
Abstract
Focusing on Sumatra, a hotspot of tropical lowland rainforest transformation, we investigated effects of the conversion of rainforests into rubber agroforests (“jungle rubber”), intensive rubber, and oil palm plantations on the communities of litter and soil microorganisms and identified factors responsible for these changes. Litter basal respiration, microbial biomass, total bacterial phospholipid fatty acids (PLFAs), and fungal PLFAs did not vary significantly with rainforest conversion. In litter of converted ecosystems, the concentration of certain PLFAs including the Gram-negative bacteria marker PLFA cy17:0 and the Gram-positive bacteria marker PLFA i17:0 was reduced as compared to rainforest, whereas the concentration of the arbuscular mycorrhizal fungi (AMF) marker neutral lipid fatty acid (NLFA) 16:1ω5c increased. As indicated by redundancy analysis, litter pH and carbon concentration explained most of the variation in litter microbial community composition. In soil, microbial biomass did not vary significantly with rainforest conversion, whereas basal respiration declined. Total PLFAs and especially that of Gram-negative bacteria decreased, whereas PLFA i17:0 increased with rainforest conversion. The concentration of fungal PLFAs increased with rainforest conversion, whereas NLFA 16:1ω5c did not change significantly. Redundancy analysis indicated that soil pH explained most of the variation in soil microbial community composition. Overall, the data suggest that conversion of rainforests into production systems results in more pronounced changes in microbial community composition in soil as compared to litter. In particular, the response of fungi and bacteria was more pronounced in soil, while the response of AMF was more pronounced in litter. Notably, only certain bacterial markers but not those of saprotrophic fungi and AMF were detrimentally affected by rainforest conversion.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Biodiversity and services of tropical forests are of global importance (Naeem et al. 2012; Cole et al. 2014); however, driven by the demand for cropland of a growing human population, they are among the most threatened ecosystems of the world, especially in Southeast Asia (Sodhi et al. 2004; Koh et al. 2011; Wilcove et al. 2013). This applies particularly to Indonesia, known as a hotspot of biodiversity, but now being one of the top world’s producer of palm oil (Fitzherbert et al. 2008; Koh et al. 2011) and second in rubber (Marimin et al. 2014). The conversion of rainforests into agricultural land is associated by deforestation and peatland degradation (Miettinen et al. 2013), resulting in nonreversible alteration in biodiversity and ecosystem functions (Sala et al. 2000; Gibbs et al. 2010). However, studies on effects of land use changes on biodiversity of plants, invertebrates and vertebrates, and ecosystem functioning in Southeast Asia are scarce; long-term studies are missing almost entirely (Fitzherbert et al. 2008; Sodhi et al. 2010). Thus, despite the large-scale conversion of rainforests worldwide, little is known on biodiversity and ecological functions in converted ecosystems, and this applies in particular to the belowground system (Giam et al. 2010; Wilcove and Koh 2010).
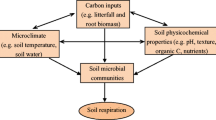
Lowland tropical rainforests are particularly species-rich, and soil biota account for a significant fraction of their overall biodiversity (Brussaard et al. 1997). Microorganisms form an essential component of any ecosystem reaching high diversity, abundance, and biomass in tropical rainforests (Whitman et al. 1998; Fierer et al. 2007; Roesch et al. 2007). A number of studies investigated effects of land use changes on soil microbial biomass, diversity, and community composition; however, the majority of data has been collected in South and Central America, e.g., Brazil, Ecuador, and Costa Rica (Borneman and Triplett 1997; Carney et al. 2004; Rodrigues et al. 2013; Tischer et al. 2014); less information is available from tropical Asia (Tripathi et al. 2012; Lee-Cruz et al. 2013). Further, the great majority of studies focused on mineral soil and neglected the litter layer. This is surprising since litter material in organic layers forms the living space for a variety of microorganisms and invertebrates and represents the focal compartment for organic matter decomposition and nutrient mineralization. Land use change is known to alter physicochemical properties in litter, and community composition and functioning of microorganisms are susceptible to these changes (Murty et al. 2002; Rousk et al. 2010; Chapin et al. 2012; Rasche and Cadisch 2013). However, there is hardly any information on how the structure of microbial communities responds to the conversion of rainforests into major agricultural systems such as oil palm and rubber in Southeast Asia. To fill this gap of knowledge, we investigated these changes focusing on Jambi province in southwest Sumatra with a total area of 50,058 km2, of which 16 % are covered into oil palm and 12 % into rubber plantations (Gatto et al. 2015). Forest conversion has increased markedly in the last 20 years, and this is predicted to continue in the future (Koh and Ghazoul 2010; Gatto et al. 2015). Therefore, Sumatra represents an ideal model region to investigate effects of changes in land use on biodiversity and ecosystem functioning at local but also regional scales. Knowledge on effects of rainforest conversion on microbial community composition and functioning is essential for understanding and predicting the response of tropical ecosystems to anthropogenic disturbances and environmental changes.
