
Abstract
Lithium, a widely used drug for treating affective disorders, is known to inhibit glycogen synthase kinase-3 (GSK-3), which is one of the major tau kinases. Thus, lithium could have therapeutic benefit in neurodegenerative tauopathies by reducing tau hyperphosphorylation. We tested this hypothesis and showed that long-term administration of lithium at relatively low therapeutic concentrations to transgenic mice that recapitulate Alzheimer’s disease (AD)-like tau pathologies reduces tau lesions, primarily by promoting their ubiquitination rather than by inhibiting tau phosphorylation. These findings suggest novel mechanisms whereby lithium treatment could ameliorate tauopathies including AD. Because lithium also has been shown to reduce the burden of amyloid-β pathologies, it is plausible that lithium could reduce the formation of both amyloid plaques and tau tangles, the two pathological hallmarks of AD, and thereby ameliorate the behavioral deficits in AD.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Abnormal tau proteins are implicated in the mechanisms of brain degeneration in Alzheimer’s disease (AD), frontotemporal dementia with parkinsonism linked to chromosome 17, Pick’s disease and a group of neurodegenerative diseases that are collectively known as tauopathies [20]. A number of well-defined functions of tau proteins have been discovered and extensively characterized [4]. Most notably, tau binds to and stabilizes microtubules (MTs), in addition to promoting MT polymerization [7, 10, 37]. Aberrant phosphorylation of tau, which is seen in most tauopathies, causes loss of MT binding (loss of function) and/or promotes tau aggregation (gain of toxic function) [1, 20]. We have reported that transgenic (Tg) mice overexpressing human tau protein develop filamentous tau aggregates in the CNS. We overexpressed the smallest human tau isoform (T44) in the mouse CNS with the mouse prion protein (PrP) promoter to model tauopathies. These tau Tg mice (PrP T44 Tg) acquire age-dependent CNS pathologies, including insoluble, hyperphosphorylated tau and argyrophilic intraneuronal inclusions formed by tau-immunoreactive filaments [16, 17, 18, 28]. Therefore, these Tg mice are a model that can be exploited for drug discovery in studies that target amelioration of tau-induced neurodegeneration, as well as for elucidating mechanisms of tau pathology in various neurodegenerative tauopathies [19].
Many studies have shown that lithium, one of the most widely used drugs for treating affective disorder [2], inhibits glycogen synthase kinase-3 (GSK-3), and thereby affects multiple cellular functions [2, 3]. Because GSK-3 is one of the major kinases for tau, its inhibition is expected to protect against tauopathies. Indeed, some studies have shown that lithium reduced tau phosphorylation and cytotoxicity both in cultured cells and in rat brain in vivo [14, 26, 27, 36]. There are also accumulating data from clinical studies that correlate GSK-3 with tau pathologies. For example, active GSK-3 localizes to pretangle neurons, dystrophic neuritis, and neurofibrillary tangles (NFTs) in AD brains [29], and a spatial and temporal pattern of increased active GSK-3 expression coinciding with the progression of NFT and neurodegeneration has been observed [21], although increased levels of total GSK-3 have not been consistently observed.
Here, we tested the hypothesis that lithium treatment could have therapeutic benefit in neurodegenerative tauopathies by reducing tau hyperphosphorylation. We treated PrP T44 Tg mice with lithium chloride (LiCl) for 5 months, and evaluated the consequences of the treatment on tau inclusions, the phosphorylation state of tau, and motor behavior. Indeed, long-term administration of lithium at therapeutic concentrations reduced tau lesions, unexpectedly and interestingly, by promoting their ubiquitination rather than by inhibiting tau phosphorylation. These findings suggest novel mechanisms whereby lithium treatment could ameliorate tauopathies, including AD.
Materials and methods
Generation of mice
A transgene including a cDNA of the shortest human tau isoform (T44) driven by the mouse PrP promoter and 3’ untranslated sequences was used to create tau Tg mice on a B6D2/F1 background. Studies characterizing three lines of PrP T44 Tg mice generated with this transgene were described earlier [16]. The heterozygous Tg mouse lines 7, 43, and 27 overexpress human tau proteins at levels approximately 5-, 10-, and 15-fold higher than endogenous mouse tau, respectively. The heterozygous line 27 with the highest levels of Tg tau is not viable beyond 3 months, and none of the homozygous mice generated from any of these lines survive longer than 3 months. Therefore, we conducted the studies described here on heterozygous line 7 PrP T44 Tg mice and WT littermate control mice. Many studies and experiments have been conducted on the T44 Tg mice over the past 5 years, and we have found that neither gender differences among the phenotypes in these Tg mice nor whether the transgene is inherited from the father or the mother affects features of the phenotype in these mice or their response to the treatments described here.
