
Abstract
Soil labile organic carbon (C) oxidation drives the flux of carbon dioxide (CO2) between soils and the atmosphere. However, the impact of grazing management and the contribution soil aggregate size classes (ASCs) to labile organic C from grassland soils is unclear. We evaluated the effects of grazing intensity and soil ASC on the soil labile organic C, including CO2 production, microbial biomass C, and dissolved organic C and nitrogen (N) mineralization in topsoils (0–10 cm) in Inner Mongolia, Northern China. Soil samples were separated into ASCs of 0–630 μm [fine ASC (fASC)], 630–2000 μm [medium ASC (mASC)] and >2000 μm [coarse ASC (cASC)]. The results showed that heavy grazing (HG) and continuous grazing (CG) increased soil labile organic C significantly compared to an ungrazed site since 1999 (UG99) and an ungrazed site since 1979 (UG79). For winter grazing site (WG), no significant differences were found. CO2 production was highest in cASC, while lowest in fASC. Microbial biomass C and dissolved organic C showed the highest values in mASC and were significantly lower in fASC. Grazing increased N mineralization in bulk soils, while it exhibited complex effects in the three ASCs. The results suggest that the rate of C mineralization was related to the rate of N accumulation. To reduce CO2 emission and nutrient loss, and to improve soil quality and productivity, a grazing system with moderate intensity is suggested.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Soil labile organic C is a soil fraction with turnover time of less than a few years (even less than weeks) as compared to recalcitrant C with a turnover time of several thousand years (Parton et al. 1987; Schimel et al. 1985). Soil labile organic C as the most active fraction of soil organic C can be readily influenced by disturbance and management (Harison et al. 1993). Therefore, soil labile organic carbon oxidation drives the flux of carbon dioxide (CO2) between soils and the atmosphere (Zou et al. 2005) and makes a greater contribution to nutrient cycling than stable soil organic C (Whalen et al. 2000). As most studies are focusing on total organic C storage and sequestration, mineralization of soil labile organic C is not well understood, particularly for typical grassland soils.
The Inner Mongolian grasslands in North China, covering approximately 110 × 106 ha, are of denotative ecological and economical importance (He et al. 2011). Most grasslands in China have undergone some degree of degradation or desertification, following the rapid expansion of the livestock industry after 1980 (Tong et al. 2003). Grazing is one of the most important factors that could change the soil C stock in grassland ecosystems (Cui et al. 2005), which influences organic matter input and associated soil properties (Steffens et al. 2009; Wiesmeier et al. 2009). Previous studies have shown that heavy grazing (HG) could reduce soil organic C contents and stocks associated with higher bulk densities in topsoil in semiarid steppes in Inner Mongolia (Cui et al. 2005; He et al. 2011; Steffens et al. 2008; Wiesmeier et al. 2011). However, light grazing pressure for 20 years caused no significant decrease of soil organic C contents (Cui et al. 2005). The influence of grazing on soil C turnover in grasslands is complex and difficult to predict. Until now, only few studies have been conducted on the impact of grazing on soil labile organic C.
Soil organic carbon protection mechanisms are intimately tied to the processes of aggregate turnover and stabilization at multiple scales (Steffens et al. 2009). The deposition and transformation of organic matter plays a major role in aggregate stabilization, and there are strong feedbacks between aggregate turnover and soil organic C dynamics (Jastrow et al. 2007; Lützow et al. 2006). Differences in turnover rates of soil organic matter fractions may be due to physical protection of organic matter within soil aggregates as well as chemical protection from humification (Cambardella and Elliott 1993). The fractions of soil organic matter that turn over rapidly are believed to make a greater contribution to nutrient cycling than fractions that turn over slowly because they provide a more readily accessible source of energy for the saprotrophic soil organisms responsible for nutrient cycling (Janzen et al. 1992). It is becoming increasingly important to determine not only how land management practices affect the retention or loss of these fractions of SOM but also how they affect nutrient cycling from soil organic matter fractions. Improved understanding of these processes will provide valuable information for maintaining or implementing environmentally sustainable land management practices in agricultural and forest soils (Whalen et al. 2000).
