
Abstract
Origins of all hearing impairment forms may be divided into genetic mutations and acquired influence. Both carry damage to the inner ear structure resulting in a mild to profound dysfunction of the auditory system. The purpose of this study was to assess the different etiologies of deafness in two reference centers for hearing-impaired children in Nouakchott/Mauritania. Data on gender, age, consanguinity, etiology and family history of deafness were gathered by interviewing the custodians of 139 children with hearing loss. DNA of pupils with hereditary non-syndromic deafness was then screened for GJB2 mutations by sequencing methods. Postnatal hearing loss was found in 36 (25.8 %) out of the 139 children surveyed. The main etiologies of this group were infections caused by meningitis (12.9 %) and measles (2.8 %). Unknown and ototoxic origins accounted for, respectively, 5.7 and 3.5 %. In 103 (74.1 %) children, deafness was identified near after the time of birth and, therefore, presumed as congenital. 56.8 % of deaf children had consanguineous parents. Two GJB2 mutations, c.del35G with an allele frequency of 4.7 % and R32C (3.7 %) were detected. Infections such as meningitis and measles were the most prevalent causes of postnatal deafness. In cases of congenital hearing impairment, two GJB2 allele variants, i.e., del35G and R32C (3.7 %) were detected. Extended genetic testing is recommended for a more comprehensive determination of congenital causes.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
About 5 % of the global population has hearing impairment [1]. Although this disability is commonly regarded as an aging-related change, it affects more than 32 million children worldwide. Deafness is indeed one of the most known congenital disorders as it occurs in 1 in 1000 newborns [2]. By the age of four, this prevalence could rise up to 1 in 300 children following damage to the hearing system prompted by numerous postnatal events such as viral or bacterial infections, shock, medication or exposure to noise [3]. Both inborn and acquired postnatal forms of inner ear deafness are irreversible. Sensorineural hearing loss (SNHL) results from injury to the cochlea, the hair or nerve cells of the inner ear. Most of genetic SNHL are of autosomal recessive inheritance and associated with GJB2, a gene located in the DNFB1 locus and encoding the gap junction connexin 26, the major connexin isoform of the cochlea [4]. Allele variants of this gene, e.g., c.167delT and R143W are rare [5, 6] while others such as c.35delG, c.235delC and W24X have a wider distribution [7–9]. c.35delG is a frameshift-inducing deletion which results in an early termination at the 12th residue of the polypeptide chain. On its own, this change accounts for two-thirds of all GJB2 alterations in European and American Caucasian populations [10, 11]. In North Africa, studies showed that, with prevalence up to 37 %, c.35delG is the most common GJB2 mutation in Tunisian and Moroccan deaf populations [12, 13].
The Mauritanian population takes its name from the Berber kingdom of Mauritania which existed from the third century BC until the 7th AD in what is called now Morocco. However, the present Mauritanian population is made of two racial categories: the Moor (Maures) of Arab–Berber origin composed of white and black Moor both speaking Hassaniya, a Berber-influenced Arabic dialect. This group self-identifies with the geographically close populations of North Africa as they speak related languages and share the same Arab-Islamic culture. The Mauritanian black African group is composed of three ethnicities (Pulhar, Soninké and Wolofs) all from the same sub-Saharan black African descent but each with its own language, geographic origin and cultural specificities. Although these two classes (the Moor and black Africans) are racially different, they share the same Islam religion and lived together for centuries. This cohabitation allowed not only cultural exchange but also interracial marriage, which greatly contributed in shaping a mixed society with specific genetic features.
This study was aimed to provide the etiology profile of hearing impairment in a cohort of Mauritanian deaf children. A GJB2 screening carried out in patients with congenital deafness was also the first attempt to evaluate the genotype–phenotype correlation of hearing loss in our population.
Materials and methods
Patients selection
Approval to this study was given by the boards of the two schools where the screening was conducted and the ethics committee of the University of Sciences, Technologies and Medicine, Nouakchott, Mauritania. Informed consent was obtained from children’s parents or guardians who were then interviewed to complete the study questionnaire. The 139 deaf children (from 113 families) included in this study were from the two race categories: 119 Maures and 20 black Africans.
