
Abstract
Improved repigmentation of generalized vitiligo in skin types IV–VI has been reported in clinical response to combined therapy with apremilast and narrowband (NB)-UVB; however, tissue responses to combined therapy versus NB-UVB monotherapy have not been elucidated. We compared the change from baseline in cellular and molecular markers in vitiligo skin after combined therapy versus NB-UVB monotherapy. We assessed lesional and nonlesional skin samples from enrolled subjects and evaluated for immune infiltrates, inflammatory, and melanogenesis-related markers which were compared across different treatment groups. Combined therapy resulted in significant reduction of CD8+T cells and CD11c+ dendritic cells, downregulation of PDE4B and Th17-related markers, and upregulation of melanogenesis markers. This study was limited to small sample size, skin types IV–VI, and high dropout rate. Our molecular findings support the clinical analysis that apremilast may potentiate NB-UVB in repigmentation of generalized vitiligo in skin types IV–VI.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Vitiligo is a disorder affecting pigmentation, acquired by 0.5%–1% of the global population [1]. It is often associated with other autoimmune conditions with T helper-17 (Th17) dysregulation such as psoriasis and alopecia areata, but can also occur in the setting of drug or vaccination-induced adverse events [2,3,4,5]. Several studies have reported the role of T helper-1 (Th1) and Th17 immune axes in generalized vitiligo [6,7,8,9,10], and the role of interferon gamma (IFNγ) pathway [11, 12]. IFNγ signaling stimulates keratinocytes and melanocytes to produce chemokines, including CXCL9 and CXCL10, which induce initial apoptosis of melanocytes and promote CD8 + T-cell recruitment to the epidermis [11, 13, 14]. Also, innate immune cells have been identified as the source of IFNγ, suggesting a key role in early stages of vitiligo [15]. Circadian clock dysfunction may also play a role as it has been linked to dysregulation of immune cells and Th17 cell differentiation [16, 17]. Currently used therapies for vitiligo include topical and systemic corticosteroids, topical immunomodulators, phototherapy and 308-nm excimer laser [18]. Narrowband UVB (NB-UVB) is considered the preferred first-line treatment of choice for generalized vitiligo affecting more than 10–20% of the body surface area (BSA) [19,20,21,22]. Recently, Janus Kinase (JAK) inhibitors are being explored as treatment option for vitiligo, since they downregulate IFNγ signaling [23,24,25]. However, there are long-term safety concerns with broad-acting JAK inhibitors and more selective JAK target for vitiligo still needs to be investigated [23].
Apremilast is a small oral molecule that inhibits phosphodiesterase-4 (PDE4), which has been shown to be effective in the treatment of psoriasis, psoriatic arthritis, and Behçet disease [26,27,28,29]. PDE4 inhibition activates the cyclic adenosine monophosphate (cAMP) pathway, which leads to downregulation of inflammatory mediators such as tumor necrosis factor (TNF)-α, IFN-γ, CXCL9, CXCL10, interleukin (IL)-2, IL-12, IL-17 and IL-23, and upregulation of anti-inflammatory cytokines such as IL-10 [26, 27, 30]. It also modulates innate immunity and reduces infiltration of myeloid dendritic cells, T cells, and NK cells into the epidermis and dermis [27, 31, 32]. Moreover, upregulation of cAMP pathway in melanocytes has been shown to activate melanocyte inducing transcription factor (MITF) and promote skin pigment production [33, 34]. PDE4 inhibition and subsequent cAMP upregulation suppress key mediators involved in the Th17 pathway, IFNγ pathway, and innate immune system, all of which have been reported to play a role in the pathophysiology of vitiligo.
Several studies have suggested beneficial clinical effects of apremilast in vitiligo [35, 36], while a recent study showed that apremilast does not bring any additional benefit to NB-UVB [13]. That study, however, lacked the advantage of a bilateral comparison study design which is optimal of vitiligo and tissue analysis [13]. We performed a randomized, split-body, pilot study to investigate the clinical and mechanistic response to combined therapy with apremilast and NB-UVB compared to NB-UVB monotherapy in SPTs IV-VI with generalized vitiligo. An analysis on clinical response has been reported in a separate publication, which suggested expedited clinical improvement with combined therapy [37]. In this mechanistic study, we investigated lesional and nonlesional skin biopsy specimens from subjects’ body sides before and after treatment with combined therapy versus NB-UVB monotherapy followed by apremilast monotherapy.
