larger than in test blocks in which only one sequence was abolished. Moreover, response errors were mostly hand-related, that is, error costs were evident only for hands executing a pseudo-random sequence, but not for hands executing a practiced sequence. Both results suggest the acquisition of independent knowledge about the sequences of stimuli and responses of the left and the right hand. Although coordinated responding with both hands was required, participants seem to have acquired separate knowledge about the sequential repetitions in each hand. As such learning of hand-related sequences was also indicated in sub-samples with only fragmentary or negligible explicit sequence knowledge, the acquisition of hand-related sequences appears to be independent of sequence recognition.
In addition to that, the data revealed complete intermanual transfer of the acquired sequence knowledge when responses of both hands were triggered by distinct but identically arranged stimuli (Experiment 1). Reducing the distinctiveness of the hand-related stimulus sequences (dual-stimulus condition of Experiment 2), also reduced the amount of intermanual transfer. Finally, when there was only one stimulus for both sequences, intermanual transfer was no longer indicated. These findings suggest that under the present conditions the transferable sequence knowledge relies mainly on stimulus codes which are acquired in a hand-related manner but which nevertheless are available for intermanual transfer.
Some aspects of the reported data pattern could also have been due to learning of the compound bimanual sequence instead of separate learning of two hand-related sequences.Footnote 3 When, after learning, only the part of the compound sequence pertaining to one hand is randomized, and the learned part pertaining to the other hand remains correct, hand-related error costs as well as reduced RT costs compared to when the whole compound sequence is randomized would result even if only compound learning had occurred. Therefore, this pattern of findings does not unambiguously indicate independent learning of two hand-related sequences. We cannot rule out that at least parts of the compound sequence may have been learned but we doubt that compound learning provides a satisfactory account for all our findings.
First, it stands to reason that compound learning in Experiment 2 should have been easier and thus more pronounced in the single-stimulus condition than in the dual-stimulus condition, which was clearly not the case. Numerically, the difference was even in the opposite direction. Second, the transitions of the compound sequence are changed in equal measure both in transfer blocks (L trans and R trans) and in test block LR rand, so that the same costs should result. However, we found in Experiment 1 and in the dual-stimulus condition of Experiment 2 substantially greater RT costs in block LR rand than in transfer blocks. One may argue that the reduced costs in the transfer blocks are due to within-block learning. However, there were no reliable indications of within-block learning. Furthermore, within-block learning of a compound sequence should have reduced error costs for both hands whereas we found the error costs selectively reduced for only that hand which performed the transferred sequence. In sum, we argue that the data pattern reported here is more consistent with the independent sequence learning than with the compound learning account.
It remains to discuss the transfer issue: Experiment 2 provided data in support of hand-related sequence knowledge which could be transferred to the other hand only partly (dual-stimulus condition) or not at all (single-stimulus condition). This suggests sequence knowledge which is bound to the hand with which it has been acquired (cf. Verwey & Clegg, 2005; Verwey & Wright, 2004; see also Park & Shea, 2005).
Such hand-specific sequence knowledge may rely on the order in which the keys assigned to the respective hand are to be pressed (cf. Willingham et al., 2000), or it might rely on the order in which the fingers of the respective hand have to act. Under the present conditions the order of keys is equally transferable as the order of stimulus locations. Therefore, we consider the latter alternative to be more likely. The notion that after extensive practice our participants may have acquired hand-specific representations of the order in which the fingers are to be moved fits well with the speculation of Keele et al. (2003, p. 317) that “distinctions within the motor system (e.g., hands vs. feet) may also constitute dimensions”, that is, separate unidimensional learning modules.
The possibility that the execution of highly trained coordinated movements may rely partly on sequence knowledge which is stored ‘in’ the involved limbs opens interesting research perspectives. On the one hand, it is necessary to explore the conditions for the acquisition of such embodied sequence knowledge and to clarify its representational format. On the other hand, the existence of effector-specific sequence knowledge points to a hierarchy of movement control in which effector-independent and effector-specific mechanisms cooperate in a way we do not yet understand and which to elucidate seems to us a worthwhile endeavor.
Notes
We would like to thank Willem Verwey for pointing this out.