In addition to lowland rainforests, we investigated major rainforest transformation systems in Sumatra including rubber agroforests (“jungle rubber”) and intensively managed rubber and oil palm plantations. We aimed at quantifying the impact of rainforest conversion on the community of soil microorganism and identify the drivers responsible for these changes. To achieve this goal, we investigated biomass and community composition of microorganisms and explored potential driving factors, such as carbon and nitrogen concentrations, pH and water content in soil and litter. We hypothesized that (1) microbial communities in litter and soil are differentially affected, with the response of fungal communities being more pronounced in litter while that of bacterial communities being more pronounced in soil, as the latter is buffered against changes in factors associated with land transformation. Further, we hypothesized that (2) composition of bacterial and fungal communities differentially respond to land use changes and agricultural management as fungi are known to thrive in forests and to be detrimentally affected by agricultural management, while bacteria are less affected by disturbances associated with agricultural practices (van der Wal et al. 2006; García-Orenes et al. 2013; Baldrian 2014).
Materials and methods
Study sites and sampling
The study sites are located in tropical lowland rainforest in the Jambi province of southwest Sumatra, Indonesia. Four transformation systems, i.e., secondary rainforest, jungle rubber, rubber, and oil palm were replicated eight times across two landscapes, Bukit Duabelas and Harapan, resulting in 32 sampling sites. Secondary rainforest was considered as reference site and comprised old growth forests with minor logging; jungle rubber represented rubber agroforestry systems originating from enrichment of rainforest by rubber; rubber comprised monoculture rubber plantations with an average age of 13 years; oil palm comprised monoculture oil palm plantations with an average age of 14 years. Combined NPK, urea, and potassium chloride are used as fertilizers by smallholders and added in small amounts to jungle rubber, but at high rates to rubber and oil palm plantations. Further, lime is added at irregular intervals to rubber and oil palm plantations. Weeds typically are controlled by using glyphosate and/or paraquat in rubber and oil palm plantations but in part also manually. Soils in the Bukit Duabelas region are represented by clay Acrisols and in the Harapan region by loam Acrisols (Guillaume et al. 2015); for further information on the study sites, see Barnes et al. (2014) and Guillaume et al. (2015).
Samples from the litter/fermentation and top soil layers were taken in October/November 2013, using a corer of a diameter of 5 cm. To account for spatial variability in the thickness of the litter layer, litter samples from five cores from each plot were taken and pooled resulting in 32 pooled samples. Material from the litter layer consisted of a mixture of organic material including fresh and partly decomposed leaves from trees, herbs, and mosses (L/F horizon); twigs, roots, and coarse woody debris were removed by the hand. For mineral soil (Ah horizon), three soil cores to a depth of 5 cm were taken and pooled, again resulting in 32 pooled samples. Roots and large litter particles including seeds and twigs were removed by the hand. Samples were kept on ice during the sampling campaign and stored at −20 °C until used for the analyses.
Environmental factors
Litter and soil pH (CaCl2) were measured using a digital pH meter. Aliquots of litter and soil material were dried at 65 °C for 72 h, milled, and analyzed for total C and N concentrations using an elemental analyzer (Carlo Erba, Milan, Italy). Amount of litter and water content of litter and soil were determined gravimetrically.
Microorganisms
Basal respiration and microbial biomass were determined by measuring O2 consumption using an automated respirometer system (Scheu 1992; SIR method, Anderson and Domsch 1978; for details see Krashevska et al. 2012). Microbial community composition was analyzed by fatty acid analysis commonly used for analyzing major microbial groups in soil such as bacteria and fungi (Frostegård et al. 2011). The method has been proven to be among the most sensitive techniques for analyzing changes in microbial community composition (Ramsey et al. 2006; Joergensen and Wichern 2008; Strickland and Rousk 2010). For measuring phospholipid fatty acids (PLFAs) and neutral lipid fatty acids (NLFAs), 2 and 4 g wet litter and soil, respectively, were extracted following the procedure of Frostegård et al. (1993); for details, see Krashevska et al. (2010). Individual PLFAs were calculated as percentages of total PLFAs (relative nmol g−1 dry litter/soil material). The sum of PLFAs i15:0, a15:0, 15:0, i16:0, 16:1ω7, 17:0, i17:0, cy17:0, 18:1ω7, and cy19:0 represented total bacterial PLFAs (Frostegård and Bååth 1996). The sum of 16:1ω7, cy17:0, and cy19:0 was used as relative marker for Gram-negative, and the sum of i15:0, a15:0, i16:0, and i17:0 as relative marker for Gram-positive bacteria (Zelles 1997, 1999). The sum of 18:2ω6,9, 18:3ω6, 18:3ω3 was used to represent total fungal PLFAs (Frostegård and Bååth 1996; Ruess and Chamberlain 2010). The NLFA 16:1ω5c was used as marker for arbuscular mycorrhizal fungi (AMF; nmol g−1 dry litter/soil material; Lekberg et al. 2012; Ngosong et al. 2012).
Calculations and statistical analyses
Since litter and soil samples were taken from independent soil cores and the amount of litter varied markedly between systems, data from litter and soil samples were analyzed separately using analysis of variance. Effects of forest conversion on the measured abiotic and biotic variables of litter and soil were analyzed using forest transformation system (rainforest, jungle rubber, rubber, oil palm) as fixed factor (type 3 sum of squares) including landscape (Harapan, Bukit Duabelas) as random factor in SAS version 9.3 (SAS Institute, Cary, NC, USA). Tukey’s HSD test was used to identify significant differences between means. Levels of significance in text and figures are indicated as ns = P > 0.05, *P < 0.05, **P < 0.01, and ***P < 0.001.
