
Abstract
Decreased oxygen (O2) availability (hypoxia) is common in rock pools and challenges the aerobic metabolism of fishes living in these habitats. In this study, the critical O2 tension (Pcrit), a whole animal measure of the aerobic contribution to hypoxia tolerance, was compared between four New Zealand triplefin fishes including an intertidal specialist (Bellapiscis medius), an occasional intertidal inhabitant (Forsterygion lapillum) and two exclusively subtidal species (F. varium and F. malcolmi). The intertidal species had lower Pcrit values than the subtidal species indicating traits to meet resting O2 demands at lower O2 tensions. While resting O2 demand (standard metabolic rate; SMR) did not show a major difference between species, the intertidal species had higher maximal rates of O2 consumption (\( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \)) and higher aerobic metabolic scope (MS). The high O2 extractive capacity of the intertidal species was associated with increased blood O2 carrying capacity (i.e., higher Hb concentration), in addition to higher mass-specific gill surface area and thinner gill secondary lamellae that collectively conveyed a higher capacity for O2 flux across the gills. The specialist intertidal species B. medius also had higher glycogen stores in both white muscle and brain tissues, suggesting a greater potential to generate ATP anaerobically and survive in rock pools with O2 tensions less than Pcrit. Overall, this study shows that the superior Pcrit of intertidal triplefin species is not linked to a minimisation of SMR, but is instead associated with an increased O2 extractive capacity of the cardiorespiratory system (i.e., \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \), MS, Hb and gill O2 flux).
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
To carry out the necessary requirements of life, all aerobic organisms need to acquire enough O2 to drive ATP production via oxidative phosphorylation. Therefore, decreased O2 availability (environmental hypoxia), which is a common occurrence in a range of aquatic habitats, presents as a challenge for organisms such as fish (Diaz and Breitburg 2009; Farrell and Richards 2009). Intertidal fish can be exposed to hypoxic conditions when rock pool organisms consume O2 at a faster rate than it can be replaced by either algal photosynthesis or diffusion from air (Berschick et al. 1987; Bridges 1988; Richards 2011; Truchot and Duhamel-Jouve 1980). Thus, hypoxia is prevalent in rock pools at night when there is no photosynthetic activity to buffer O2 depletion due to respiration (Table 1; Fig. 1A, B). Well-mixed coastal subtidal habitats on the other hand have relatively stable O2 conditions (Fig. 1C) and, while hypoxia can develop in subtidal habitats (e.g., due to eutrophication and algal blooms or poor mixing), the frequent and large change in O2 availability typical of rock pools does not occur to the same extent. Notably, comparisons of closely related intertidal and subtidal species have proven to be a useful system in which to examine physiological mechanisms of hypoxia tolerance in fish (Brix et al. 1999; Hilton et al. 2010; Mandic et al. 2012; Mandic et al. 2009; Richards 2011; Speers-Roesch et al. 2013).

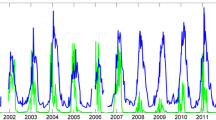
Changes in oxygen availability (% of air saturation) and temperature (°C) in two rock pools inhabited by intertidal triplefin fish (A, B) and at a shallow subtidal site (C). Values represent measurements logged every 5 min between the 5th and 6th of February 2019. LT low tide, HT high tide. Yellow and black shaded areas in panel C show daytime and nighttime hours, respectively. The two intertidal rock pools were located at Goat Island, Leigh, New Zealand (36°16′S, 174°47′E) and the subtidal site was located immediately adjacent to the rock pools at a depth of ~ 2 m (color figure online)