Moreover, the rate of changes in soil C over time in the processes of biotic community development is tightly coupled with soil N mechanisms (Knops and Tilman 2000). The effects on soil N mineralization have been intensively studied. However, variable grazing effects have been reported with both increasing (Groffman et al. 1993; Le Roux et al. 2003) and decreasing (Bardgett and Wardle 2003; Biondini et al. 1998) N mineralization. Furthermore, N mineralization of soil aggregate size classes (ASCs) is not well understood, especially under the influence of grazing (He et al. 2011). Estimating N mineralization would be helpful to understand C mineralization of ASCs in semiarid grasslands affected by grazing management.
The objectives of this research were: (1) to evaluate the influence of different grazing intensities on soil labile organic C and N mineralization, (2) to estimate the effect of different ASCs on soil labile organic C and N mineralization, (3) to evaluate the interactions of soil labile organic C and N mineralization.
Materials and methods
Study area
The study was performed near the Inner Mongolia Grassland Ecosystem Research Station (IMGERS, administered by the Chinese Academy of Sciences; 43°38′N, 116°42′E), which is located in the Xilin River Area, Inner Mongolia Autonomous Region, north China, with 78.8 million hectares of natural grasslands, a typical steppe ecosystem biogeographically belonging to the Eurasia Steppe region. The growing season runs from early April to late September. The mean annual temperature in the study area is 0.3°C with mean monthly temperatures ranging from −21.6°C in January to 19.0°C in July. The mean annual precipitation is 346.1 mm with 60–80% falling during the growing season from May to August and approximately 10% of which falls as snow. The soil is classified as Calcic Chernozem (IUSS Working Group WRB, 2006).
Plot description and sampling design
Five experimental sites with different grazing intensities were chosen in this study. The whole area is grazed by herds that are composed of 70–90% sheep and 10–30% goats. In 1979, one plot (24 ha) was fenced and excluded from grazing [ungrazed site since 1979 (UG79)]. The whole experimental area was grazed before with low intensity. After 20 years of moderate grazing, two plots were fenced; one was completely excluded from grazing [25 ha; ungrazed site since 1999 (UG99)], the other is still grazed during winter [34 ha; winter grazing (WG)], equivalent to a grazing intensity of 0.5 sheep units ha-1 year-1 (1 sheep unit = 1 ewe and 1 lamb). Another site (24 ha) was grazed during the whole year [continuous grazing (CG)], equivalent to a grazing intensity of 1.2 sheep units ha-1 year-1. An unfenced site was grazed with approximately 5–6 sheep units ha-1 (HG) during the vegetation period that is located approximately 2 km away from the other sites. At all experimental sites, three randomly located soil pits were sampled at the upper 10 cm of the topsoil using a steel cylinder with a volume of 100 cm³ to determine soil properties.
Physical fractionation
The upper 10 cm of one pit in each of the five plots were used for physical fractionation (4 kg for each sample). The air-dried soil was dry-sieved gently by hand to three ASCs: 2,000–6,300 mm, 630–2,000 mm, and < 630 mm (referred to as coarse ASC (cASC), medium ASC (mASC), and fASC; Steffens et al. 2009).
Soil incubations
C and N mineralization of the ASCs were determined after soil incubation at 25 ± 0.5°C for 30 days. Dry soil (50 g) was adjusted to 60% field moisture capacity in100 ml glass bottle with an open mouth, which was put in 1 l glass jars with a septum to keep it gas-tight. ASCs were inoculated with 1 ml dilute suspensions (soil to solution ratio 1:10) of the corresponding fresh whole soil. A thin film of water was put into the bottom of the jars to prevent the soil drying.
Determination of soil properties
In order to characterize soil labile organic C mineralization, CO2 production after incubation, microbial biomass C, and dissolved organic C were determined which were related to soil organic C contents of the samples. Additionally, ammonium (NH +4 ) and nitrate (NO -3 ) contents were analyzed which were related to total N contents of the samples. All the analyses were performed in triplicate.