Percentages of total PLFAs were analyzed by discriminant function analysis (DFA) to identify effects of conversionon PLFA composition in litter and soil layers. Squared Mahalanobis distances (MD2) between group centroids (rainforest, jungle rubber, rubber, oil palm) and the reliability of sample classification were determined. DFA was performed using STATISTICA 7.0 for Windows (StatSoft, Tulsa, USA). For identifying which of the fatty acids were responsible for significant differences between transformation systems, analyses of variance (ANOVA) for individual PLFAs as well as for groups of PLFAs (see subsection “Microorganisms”) were carried out in SAS (Scheiner and Gurevitch 2001). Prior to the analyses, relative abundances of PLFAs expressed as percentages were arcsine-square root transformed.
Relationships between microorganisms (dependent variables) and environmental factors (independent variables) were analyzed using redundancy analysis (RDA). RDA included only variables significantly affected by forest conversion (according to ANOVAs; P < 0.05). Monte-Carlo tests (999 permutations) were performed to evaluate overall significance and the significance of environmental variables and individual axes. Transformation systems (forest, jungle rubber, rubber, and oil palm) were included as passive variables. RDA was implemented in CANOCO 5.02 (Ter Braak and Šmilauer P 2012). Figures were prepared using SigmaPlot for Windows version 11.0.
Results
Environmental factors in litter
The amount of litter in the litter layer declined in the order F > J > R > O with 0.18 ± 0.05, 0.12 ± 0.03, 0.09 ± 0.04, and 0.03 ± 0.02 g cm−2, respectively. Therefore, as compared to rainforest, the amount of litter in jungle rubber, rubber, and oil palm was reduced by 30, 50, and 84 %, respectively (F3,27 = 30.2***). Water content declined in the order J ≥ F > O = R with 208 ± 82.8, 175 ± 74.9, 136 ± 71.3, and 118 ± 42.8 % of litter dry weight, respectively (F3,27 = 5.45**). Litter pH was generally higher in converted systems and decreased in the order O ≥ R > J > F with 5.62 ± 0.37, 5.57 ± 0.26, 5.08 ± 0.44, and 4.41 ± 0.67, respectively (F3,27 = 12.4***). Total litter C concentration was similar in jungle rubber (39.8 ± 5.85 %) and rainforest (38.4 ± 7.96 %) but lower in rubber (32.8 ± 10.1 %) and oil palm (29.3 ± 5.51 %; F3,27 = 9.71***). Total litter N concentration significantly declined from jungle rubber (1.43 ± 0.09 %) to rainforest (1.31 ± 0.23 %) to oil palm (1.13 ± 0.16 %) to rubber (1.06 ± 0.29 %; F3,27 = 9.84***). Litter C-to-N ratio varied little between transformation systems, but the differences were significant; it decreased from rubber (31.2 ± 2.82) to rainforest (29.5 ± 3.84) to jungle rubber (28.3 ± 4.06) to oil palm (26.6 ± 3.42; F3,27 = 3.44**).
Environmental factors in soil
Water content varied little between transformation systems and was on average 73.7 ± 49.8 % (F3,27 = 1.00ns). Soil pH generally was higher in converted systems and decreased in the order oil palm (4.70 ± 0.36), rubber (4.39 ± 0.10), jungle rubber (4.34 ± 0.14), and rainforest (3.78 ± 0.32; F3,27 = 23.0***). Total soil C concentration was similar in rainforest (6.04 ± 3.77 %) and jungle rubber (5.70 ± 2.37 %), but markedly lower in rubber (3.51 ± 1.62 %) and oil palm (3.42 ± 1.08 %; F3,27 = 4.10*). Similarly, total soil N concentration was similar in rainforest (0.36 ± 0.15 %) and jungle rubber (0.38 ± 0.13 %), but considerably lower in rubber (0.24 ± 0.07 %) and oil palm (0.25 ± 0.10 %; F3,27 = 5.96**). Soil C-to-N ratio did not vary significantly between transformation systems and was on average 14.6 ± 3.61 (F3,27 = 0.73ns).
Microorganisms in litter
In litter neither basal respiration (overall mean 80.5 ± 5.69 μg O2 g−1 dw h−1; F3,27 = 1.71ns) nor microbial biomass (8.69 ± 0.72 mg Cmic g−1 dw; F3,27 = 0.65ns) varied significantly between transformation systems. DFA of litter fatty acids separated rainforest from oil palm (MD2 = 64.4***) and rubber plantations (MD2 = 84.4***), but less from jungle rubber (MD2 = 33.0**; Wilk’s λ = 0.014, F102,78 = 7.41**). Based on ANOVA total bacterial PLFAs (35.4 ± 8.32 %; F3,27 = 0.70ns), Gram-negative (9.38 ± 3.76 %; F3,27 = 1.10ns) and Gram-positive bacterial PLFAs (12.2 ± 3.69 %; F3,27 = 0.05ns), sum of fungal PLFAs (26.09 ± 6.93 %; F3,27 = 1.59ns) did not vary significantly between transformation systems. In contrast, PLFAs cy17:0 (F3,27 = 6.44**) and i17:0 (F3,27 = 3.76*), indicators of Gram-negative and Gram-positive bacteria, respectively, varied significantly between transformation systems and were higher in rainforest as compared to jungle rubber, oil palm, and rubber, on average by 25 and 16 %, respectively (Fig. 1a, b). In contrast to PLFAs, NLFA 16:1ω5c was at a minimum in rainforest and higher in jungle rubber, rubber, and oil palm; on average, in these systems, it exceeded that in rainforests by 59 % (F3,27 = 4.60*; Fig. 1c).