Treatment with lithium
After a preliminary experiment to examine the phosphorylation state of tau after 2- or 4-week LiCl treatment in approximately 6-month-old Tg mice (Li-2w or Li-4w, respectively), untreated mice were randomly divided into three experimental groups. Mice in the control group (Cont) were fed standard chow. A second group of mice was treated early with lithium (Li-E group) by feeding them chow containing 2.0 g LiCl per kg chow from 2 to 6 months of age to evaluate the ability of LiCl in preventing formation of tau pathologies. A third group of mice was treated later (Li-L) with the same dose of LiCl in their chow from 5 to 9 months of age to determine if LiCl could reduce the burden of existing tau pathologies. To prevent hyponaturemia (a side effect of long-term lithium treatment), 1.8 g NaCl per kg chow was added to the chow supplemented with LiCl. All animals were treated in accordance with the Guidelines for Animal Experimentation of Okayama University.
Immunohistochemical analyses
Tau Tg and WT mice were anesthetized and perfused transcardially with 20 ml phosphate-buffered saline (PBS) followed by 20 ml 70% ethanol in isotonic saline. A representative series of 6-μm-thick paraffin sections of Tg and WT mouse spinal cord were immunostained by standard streptavidin-biotin-peroxidase methods, as described [16], using well-characterized antibodies (Table 1). Axonal tau pathology in the tau Tg mice was quantified by counting the number of tau-positive spheroids with a diameter of larger than 10 μm in 9–12 lumbar spinal cord sections of Tg mice from each group (n=4–6), and the average number of spheroids per section was used as a representative value. The histological quantification of the spheroids was done by a person who was blind to the experimental status of the animals. In this experiment, the mean and SEM values of the number of spheroids at the age of 6 and 9 months were calculated, and the difference in the number of spheroids among the groups was examined statistically by one-way ANOVA.
Confocal laser scanning microscopy
Double-labeling immunofluorescence staining was performed with a combination of primary antibodies Tau5/ubiquitin diluted 1:2,000 and 1:500 respectively. Sections were deparaffinized and nonspecific binding was blocked. Sections were first incubated in a mixture of the two primary antibodies overnight at 4°C and then in fluorescence-labeled secondary antibodies [Alexa 488-labeled anti-rabbit IgG (H+L) and Alexa 594-labeled anti-mouse IgG (H+L) (Molecular Probes, Eugene, OR)] for 1 h. Sections were viewed with a confocal microscope (FV300, OLYMPUS, Tokyo, Japan).
Tau protein solubility in the CNS of mice
Brains and spinal cords were dissected from lethally anesthetized 6- or 9-month-old Tg and WT mice, and methods similar to those described recently were used in the isolation procedures here [13, 16, 17, 18, 28]. Briefly, brain and spinal cord tissues were sequentially extracted with ice-cold high-salt RAB buffer [0.1 M MES, 1 mM EGTA, 0.5 mM MgSO4, 0.75 M NaCl, 0.02 M NaF, 1 mM PMSF, and 0.1% protease inhibitor cocktail: 100 μg/ml each of pepstatin A, leupeptin, TPCK, TLCK, soybean trypsin inhibitor, and 100 mM EDTA (Sigma-Aldrich, St. Louis, MO), pH 7.0], followed by RIPA buffer (50 mM Tris, 150 mM NaCl, 1% NP40, 5 mM EDTA, 0.5% sodium deoxycholate, and 0.1% SDS, pH 8.0) and finally with 70% formic acid (FA). Equal amounts of samples were subsequently resolved on 7.5% SDS-PAGE gels and transferred onto nitrocellulose membranes. Western blot analysis was performed, and the bands detected by enhanced chemiluminescence (ECL) reagent (Amersham, Bucks, UK) were analyzed quantitatively.
Determination of the phosphorylation state of tau in Tg mice
To assess the phosphorylation state of tau in lithium-treated T44 Tg mice biochemically, spinal cord tissues from 6-month-old Tg mice from Cont or Li-E groups were extracted as described above. As previously reported [16], a simplified protocol was adopted to generate the RAB-insoluble fractions. The RAB-insoluble pellets were sonicated in sample buffer containing 0.2 g/ml sucrose, 18.5 mM Tris pH 6.8, 2 mM EDTA, 80 mM DTT, and 2% SDS, and the 50,000 g supernatants were used as insoluble tau samples. Western blot analysis was performed using phosphorylation-dependent tau antibodies. GSK-3β was also assessed biochemically. Briefly, fresh spinal cord tissues were dissected and homogenized in RIPA buffer; approximately 15-μg samples were loaded on 7.5% SDS-PAGE gels, and the activation state of GSK-3β was detected with GSK-3β (Ser 9) antibody (Table 1).