Soil organic C and total N were determined in duplicate by dry combustion on a Vario Max CNS elemental analyser (Elementar, Hanau, Germany). The measured C concentrations of the samples that were free of carbonate represent the soil organic C concentration. Samples that contained CaCO3 were heated to 500°C for 4 h to remove organic C, and the concentration of inorganic C of the residual material was determined by dry combustion. The content of inorganic C was subtracted from the C concentration of the untreated material and represents the soil organic C content.
CO2 production after incubation was determined by incubating the soil fractions in 1 l airtight jars with a vial of 20 ml 0.1 M NaOH. The NaOH solution was removed and replaced with fresh solution during sampling. The NaOH solution was sampled on 2th, 5th, 8th, 11th, 14th, 17th, 23th, 30th day after the incubation started. At the sampling dates, the captured CO2 was determined by titration with 0.1 M HCl (Zibilske 1994) after precipitation of the carbonate with excess BaCI2. The CO2 produced after the incubation was used to calculate the C mineralization rate.
Microbial biomass C was determined using the fumigation–extraction (FE) method (Vance et al. 1987; Dannenmann et al. 2006). The sample (10 g) was immediately extracted with 30 ml 0.5 M K2SO4 for 60 min on a rotary shaker at 150 rpm. The second sample was fumigated under chloroform vapor for 24 h in a desiccator and then extracted as described above. Extracts were frozen under −20°C and analyzed within 1 month for dissolved organic C (Dimatec Analysentechnik, Essen, Germany). Correction factors (0.379 for microbial biomass C, Vance et al. 1987) were applied to the difference in total dissolved organic C between untreated and fumigated subsamples to estimate microbial biomass C.
After rewetting the soil fractions, 10 g subsample was immediately (t 1) extracted as described above in order to analyze NH +4 and NO -3 concentrations. The other subsample were incubated in the glass jar for 1 month and thereafter extracted with 0.5 M K2SO (t 2). The extract solution was immediately frozen until the colorimetrical determination of NH +4 and NO –3 concentrations (FIAstar 5000 Analyzer, Foss Tecator, Denmark). The difference of NH +4 , NO -3 , and inorganic N between t 2 and t 1 were the net ammonification, nitrification, and mineralization, respectively.
Statistical methods
All statistical analyses were carried out using SPSS 15.0 (SPSS, Chicago, IL). To test the significance of grazing and soil aggregate size effects on the examined parameters, a two-way analysis of variance (ANOVA) was applied.
Results
Labile organic C
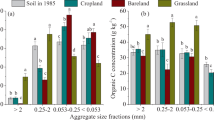
CO2 production after incubation ranged from 1 to 12 mg g-1 between all grazing intensities and ASCs (Fig. 1). HG and CG showed a much higher CO2 production than WG, UG99, and UG79. HG was significantly (P < 0.05) higher than CG, while there were no differences among WG, UG99, and UG79. Grazing intensity had a consistent effect on CO2 production across all ASCs. CO2 production was highest for cASC, while fine ASC (fASC) showed the lowest value (P < 0.05). CO2 production of the bulk soil was 3.75 ± 1.60 mg g-1, which was between mASC and fASC.

CO2 production (a), microbial biomass C (MBC, b), and dissolved organic C (c) influenced by soil aggregate size and grazing
Microbial biomass C and dissolved organic C showed similar patterns after incubation. Both microbial biomass C and dissolved organic C in CG and HG were considerably higher compared to WG, UG99, and UG79 (Fig. 1). CG showed significantly (P < 0.01) higher value than HG both for microbial biomass C and dissolved organic C. No differences were found between WG, UG99, and UG79. Grazing intensities had a consistent effect on microbial biomass C and dissolved organic C across all ASCs (Fig. 1). Among the three ASCs, microbial biomass C and dissolved organic C of mASC showed the highest values, microbial biomass C of fASC was the lowest (P < 0.05), microbial biomass C of cASC was the lowest. Aggregate size, grazing, and their interaction effect all influenced CO2 production, microbial biomass C, and dissolved organic C significantly (Table 1 and Fig. 1, two-way ANOVA, P < 0.0001).
N mineralization
Land-use styles, soil aggregate sizes, and their interaction effect had significant effects on ammonification, nitrification, and mineralization (Table 2, P < 0.0001). However, there were no consistent patterns across all land-use styles and aggregate sizes.