Child identity was taken and anonymized. Data included the period of identification of hearing loss, etiology and degree of the deafness, associated symptoms which may be characterized as part of a known syndrome, consanguinity and hearing loss history in the family. All etiologies stated without medical assessment were considered as unknown. Congenital cases were referred for genetic testing. In families with multiple affected siblings, one child per family was chosen randomly for GJB2 screening. Although all custodians of the 101 children (75 families) with congenital deafness recruited accepted to complete the questionnaire, consent for genetic testing was agreed by only 53 families and, therefore, concerned 53 unrelated pupils.
Molecular analysis
Genomic DNA was extracted by salting out methods [14] from blood samples collected in ETDA tubes. Target regions in GJB2 exon2 were amplified using, respectively, the primer sets Con26F(5′GAGGTTGTGTAAGAGTTGGTGTT3′)/Con26R (5′TCTTCTCATGTCTCCGGTAG3′) for the upstream region (457 bp) of the exon and the set GJB2-2F (5′CCAGGCTGCAAGAACGTGT3′)/GJB2-2R (5′ACAGCTGAGCACGGGTTG3′) for the subsequent part (597 bp). PCRs (25 µl) contained 3 µl (20 ng) of genomic DNA, 1 µl (5 µM) of each primer, 0.25 µl (20 mM) of dNTP, 0.75 µl (50 mM) of MgCl2, 0.1 µl of Invitrogen Taq DNA polymerase 5 units/µl, 2.5 µl of 10× PCR buffer and 17.4 µl distilled H2O. The amplification program presented a DNA denaturation at 96 °C for 4 min followed by 35 cycles of denaturation at 94 °C for 30 s, annealing at 53 °C for 35 s and extension at 72 °C for 40 s tailed by a 10-min final extension at 72 °C. PCR products were sequenced on capillary AB 3130 Genetic Analyzer (Applied Biosystems, California, USA). Data obtained were then matched with reference sequences of the GJB2 gene (NM_004004.5) using the Sequencher 5.0 software program package (Gene Codes, MI, and USA).
Results
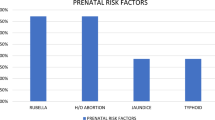
The 139 deaf children (from 113 families) surveyed in this study included 78 boys (56 %) and 61 girls (44 %) with an age between 5 and 25 years. Reports examination showed that in 101 pupils (75 families) deafness was noticed in the first weeks after birth and considered as congenital (72.6 %), while in 36 children (36 families), the disability occurred in the early childhood and following a health-affecting event (25.8 %) (Table 1). In two children, the period of onset could not be obtained. Postnatal deafness was mainly attributed to complication of infectious diseases such as meningitis (12.9 %) which was the most common reported cause followed by measles (2.9 %). Ototoxic drugs and exposure to loud noise were also stated as cause of hearing loss. In eight children (5.7 %), it was not possible to identify the etiology (Table 2).
In the 101 children with congenital hearing loss, 66 were born from consanguineous unions (46 families) against 35 children from non-consanguineous unions (29 families). Multiple occurrence of hearing loss was encountered in 54.6 % (25/46) of consanguineous families against only 27.5 % (8/29) in non-consanguineous families (Fig. 1).

Effect of consanguinity on family history of deafness
GJB2 sequence data of children chosen from the 53 families with congenital SNHL revealed two mutations c.del35G and R32C, present, respectively, in 3 and 2 families, all from the Maure group. No second associated mutation was identified. Both mutations were detected in children born from consanguineous unions. The variant c.del35G had an allele frequency of 4.71 % (two homozygotes and one heterozygote) while R32C was in homozygote state in the two individuals identified giving an allele frequency of 3.77 %. Polymorphism rs9578260 was detected in heterozygote state in three families (Table 3). Overall, eight families (15.1 %) were carrying at least one mutant Cx26 allele.
Discussion
Early identification of the etiology is of great importance in restoring the hearing ability. In this study, the first conducted in Mauritania to assess the main causes of childhood hearing loss, we found that among children who became deaf after birth, the impairment resulted largely from complication of infections such as measles and meningitis. The latter accounted for 13 % of all causes of deafness in this study. Meningitis remains one of the most common reasons of deafness worldwide, although its prevalence varied among populations, mostly through immunization, [15, 16]. This avoiding effect is underlined by the wide difference between the prevalence of the disease as diagnosed etiology of deafness in the US [17] (3.2 %) and the Gambia [18] (31.7 %).