Materials and methods
Skin biopsy specimens were obtained from SPTs IV–VI with generalized vitiligo in a randomized, split-body, single-center, pilot study (NCT03123016) evaluating clinical and mechanistic effect of combined therapy with apremilast and NB-UVB versus NB-UVB monotherapy followed by apremilast monotherapy [37]. Each eligible subject’s right side of the body was randomized 1:1 to either Regimen A or B. Complement Regimen was assigned to each subject’s left side of the body. The body side assigned to Regimen A received NB-UVB monotherapy during Phase 1 (weeks 0–16), then was concealed from NB-UVB with a half-sided skin covering while exposed to oral apremilast monotherapy during Phase 2 (weeks 16–32). The body side assigned to Regimen B was untreated and occluded from NB-UVB with a half-sided skin covering during Phase 1, but received combined therapy with oral apremilast and NB-UVB during Phase 2. In Phase 3 (weeks 32–48), all subjects received combined therapy to their whole body. The scheme of the study has been presented in separate publication [37].
Lesional and nonlesional skin biopsies were obtained before treatment at baseline/week 0, and only lesional biopsies were obtained after 16, 32, and 48 weeks of treatment as above (Fig. 1). Lesional biopsies were taken from the edges of the active border of vitiligo skin and nonlesional biopsies were taken from adjacent normally pigmented skin. The expression levels of the markers of different immune and melanogenesis pathways were evaluated and compared among different treatment arms. Skin infiltration by T cells, dendritic cells (DCs), and Langerhans cells was assessed by immunohistochemistry.

Biopsy Collection Diagram. Skin samples were obtained at: Baseline (lesional skin from the active edge of vitiligo [Side X] and non-lesional—normally pigmented skin [Side X]); Week 16 (lesional skin in the same proximity to the prior biopsy but at least 1 cm away from scar [Side X]); Week 32 (lesional skin in the same proximity to the prior biopsy but at least 1 cm away from scar [Side X], and lesional skin from the contralateral side [Side Y]); and Week 48 or Early Termination visit (lesional skin in the same proximity of the prior biopsy from Side Y at Week 32 but at least 1 cm away from the scar). All skin biopsies were 4.5 mm in diameter
Immunohistochemistry
Immunohistochemistry (IHC) staining was performed on frozen OCT-embedded cryostat tissue sections of lesional and nonlesional biopsy specimens using purified mouse anti-human mAbs, as previously described (Table 1) [38, 39]. Human skin biopsies were dried and then fixed in acetone. The samples were blocked using 10% normal serum of the species the secondary antibody was made in. The samples were then incubated with primary antibody overnight in 4 °C. The primary antibodies’ signals were detected using biotin-labeled secondary antibodies (Vector Laboratories, Burlingame, CA). Chromogen 3-amino-9-ethylcarbazole (AEC, Sigma-Aldrich, Burlington, MA) was used to develop the staining intensity and signal. Cell counts were quantified using ImageJ V1.42 software (National Institutes of Health, Bethesda, MD).
Quantitative RT-PCR
RNA from skin biopsy specimens was extracted for real-time polymerase chain reaction (RT-PCR) using the miRNAeasy Mini Kit (Qiagen, Hilden, Germany). Reverse transcription to complementary DNA (cDNA) from RNA was carried out using the High Capacity cDNA reverse transcription (Thermo fisher). Pre-amplification was performed on all samples. Primers are listed in Table 2. 100 ng total RNA was used for PreAMP pool. Rplp0 was used as endogenous control. Expression values were normalized to Rplp0.
Statistical analyses
All analyses for immunohistochemistry and gene expression studies were performed in the statistical language R (www.R-Project.org). qRT-PCR values were normalized using RPLP0 housekeeping gene by transforming Ct values to -ΔCt. Then log2-scale qRT-PCR expression and IHC data were modeled by a linear mixed-effect model using nlme R framework.