Relative amounts of PLFA cy17:0 (a), PLFA i17:0 (b), and amounts of AMF NLFA 16:1ω5c (c) in litter of rainforest (F), jungle rubber (J), rubber (R), and oil palm (O); means with s.d. (n = 8). Bars sharing the same letter do not differ significantly (Tukey’s HSD test, p < 0.05)
Microorganisms in soil
Soil basal respiration was similar in rainforest and jungle rubber but lower in rubber and oil palm, on average by 32 % (F3,27 = 7.34***; Fig. 2a). Soil microbial biomass did not vary significantly between transformation systems (overall mean 0.47 ± 0.14 mg Cmic*g−1 dw; F3,27 = 1.86ns). DFA based on PLFAs separated rainforest from oil palm (MD2 = 38.0***) and rubber (MD2 = 37.9***), but less from jungle rubber (MD2 = 21.1**; Wilk’s λ = 0.011, F102,78 = 8.41**). Concentration of total bacterial PLFAs varied significantly between transformation systems; it was similar in rainforest and oil palm and reduced in jungle rubber and rubber by an average of 17 % (F3,27 = 4.69**; Fig. 2b). Also, Gram-negative bacterial PLFAs varied significantly between transformation systems; as compared to rainforest, they were reduced by 52 % in jungle rubber and in both oil palm and rubber by on average 29 % (F3,27 = 4.47**; Fig. 2c). In contrast, the sum of Gram-positive bacterial PLFAs did not differ significantly between transformation systems and was on average 28.9 ± 5.53 % (F3,27 = 1.45ns). However, PLFA i17:0 was at a minimum in rainforest and higher in jungle rubber, rubber, and oil palm; on average, in these systems, it exceeded that in rainforest by 19 % (F3,27 = 6.10**; Fig. 2d). In contrast, total fungal PLFAs were at a maximum in jungle rubber and exceeded those in rainforest, rubber, and oil palm by an average of 29 % (F3,27 = 4.16*; Fig. 2e). NLFA 16:1ω5c was on average 12.0 ± 6.94 nmol g−1 dw and did not differ significantly between transformation systems (F3,27 = 2.53ns).

Basal respiration (a), total bacterial PLFAs (b), Gram-negative bacterial PLFAs (c), PLFA i17:0 (d), total fungal PLFAs (e) in soil of rainforest (F), jungle rubber (J), rubber (R), and oil palm (O); means with s.d. (n = 8). Bars sharing the same letter do not differ significantly (Tukey’s HSD test, p < 0.05)
Explanatory factors in litter
In the forward selection procedure of RDA, two of the five quantitative explanatory variables were significantly related to the community composition of microorganisms (P < 0.05; Fig. 3a). Together, the variables explained 58.8 % of the variation in microbial data, with the trace being significant (F = 2.90*). Litter pH accounted for 28.9 % of total variation in microbial data (F = 5.70**). Carbon concentration was the second environmental variable with significant explanatory power, accounting for an additional 16.8 % of the variation (F = 4.00*). RDA of microbial and environmental variables separated rainforest from each of the converted systems (Fig. 3a). Generally, rainforest was positioned at the end of axis 1 separated from the other systems by higher amount of litter and i17:0, cy17:0 bacterial PLFAs, while rubber and oil palm plantations were positioned at the opposite end and were associated with high pH and high AMF concentration. The second axis mainly separated jungle rubber from rubber and oil palm by high water content and high N and C concentration, and jungle rubber from rainforest by high AMF concentration.

RDA ordination of litter (a) and soil (b) microbial markers and environmental factors [water content (water), pH, carbon concentration (C), nitrogen concentration (N), amount of litter (litter), fatty acids PLFA cy17:0, an indicator of Gram-negative bacteria, PLFA i17:0, an indicator of Gram-positive bacteria, total fungal PLFAs (total fungi), total bacterial PLFAs (total bacteria), NLFA 16:1ω5c, an indicator for arbuscular mycorrhizal fungi (AMF), basal respiration (BR)] in rainforest, jungle rubber, rubber, and oil palm (included as passive variables, n = 8). For litter, axes 1 and 2 explained 47.5 and 8.8 % of the total variation of the microbial data, respective values for soil were 25.4 and 9.1 %. Dashed arrows represent microbial markers, solid filled arrows represent environmental factors significantly related to microbial markers, and empty arrows represent environmental factors that do not significantly contribute to the model
Explanatory factors in soil
In the forward selection procedure of RDA, one of the three explanatory variables was significantly related to the community composition of microorganisms (P < 0.05; Fig. 3b). Together, the variables explained 36.0 % of the variation in microbial data, with the trace being significant (F = 2.20*). Soil pH accounted for 24.1 % of total variation in microbial data (F = 4.40*). Similar to the litter layer, RDA of microbial and environmental variables separated rainforest from each of the converted systems (Fig. 3b). However, the associations of the transformation systems with environmental and microbial variables were more complex than in the litter layer. Generally, rainforest was positioned at the end of axis 1 separated from each of the conserved systems by high concentration of total bacterial PLFAs, by high C and N concentrations, and high basal respiration, while rubber and oil plantations were positioned at the opposite end and were associated with high pH and PLFA i17:0 concentration. Further, jungle rubber was separated from rainforest and each of the transformed systems by high fungal PLFAs.