The almost fivefold difference in the prevalence of meningitis and measles as cause of deafness (Table 2) is also more likely the result of immunization as unlike measles, meningitis is yet to be added to the national children vaccination program. Our data emphasize the adverse effect of this illness and the urgent need to include it in this program. Deafness cases consecutive to measles could be explained by vaccine supply conditions, a generic cause of loss of protection in developing countries or by inherent factors in children linked to socio economic conditions.
Hearing loss caused by ototoxic medicines represented 13 %. This prevalence took into account only children in whom the damage to the hearing was not reversed when they stopped taking their medicine. It is slightly higher than the 8 % reported in sub-Saharan populations [19]. A close monitoring for pediatric patients receiving known ototoxic agents is recommended.
In the unknown etiology group, we classified the impairments which cause could not be identified through the questionnaire. A likely reason, as in many developing countries, may be that the etiology was not properly diagnosed or the parents could not reach medical care for various reasons.
In this study, all hearing loss cases reported by the parents in the first weeks of childhood and not associated to a diagnosed postnatal illness were considered as congenital. Acquired deafness resulting from infections or traumatic events during fetal development or at the time of birth is well documented [20]. Therefore, the 101 (72.6 %) congenital hearing loss cases identified in 75 families may not be all of genetic origin. However, 33 families of them had history of deafness giving a minimum prevalence of 44 % for hereditary deafness. Deafness in multiple family members doubled from 27.5 % in non-consanguineous families to 54.3 % in consanguineous families. By increasing the probability of defective gene sharing and, therefore, the level of homozygote, consanguineous union was known to increase the risk of autosomal recessive disorders such as SNHL in different populations [21]. Although considered of low influence in western populations [22, 23], the consanguinity impact was more noticeable in developing countries [24, 25]. For instance, 70 % of deaf children in Oman had consanguineous parents [26]. A lower rate of consanguinity (56.8 %) was found here. Even though not very representative given the design of the study, this percentage found among the deaf population may be used as indication of the prevalence of consanguinity in the general population before a proper study is conducted.
As stated above, genetic origin represented at least 44 % of all cases of congenital deafness. In the 53 DNA samples referred for GJB2 variant search, c.del35G and R32C mutations were almost equally prevalent. Although c.del35G is reported as the most prevalent GJB2 mutant allele in North America, Europe, Asia and North Africa populations [27], this variant was detected here only among 3 of the 53 families with supposed congenital deafness. c.delG35 was also not found in cohorts of deaf children from many black African countries including South Africa [28], Cameroon [29], Kenya and Sudan [30]. The intermediary prevalence found in our study between a high frequency of about 35 % in North Africa [12, 13] and a total absence of this mutation in the sub-Saharan deaf populations supports the previously reported specific heterogeneity of our population using others biomarkers [31, 32] and briefly reviewed above. Indeed, although Mauritania is geographically and culturally close to the North African region, there is an important genetic setting particular to our population induced by the same geographic nearness of our country to the sub-Saharan populations.
Genetic variation in GJB2 gene was also encountered even among the others North African populations. The prevalence of c.35delG mutation in Moroccan deaf population (43.20 %) [13] is double of that found in Tunisia (21.57 %) [33]. The carrier frequencies are also different with, respectively, 2.07 % in Morocco and 1.3 % in Tunisia [34, 35]. Mutations in LRTOMT gene, another hearing loss candidate gene, have shown also difference in prevalence between the two populations [36].
A contribution to the relatively low prevalence of c.delG35 in our population with regard to those described in North African populations may also be of statistical source. Indeed, as said above, only 33 families from the 75 with supposed congenial hearing loss had history family of the disorder. As a result, the prevalence of all DNA changes found in this study, including c.delG35, would be higher if brought to 33 families instead of 53 as the total of families with congenital deafness.