Results
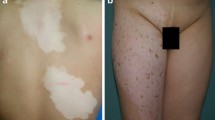
A total of 28 subjects were screened, and 23 subjects who met eligibility criteria were randomized 1:1 to receive NB-UVB to either the right side (n = 11) or the left side (n = 12) of the body in Phase 1. A total of 14 (60%) subjects completed visits through week 32. (Fig. 2). As recently published, clinical outcomes (probability of achieving grade 3 or 4 repigmentation, Vitiligo Area Scoring Index, and Body Surface Area scores) significantly improved after 16 weeks of combined therapy with apremilast and NB-UVB compared to NB-UVB monotherapy [37].

CONSORT flow diagram of study patients
We performed IHC to quantify changes in immune infiltrates across treatment groups and PCR analysis to assess differences in expression of inflammatory and melanogenesis-related markers. CD8+T cells and Langerin + Langerhans Cells (LCs) showed a significant reduction at week 48 compared to baseline in the body sides that received apremilast plus NB-UVB for 32 weeks during Phases 2 and 3. CD11c+ myeloid Dendritic Cells (DCs) achieved significant decrease in all groups treated with apremilast, with maximal reduction at week 48 in the body sides that received apremilast and NB-UVB for 32 weeks (p < 0.01; Fig. 3).

Representative immunostaining in skin biopsies from vitiligo patients. CD8+, Cd11c+ and Langerin+ immunostaining was performed on lesional (LES) and non-lesional (NON-LES) biopsies at baseline, and on LES biopsies collected from NB-UVB (Week 16), apremilast (Week 32), and NB-UVB plus apremilast (Weeks 32 and 48) arms. Cell count differences from baseline are depicted in the boxplots. Black stars, significance of comparison vs NB-UVB arm; red stars, significance of comparison versus baseline. ***P < 0.001, **P < 0.01, *P < 0.05, +P < 0.1
Our PCR analysis revealed a downregulation of PDE4B, a direct target of apremilast [40], with all treatment arms except for the body sides treated with 16 weeks of NB-UVB monotherapy (p < 0.05; Fig. 4). Th17-related markers such as PI3, S100A9, and IL23p19 showed significant reductions with all groups exposed to apremilast compared to NB-UVB monotherapy (p < 0.05). The greatest upregulation of melanogenesis/melanocyte markers, such as TYR, TRP1 and TRP2, the main melanogenic markers [38, 41], and MLANA/MART1, was observed in the body sides that received 32 weeks of combined therapy with apremilast and NB-UVB (p < 0.01). MITF and PAX3, which encode transcriptomic factors integral for both melanogenesis and melanocyte survival [39], and c-Kit, a tyrosinase kinase receptor involved in melanoblast expansion and migration,[42] were significantly upregulated only in the body sides that received combined therapy with apremilast and NB-UVB for 32 weeks (p < 0.01). The regulatory/fibrosis marker TGFβ that is central for Th17 differentiation in humans [10], showed a similar trend (p < 0.05).

Immune and melanocyte/melanogenesis biomarkers in lesional skin of patients with vitiligo. Log2 fold-changes (log2FCH) in lesional skin in NB-UVB, apremilast, and NB-UVB plus apremilast treatment arms compared to baseline. Black stars, significance of comparison between the NB-UVB monotherapy and treatment arms; red stars, significance of comparison versus baseline. ***P < 0.001, **P < 0.01, *P < 0.05, +P < 0.1
Discussion
Recent evidence suggests the role of Th17 axis, IFNγ pathway, and innate immune system in the pathophysiology of vitiligo [8, 9, 15, 26, 31, 32]. Apremilast is an oral PDE4 inhibitor with anti-inflammatory and immune modulating properties that targets key mediators involved in the pathogenesis of vitiligo [26, 30,31,32]. As reported in separate publication, analysis of clinical response to combined therapy with apremilast and NB-UVB versus NB-UVB monotherapy showed that apremilast may potentiate NB-UVB in repigmentation of generalized vitiligo in SPTs IV-VI [37]. In this study, we investigated molecular effects of combined therapy with apremilast and NB-UVB in vitiligo skin and correlated clinical disease improvement with histologic tissue response before and after treatment.