Discussion
Impact of rainforest transformation on microorganisms in litter and soil
Despite more pronounced changes in the structure of the litter layer as compared to the soil after conversion of rainforests into rubber and oil palm plantations, changes in microbial community composition in soil exceeded those in litter contrasting our hypothesis (1). Of the nine microbial parameters studied, only three were significantly affected by rainforest conversion in litter and five in soil. In litter, only two bacterial PLFAs and the NLFA AMF marker significantly responded to rainforest conversion, whereas in soil, total and especially Gram-negative bacterial and fungal marker PLFAs were significantly affected. The more pronounced response of AMF in the litter layer as compared to the soil is surprising, since AMF are associated with roots and are expected to dominate in low organic matter soils (Allen et al. 1995). Notably, at our study site, Acrisols predominate, i.e., soils that do not hold large amounts of nutrients. Therefore, AMF may be forced to exploit nutrients in litter. Indeed, in rubber and oil palm plantations, fertilizer is applied on the soil surface, and the nutrients applied are likely to be captured by litter microorganisms (Lummer et al. 2012; Geisseler and Scow 2014; Lemanski and Scheu 2014). Therefore, nutrients in litter and litter decomposer microorganisms are likely to form an important pool of nutrients being captured by AMF (Aristizabal 2008; Nuccio et al. 2013). Similar to the present study, earlier studies in tropical forest ecosystems also found high concentrations of AMF in the litter layer (Krashevska et al. 2008, 2014), but colonization of litter by AMF is rarely studied (Posada et al. 2012).
For identifying factors responsible for changes in microbial community composition with rainforest conversion, we measured chemical and physical litter and soil characteristics. Rainforest conversion strongly reduced the amount of litter in the litter layer, litter C and N concentrations, and water content, but increased litter pH and C-to-N ratio. Similar to litter, pH in soil increased, and C and N concentrations decreased; however, changes in soil were less pronounced than in litter, suggesting that the impact of forest conversion on abiotic factors is more pronounced in litter than in soil. Generally, the results are in line with earlier studies reporting low water content, acidity, and C concentration in rubber and oil palm plantations (Li et al. 2012; Lee-Cruz et al. 2013; but see Templer et al. 2005). However, as indicated by redundancy analysis, only some of these changes contributed to changes in soil microbial community composition with pH and C concentration being most important. The strong impact of soil pH on composition of litter and soil microbial communities is consistent with earlier studies in tropical and temperate ecosystems indicating that pH is among the most important driver for microbial community composition (Bååth and Anderson 2003; Jesus et al. 2009; Tripathi et al. 2012).
As indicated by RDA, the abiotic factors studied explained a large fraction of the variability in microbial community composition, but this more pronounced in litter (59 %) than in soil (36 %), suggesting that, as compared to litter, soil is more intensively buffered against changes in abiotic factors associated with land transformation.
Differential response of microbial groups
We hypothesized that, as compared to bacteria, fungi are more heavily affected by conversion of rainforest into production systems. In contrast to this hypothesis, fatty acid analysis suggested that only certain bacterial groups, but not saprotrophic fungi and AMF, are detrimentally affected by rainforest conversion. Soils of converted systems had lower concentration of Gram-negative bacteria, whereas Gram-positive bacteria (based on PLFA i17:0) were more abundant in oil palm plantations than in rainforest soil. Similar results have been found in Malaysian Borneo (Tripathi et al. 2012; Lee-Cruz et al. 2013), where forest soils had lower pH than in oil palm plantations and this favored the dominance of Gram-negative bacteria, which, compared to Gram-positive bacteria and AMF, are assumed to be less sensitive to low pH conditions (Bååth and Anderson 2003). Increased concentrations of AMF at higher pH in converted systems in litter and (slightly) in soil suggests that low pH limits AMF in rainforests. On the other hand, increased pH presumably is a key factor favoring nutrient capture in rubber and oil palm production systems in particular in heavily weathered Acrisols (cf. Feldmann et al. 2000; Galindo-Castañeda and Romero 2013; Damayanti et al. 2015). Limited agricultural productivity in Acrisols is presumed to be due to high Al and Mn, and low nutrient concentrations, in particular P; presumably, AMF is of paramount importance for mitigating the negative effects of these conditions (Von Uexkull 1986; Smith and Read 2008; Damayanti et al. 2015).
Surprisingly, fungal PLFAs in soil (as percentages of total) were lower in rainforest as compared to jungle rubber and rubber. Low concentrations of fungi in rainforest soils were associated by high concentrations of bacteria, suggesting that the latter are more effective in exploiting resources in rainforest as compared to the studied production systems. High water content in litter and more developed litter layer in rainforests as compared to converted production systems likely contributed to the increased dominance of bacteria known to be more sensitive to water stress than fungi (Manzoni et al. 2012). Fertilizer addition to converted systems, on the other hand, likely favored fungi at the expense of bacteria as shown for tropical forests in Panama and Ecuador (Kerekes et al. 2013; Krashevska et al. 2014). Moreover, oil palm and rubber leaves contain high concentrations of Fe, Al, Zn, and Cd (V. Krashevska, unpubl. data), and oil palm roots also are richer in Al than roots in other land use systems (J. Sahner, unpubl. data), and this also may have favored fungi at the expense of bacteria (Frostegård et al. 1993).