Tail suspension test
The tail suspension test was performed as described previously [17, 28, 38]. Briefly, mice from each experimental group (n=9–12) were videotaped while being suspended by their tails for 15 s at 6 and 9 months of age. Animals were assessed for clasping behavior. The test period was divided into 2 s segments. An animal received a score of 1 point for each abnormal movement it displayed during each time segment. An abnormal movement was defined as dystonic movements of the hind limbs, or a combination of hind limbs, forelimbs and trunk, during which the limbs were pulled into the body in a manner not observed in WT mice. Unbiased sampling was used in this test.
Co-immunoprecipitation of tau with ubiquitin or CHIP
Spinal cord tissues from 6-month-old Tg mice from Cont, Li-4w, and Li-E groups and 9-month-old Tg mice from the Li-L group were extracted, and supernatants of RIPA buffer homogenates were prepared as described above. Samples were incubated with ~2 μg antibodies against ubiquitin or the C terminus of the Hsc70-interacting protein (CHIP) and 50 μl protein A (Amersham Bioscience, Uppsala, Sweden) with rocking at 4°C overnight. The protein A beads were pelleted and washed three times with immunoprecipitation buffer. The precipitates were resolved on SDS-PAGE gels and subjected to Western blotting analysis using Tau5 or PHF1 antibodies.
Results
Mice treated with LiCl did not show any apparent clinical symptoms (e.g., weight loss, bad fur condition), and their plasma LiCl levels (0.523±0.045 mM) were in the therapeutic range of clinical use (0.4–1.2 mM). There was no significant difference in plasma levels of Na+ between LiCl-treated and -untreated mice (data not shown). Because the major pathology found in the PrP T44 Tg mice is the presence of tau-immunopositive spheroids in the spinal cord, we sought to determine whether treatment with LiCl would have any effect on the number of spheroids. As reported previously [16, 17], the number of tau-positive spheroids in the spinal cord of the Tg mice increased with age until 6 months and decreased thereafter. In the older Tg mice, many vacuolar lesions of the same size or larger than the inclusions were also observed in the spinal cord, which may reflect the degradation of tau-positive spheroids and/or degeneration of affected neurons. As shown in Fig. 1, the number of tau-positive spheroids unexpectedly increased in the Tg mice from the Li-E group compared with those of the Cont group at 6 months, but by 9 months, the number of inclusions had decreased dramatically, and instead, many vacuolar lesions were observed (Fig. 2). In the spinal cord of these Tg mice, the number of neurons had decreased ~9% or ~16% at 6 or 9 months, respectively, compared with WT littermates. In this study, although the decrease was observed in Tg mice from all experimental groups compared with WT littermates, no significant differences in the number of neurons in the spinal cords were detected among the Tg mice at 9 months (Cont:Li-E:Li-L=1:1.11:0.98), suggesting that vacuolar lesions found in the spinal cord of Tg mice from Li-E group reflect the degradation of tau-positive spheroids rather than degeneration of affected neurons.

Quantification of tau-positive spheroids in the spinal cord of each experimental group. Mice in the control group (Cont) were fed standard chow. A second group of mice was treated early with lithium (Li-E) by feeding them chow containing 2.0 g LiCl/kg chow from 2 to 6 months of age. A third group of mice was treated later (Li-L) with the same dose of LiCl from 5 to 9 months of age. Quantification of spheroids is described under Materials and methods. The mean numbers of spheroids among experimental groups are summarized here. The error bars represent SEM. Statistical analysis was performed by ANOVA using mice at 6 and 9 months (n=4–6). A significant difference between each experimental group is indicated by asterisks (*P<0.05; ***P<0.001)

Vacuolar lesions in the spinal cord of LiCl-treated mice. Many vacuolar lesions are observed in the spinal cord of Tg mice from Li-E group at 9 months (arrows). No obvious differences in the number or distribution of neurons are observed between the Tg mice from Cont and Li-E groups. Bar 100 μm
We have reported that tau protein becomes progressively more insoluble with age in the PrP T44 Tg mice, as in human tauopathies [16, 17, 18]. As previously reported [16], the expression level of tau in the CNS of these Tg mice is stable throughout their lifetime, and no change was detected in the expression level of tau with LiCl treatment (data not shown). To determine the effect of LiCl treatment on the accumulation of insoluble tau, we analyzed the solubility of tau protein in the different experimental groups by extracting brain and spinal cord samples using buffers with increasing extraction strengths. The brain and spinal cord samples from 6- or 9-month-old mice from each group were sequentially extracted with RAB, RIPA buffer, and 70% FA, as described in Materials and methods. The three fractions were then analyzed by quantitative Western blotting with antibody 17026, a polyclonal antibody to recombinant tau protein. As shown in Fig. 3 and reported previously [16, 17, 18], over 90% of endogenous mouse tau from both the brain and spinal cord of the WT mouse was primarily RAB soluble, and no tau immunoreactivity was detected in the FA-soluble fraction. At 6 months of age, ~76% and ~74% of total tau proteins were RAB soluble, and ~1.2% and ~1.7% were found in the FA-soluble fraction of the brain and spinal cord, respectively, of the Tg mice [17]. Although the RAB-soluble tau remained relatively constant in Tg mice in all experimental groups, the intensity of the FA-soluble fractions was significantly decreased in the spinal cords of Tg mice from Li-E group at both 6 and 9 months. A solubility study of brain samples showed similar results (data not shown). Thus, these results suggest that the Tg mice in the Li-E group develop tau aggregates in the spinal cord that are not as insoluble as those in untreated tau Tg mice.