Combined therapy with apremilast and NB-UVB for 32 weeks resulted in significant histologic changes in vitiligo skin, compared to baseline or NB-UVB monotherapy followed by apremilast monotherapy. Compared to baseline, combined therapy progressively reduced immune infiltrates, including CD8+T cells, LCs, and CD11c+ myeloid DCs, in vitiligo skin at week 48. Inhibition of Th17-related markers (IL23p19, PI3, S100s) occurred with all groups exposed to apremilast, with strongest reduction after 32 weeks of combined therapy. Although increase in melanogenesis/melanocyte markers (TYR, TRP1, MLANA/MART1) was seen in all treatment arms, combined therapy arm showed the greatest upregulation at week 48. Some melanogenesis and melanoblast expansion markers (MITF, PAX3, c-KIT) were significantly upregulated only in the body sides that received combined therapy for 32 weeks, suggesting potential long-term benefit of combined therapy in repigmentation. Although histologic findings support clinical results that adding apremilast to NB-UVB enhances repigmentation, perhaps the duration of each phase of the study was not long enough to see maximal clinical and molecular effect of each treatment. Future longer studies are needed to evaluate for optimal treatment duration and long-term clinical and histologic responses to combination therapy.
In the previous analysis of clinical responses, progressive repigmentation was observed in the side of the body exposed to apremilast monotherapy from week 16 to week 32 after initial treatment with 16 weeks of NB-UVB monotherapy [37]. This was supported by tissue response which showed continuous reduction in CD8+ T-cell and CD11c+ DC infiltrates, downregulation of Th17 axis, and promotion of melanogenesis markers [26, 31]. However, it cannot be clearly demonstrated whether the progressive repigmentation was due to apremilast alone or residual efficacy from initial treatment with 16 weeks of NB-UVB monotherapy. More studies are needed to evaluate the potential role of apremilast monotherapy in the treatment of vitiligo.
Limitations of this study include small sample size and high dropout rate due to the demanding phototherapy schedule of multiple treatments per week as reported by patients. This suggests that the subjects who remained in the study may not be representative of general population and therefore, our results may not be generalizable. Also, only patients with dark skin phototypes IV–VI were included in the study to maximize chance of repigmentation since this group of patients are thought to be more responsive to treatment than light-skinned patients [43]. Furthermore, the methods used for tissue analysis such as immunohistochemistry and PCR for mRNA expression of inflammatory and melanogenesis markers may be insufficient to draw definitive conclusion. Future studies should assess related proteins by Western blot. Finally, we did not conduct a dose–response study specific for vitiligo. The dosage and frequency of apremilast was based on the data from psoriasis trials [44, 45.
In conclusion, our mechanistic findings support the clinical notion that apremilast may potentiate NB-UVB in repigmentation of generalized vitiligo in skin types IV–VI more than NB-UVB monotherapy followed by apremilast monotherapy, and suggest the potential role of combined therapy in progressive repigmentation of vitiligo and enhanced upregulation of melanogenesis markers in vitiligo skin. This trial serves as a clear proof-of-concept. We recommend further trials with larger sample sizes, longer treatment period, more comprehensive methods for tissue analysis, and investigation of apremilast monotherapy.
References
Taïeb A, Picardo M (2007) The definition and assessment of vitiligo: a consensus report of the Vitiligo European Task Force. Pigment Cell Res 20(1):27–35
Conic RZ, Tamashunas NL, Damiani G, Fabbrocini G, Cantelli M, Bergfeld WF (2020) Comorbidities in pediatric alopecia areata. J Eur Acad Dermatol Venereol 34(12):2898–2901
Malerba M, Damiani G, Radaeli A, Ragnoli B, Olivini A, Calzavara-Pinton PG (2015) Narrowband ultraviolet B phototherapy in psoriasis reduces proinflammatory cytokine levels and improves vitiligo and neutrophilic asthma. Br J Dermatol 173(6):1544–1545
Mineiro Dos Santos Garrett NF, Carvalho da Costa AC, Barros Ferreira E, Damiani G, Diniz Dos Reis PE, Inocêncio Vasques C. Prevalence of dermatological toxicities in patients with melanoma undergoing immunotherapy: Systematic review and meta-analysis. PLoS One. 2021;16(8):e0255716.