One reviewer correctly pointed out that the performance decrements in transfer blocks might not be due to a failure of transfer but rather to interference: first, for the transfer hand the formerly practiced sequence might interfere with the new transferred sequence; second, the transferred sequence might become suppressed because the hand which has practiced this sequence before now has to perform a random sequence. However, the interference account can hardly explain why in transfer blocks of Experiment 1 no interference at all is indicated. In particular, in Experiment 1 the error data suggested nearly complete transfer as the hands which performed the transferred sequence never produced error costs, whereas the hands which performed the random sequence always produced error costs. Therefore we consider the account in terms of transfer at least as plausible as the interference account.
We would like to thank an anonymous reviewer for alerting us to this alternative account.
References
Bapi, R. S., Doya, K., & Harner, A. M. (2000). Evidence for effector independent and dependent representations and their differential time course of acquisition during motor sequence learning. Experimental Brain Research, 132, 149–162.
Baddeley, A. D., & Hitch, G. J. (1974). Working memory. In G. Bower (Ed.), The psychology of learning and motivation (Vol. 8, pp. 47–90). San Diego, CA: Academic Press.
Clegg, B. A., DiGirolamo, G. J., & Keele, S. W. (1998). Sequence learning. Trends in Cognitive Sciences, 2, 275–281.
Cohen, A., Ivry, R. B., & Keele, S. W. (1990). Attention and structure in sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 16, 17–30.
Deroost, N., & Soetens, E. (2006). Perceptual or motor learning in SRT tasks with complex sequence structures. Psychological Research, 70, 88–102.
Destrebecqz, A., & Cleeremans, A. (2001). Can sequence learning be implicit? New evidence with the process dissociation procedure. Psychonomic Bulletin and Review, 8, 343–350.
Frensch, P. A., & Miner C. S. (1995). Zur Rolle des Arbeitsgedächtnisses beim impliziten Sequenzlernen (The role of working memory in implicit sequence learning). Zeitschrift für Experimentelle Psychologie, 42, 545–575.
Grafton, S. T., Hazeltine, E., & Ivry, R. B. (1998). Abstract and effector-specific representations of motor sequences identified with PET. Journal of Neuroscience, 18, 9420–9428.
Grafton, S. T., Hazeltine, E., & Ivry, R. B. (2002). Motor sequence learning with the nondominant left hand. A PET functional imaging study. Experimental Brain Research, 146, 369–378.
Hazeltine E. (2002). The representational nature of sequence learning: Evidence for goal-based codes. In W. Prinz, & B. Hommel (Eds.), Common mechanisms in perception and action (Attention and Performance XIX, pp. 673–689). New York, NY: Oxford University Press.
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002). Central mechanisms of motor skill learning. Current Opinion in Neurobiology, 12, 217–222.
Hoffmann, J. (2001). Representation of serial order, cognitive psychology of. In N. Smelser, & P. Baltes (Eds.), International encyclopedia of the social and behavioral sciences (Vol. 19, pp. 13171–13174). Oxford, UK: Elsevier.
Hoffmann, J., & Koch, I. (1997). Stimulus-response compatibility and sequential learning in the serial reaction time task. Psychological Research, 60, 87–97.
Hoffmann, J., & Koch, I. (1998). Implicit learning of loosely defined structures. In M. A. Stadler, & P. Frensch (Eds.), Handbook of implicit learning (pp. 161–199). Thousand Oaks, CA: Sage Publications.
Hoffmann, J., Martin, C., & Schilling, A. (2003). Unique transitions between stimuli and responses in SRT tasks: Evidence for the primacy of response predictions. Psychological Research, 67, 160–173.
Hoffmann, J., & Sebald, A. (1996). Reiz- und Reaktionsmuster in seriellen Wahlreaktionen [Stimulus and response patterns in serial reaction time tasks]. Zeitschrift für Experimentelle Psychologie, 43, 40–68.
Hoffmann, J., & Sebald, A. (2005). When obvious covariations are not even learned implicitly. European Journal of Cognitive Psychology, 17, 449–480.
Hoffmann, J., Sebald, A., & Stöcker, C. (2001). Irrelevant response effects improve serial learning in serial reaction time tasks. Journal of Experimental Psychology: Learning, Memory, and Cognition, 27, 470–482.
Howard, J. H., Mutter, S. A., & Howard, D. V. (1992). Serial pattern learning by event observation. Journal of Experimental Psychology: Learning, Memory, and Cognition, 18, 1029–1039.
Jiménez, L., & Méndez, C. (1999). Which attention is needed for implicit sequence learning? Journal of Experimental Psychology: Learning, Memory, and Cognition, 25, 236–259.
Keele, S. W., Ivry, R. B., Mayr, U., Hazeltine, E., & Heuer, H. (2003). The cognitive and neural architecture of sequence representation. Psychological Review, 110, 316–339.