For bulk soil, net ammonification (Fig. 2) of all experimental sites was negative. Net ammonification of CG was significantly lower (P < 0.05) compared to all other grazing intensities. Furthermore, net ammonification of HG and WG was significantly higher (P < 0.05) than that of UG99 and UG79. For mASC, all net ammonification rates were positive. For cASC and fASC, net ammonification of WG and UG79 was positive while that of UG99, CG, and HG was negative.

Ammonification (a), nitrification (b), and mineralization (c) affected by soil aggregate size and grazing
Net nitrification revealed the highest values for HG (Fig. 2). Net nitrification in UG79 was the lowest between all experimental sites. Winter grazing showed a high net nitrification for bulk soil, but for three ASCs, WG had the lowest nitrification. In general, bulk soil showed higher nitrification rates than the ASCs. fASC was significantly higher than mASC, but cASC had no difference with them.
Grazing increased net N mineralization significantly (Fig. 2 and Table 2), especially in bulk soils. Net N mineralization of bulk soil was the highest among all ASCs, but no difference was found among cASC, mASC, and fASC.
Inorganic N
Both grazing intensities and ASC significantly influenced NH +4 , NO -3 , and inorganic N concentrations Table 3. For bulk soils, only CG exhibited a significant effect on NH +4 concentration. There was no significant difference among HG, WG, UG99, and UG79 (Fig. 3a). There were also no clear patterns of grazing intensity effects across the three ASCs. However, the NH +4 concentrations in cASC, mASC, and fASC were much higher than that in bulk soil across all grazing intensities (Fig. 3a), while NO -3 and inorganic N concentrations in cASC, mASC, and fASC was much lower than that in bulk soil (Fig. 3b and c). Grazing significantly increased NO -3 and inorganic N in bulk soil (Fig. 3b and c).

Ammonium-N (a), NO -3 (b), and inorganic N (c) affected by soil aggregate size and grazing
Discussion
Effect of grazing on labile organic C
The results showed that CG and HG increased CO2 production, microbial biomass C, and dissolved organic C significantly across all ASCs and bulk soil, indicating overgrazing increased soil labile organic C in Inner Mongolia grassland. Our results correspond well with the fact that overgrazing increased C loss and decreased C storage (He et al. 2008, 2011; Ingram et al. 2008). However, the results also showed that WG exhibited no significant effect on soil labile organic C compared to UG99 and UG79, suggesting that moderately grazing would not increase C loss or even increased C storage and bacterial diversities (Milchunas and Lauenroth 1993; Schuman et al. 1999; Han et al. 2008; Zhou et al. 2010). Our results are inconsistent with the finding of a higher CO2 production in UG99 compared to WG during freezing–thawing cycles (Holst et al. 2008) and greater microbial and enzyme activities in ungrazed compared to grazed plots in semiarid Australia (Holt 1997). They attributed their findings to greater soil moisture and greater input of organic matter in ungrazed plots. Thus, the effects of grazing are complex because of the variations in climate, soil, landscape location, plant community type, and grazing management practices (Milchunas and Lauenroth 1993; Reeder and Schuman 2002).
Labile organic C of different soil aggregate sizes
Soil ASC had a consistent effect on CO2 production of all grazing intensities. CO2 production of cASC was highest, while fASC was lowest among the three ASCs, suggesting that C in cASC was most labile while it was stable in fASC. This is consistent with the findings that coarse aggregates are less stable and have faster turnover times than small aggregates (Six et al. 2004; Steffens et al. 2009). The CO2 release of cASC is high, because it contains more labile soil organic matter, and it is less protected against mineralization (Steffens et al. 2009).
For mASC, high values of microbial biomass C and dissolved organic C were found, whereas fASC revealed much lower amounts. This can be explained by a high microbial biomass in mASC due to a high efficient usage of SOM within this fraction (Sainju et al. 2009). Carbon mineralization of fASC was low, suggesting that aggregates protect the mineralization of soil organic C by reducing microbial access to the substrates that bind them (Elliott 1986; Six et al. 2000). Coarse aggregates had higher C and N concentrations than small aggregates because coarse aggregates are composed of microaggregates and organic binding agents (Elliott 1986), while aggregate-protected C and N pools are less labile than unprotected pools because protected pools are less exposed to microbial decay (Beare et al. 1994; Cambardella and Elliott 1993).