Ciccarese G, Drago F, Boldrin S, Pattaro M, Parodi A (2022) Sudden onset of vitiligo after COVID-19 vaccine. Dermatol Ther. 35(1):e15196
Kotobuki Y, Tanemura A, Yang L, Itoi S, Wataya-Kaneda M, Murota H et al (2012) Dysregulation of melanocyte function by Th17-related cytokines: significance of Th17 cell infiltration in autoimmune vitiligo vulgaris. Pigment Cell Melanoma Res 25(2):219–230
Antonelli A, Ferrari SM, Fallahi P (2015) The role of the Th1 chemokine CXCL10 in vitiligo. Ann Transl Med 3(Suppl 1):S16
Sushama S, Dixit N, Gautam RK, Arora P, Khurana A, Anubhuti A (2019) Cytokine profile (IL-2, IL-6, IL-17, IL-22, and TNF-α) in vitiligo-New insight into pathogenesis of disease. J Cosmet Dermatol 18(1):337–341
Wang CQ, Cruz-Inigo AE, Fuentes-Duculan J, Moussai D, Gulati N, Sullivan-Whalen M et al (2011) Th17 cells and activated dendritic cells are increased in vitiligo lesions. PLoS One. 6(4):e18907
Zhou L, Shi YL, Li K, Hamzavi I, Gao TW, Huggins RH et al (2015) Increased circulating Th17 cells and elevated serum levels of TGF-beta and IL-21 are correlated with human non-segmental vitiligo development. Pigment Cell Melanoma Res 28(3):324–329
Richmond JM, Bangari DS, Essien KI, Currimbhoy SD, Groom JR, Pandya AG et al (2017) Keratinocyte-derived chemokines orchestrate T-Cell positioning in the epidermis during vitiligo and may serve as biomarkers of disease. J Invest Dermatol 137(2):350–358
Harris JE, Harris TH, Weninger W, Wherry EJ, Hunter CA, Turka LA (2012) A mouse model of vitiligo with focused epidermal depigmentation requires IFN-γ for autoreactive CD8+ T-cell accumulation in the skin. J Invest Dermatol. 132(7):1869–76
Khemis A, Fontas E, Moulin S, Montaudié H, Lacour JP, Passeron T (2020) Apremilast in combination with narrowband UVB in the treatment of vitiligo: a 52-week monocentric prospective randomized placebo-controlled study. J Invest Dermatol 140(8):1533–7.e2
Wang XX, Wang QQ, Wu JQ, Jiang M, Chen L, Zhang CF et al (2016) Increased expression of CXCR3 and its ligands in patients with vitiligo and CXCL10 as a potential clinical marker for vitiligo. Br J Dermatol 174(6):1318–1326
Tulic MK, Cavazza E, Cheli Y, Jacquel A, Luci C, Cardot-Leccia N et al (2019) Innate lymphocyte-induced CXCR3B-mediated melanocyte apoptosis is a potential initiator of T-cell autoreactivity in vitiligo. Nat Commun 10(1):2178
Orihara K (2021) Skin and immune cells crosstalk via circadian regulations. Explor Immunol 1(4):285–294
Yu X, Rollins D, Ruhn KA, Stubblefield JJ, Green CB, Kashiwada M et al (2013) TH17 cell differentiation is regulated by the circadian clock. Science 342(6159):727–730
Guerra L, Dellambra E, Brescia S, Raskovic D (2010) Vitiligo: pathogenetic hypotheses and targets for current therapies. Curr Drug Metab 11(5):451–467
Njoo MD, Westerhof W, Bos JD, Bossuyt PM (1999) The development of guidelines for the treatment of vitiligo. Clinical epidemiology unit of the Istituto Dermopatico dell’Immacolata-Istituto di Recovero e Cura a Carattere Scientifico (IDI-IRCCS) and the archives of dermatology. Arch Dermatol. 135(12):1514–21
Kanwar AJ, Dogra S, Parsad D, Kumar B (2005) Narrow-band UVB for the treatment of vitiligo: an emerging effective and well-tolerated therapy. Int J Dermatol 44(1):57–60
Parsad D, Kanwar AJ, Kumar B (2006) Psoralen-ultraviolet A vs. narrow-band ultraviolet B phototherapy for the treatment of vitiligo. J Eur Acad Dermatol Venereol. 20(2):175–7