R32C substitution (c.94C>T) was so far identified only as a novel and rare mutation [37]. The relatively high frequency found here strengthens the pathogenic significance of R32C as a hearing loss-associated variant [38] although arguably a case–control study may still need to be carried out to confirm this relevance in our population. To our knowledge, it is the first time that this mutation is encountered in an African population. As the descent of the R32C carrier was not given in the only study reporting this variant [37], we could not speculate further on its ethnic derivation. SNP c.-22-12C>T (rs9578260) found here in 3 out of the 53 families tested was described also in heterozygote state and with a benign consequence on hearing loss in other African populations [3].
Although these DNA changes have already been described in other populations, the possibility to detect hearing loss-associated mutations and others inherited diseases through single-gene screening will allow our population to benefit from this genetic test and, in due time, provide the opportunity to explore multi-gene screening which became important in the diagnosis and evaluation of hereditary disorders. The identification of these mutations may also contribute in understanding and characterizing the genetic variations of our population composed of several groups racially and ethnically distinct.
Mutations in GJB2 were detected in only 5 out of the 53 investigated families. This leaves 90 % of putative genetic causes not detected and more likely located in others candidate genes. As a result, a screening involving a wider gene panel will certainly unravel other hearing loss-associated allele variants. A high prevalence of LRTOMT gene mutations was indeed reported in the Moroccan deaf population, another North African Arab–Berber population geographically and ethnically close to our population [36].
In this study, deafness-associated mutations were searched only in DNA of patients with congenital deafness. We did not genetically test the postnatal cases even though the occurrence of such mutations could not be excluded, namely in case of sporadic mutation denoting a first appearance of deafness in the family.
An important point to be considered is the representativeness, both ethnic and numeric, of the cohort screened here. As mentioned above, pupils screened were from the different ethnic groups of the Mauritanian population. However, due to the limited number of screened children, the percentages found here could only be considered as indication on the prevalence of hearing loss and associated mutations in our population. The difficulty in patient’s recruitment, inherent to this type of study, may also explain why most of similar screening carried out in our region [13, 33, 39] and worldwide [4] had a ratio screened cohort: deaf population or general population of comparable order to that used here.
Conclusions
Among the different etiologies, this study unveiled that preventable infections play a major part as incurring factor of deafness. The GJB2 gene mutations detected in this screening were already associated with NSHL in various studies and support the introduction of this gene in deafness diagnosis protocol. Although carried out on a limited population, this study showed that, giving the low prevalence of GJB2 mutations in the hearing-impaired children tested, the genetic screening could be optimized by including others gene candidates such as LRTOMT also described in our region.
References
Fact sheet No. 300 on deafness and hearing loss [WHO Web site]. http://www.who.int/mediacentre/factsheets/fs300/en/. Accessed May 13 2015
Morton CC, Nance WE (2006) Newborn hearing screening—a silent revolution. Engl J Med 18:2151–2164
Javidnia H, Carson N, Awubwa M, Byaruhanga R, Mack D, Vaccani JP (2014) Connexin gene mutations among Ugandan patients with nonsyndromic sensorineural hearing loss. Laryngoscope 124:373–376
Chan DK, Chang KW (2014) GJB2-associated hearing loss: systematic review of worldwide prevalence, genotype, and auditory phenotype. Laryngoscope 124:34–53