Reduced accumulation of insoluble tau protein in the CNS of LiCl-treated mice. a Spinal cord tissues of 6-month-old WT and Tg mice from Cont and Li-E groups were sequentially extracted with RAB, RIPA buffer, and 70% FA, and the tau levels were determined by quantitative Western blotting with antibody 17026. b Changes in the RIPA buffer- and FA-soluble tau as intensity of Western blotting using antibody 17026 of 6-month-old Tg mice from Cont and Li-E and 9-month-old Tg mice from the three groups are summarized. The intensity of the FA-soluble tau fraction was significantly decreased in Tg mice from the Li-E group compared with that of Tg mice from the Cont group at 6 and 9 months (n=4–5, error bars; SEM, *P<0.05, **P<0.01) (WT wild type, FA formic acid)
As reported previously [16, 17], tau aggregates in the spinal cord of PrP T44 Tg mice are positive for various phosphorylation-dependent tau antibodies histochemically and biochemically. As a preliminary experiment before starting this long-term LiCl treatment study, we examined the activity of GSK-3β and also the phosphorylation state of tau after 2- or 4-week LiCl treatment in ~6-month-old Tg mice (Li-2w or Li-4w, respectively). As shown in Fig. 4a, LiCl treatment did not produce an obvious difference in the expression level of GSK-3β. After LiCl treatment for 2 weeks (Li-2w), increased reactivity to GSK-3β Ser 9 antibody (Table 1) was observed, indicating decreased activity of GSK-3β, although this alteration became almost undetectable after 4-week treatment (Li-4w), and no change was detected at all in the activity of GSK-3β after 5-month treatment (Li-E, Li-L) (data not shown). As shown in Fig. 4b, tau protein extracted from the spinal cord of Tg mice from the Li-2w and Li-4w groups had less reactivity to antibodies to phosphorylated tau. Notably, tau from Tg mice of the Li-2w group showed less reactivity than tau from Tg mice of the Li-4w group, but after 5 months of LiCl administration, no reduction of tau phosphorylation was detected histochemically (Fig. 4c) or biochemically (Fig. 4d) with phosphorylation-dependent tau antibodies. Including the experiments shown in Fig. 4, all ten different phosphorylation-dependent tau antibodies listed in Table 1 were tried, but no obvious difference in the tau phosphorylation was observed after 5 months of lithium treatment (data not shown). Taken together with the results of the preliminary experiments, lithium treatment had a transient inhibitory effect on tau phosphorylation via inhibition of GSK-3β activity in the PrP T44 Tg mice for a limited period of time; then this effect decreased, and tau phosphorylation became undetectable within months. We investigated the expression levels of tau kinases, including GSK-3, cyclin-dependent kinase 5 (Cdk5), p38, extracellular signal-related kinase (ERK) 1 and ERK2 using quantitative Western blotting in each experimental group but did not detect obvious alterations (data not shown).