Li R, Qiao M, Wang X, Zhao X, Sun Q (2017) Effect of narrow band ultraviolet B phototherapy as monotherapy or combination therapy for vitiligo: a meta-analysis. Photodermatol Photoimmunol Photomed 33(1):22–31
Relke N, Gooderham M (2019) The use of janus kinase inhibitors in vitiligo: a review of the literature. J Cutan Med Surg 23(3):298–306
Liu LY, Strassner JP, Refat MA, Harris JE, King BA (2017) Repigmentation in vitiligo using the Janus kinase inhibitor tofacitinib may require concomitant light exposure. J Am Acad Dermatol 77(4):675–82.e1
Craiglow BG, King BA (2015) Tofacitinib citrate for the treatment of vitiligo: a pathogenesis-directed therapy. JAMA Dermatol 151(10):1110–1112
Schafer PH, Parton A, Gandhi AK, Capone L, Adams M, Wu L et al (2010) Apremilast, a cAMP phosphodiesterase-4 inhibitor, demonstrates anti-inflammatory activity in vitro and in a model of psoriasis. Br J Pharmacol 159(4):842–855
Gottlieb AB, Strober B, Krueger JG, Rohane P, Zeldis JB, Hu CC et al (2008) An open-label, single-arm pilot study in patients with severe plaque-type psoriasis treated with an oral anti-inflammatory agent, apremilast. Curr Med Res Opin 24(5):1529–1538
Kavanaugh A, Gladman DD, Edwards CJ, Schett G, Guerette B, Delev N et al (2019) Long-term experience with apremilast in patients with psoriatic arthritis: 5-year results from a PALACE 1–3 pooled analysis. Arthritis Res Ther 21(1):118
Hatemi G, Melikoglu M, Tunc R, Korkmaz C, Turgut Ozturk B, Mat C et al (2015) Apremilast for Behçet’s syndrome–a phase 2, placebo-controlled study. N Engl J Med 372(16):1510–1518
Perez-Aso M, Montesinos MC, Mediero A, Wilder T, Schafer PH, Cronstein B (2015) Apremilast, a novel phosphodiesterase 4 (PDE4) inhibitor, regulates inflammation through multiple cAMP downstream effectors. Arthritis Res Ther 17(1):249
Gottlieb AB, Matheson RT, Menter A, Leonardi CL, Day RM, Hu C et al (2013) Efficacy, tolerability, and pharmacodynamics of apremilast in recalcitrant plaque psoriasis: a phase II open-label study. J Drugs Dermatol 12(8):888–897
Schafer PH, Parton A, Capone L, Cedzik D, Brady H, Evans JF et al (2014) Apremilast is a selective PDE4 inhibitor with regulatory effects on innate immunity. Cell Signal 26(9):2016–2029
Khaled M, Levy C, Fisher DE (2010) Control of melanocyte differentiation by a MITF-PDE4D3 homeostatic circuit. Genes Dev 24(20):2276–2281
D’Orazio JA, Nobuhisa T, Cui R, Arya M, Spry M, Wakamatsu K et al (2006) Topical drug rescue strategy and skin protection based on the role of Mc1r in UV-induced tanning. Nature 443(7109):340–344
Majid I, Imran S, Batool S (2019) Apremilast is effective in controlling the progression of adult vitiligo: A case series. Dermatol Ther. 32(4):e12923
Huff SB, Gottwald LD (2017) Repigmentation of Tenacious Vitiligo on Apremilast. Case Rep Dermatol Med 2017:2386234
Kim HJ, Singer G, Abittan BJ, Chima MA, Kimmel G, Bares J et al (2021) Combination of apremilast and narrowband ultraviolet B light in the treatment of generalized vitiligo in skin phototypes IV to VI: A randomized split-body pilot study. J Am Acad Dermatol. 85(6):1657–1660
Saternus R, Pilz S, Gräber S, Kleber M, März W, Vogt T et al (2015) A closer look at evolution: Variants (SNPs) of genes involved in skin pigmentation, including EXOC2, TYR, TYRP1, and DCT, are associated with 25(OH)D serum concentration. Endocrinology 156(1):39–47