Overall, these results suggest that the composition of microbial communities sensitively responds to rainforest conversion, with the response being more pronounced in soil as compared to litter. However, unexpectedly, the response of AMF was more pronounced in litter than in soil. Further, as indicated by marker PLFAs, only some bacteria groups were detrimentally affected by rainforest conversion, and saprotrophic fungi and AMF even increased in converted production systems. Apart from the significant changes in microbial community composition, results of the present study suggest that microbial communities in litter and soil are remarkably resistant to conversion of rainforest into agricultural production systems such as rubber and oil palm, with the resistance of litter exceeding that of soil microbial communities. Further studies on the susceptibility of microorganisms are needed for understanding how land-use changes and management practices in converted production systems impact decomposer community composition and thereby ecosystem functioning, such as nutrient cycling and decomposition of plant residues. Currently, at our field sites, two experiments are on the way to explore relationships between the composition of the decomposer system and the functioning of tropical rainforests and rubber and oil palm production systems.
References
Allen E, Allen M, Helm D, Trappe J, Molina R, Rincon E (1995) Patterns and regulation of mycorrhizal plant and fungal diversity. Plant Soil 170:47–62
Anderson JPE, Domsch KH (1978) A physiological method for the quantitative measurement of microbial biomass in soils. Soil Biol Biochem 10:215–221
Aristizabal C (2008) Arbuscular mycorrhizal fungi enhance the acquisition of mineral nutrients from leaf litter by Morella cerifera. PhD, University of Miami, Coral. Open Access Dissertations, paper 94
Bååth E, Anderson T-H (2003) Comparison of soil fungal/bacterial ratios in a pH gradient using physiological and PLFA-based techniques. Soil Biol Biochem 35:955–963
Baldrian P (2014) Distribution of extracellular enzymes in soils: spatial heterogeneity and determining factors at various scales. Soil Sci Soc Am J 78:11
Barnes AD, Jochum M, Mumme S, Haneda NF, Farajallah A, Widarto TH, Brose U (2014) Consequences of tropical land use for multitrophic biodiversity and ecosystem functioning. Nat Commun 5:5351
Borneman J, Triplett E (1997) Molecular microbial diversity in soils from eastern Amazonia: evidence for unusual microorganisms and microbial population shifts associated with deforestation. Appl Environ Microbiol 63:2647–2653
Brussaard L, Behan-Pelletier VM, Bignell DE, Brown VK, Didden W, Folgarait P, Fragoso C, Wall Freckman D, Gupta VVSR, Hattori T, Hawksworth DL, Klopatek C, Lavelle P, Malloch DW, Rusek J, Söderström B, Tiedje JM, Ross AV (1997) Biodiversity and ecosystem functioning in soil. Ambio 26:563–570
Carney KM, Matson PA, Bohannan BJM (2004) Diversity and composition of tropical soil nitrifiers across a plant diversity gradient and among land-use types. Ecol Lett 7:684–694
Chapin S, Matson PA, Vitousek P (2012) Principles of terrestrial ecosystem ecology. Springer, New York
Cole LES, Bhagwat SA, Willis KJ (2014) Recovery and resilience of tropical forests after disturbance. Nat Commun 5:3906
Damayanti ND, Rini MV, Evizal R (2015) Growth response of oil palm seedling (Elaeis guineensis Jacq.) to give five types of arbuscular mycorrhizal fungi on two level NPK fertilize. J Penelitian Pertanian Terapan 15:33–40, (in Indonesian) ISSN 1410-5020
Feldmann F, da Silva Junior JP, Idczak E, Lieberei R (2000) AMF spore community composition at natural and agricultural sites in Central Amazonia - a long term study. Neotropical Ecosyst Hamburg, 669–682
Fierer N, Breitbart M, Nulton J, Salamon P, Lozupone C, Jones R, Robeson M, Edwards RA, Felts B, Rayhawk S, Knight R, Rohwer F, Jackson RB (2007) Metagenomic and small-subunit rRNA analyses reveal the genetic diversity of bacteria, archaea, fungi, and viruses in soil. Appl Environ Microbiol 73:7059–7066
Fitzherbert EB, Struebig MJ, Morel A, Morel A, Danielsen F, Brühl CA, Donald PF, Phalan B (2008) How will oil palm expansion affect biodiversity? Trends Ecol Evol 23:538–545
Frostegård A, Bååth E (1996) The use of phospholipid fatty acid analysis to estimate bacterial and fungal biomass in soil. Biol Fertil Soils 22:59–65
Frostegård A, Tunlid A, Bååth E (1993) Phospholipid fatty acid composition, biomass, and activity of microbial communities from two soil types experimentally exposed to different heavy metals. Appl Environ Microbiol 59:3605–3617
Frostegård Å, Tunlid A, Bååth E (2011) Use and misuse of PLFA measurements in soils. Soil Biol Biochem 43:1621–1625
Galindo-Castañeda T, Romero HM (2013) Mycorrhization in oil palm (Elaeis guineensis and E. oleifera x E. guineensis) in the pre-nursery stage. Agron Colomb 31:95–102, ISSN 0120-9965
García-Orenes F, Morugán-Coronado A, Zornoza R, Scow K (2013) Changes in soil microbial community structure influenced by agricultural management practices in a mediterranean agro-ecosystem. PLoS One 8, e80522