Phosphorylation state of tau in the spinal cord of mice. a GSK-3β activity was assessed on RIPA buffer-extracted samples from the spinal cords of 6-month-old Tg mice from the Cont, Li-2w, and Li-4w groups. No alteration in expression level of total GSK-3β was detected after LiCl treatment (upper panel). After LiCl treatment for 2 weeks, increased reactivity to GSK-3β Ser 9 antibody (Table 1) is observed, indicating decreased activity of GSK-3β, although this alteration becomes almost undetectable after 4-week treatment (lower panel). b Tau protein was extracted with RIPA buffer from the spinal cords of 6-month-old Tg mice from the Cont, Li-2w, and Li-4w groups, followed by Western blotting using the phosphorylation-dependent tau antibody AT180 or PHF1. Tau protein extracted from the spinal cords of Tg mice from Li-2w and Li-4w groups has less reactivity to antibodies to phosphorylated tau. Tau from Tg mice of the Li-2w group shows less reactivity than that of the Li-4w group. c Tau aggregates in the spinal cords of 6-month-old Tg mice from Cont or Li-E groups are stained with the antibody PHF1. Bar 100 μm, insets 10 μm. d Western blotting of tau extracted from the spinal cords of 6-month-old Tg mice with a panel of phosphorylation-dependent tau antibodies. No reduction of tau phosphorylation is detected histochemically or biochemically after 5-month treatment with LiCl (c, d)
T44 Tg mice develop progressive motor weakness, as demonstrated by an impaired ability to stand on a slanted surface and by clasping their hind limbs when lifted by the tail. Previous studies on T44 Tg mice have shown an overall correlation between the number of tau-positive inclusions in the spinal cord and severity of the behavioral phenotype [16, 17, 28]. However, whereas long-term LiCl treatment of the Tg mice from the Li-E group increased the number of tau-positive inclusions in the spinal cord at 6 months, clasping behavior scores of Tg mice in the Li-E group were lower, although not significantly, than scores of the control group. Further, a significant ameliorative effect of LiCl on motor impairments was observed at 9 months (Fig. 5).

Clasping phenotype of mice. Mice were videotaped during a 15-s tail suspension test at 6 or 9 months of age and observed for clasping behavior as described under Materials and methods. Tau Tg mice from the Li-E group show a lower score than Tg mice from the Cont group at 6 and 9 months, although not significantly at 6 months. Bar graphs show means ± SEM of five trials (n=9-12, *P<0.05, **P<0.01, ***P<0.001)
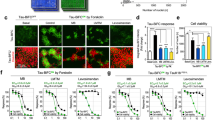
To examine the mechanism(s) of the dramatic changes in the number of tau-positive spheroids in the spinal cord of PrP T44 Tg mice in the Li-E group from 6 to 9 months, spinal cord sections were stained with a ubiquitin antibody. As previously reported [16, 18] and shown in Fig. 6a, tau-positive spheroids in the spinal cord of these Tg mice are ubiquitin negative, but after 5 months of LiCl treatment, about 50% of tau-positive spheroids of the Li-E group were ubiquitin positive (Fig. 6a, b). This immunoreactivity was also detected in Tg mice from the Li-4w and Li-L groups at 6 and 9 months of age, respectively, although it was weaker than that of Tg mice from the Li-E group. Because the CHIP has been identified as a ubiquitin ligase for tau [31, 33], we examined CHIP and found that tau lesions positive for ubiquitin were generally also positive for CHIP (Fig. 6b). The increased ubiquitination of tau with CHIP as a ligase is also supported by the results of immunoprecipitation studies (Fig. 7). Figure 7 shows that after immunoprecipitation with ubiquitin or CHIP antibodies, immunoreactivity to phosphorylated tau significantly increased with long-term LiCl treatment. Quantitative Western blotting showed no alteration in the expression levels of ubiquitin or CHIP with LiCl treatment (data not shown).

Tau aggregates in the spinal cord of T44 Tg mice were ubiquitinated with LiCl treatment. a Spinal cord sections of 6-month-old Tg mice from Cont, Li-E, and Li-4w groups or 9-month-old Tg mice from the Li-L group were stained with a ubiquitin antibody (DakoCytomation, Denmark). As previously reported [16, 18], tau-positive spheroids in the spinal cord of these Tg mice are ubiquitin negative (Cont 6m), but after long-term LiCl treatment, a substantial portion of the tau-positive spheroids become ubiquitin positive. b Double-immunofluorescence staining was performed with a combination of primary antibodies, Tau5 and a ubiquitin antibody. After confocal laser scanning microscopy, antibodies were stripped and sections were stained by regular immunostaining using an antibody to CHIP (Calbiochem, San Diego, CA). Colocalization of tau, ubiquitin, and CHIP is seen in the spinal cord lesions of PrP T44 Tg mice

Tau ubiquitination with LiCl treatment was mediated by CHIP. Tau immunoprecipitated with antibodies to ubiquitin or CHIP was eluted and analyzed by quantitative Western blotting using antibodies Tau5 or PHF1. The intensity of the phosphorylation-dependent antibody PHF1 signal is summarized (n=4–5, error bars; SEM, *P<0.05, **P<0.01). Phosphorylated tau was preferentially immunoprecipitated with ubiquitin and CHIP antibodies after long-term LiCl treatment. Input signals were probed with antibodies to ubiquitin or CHIP (upper or lower panels, respectively)
Discussion
Treatment of tau Tg mice with LiCl for as long as 5 months has not been previously reported; thus, there are no data about the phosphorylation state of tau after long-term LiCl treatment. The inhibitory effect of LiCl on tau phosphorylation became undetectable after 5-month LiCl treatment (Fig. 4), suggesting the existence of some compensatory mechanisms, although we did not detect any alteration in the expression levels of tau kinases (including GSK-3, Cdk5, p38, ERK1 and ERK2) or in the GSK-3 activity. Other research groups have reported that tau phosphorylation was attenuated in tau Tg mice after 30-day or 6-week LiCl treatment [29, 31], and that phenomenon seems to be similar to the data from our preliminary study in which the reduction of tau phosphorylation was detected after 2- or 4-week LiCl treatment. However, we cannot conclude that their data are consistent with ours presented here because of differences in the LiCl dose, method of administration, treatment duration and tau Tg mouse line. For example, it was reported that the plasma concentration of lithium of treated mice in one of their studies was 1.24±0.1 mM [31], while in our pilot study, mice showed weight loss and bad fur condition, and moreover, about 50% of mice died within 2 months at plasma concentrations above 1 mM (data not shown), suggesting that such concentrations were possibly toxic. Nonetheless, it seems that the long-term effect of lithium on tau phosphorylation in vivo is different from that predicted from cell culture studies or acute in vivo studies [14, 25, 26, 27, 36], although further studies are needed to elucidate the mechanism(s) of this disparity.