Environmental hypoxia represents a continuum of diminishing O2 availability under which it becomes increasingly challenging for organisms to take up adequate O2 to meet energetic demands via aerobic metabolism. If O2 availability is critically low, organisms cannot extract enough O2 to meet even basal energetic demands aerobically and O2 independent (anaerobic) ATP production is recruited to maintain energy balance (Richards 2011). O2 independent ATP production, however, is relatively inefficient, dependent on a finite pool of endogenous fermentable fuels (e.g., glycogen) and results in the accumulation of by-products (e.g., ADP, lactate and H+) which can be deleterious (Nilsson and Östlund-Nilsson 2008; Richards et al. 2007; Richards 2011; Speers-Roesch et al. 2013). Thus, a key strategy for hypoxia survival is to continue to meet O2 demand even at low environmental O2 tensions (PO2) (Mandic et al. 2009). In fish, the capacity to meet O2 demand under hypoxia can be measured as the critical PO2 (Pcrit), which is defined as the lowest O2 tension at which a resting O2 consumption rate (\( \dot{M}{\text{O}}_{2} \)) can be maintained. In practical terms, Pcrit is the PO2 at which \( \dot{M}{\text{O}}_{2} \) is forced lower than standard metabolic rate (SMR) (Claireaux and Chabot 2016; Rogers et al. 2016). Importantly, fish exposed to PO2 equivalent to 30% of their known Pcrit accumulate lactate, deplete tissue glycogen and ATP stores and eventually lose equilibrium (Speers-Roesch et al. 2013). In the sculpins, intertidal species with a lower Pcrit survive extreme hypoxia for longer (Mandic et al. 2012), showing that a low Pcrit makes an important contribution to overall hypoxia tolerance in at least some intertidal fishes. Theoretically, species with a lower Pcrit should be more hypoxia tolerant as a high extractive capacity for O2 allows energy demand to be met through the efficient aerobic pathway at lower PO2, thus avoiding or limiting the extent to which a time-limited anaerobic strategy must be relied upon for survival. It should be acknowledged, however, that some fishes can be very hypoxia tolerant and also have a relatively high Pcrit. This phenomenon is seen in the Amazonian oscar (Astronotus ocellatus) which can survive up to 6 h anoxia at 28 °C but has a Pcrit of 6.1 kPa (Scott et al. 2008). However, rather than relying on a high extractive capacity for O2, the Amazonian oscar is thought to achieve hypoxia tolerance through a combination of metabolic rate depression and a high tolerance of the end-products of anaerobic metabolism (Scott et al. 2008).
According to Mandic et al. (2009), intertidal sculpins have a lower Pcrit than their subtidal counterparts, indicating that the ability to meet resting O2 demand at low PO2 has been selected for fishes occupying intertidal habitats with variable O2 conditions. Mandic et al. (2009) also showed that sculpin species with a low Pcrit have red blood cells (RBC) with a higher affinity to bind O2 (low whole RBC P50), a larger mass-specific gill surface area and a lower routine \( \dot{M}{\text{O}}_{2} \). Thus, it appears that a low Pcrit requires a high extractive capacity for O2, which can then also be put to use to meet a low basal requirement for O2. Therefore, one prediction is that species with a lower Pcrit will have a cardiorespiratory system with a high capacity to take up O2 (i.e., a high maximum metabolic rate [\( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \)]) and also a low SMR (the best estimate of resting O2 demand in a recovered post-absorptive animal). This combination of respiratory characteristics would mean that species with a lower Pcrit will also have higher aerobic metabolic scope (MS), representing the difference between \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and SMR.MS reflects the capacity of an organism to perform O2 demanding activities (e.g., growth, activity, feeding, etc.) beyond maintenance requirements and a constraint upon MS has been proposed as a crucial determinant of performance in fish facing hypoxia (Chabot and Claireaux 2008; Claireaux and Chabot 2016; Claireaux and Lefrancois 2007). Intertidal rock pool fish with a high MS might therefore avert impaired aerobic performance during rock pool hypoxia (Fig. 1; Table 1), but MS has yet to be compared among intertidal and subtidal fishes.