Zelante L, Gasparini P, Estivill X, Melchionda S, D’Agruma L, Govea N et al (1997) Connexin26 mutations associated with the most common form of non-syndromic neurosensory autosomal recessive deafness (DFNB1) in Mediterraneans. Hum Mol Genet 6:1605–1609
Brobby GW, Muller-Myhsok B, Horstmann RD (1997) Connexin 26 R143W mutation associated with recessive nonsyndromic sensorineural deafness in Africa. N Engl J Med 19:548–550 (no abstract available)
Denoyelle F, Weil D, Maw MA, Wilcox SA, Lench NJ, Allen-Powell DR et al (1997) Prelingual deafness: high prevalence of a 30delG mutation in the connexin 26 gene. Hum Mol Genet 6:2173–2177
Fuse Y, Doi K, Hasegawa T, Sugii A, Hibino H, Kubo T (1999) Three novel connexin26 gene mutations in autosomal recessive non-syndromic deafness. Neuroreport 10:1853–1857
Kelley PM, Harris DJ, Comer BC, Askew JW, Fowler T, Smith SD et al (1998) Novel mutations in the connexin 26 gene (GJB2) that cause autosomal recessive (DFNB1) hearing loss. Am J Hum Genet 62:792–799
Gasparini P, Rabionet R, Barbujani G, Melçhionda S, Petersen M, Brøndum-Nielsen K et al (2000) High carrier frequency of the 35delG deafness mutation in European populations. Genetic Analysis Consortium of GJB2 35delG. Eur J Hum Genet 8:19–23
Green GE, Scott DA, McDonald JM, Woodworth GG, Sheffield VC, Smith RJ (1999) Carrier rates in the midwestern US for GJB2 mutations causing inherited deafness. JAMA 16:2211–2216
Riahi Z, Hammami H, Ouragini H, Messai H, Zainine R, Bouyacoub Y et al (2013) Update of the spectrum of GJB2 gene mutations in Tunisian families with autosomal recessive nonsyndromic hearing loss. Gene 525:1–4
Abidi O, Boulouiz R, Nahili H, Ridal M, Alami MN, Tlili A et al (2007) GJB2 (connexin 26) gene mutations in Moroccan patients with autosomal recessive non-syndromic hearing loss and carrier frequency of the common GJB2-35delG mutation. Int J Pediatr Otorhinolaryngol 71:1239–1245
Miller SA, Dykes DD, Polesky HF (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic Acids Res 16:1215
Derekoy FS (2000) Etiology of deafness in Afyon school for the deaf in Turkey. Int J Pediatr Otorhinolaryngol 55:125–131
da Silva LP, Queiros F, Lima I (2006) Etiology of hearing impairment in children and adolescents of a reference center APADA in the city of Salvador, state of Bahia. Rev Bras Otorhinolaringol 72:33–36
Meningitis and hearing loss [Beltone Web site]. http://www.beltone.com/hearing-health/meningitis-and-hearing-loss.aspx. Accessed May 14 2015
Mc Pherson B, Holborrow CA (1985) A study of deafness in West Africa: the Gambian hearing Project. Int J Pediatr Otorhinolaryngol 10:115–135
Wonkam A, Noubiap JJ, Djomou F, Fieggen K, Njock R, Toure GB (2013) Aetiology of childhood hearing loss in Cameroon (sub-SaharanAfrica). Eur J Med Genet 56:20–25
Haddad J Jr (2011) Hearing loss. In: Kliegman RM, Behrman RE, Jenson HB, Stanton BF (eds) Nelson textbook of pediatrics, 19th edn. Elsevier, Philadelphia, p 629
Smith RG, Vancamp G (2013) Deafness and hereditary hearing loss. Gene reviews (2007). http://www.cridi.com.br/testedesurdez1.pdf
Cone-Wesson B, Vohr BR, Sininger YS, Widen JE, Folsom RC, Gorga MP (2000) Identification of neonatal hearing impairment: infants with hearing loss. Ear Hear 21:488–507
Van Riper LA, Kileny PR (1999) ABR hearing screening for high-risk infants. Am J Otol 20:516–521
Zakzouk S (2002) Consanguinity and hearing impairment in developing countries: a custom to be discouraged. J Laryngol Otol 116:811–816
Tadmouri GO, Nair P, Obeid T, Al Ali MT, Al Khaja N, Hamamy HA (2009) Consanguinity and reproductive health among Arabs. Reprod 8(6):17. doi:10.1186/1742-4755-6-17
Khabori MA, Patton MA (2008) Consanguinity and deafness in Omani children. Int J Audiol 47:30–33
Snoeckx RL, Huygen PL, Feldmann D, Marlin S, Denoyelle F, Waligora J et al (2005) GJB2 mutations and degree of hearing loss: a multicenter study. Am J Hum Genet 77:945–957