Gatto M, Wollni M, Qaim M (2015) Oil palm boom and land-use dynamics in Indonesia: the role of policies and socioeconomic factors. Land Use Policy 46:292–303
Geisseler D, Scow KM (2014) Long-term effects of mineral fertilizers on soil microorganisms – a review. Soil Biol Biochem 75:54–63
Giam X, Ng TH, Yap VB, Tan HTW (2010) The extent of undiscovered species in Southeast Asia. Biodivers Conserv 19:943–954
Gibbs HK, Ruesch AS, Achard F, Clayton MK, Holmgren P, Ramankutty N, Foley JA (2010) Tropical forests were the primary sources of new agricultural land in the 1980s and 1990s. Proc Natl Acad Sci U S A 107:16732–16737
Guillaume T, Muhammad D, Kuzyakov Y (2015) Losses of soil carbon by converting tropical forest to plantations: erosion and decomposition estimated by δ13C. Glob Chang Biol. doi:10.1111/gcb.12907
Jesus EC, Marsh TL, Tiedje JM, Moreira FM (2009) Changes in land use alter the structure of bacterial communities in Western Amazon soils. ISME J 3:1004–1011
Joergensen RG, Wichern F (2008) Quantitative assessment of the fungal contribution to microbial tissue in soil. Soil Biol Biochem 40:2977–2991
Kerekes J, Kaspari M, Stevenson B, Nilsson RH, Hartmann M, Amend A, Bruns TD (2013) Nutrient enrichment increased species richness of leaf litter fungal assemblages in a tropical forest. Mol Ecol 22:2827–2838
Koh LP, Ghazoul J (2010) Spatially explicit scenario analysis for reconciling agricultural expansion, forest protection, and carbon conservation in Indonesia. Proc Natl Acad Sci U S A 107:11140–11144
Koh LP, Miettinen J, Liew SC, Ghazoul J (2011) Remotely sensed evidence of tropical peatland conversion to oil palm. Proc Natl Acad Sci U S A 108:5127–51232
Krashevska V, Bonkowski M, Maraun M, Ruess L, Kandeler E, Scheu S (2008) Microorganisms as driving factors for the community structure of testate amoebae along an altitudinal transect in tropical mountain rain forests. Soil Biol Biochem 40:2427–2433
Krashevska V, Maraun M, Ruess L, Scheu S (2010) Carbon and nutrient limitation of soil microorganisms and microbial grazers in a tropical montane rain forest. Oikos 119:1020–1028
Krashevska V, Sandmann D, Maraun M, Scheu S (2012) Consequences of exclusion of precipitation on microorganisms and microbial consumers in montane tropical rainforests. Oecologia 170:1067–1076
Krashevska V, Sandmann D, Maraun M, Scheu S (2014) Moderate changes in nutrient input alter tropical microbial and protist communities and belowground linkages. ISME J 8:1126–1134
Lee-Cruz L, Edwards DP, Tripathi BM, Adams JM (2013) Impact of logging and forest conversion to oil palm plantations on soil bacterial communities in Borneo. Appl Environ Microbiol 79:7290–7297
Lekberg Y, Rosendahl S, Michelsen A, Olsson PA (2012) Seasonal carbon allocation to arbuscular mycorrhizal fungi assessed by microscopic examination, stable isotope probing and fatty acid analysis. Plant Soil 368:547–555
Lemanski K, Scheu S (2014) Incorporation of 13C labelled glucose into soil microorganisms of grassland: effects of fertilizer addition and plant functional group composition. Soil Biol Biochem 69:38–45
Li H, Ma Y, Liu W, Liu W (2012) Soil changes induced by rubber and tea plantation establishment: comparison with tropical rain forest soil in Xishuangbanna, SW China. Environ Manag 50:837–848
Lummer D, Scheu S, Butenschoen O (2012) Connecting litter quality, microbial community and nitrogen transfer mechanisms in decomposing litter mixtures. Oikos 121:1649–1655
Manzoni S, Schimel JP, Porporato A (2012) Responses of soil microbial communities to water stress: results from a meta-analysis. Ecology 93:930–938
Marimin M, Darmawan MA, Islam Fajar Putra MP, Machfund, Wiguna B (2014) Value chain analysis for green productivity improvement in the natural rubber supply chain: a case study. J Clean Prod 85:201–211
Miettinen J, Wang J, Hooijer A, Liew S (2013) Peatland conversion and degradation processes in insular southeast Asia: a case study in Jambi, Indonesia. Land Degrad Dev 341:334–341
Murty D, Kirschbaum M, Mcmurtrie R, Mcgilvray H (2002) Does conversion of forest to agricultural land change soil carbon and nitrogen? A review of the literature. Glob Chang Biol 8:105–123
Naeem S, Duffy JE, Zavaleta E (2012) The functions of biological diversity in an age of extinction. Science 336:1401–1406
Ngosong C, Gabriel E, Ruess L (2012) Use of the signature fatty acid 16:1ω5 as a tool to determine the distribution of arbuscular mycorrhizal fungi in soil. J Lipids 2012:236807
Nuccio EE, Hodge A, Pett-Ridge J, Herman DJ, Weber PK, Firestone MK (2013) An arbuscular mycorrhizal fungus significantly modifies the soil bacterial community and nitrogen cycling during litter decomposition. Environ Microbiol 15:1870–1881
Posada RH, Madriñan S, Rivera EL (2012) Relationships between the litter colonization by saprotrophic and arbuscular mycorrhizal fungi with depth in a tropical forest. Fungal Biol 116:747–755
Ramsey PW, Rillig MC, Feris KP, Holben WE, Gannon JE (2006) Choice of methods for soil microbial community analysis: PLFA maximizes power compared to CLPP and PCR-based approaches. Pedobiologia 50:275–280