New Zealand triplefins (family: Tripterygiidae) are a highly specious and closely related group of marine blennoid fishes (Hickey and Clements 2005) which show habitat partitioning by depth and exposure across the nearshore coastal environment (Wellenreuther et al. 2007). Comparisons among New Zealand triplefin fishes already show that intertidal species have a lower Pcrit than subtidal species (Hilton et al. 2008, 2010; Hilton 2010). However, it is possible that the protocols and respirometry methods used in this previous work produced elevated estimates of Pcrit. For example, fish within these studies were allowed to recover for only 2.5 h within respirometers prior to hypoxia exposure and Pcrit measurement. The problems associated with the use of a short recovery time when estimating Pcrit are twofold. First, a reliable estimate of resting \( \dot{M}{\text{O}}_{2} \) (i.e., SMR) may be precluded and, if this is the case, Pcrit has to be identified as the breakpoint in \( \dot{M}{\text{O}}_{2} \) during hypoxia exposure using segmented regression. Segmented regression is not recommended for Pcrit determination because, if for any reason the \( \dot{M}{\text{O}}_{2} \) of fish is elevated at the time of hypoxia exposure (e.g., due to incomplete recovery from handling stress or spontaneous activity), this methodology will overestimate Pcrit (Claireaux and Chabot 2016). This problem is exacerbated when making comparisons of hypoxia tolerance among multiple species that may recover from handling stress at different rates, and/or be more or less active within respirometers. Furthermore, previous estimates of Pcrit in triplefins (i.e., Hilton et al. 2008; Hilton 2010; Hilton et al. 2010) were made using the closed-system respirometry, which in some cases has been demonstrated to confound \( \dot{M}{\text{O}}_{2} \) measurements due to the accumulation of waste products (e.g., CO2) within respirometers (Rogers et al. 2016; Snyder et al. 2016). Therefore, in the present investigation, we determined it necessary to re-examine the Pcrit of triplefin species, but this time using intermittent stop-flow respirometry (Steffensen 1989) and more established methods for Pcrit measurement (see Claireaux and Chabot 2016; Snyder et al. 2016).
In this study, the capacity to meet resting O2 demands under hypoxia (i.e., Pcrit) was compared among four New Zealand triplefin fish species (Bellapiscis medius, Forsterygion lapillum, F. varium and F. malcolmi). These four species were included as they occupy different habitat depths (intertidal to subtidal; Table 2) and represented a broad range of Pcrit for this group (i.e., low, medium and high Pcrit) in a previous investigation (Hilton 2010). B. medius is an intertidal specialist which occupies rock pools located on the shoreline, on average 1.39 m (range 0.31–3.39 m) above chart datum (ACD) (Hilton et al. 2008). F. lapillum is an occasional occupant of intertidal rock pools, but is most abundant in relatively shallow (mean depth of occurrence 3.5 m; Table 2) subtidal habitats. In the intertidal zone, F. lapillum is less abundant than B. medius, however, their distributions, at least in terms of shoreline elevation, do overlap as F. lapillum is found in rock pools on average 1.21 m (range 0.19–3.09 m) ACD (Wellenreuther 2007). Forsterygion varium and F. malcolmi are both exclusively subtidal species with mean depth of occurrences of 8 and 11 m, respectively (Table 2). The inclusion of these four species allowed the physiological characteristics of a specialist intertidal species (B. medius) to be compared to three other relatively closely related triplefin species which are either occasionally intertidal (F. lapillum) or exclusively subtidal (F. varium and F. malcolmi). Additionally, we could compare the physiological characteristics of the occasional intertidal rock pool inhabitant F. lapillum to that of two exclusively subtidal species from the same genus (F. varium and F. malcolmi). This comparison was advantageous because if the same physiological characteristics distinguishing B. medius from the exclusively subtidal species were also seen in F. lapillum, this would provide evidence that the observed characteristics of intertidal species result from selection for intertidal conditions rather than phylogeny.
Specifically, the aims of this study were: (1) to re-examine if there are differences in Pcrit between intertidal and subtidal triplefin fishes, and (2) to identify physiological characteristics associated with Pcrit among intertidal and subtidal triplefin fishes. To address these aims, the Pcrit for SMR (Claireaux and Chabot 2016) was measured to establish differences in the ability to meet resting O2 demands under hypoxia among species. Whole animal metabolic rate (SMR, \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and MS) was assessed to determine if a low Pcrit is associated with a low resting O2 demand and/or a high extractive capacity for O2 as shown in other fishes (Richards 2011). Blood Hb concentration and gill morphometry were also measured as these may be associated with extractive capacity for O2 and Pcrit (Mandic et al. 2009). Finally, tissue glycogen was measured in brain, liver and white muscle, as it has been hypothesised that hypoxia-tolerant fishes have higher endogenous stores of fermentable fuels (Mandic et al. 2012; Richards 2011).