Seberg HE, Van Otterloo E, Cornell RA (2017) Beyond MITF: Multiple transcription factors directly regulate the cellular phenotype in melanocytes and melanoma. Pigment Cell Melanoma Res 30(5):454–466
Bianchi L, Del Duca E, Romanelli M, Saraceno R, Chimenti S, Chiricozzi A (2016) Pharmacodynamic assessment of apremilast for the treatment of moderate-to-severe plaque psoriasis. Expert Opin Drug Metab Toxicol 12(9):1121–1128
Alonso S, Izagirre N, Smith-Zubiaga I, Gardeazabal J, Díaz-Ramón JL, Díaz-Pérez JL et al (2008) Complex signatures of selection for the melanogenic loci TYR, TYRP1 and DCT in humans. BMC Evol Biol 8:74
Wehrle-Haller B (2003) The role of Kit-ligand in melanocyte development and epidermal homeostasis. Pigment Cell Res 16(3):287–296
Lim HW, Grimes PE, Agbai O, Hamzavi I, Henderson M, Haddican M et al (2015) Afamelanotide and narrowband UV-B phototherapy for the treatment of vitiligo: a randomized multicenter trial. JAMA Dermatol 151(1):42–50
Papp K, Reich K, Leonardi CL, Kircik L, Chimenti S, Langley RG et al (2015) Apremilast, an oral phosphodiesterase 4 (PDE4) inhibitor, in patients with moderate to severe plaque psoriasis: Results of a phase III, randomized, controlled trial (Efficacy and Safety Trial Evaluating the Effects of Apremilast in Psoriasis [ESTEEM] 1). J Am Acad Dermatol 73(1):37–49
Paul C, Cather J, Gooderham M, Poulin Y, Mrowietz U, Ferrandiz C et al (2015) Efficacy and safety of apremilast, an oral phosphodiesterase 4 inhibitor, in patients with moderate-to-severe plaque psoriasis over 52 weeks: a phase III, randomized controlled trial (ESTEEM 2). Br J Dermatol 173(6):1387–1399
Acknowledgements
This study received funding from Celgene Corporation.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
Dr. Lebwohl has received research funds from Abbvie, Amgen, Arcutis, AstraZeneca, Boehringer Ingelheim, Celgene, Clinuvel, Corrona.Inc, Eli Lilly, Foundation for Research & Education in Dermatology, Incyte, Janssen Research & Development, Kadmon Corp, Leo Pharmaceuticals, MedImmune, Novartis, Ortho Dermatologics, Pfizer, Sciderm, UCB.Inc, and ViDac, and is a consultant for Allergan, Almirall, Arcutis, Boehringer Ingelheim, Bristol-Myers Squibb, Castle Biosciences, Leo Pharma, Menlo, Mitsubishi, Neuroderm, Pfizer, Promius/Dr. Reddy's Laboratories, Theravance, and Verrica. Dr. Guttman has received research funds from Abbvie, Celgene, Eli Lilly, Janssen, MedImmune/AstraZeneca, Novartis, Pfizer, Regeneron, Vitae, Glenmark, Galderma, Asana, Innovaderm, Dermira and UCB, and is also a consultant for Sanofi Aventis, Regeneron, Stiefel/GlaxoSmithKline, MedImmune, Celgene, Anacor, AnaptysBio, Dermira, Galderma, Glenmark, Novartis, Pfizer, Vitae, Leo Pharma, Abbvie, Eli Lilly, Kyowa, Mitsubishi Tanabe, Asana Biosciences, and Promius. Other authors have no conflicts of interest to declare.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Kim, H.J., Del Duca, E., Pavel, A.B. et al. Apremilast and narrowband ultraviolet B combination therapy suppresses Th17 axis and promotes melanogenesis in vitiligo skin: a randomized, split-body, pilot study in skin types IV–VI. Arch Dermatol Res 315, 215–221 (2023). https://doi.org/10.1007/s00403-022-02343-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00403-022-02343-1