Kabahuma RI, Ouyang X, Du LL, Yan D, Hutchin T, Ramsay M et al (2011) Absence of GJB2 gene mutations, the GJB6 deletion (GJB6-D13S1830) and four common mitochondrial mutations in nonsyndromic genetic hearing loss in a South African population. Int J Pediatr Otorhinolaryngol 75:611–617
Trotta L, Iacona E, Primignani P, Castorina P, Radaelli C, Del Bo L et al (2011) GJB2 and MTRNR1 contributions in children with hearing impairment from Northern Cameroon. Int J Audiol 50:133–138
Gasmelseed NM, Schmidt M, Magzoub MM, Macharia M, Elmustafa OM, Ototo B et al (2004) Low frequency of deafness-associated GJB2 variants in Kenya and Sudan and novel GJB2 variants. Hum Mutat 23:206–207
Hamed CT, Bollahi MA, Abdelhamid I, Med Mahmoud MA, Ba B, Ghaber S et al (2012) Frequencies and ethnic distribution of ABO and Rh(D) blood groups in Mauritania: results of first nationwide study. Int J Immunogenet 39:151–154
Veten FM, Abdelhamid IO, Meiloud GM, Ghaber SM, Salem ML, Abbes S et al (2012) Hb S [p6(A3)Glu→Val, GAG>GTG] and β-globin gene cluster haplotype distribution in Mauritania. Hemoglobin 36:311–315
Belguith H, Tlili A, Dhouib H, Ben Rebeh I, Lahmar I, Charfeddine I, Driss N, Ghorbel A, Ayadi H, Masmoudi S (2009) Mutation in gap and tight junctions in patients with non-syndromic hearing loss. Biochem Biophys Res Commun 385(1):1–5
Abidi O, Boulouiz R, Nahili H, Bakhouch K, Wakrim L, Rouba H, Chafik A, Hassar M, Barakat A (2008) Carrier frequencies of mutations/polymorphisms in the connexin 26 gene (GJB2) in the Moroccan population. Genet Test 12(4):569–574
Masmoudi S, Elgaied-Boulila A, Kassab I, Ben Arab S, Blanchard S, Bouzouita JE, Drira M et al (2000) 2000 Determination of the frequency of connexin26 mutations in inherited sensorineural deafness and carrier rates in the Tunisian population using DGGE. J Med Genet 37(11):E39
Charif M, Bounaceur S, Abidi O, Nahili H, Rouba H, Kandil M, Boulouiz R, Barakat A (2012) The c.242G>A mutation in LRTOMT gene is responsible for a high prevalence of deafness in the Moroccan population. Mol Biol Rep 39:11011–11016
Prasad S, Cucci RA, Green GE, Smith RJ (2000) Genetic testing for hereditary hearing loss: connexin 26 (GJB2) allele variants and two novel deafness-causing mutations (R32C and 645-648delTAGA). Hum Mutat 16:502–508
Rabionet R, Gasparini P, Estivill X (2001). http://www.iro.es/deafness/
Trabelsi M, Bahri W, Habibi M, Zainine R, Maazoul F, Ghazi B, Chaabouni H (2013) Mrad R GJB2 and GJB6 screening in Tunisian patients with autosomal recessive deafness. Int J Pediatr Otorhinolaryngol 77(5):714–716. doi:10.1016/j.ijporl.2013.01.024 (Epub 2013 Feb 19)
Acknowledgments
The authors wish to thank the boards of the schools for deaf children in Nouakchott, in particular Mr Habib and Mrs Aminetou for their kind and valuable support in the observational study.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors do not have financial disclosure to report and declare no conflict of interest. The research was carried within the Université des Sciences de Technologies et de médecine (USTM), Nouakchott, Mauritanie. No extra funding to state. The authors declare that they have no conflict of interest.
Research involving human participants and/or animals
All procedures performed in studies involving human participants were in accordance with the ethical standards of the Université des Sciences de Technologies et de médecine (USTM), Nouakchott, Mauritania national ethics committee to whom all aspects of the study were explained. This article does not contain any studies with animals performed by any of the authors.
Informed consent
Informed consent was obtained from all legal guardians of individual participants included in the study.
Rights and permissions
About this article

Cite this article
Moctar, E.C.M., Riahi, Z., El Hachmi, H. et al. Etiology and associated GJB2 mutations in Mauritanian children with non-syndromic hearing loss. Eur Arch Otorhinolaryngol 273, 3693–3698 (2016). https://doi.org/10.1007/s00405-016-4036-z
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00405-016-4036-z