In WG, UG99, and UG79, ASC exhibited no significant effect on microbial biomass C and dissolved organic C, while there is a significant effect on CG and HG, indicating that overgrazing increased microbial biomass C and dissolved organic C in ASCs more than those in bulk soil. It also supports the findings that small aggregates are more stable, while coarse aggregates are more likely to be influenced by grazing (Cambardella and Elliott 1993; Six et al. 2004). To reduce CO2 emission, soil erosion, and nutrient loss, and to improve soil quality and productivity, the management practices should adopt moderate grazing to promote the fASC aggregation size (Sainju et al. 2009).
N mineralization and inorganic N of soil ASCs affected by grazing and its interaction with C mineralization
The effects of grazing intensity and soil ASC on net ammonification, nitrification, and N mineralization were complex. Grazing increased N mineralization significantly in bulk soils, which is consistent with other studies (Groffman et al. 1993; Le Roux et al. 2003; Wu et al. 2011a). However, grazing exhibited indefinite effects in the three soil ASCs. Most of them were not significant with the exceptions that net nitrification and mineralization of fASC in CG increased significantly compared to UG99 and UG79, and net nitrification in WG decreased significantly for the three ASCs. The inconsistent effect of bulk soil and soil ASC on N mineralization suggests the interactions of soil ASCs in bulk soil. Further research is needed in terms of N mineralization in soil ASCs, especially under the effect of grazing.
Generally, HG increased C and N mineralization in bulk soils that is consistent with the finding that HG decreases C and N sequestration (He et al. 2011), suggesting that the rate of C mineralization was related to the rate of N accumulation (Knops and Tilman 2000). Based on a 16 year grazing experiment, Zhou et al. (2010) found that intermediate grazing intensities by sheep increase soil bacterial diversities in an Inner Mongolian steppe, suggesting that microbial activities and their diversities may play an important role in moderating the effects of grazing on C and N turnover. As net rates of N mineralization do not at all reflect the dynamic of gross rates of N mineralization (Wu et al. 2011a, b), future studies should foucus on the linkage between gross N mineralization and C mineralization.
Conclusion
This study showed that heavy grazing (i.e., HG and CG) increased soil labile organic C significantly as compared to ungrazed sites, while moderate grazing (i.e., WG) exhibited no significant effect. CO2 production was highest in cASC while lowest in fASC. Microbial biomass C and dissolved organic C showed the highest values in mASC and were significantly lower in fASC. Grazing increased N mineralization in bulk soils while exhibited complex effects in the three ASCs. Moderate grazing exhibited no significant negative effects on C and N mineralization in bulk soils, which is consistent with the finding that moderate grazing increases C and N sequestration (He et al. 2011) and bacterial diversities (Zhou et al. 2010). Thus, we recommend moderate grazing as a proper way to protect C and N losses in semiarid grasslands.