Keele, S. W., Jennings, P., Jones, S., Caulton, D., & Cohen, A. (1995). On the modularity of sequence representation. Journal of Motor Behavior, 27, 17–30.
Koch, I., & Hoffmann, J. (2000). The role of stimulus-based and response-based spatial information in sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26, 863–882.
Mayr, U. (1996). Spatial attention and implicit sequence learning: Evidence for independent learning of spatial and nonspatial sequences. Journal of Experimental Psychology: Learning, Memory, and Cognition, 22, 350–364.
Nattkemper, D., & Prinz, W. (1997). Stimulus and response anticipation in a serial reaction task. Psychological Research, 60, 98–112.
Nissen, M. J., & Bullemer, P. (1987). Attentional requirements of learning: Evidence from performance measures. Cognitive Psychology, 19, 1–32.
Park, J.-H., & Shea, C. H. (2005). Sequence learning: Response structure and effector transfer. Quarterly Journal of Experimental Psychology, 58A, 387–419.
Remillard, G. (2003). Pure perceptual-based sequence learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 29, 581–597.
Rhodes, B. J., Bullock, D., Verwey, W. B., Averbeck, B. B., & Page, M. P. A. (2004). Learning and production of movement sequences: Behavioral, neurophysiological, and modelling perspectives. Human Movement Science, 23, 699–746.
Riedel, B., & Burton, A. M. (2006). Auditory sequence learning: Differential sensitivity to task relevant and task irrelevant sequences. Psychological Research, 70, 337–344.
Rüsseler, J., Münte, T. F., & Rösler, F. (2002). Influence of stimulus distance in implicit learning of spatial and nonspatial event sequences. Perceptual and Motor Skills, 95, 973–987.
Rüsseler, J., & Rösler, F. (2000). Implicit and explicit learning of event sequences: Evidence for distinct coding of perceptual and motor representations. Acta Psychologica, 104, 45–67.
Schmidtke, V., & Heuer, H. (1997). Task integration as a factor in secondary-task effects on sequence learning. Psychological Research, 60, 53–71.
Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-Prime, version 1.1. Pittsburgh, PA: Psychology Software Tools.
Stöcker, C., & Hoffmann, J. (2004). The ideomotor principle and motor sequence acquisition: Tone effects facilitate movement chunking. Psychological Research, 68, 126–137.
Stöcker, C., Sebald, A., & Hoffmann, J. (2003). The influence of response-effect compatibility in a serial reaction time task. The Quarterly Journal of Experimental Psychology, 56A, 586–703.
van der Graaf, F. H. C. E., de Jong, B. M., Maguire, R. P., Meiners, L. C., & Leenders, K. L. (2004). Cerebral activation related to skills practice in a double serial reaction time task: Striatal involvement in random-order sequence learning. Cognitive Brain Research, 20, 120–131.
Verwey, W. B., & Clegg, B. A. (2005). Effector dependent sequence learning in the serial RT task. Psychological Research, 69, 242–251.
Verwey, W. B., & Wright, D. L. (2004). Effector-independent and effector-dependent learning in the discrete sequence production task. Psychological Research, 68, 64–70.
Willingham, D. B., Wells, L. A., Farrell, J. M., & Stemwedel, M. E. (2000). Implicit motor sequence learning is represented in response locations. Memory & Cognition, 28, 366–375.
Ziessler, M. (1998). Response-effect learning as a major component of implicit serial learning. Journal of Experimental Psychology: Learning, Memory, and Cognition, 24, 962–978.
Ziessler, M., & Nattkemper, D. (2001). Learning of event sequences is based on response-effect learning: Further evidence from a serial reaction task. Journal of Experimental Psychology: Learning, Memory, and Cognition, 27, 595–613.
Acknowledgments
This research was supported by grant HO 1301/12-1 awarded to J. Hoffmann by the German Research Council (DFG). Parts of this research were presented at the XIVth Conference of the European Society for Cognitive Psychology (ESCoP), Leiden, The Netherlands, August 31–September 3, 2005. The authors would like to thank Herbert Heuer, Willem B. Verwey, and an anonymous reviewer for valuable comments on earlier versions of this article.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Berner, M.P., Hoffmann, J. Effector-related sequence learning in a bimanual-bisequential serial reaction time task. Psychological Research 72, 138–154 (2008). https://doi.org/10.1007/s00426-006-0097-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00426-006-0097-8