The proteasome pathways involved in tau degradation have been reported but not fully elucidated. For example, although tau can be degraded via the ubiquitination cascade [32, 34], some cell culture studies reported the existence of ubiquitin-independent pathways for the proteasomal degradation of tau [8, 9]. Notably, it is also difficult to say whether the ubiquitination observed in diseases is a consequence of a neuroprotective event or neurodegenerative one. Ubiquitination can primarily be a neuroprotective event that contributes to clearance of pathogenic proteins, but on the other hand, it is possibly a sign of disturbance of proteasomal pathways as a result of overwhelming damage. Our data suggest that the ubiquitination of tau lesions in the spinal cord of LiCl-treated PrP T44 Tg mice may account for the dramatic decrease in the number of tau-positive spheroids in the spinal cord of the Li-E group. However, the mechanism(s) by which long-term lithium treatment induces ubiquitination of tau lesions is unclear, and we cannot determine whether or not the effect of long-term lithium treatment is caused by alteration of GSK-3 activity. Although induction of ubiquitination of tau lesions was detected in the Li-L group (in which LiCl treatment was started after development of tau pathologies), clearance of tau aggregates was limited, suggesting that LiCl treatment should be started at an early stage of the development of tau pathologies.
Lithium inhibits one of the major tau kinases, GSK-3, and it also affects neuroprotection or neurogenesis by mechanisms that are not fully understood [3, 11]. For example, some clinical studies showed that lithium treatment increased neuronal viability and/or function, and, moreover, that it could increase the volume of gray matter via neurotrophic effects [23, 24]. These and possibly yet-unknown effects of lithium may have contributed to the results presented here. For example, the inhibition of tau phosphorylation with LiCl, which occurred for at least several weeks, may reduce tau-induced neurotoxicity. Further, lithium may have a neuroprotective effect that is independent of its effect on tau phosphorylation by synergistically decreasing the vulnerability of cells and allowing them to bear more tau aggregates “safely” without cell degeneration until clearance via the ubiquitination cascade. Although lithium has effects that do not relate to tau phosphorylation, it should be noted that these effects (including neuroprotection) likely also involve GSK-3 inhibition [5, 6, 15, 22, 35]. Therefore, for treatment of neurodegenerative diseases by inhibiting of GSK-3, more-selective GSK-3 inhibitors may be considered as a possible alternative.
The clinically effective plasma concentration of lithium for treating affective disorders is about 0.4–1.2 mM, and levels in excess of about 1.5 mM cause nausea, vomiting, diarrhea, hand tremors or even convulsions and coma, and may be toxic. Thus, the safety of lithium treatment requires close monitoring, especially when it is used for long periods in the elderly, who may be more vulnerable to the toxic effects of lithium. However, the beneficial effects of lithium treatment shown here in PrP T44 Tg mice may imply that lithium could have therapeutic benefit in human tauopathies. Although the mechanisms whereby long-term lithium treatment promotes the ubiquitination of tau-positive inclusions and reduces tau-induced pathologies remain to be elucidated, these findings provide new insights into potential therapeutic interventions for patients with tauopathies. Indeed, since lithium has been shown to also reduce the burden of amyloid-β pathologies [3, 33], it is plausible that lithium administration could reduce the formation of both amyloid plaques and NFTs, the two pathological hallmarks of AD.