References
Bardgett RD, Wardle DA (2003) Herbivore-mediated linkages between aboveground and belowground communities. Ecology 84:2258–2268
Beare MH, Hendrix PF, Coleman DC (1994) Water-stable aggregates and organic-matter fractions in conventional-tillage and no-tillage soils. Soil Sci Soc Am J 58:777–786
Biondini ME, Patton BD, Nyren PE (1998) Grazing intensity and ecosystem processes in a northern mixed-grass prairie, USA. Ecol Appl 8:469–479
Cambardella CA, Elliott ET (1993) Carbon and nitrogen distribution in aggregates from cultivated and native grassland soils. Soil Sci Soc Am J 57:1071–1076
Cui X, Wang Y, Niu H, Wu J, Wang S, Schnug E, Rogasik J, Fleckenstein J, Tang Y (2005) Effect of long-term grazing on soil organic carbon content in semiarid steppes in Inner Mongolia. Ecol Res 20:519–527
Dannenmann M, Gasche R, Ledebuhr A, Papen H (2006) Effects of forest management on soil N cycling in beech forests stocking on calcareous soils. Plant Soil 287:279–300
Elliott ET (1986) Aggregate structure and carbon, nitrogen, and phosphorus in native and cultivated soils. Soil Sci Soc Am J 50:627–633
Groffman PM, Rice CW, Tiedje JM (1993) Denitrification in a tallgrass prairie landscape. Ecology 74:855–862
Han G, Hao X, Zhao M, Wang M, Ellert BH, Walter W, Wang M (2008) Effect of grazing intensity on carbon and nitrogen in soil and vegetation in a meadow steppe in Inner Mongolia. Agr Ecosyst Environ 125:21–32
Harison KG, Broecker WS, Bonani G (1993) The effect of changing land use of soil radiocarbon. Science 262:725–726
He NP, Yu Q, Wu L, Wang YS, Han XG (2008) Carbon and nitrogen store and storage potential as affected by land-use in a Leymus chinensis grassland of northern China. Soil Biol Biochem 40:2952–2959
He NP, Zhang YH, Yu Q, Chen QS, Pan QM, Zhang GM, Han XG (2011) Grazing intensity impacts soil carbon and nitrogen storage of continental steppe. Ecosphere 2:art8
Holst J, Liu C, Yao Z, Brueggemann N, Zheng X, Giese M, Butterbach-Bahl K (2008) Fluxes of nitrous oxide, methane and carbon dioxide during freezing–thawing cycles in an Inner Mongolian steppe. Plant Soil 308:105–117
Holt JA (1997) Grazing pressure and soil carbon, microbial biomass and enzyme activities in semi-arid northeastern Australia. Appl Soil Ecol 5:143–149
Ingram LJ, Stahl PD, Schuman GE, Buyer JS, Vance GF, Ganjegunte GK, Welker JM, Derner JD (2008) Grazing impacts on soil carbon and microbial communities in a mixed-grass ecosystem. Soil Sci Soc Am J 72:939–948
Janzen HH, Campbell CA, Brandt SA, Lafond GP, Townleysmith L (1992) Light-fraction organic-matter in soils from long-term crop rotations. Soil Sci Soc Am J 56:1799–1806
Jastrow JD, Amonette JE, Bailey VL (2007) Mechanisms controlling soil carbon turnover and their potential application for enhancing carbon sequestration. Clim Chang 80:5–23
Knops JMH, Tilman D (2000) Dynamics of soil nitrogen and carbon accumulation for 61 years after agricultural abandonment. Ecology 81:88–98
Le Roux X, Bardy M, Loiseau P, Louault F (2003) Stimulation of soil nitrification and denitrification by grazing in grasslands: do changes in plant species composition matter? Oecologia 137:417–425
Lützow MV, Kögel-Knabner I, Ekschmitt K, Matzner E, Guggenberger G, Marschner B, Flessa H (2006) Stabilization of organic matter in temperate soils: mechanisms and their relevance under different soil conditions — a review. Eur J Soil Sci 57:426–445
Milchunas DG, Lauenroth WK (1993) Quantitative effects of grazing on vegetation and soils over a global range of environments. Ecol Monogr 63:327–366
Parton WJ, Schimel DS, Cole CV, Ojima DS (1987) Analysis of factors controlling soil organic-matter levels in great-plains grasslands. Soil Sci Soc Am J 51:1173–1179
Reeder JD, Schuman GE (2002) Influence of livestock grazing on C sequestration in semi-arid mixed-grass and short-grass rangelands. Environ Pollut 116:457–463
Sainju UM, Caesar-TonThat T, Jabro JD (2009) Carbon and nitrogen fractions in dryland soil aggregates affected by long-term tillage and cropping sequence. Soil Sci Soc Am J 73:1488–1495