Rasche F, Cadisch G (2013) The molecular microbial perspective of organic matter turnover and nutrient cycling in tropical agroecosystems - what do we know? Biol Fertil Soils 49:251–262
Rodrigues JLM, Pellizari VH, Mueller R, Baek K, Jesus EDC, Paula FS, Mirza B, Hamaoui GS, Tsai SM, Feigl B, Tiedje JM, Bohannan BJM, Nüsslein K (2013) Conversion of the Amazon rainforest to agriculture results in biotic homogenization of soil bacterial communities. Proc Natl Acad Sci U S A 110:988–993
Roesch LFW, Fulthorpe RR, Riva A, Casella G, Hadwin AKM, Kent AD, Daroub SH, Camargo FAO, Farmerie WG, Triplett EW (2007) Pyrosequencing enumerates and contrasts soil microbial diversity. ISME J 1:283–290
Rousk J, Bååth E, Brookes PC, Lauber CL, Lozupone C, Caporaso JG, Knight R, Fierer N (2010) Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J 4:1340–1351
Ruess L, Chamberlain PM (2010) The fat that matters: soil food web analysis using fatty acids and their carbon stable isotope signature. Soil Biol Biochem 42:1898–1910
Sala OE, Chapin FS, Armesto JJ, Berlow E, Bloomfield J, Dirzo R, Huber-Sanwald E, Huenneke LF, Jackson RB, Kinzig A, Leemans R, Lodge DM, Mooney HA, Oesterheld M, Poff NL, Sykes MT, Walker BH, Walker M, Wall DH (2000) Global biodiversity scenarios for the year 2100. Science 287:1770–1774
Scheiner SM, Gurevitch J (2001) Design and analysis of ecological experiments, 2nd edn. Oxford University Press, Oxford
Scheu S (1992) Automated measurement of the respiratory response of soil microcompartments: active microbial biomass in earthworm faeces. Soil Biol Biochem 24:1113–1118
Smith SE, Read DJ (2008) Mycorrhizal symbiosis, 3rd edn. Academic press, New York
Sodhi NS, Koh LP, Brook BW, Ng PKL (2004) Southeast Asian biodiversity: an impending disaster. Trends Ecol Evol 19:654–660
Sodhi NS, Koh LP, Clements R, Wanger TC, Hill JK, Hamer KC, Clough Y, Tscharntke T, Posa MRC, Lee TM (2010) Conserving Southeast Asian forest biodiversity in human-modified landscapes. Biol Conserv 143:2375–2384
Strickland MS, Rousk J (2010) Considering fungal:bacterial dominance in soils – methods, controls and ecosystem implications. Soil Biol Biochem 42:1385–1395
Templer PH, Groffman PM, Flecker AS, Power AG (2005) Land use change and soil nutrient transformations in the Los Haitises region of the Dominican Republic. Soil Biol Biochem 37:215–225
Ter Braak CJF, Šmilauer P (2012) Canoco reference manual and user’s guide: software for ordination, version 5.0. Microcomputer Power, Ithaca, USA. 496
Tischer A, Potthast K, Hamer U (2014) Land-use and soil depth affect resource and microbial stoichiometry in a tropical mountain rainforest region of southern Ecuador. Oecologia 175:375–393
Tripathi BM, Kim M, Singh D, Lee-Cruz L, Lai-Hoe A, Ainuddin AN, Go R, Rahim RA, Husni MHA, Chun J, Adams JM (2012) Tropical soil bacterial communities in Malaysia: pH dominates in the equatorial tropics too. Microb Ecol 64:474–484
Van der Wal A, van Veen JA, Smant W, Boschker HTS, Bloem J, Kardol P, van der Putten WH, de Boer W (2006) Fungal biomass development in a chronosequence of land abandonment. Soil Biol Biochem 38:51–60
Von Uexkull HR (1986) Efficient fertilizer use in acid upland soils of the humid tropics. FAO fertilizer and plant nutrition bulletin (FAO), no 10. Food & Agriculture Org, Rome, p 69
Whitman WB, Coleman DC, Wiebe WJ (1998) Prokaryotes: the unseen majority. Proc Natl Acad Sci U S A 95:6578–6583
Wilcove DS, Koh LP (2010) Addressing the threats to biodiversity from oil-palm agriculture. Biodivers Conserv 19:999–1007
Wilcove DS, Giam X, Edwards DP, Fisher B, Koh LP (2013) Navjot’s nightmare revisited: logging, agriculture, and biodiversity in Southeast Asia. Trends Ecol Evol 28:531–540
Zelles L (1997) Phospholipid fatty acid profiles in selected members of soil microbial communities. Chemosphere 35:275–294
Zelles L (1999) Fatty acid patterns of phospholipids and lipopolysaccharides in the characterisation of microbial communities in soil: a review. Biol Fertil Soils 29:111–129
Acknowledgments
We thank the State Ministry of Research and Technology of Indonesia (RISTEK) for the research permit and the Indonesian Institute of Sciences (LIPI) and Ministry of Forestry (PHKA) for the collection permit. We thank many Indonesian organizations for granting us access to the sampling plots and use of their properties. We thank the many colleagues and helpers for their support in the field and the laboratory assistance. We thank Claudia Dislich for sharing literature database, Michael Euler for core plot farmer survey database. Financial support was provided by the German Research Foundation (DFG) in the framework of the collaborative German-Indonesian research project CRC990 (EFForTS).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Krashevska, V., Klarner, B., Widyastuti, R. et al. Impact of tropical lowland rainforest conversion into rubber and oil palm plantations on soil microbial communities. Biol Fertil Soils 51, 697–705 (2015). https://doi.org/10.1007/s00374-015-1021-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00374-015-1021-4