References
Alonso A, Zaidi T, Novak M, Grundke-Iqbal I, Iqbal K (2001) Hyperphosphorylation induces self-assembly of tau into tangles of paired helical filaments/straight filaments. Proc Natl Acad Sci USA 98:6923–6928
Belmaker RH (2004) Bipolar disorder. N Engl J Med 351:476–486
Bhat RV, Budd Haeberlein SL, Avila J (2004) Glycogen synthase kinase 3: a drug target for CNS therapies. J Neurochem 89:1313–1317
Buee L, Bussiere T, Buee-Scherrer V, Delacourte A, Hof PR (2000) Tau protein isoforms, phosphorylation and role in neurodegenerative disorders. Brain Res Brain Res Rev 33:95–130
Chalecka-Franaszek E, Chuang DM (1999) Lithium activates the serine/threonine kinase Akt-1 and suppresses glutamate-induced inhibition of Akt-1 activity in neurons. Proc Natl Acad Sci USA 96:8745–8750
Chen G, Bower KA, Ma C, Fang S, Thiele CJ, Luo J (2004) Glycogen synthase kinase 3beta (GSK3beta) mediates 6-hydroxydopamine-induced neuronal death. FASEB J 18:1162–1164
Cleveland DW, Hwo SY, Kirschner MW (1977) Purification of tau, a microtubule-associated protein that induces assembly of microtubules from purified tubulin. J Mol Biol 116:207–225
David DC, Layfield R, Serpell L, Narain Y, Goedert M, Spillantini MG (2002) Proteasomal degradation of tau protein. J Neurochem 83:176–185
Delobel P, Leroy O, Hamdane M, Sambo AV, Delacourte A, Buee L (2005) Proteasome inhibition and Tau proteolysis: an unexpected regulation. FEBS Lett 579:1–5
Drechsel DN, Hyman AA, Cobb MH, Kirschner MW (1992) Modulation of the dynamic instability of tubulin assembly by the microtubule-associated protein tau. Mol Biol Cell 3:1141–1154
Eldar-Finkelman H (2002) Glycogen synthase kinase 3: an emerging therapeutic target. Trends Mol Med 8:126–132
Greenberg SG, Davis P (1990) A preparation of Alzheimer paired helical filaments that displays distinct tau proteins by polyacrylamide gel electrophoresis.Proc Natl Acad Sci USA 87:5827–5831
Higuchi M, Ishihara T, Zhang B, Hong M, Andreadis A, Trojanowski J, Lee VMY (2002) Transgenic mouse model of tauopathies with glial pathology and nervous system degeneration. Neuron 35:433–446
Hong M, Chen DC, Klein PS, Lee VMY (1997) Lithium reduces tau phosphorylation by inhibition of glycogen synthase kinase-3. J Biol Chem 272:25326–25332
Hongisto V, Smeds N, Brecht S, Herdegen T, Courtney MJ, Coffey ET (2003) Lithium blocks the c-Jun stress response and protects neurons via its action on glycogen synthase kinase 3. Mol Cell Biol 23:6027–6036
Ishihara T, Hong M, Zhang B, Nakagawa Y, Lee MK, Trojanowski JQ, Lee VMY (1999) Age-dependent emergence and progression of a tauopathy in transgenic mice overexpressing the shortest human tau isoform. Neuron 24:751–762
Ishihara T, Higuchi M, Zhang B, Yoshiyama Y, Hong M, Trojanowski JQ, Lee VMY (2001) Attenuated neurodegenerative disease phenotype in tau transgenic mouse lacking neurofilaments. J Neurosci 21:6026–6035
Ishihara T, Zhang B, Higuchi M, Yoshiyama Y, Trojanowski JQ, Lee VMY (2001) Age-dependent induction of congophilic neurofibrillary tau inclusions in tau transgenic mice. Am J Pathol 158:555–562
Lee VMY, Trojanowski JQ (2001) Transgenic mouse models of tauopathies: prospects for animal models of Pick’s disease. Neurology 56:S26–30
Lee VMY, Goedert M, Trojanowski JQ (2001) Neurodegenerative tauopathies. Annu Rev Neurosci 24:1121–1159
Leroy K, Boutajangout A, Authelet M, Woodgett JR, Anderton BH, Brion JP (2002) The active form of glycogen synthase kinase-3beta is associated with granulovacuolar degeneration in neurons in Alzheimer’s disease. Acta Neuropathol 103:91–99
Li X, Bijur GN, Jope RS (2002) Glycogen synthase kinase-3beta, mood stabilizers, and neuroprotection. Bipolar Disord 4:137–144
Moore GJ, Bebchuk JM, Hasanat K, Chen G, Seraji-Bozorgzad N, Wilds IB, Faulk MW, Koch S, Glitz DA, Jolkovsky L, Manji HK (2000) Lithium increases N-acetyl-aspartate in the human brain: in vivo evidence in support of bcl-2’s neurotrophic effects? Biol Psychiatry 48:1–8