Schimel DS, Coleman DC, Horton KA (1985) Soil organic matter dynamics in paired rangeland and cropland toposequences in North Dakota. Geoderma 36:201–214
Schuman GE, Reeder JD, Manley JT, Hart RH, Manley WA (1999) Impact of grazing management on the carbon and nitrogen balance of a mixed-grass rangeland. Ecol Appl 9:65–71
Six J, Paustian K, Elliott ET, Combrink C (2000) Soil structure and organic matter: I. Distribution of aggregate-size classes and aggregate-associated carbon. Soil Sci Soc Am J 64:681–689
Six J, Bossuyt H, Degryze S, Denef K (2004) A history of research on the link between (micro)aggregates, soil biota, and soil organic matter dynamics. Soil Till Res 79:7–31
Steffens M, Kolbl A, Totsche KU, Kogel-Knabner I (2008) Grazing effects on soil chemical and physical properties in a semiarid steppe of Inner Mongolia (PR China). Geoderma 143:63–72
Steffens M, Kolbl A, Kögel-Knabner I (2009) Alteration of soil organic matter pools and aggregation in semi-arid steppe topsoils as driven by organic matter input. Eur J Soil Sci 60:198–212
Tong C, Xi F, Yang J, Yong W (2003) Remote sensing monitoring on degraded steppe and determination of reasonable grazing intensity for the restoration of steppe in middle reach of Xilin river basin. Acta Prataculturae Sinica 12:78–83
Vance ED, Brookes PC, Jenkinson DS (1987) An extraction method for measuring soil microbial biomass C. Soil Biol Biochem 19:703–707
Whalen JK, Bottomley PJ, Myrold DD (2000) Carbon and nitrogen mineralization from light- and heavy-fraction additions to soil. Soil Biol Biochem 32:1345–1352
Wiesmeier M, Steffens M, Kölbl A, Kögel-Knabner I (2009) Degradation and small-scale spatial homogenization of topsoils in intensively grazed steppes of Northern China. Soil Till Res 104:299–310
Wiesmeier M, Barthold FK, Blank FB, Kögel-Knabner I (2011) Digital mapping of soil organic matter stocks using Random Forest modeling in a semi-arid steppe ecosystem. Plant Soil 340:7–24
Wu H, Dannenmann M, Fanselow N, Wolf B, Yao Z, Wu X, Brüggemann N, Zheng X, Han X, Dittert K, Butterbach-Bahl K (2011a) Feedback of grazing on gross rates of N mineralization and inorganic N partitioning in steppe soils of Inner Mongolia. Plant Soil 340:127–139
Wu H, Dannenmann M, Wolf MB, Han X, Zheng X, Butterbach-Bahl K (2011b) Seasonality of soil microbial nitrogen turnover in continental steppe soils of Inner Mongolia — insights from a full year dataset of gross and net nitrogen turnover. Ecosphere in press
Zhou X, Wang J, Hao Y, Wang Y (2010) Intermediate grazing intensities by sheep increase soil bacterial diversities in an Inner Mingolia steppe. Biol Fertil Soils 46:817–824
Zibilske M (1994) Carbon mineralization. In: Angle S, Weaver RW, Bottomley P, Bezdicek D, Smith S, Tabatabai A, Wollum A (eds) Methods of soil analysis microbiological and biochemical properties. Part 2, SSSA Book Series: 5, Madison, WI, pp 835–859
Zou XM, Ruan HH, Fu Y, Yang XD, Sha LQ (2005) Estimating soil labile organic carbon and potential turnover rates using a sequential fumigation–incubation procedure. Soil Biol Biochem 37:1923–1928
Acknowledgements
The authors would like to thank Yueqin Cheng (ISC-CAS), Angelika Kölbl, Greetje Pronk, and Olivia Kreyling (TU München) for laboratory work, Tabea Dietz and Sigrid Hiesch (CAU TU München) for determination of NH +4 and NO -3 concentration. This study was funded by the Deutsche Forschungsgemeinschaft (DFG) within the MAGIM project (Forschergruppe 536 MAGIM — Matter fluxes in grasslands of Inner Mongolia as influenced by stocking rate).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Wu, H., Wiesmeier, M., Yu, Q. et al. Labile organic C and N mineralization of soil aggregate size classes in semiarid grasslands as affected by grazing management. Biol Fertil Soils 48, 305–313 (2012). https://doi.org/10.1007/s00374-011-0627-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00374-011-0627-4