Moore GJ, Bebchuk JM, Wilds IB, Chen G, Manji HK (2000) Lithium-induced increase in human brain grey matter. Lancet 356:1241–1242
Mudher A, Shepherd D, Newman TA, Mildren P, Jukes JP, Squire A, Mears A, Drummond JA, Berg S, MacKay D, Asuni AA, Bhat R, Lovestone S (2004) GSK-3beta inhibition reverses axonal transport defects and behavioural phenotypes in Drosophila. Mol Psychiatry 9:522–530
Munoz-Montano JR, Moreno FJ, Avila J, Diaz-Nido J (1997) Lithium inhibits Alzheimer’s disease-like tau protein phosphorylation in neurons. FEBS Lett 411:183–188
Munoz-Montano JR, Lim F, Moreno FJ, Avila J, Diaz-Nido J (1999) Glycogen synthase kinase-3 modulates neurite outgrowth in cultured neurons: possible Implications for neurite pathology in Alzheimer’s disease. J Alzheimers Dis 1:361–378
Nakashima H, Ishihara T, Yokota O, Terada S, Trojanowski JQ, Lee VMY, Kuroda S (2004) Effects of alpha-tocopherol on an animal model of tauopathies. Free Radic Biol Med 37:176–186
Noble W, Planel E, Zehr C, Olm V, Meyerson J, Suleman F, Gaynor K, Wang L, LaFrancois J, Feinstein B, Burns M, Krishnamurthy P, Wen Y, Bhat R, Lewis J, Dickson D, Duff K (2005) Inhibition of glycogen synthase kinase-3 by lithium correlates with reduced tauopathy and degeneration in vivo. Proc Natl Acad Sci USA 102:6990-6995
Pei JJ, Tanaka T, Tung YC, Braak E, Iqbal K, Grundke-Iqbal I (1997) Distribution, levels, and activity of glycogen synthase kinase-3 in the Alzheimer disease brain. J Neuropathol Exp Neurol 56:70–78
Perez M, Hernandez F, Lim F, Diaz-Nido J, Avila J (2003) Chronic lithium treatment decreases mutant tau protein aggregation in a transgenic mouse model. J Alzheimers Dis 5:301–308
Petrucelli L, Dickson D, Kehoe K, Taylor J, Snyder H, Grover A, De Lucia M, McGowan E, Lewis J, Prihar G, Kim J, Dillmann WH, Browne SE, Hall A, Voellmy R, Tsuboi Y, Dawson TM, Wolozin B, Hardy J, Hutton M (2004) CHIP and Hsp70 regulate tau ubiquitination, degradation and aggregation. Hum Mol Genet 13:703–714
Phiel CJ, Wilson CA, Lee VMY, Klein PS (2003) GSK-3alpha regulates production of Alzheimer’s disease amyloid-beta peptides. Nature 423:435–439
Shimura H, Schwartz D, Gygi SP, Kosik KS (2004) CHIP-Hsc70 complex ubiquitinates phosphorylated tau and enhances cell survival. J Biol Chem 279:4869–4876
Song L, De Sarno P, Jope RS (2002) Central role of glycogen synthase kinase-3beta in endoplasmic reticulum stress-induced caspase-3 activation. J Biol Chem 277:44701–44708
Takahashi M, Yasutake K, Tomizawa K (1999) Lithium inhibits neurite growth and tau protein kinase I/glycogen synthase kinase-3beta-dependent phosphorylation of juvenile tau in cultured hippocampal neurons. J Neurochem 73:2073–2083
Weingarten MD, Lockwood AH, Hwo SY, Kirschner MW (1975) A protein factor essential for microtubule assembly. Proc Natl Acad Sci USA 72:1858–1862
Yamamoto A, Lucas JJ, Hen R (2000) Reversal of neuropathology and motor dysfunction in a conditional model of Huntington’s disease. Cell 101:57–66
Acknowledgements
We thank Dr. P. Davies for PHF1 antibody. We also thank S. Fujisawa, M. Onbe, T. Kanamori and R. Wada for technical assistance. This work was supported by grants from the Japanese Ministry of Education, Culture, Sports, Science and Technology (T.I.), Taisho Pharmaceutical Co., Ltd. (T.I.), the Zikei Institute of Psychiatry (T.I., H.N.), the National Institutes of Health (V.M.-Y.L., J.Q.T.), as well as by grants from the Marian S. Ware Alzheimer Program (V.M.-Y.L., J.Q.T.).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Nakashima, H., Ishihara, T., Suguimoto, P. et al. Chronic lithium treatment decreases tau lesions by promoting ubiquitination in a mouse model of tauopathies. Acta Neuropathol 110, 547–556 (2005). https://doi.org/10.1007/s00401-005-1087-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00401-005-1087-4