Methods
Rock pool O2 measurements
The O2 content of rock pools occupied by intertidal triplefin fishes was assessed to gain an understanding of the severity of hypoxia that occurs under natural conditions. Two sets of measurements were made: (1) O2 and temperature were logged at 5 min intervals in two rock pools over a ~ 24 h period (Fig. 1A, B) and spot measurements of O2 and temperature were taken in ten rock pools at night time low tide and then again in the same rock pools the following day at low tide (Table 2). To make the continuous measurements, O2 loggers (D-Opto O2 logger, Zebra-Tech Ltd, Nelson, New Zealand) were placed in rock pools for a period of ~ 24 h between the 5th and 6th of February 2019. The night and daytime spot measurements were taken on the 26th of February 2016 approximately an hour after low tide and were made at mid-depth level in the rock pools. These measurements were made with a FireSting O2 metre and shielded temperature probe (PyroScience, Aachen, Germany). The species of fish observed in each of the ten rock pools at the time spot measurements were made was also recorded. The continuous measurements were made in rock pools located at Goat Island, Leigh, New Zealand (36°16′S, 174°47′E) and the spot measurements were made in rock pools located at Hatfields Beach, Auckland, New Zealand (36°34′S, 174°41′E). Changes in O2 and temperature over a ~ 24 h period were also assessed at a shallow subtidal site (~ 2 m deep) at Goat Island, Leigh, New Zealand (36°18′S 174°47′E). These measurements were taken between the 5th and 6th of February 2019 using the O2 logger described above.
Experimental animals and laboratory acclimation
Four triplefin species (B. medius, F. lapillum, F. varium and F. malcolmi) were collected from the wild for this study. The intertidal triplefin B. medius was netted from rock pools and F. lapillum, F. varium and F. malcolmi were hand netted from subtidal habitats by divers. The animals were housed in 30 L flow-through seawater tanks (air saturated, 200 μm filtered, 35 ppt salinity) at the Leigh Marine Laboratory. Fish were acclimated to normoxia and a temperature of 18 °C (± 0.5 °C) for a period of at least 4 weeks prior to experiments and were fed daily on a mixture of crushed aquaculture feed (Skretting, Australia) and pilchard. Food was withheld for a period of 48 h prior to experiments, which were performed under approval of the University of Auckland Animal Ethics Committee (AEC approval number 001441).
General respirometry methods
The mass-specific O2 consumption rate (\( \dot{M}{\text{O}}_{2} \)), reported as microgram of O2 consumed per gram of body weight per hour (mg O2 g−1 h−1), was determined using automated intermittent flow respirometry (Steffensen 1989). Custom-built respirometry chambers (42–210 mL) were held within a 60 L reservoir filled with filtered (1 µm) UV-sterilised seawater, which was heated or chilled to the experimental temperature (~ 18 °C) by continually pumping the seawater through a 40 L heat exchange tower containing an aluminium coil heat exchanger. The inlet of each chamber was connected to an automated Eheim compact 3000 submersible flush pump (EHEIM GmbH & Co. KG, Germany) which was switched on and off by a relay control unit (USB Power 8800 Pro, Aviosys International Inc, Taiwan) controlled by custom-coded software (Leigh Marine Laboratory). A magnetic stir bar was housed in a recess in the bottom of the respirometry chamber to ensure adequate water mixing and the O2 concentration of water within the chamber was continuously measured using contactless sensor spots followed by FireSting O2 metre (PyroScience, Aachen, Germany). The decline in O2 concentration within a respirometry chamber was used to calculate \( \dot{M}{\text{O}}_{2} \) in repeated measurement cycles according to the equation:
where V is the respirometry chamber minus fish volume, \( \Delta \% {\text{sat}}/t \) is the change in O2 saturation per unit time, α is the solubility coefficient of O2 (mg O2 %Sat−1 L−1) in sea water (35 ppt), and MB is the body mass of the fish in grams (Schurmann and Steffensen 1997). In all instances of \( \dot{M}{\text{O}}_{2} \) assessment, the repeated measurement cycles were interspersed with 1 min periods of flushing to fully replace the water volume of the chamber. Background respiration was assessed after the fish was removed from the respirometer, but remained negligible throughout the trials. In all estimates of metabolic rate, only \( \dot{M}{\text{O}}_{2} \) values with R2 of > 0.90 for the decline in O2 per unit of time were used.
Using the procedures above, \( \dot{M}{\text{O}}_{2} \) was measured to establish the standard metabolic rate (SMR), maximum metabolic rate (\( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \)), aerobic metabolic scope (MS) and Pcrit of fish (NB. SMR was measured and reported twice as part of two separate procedures. See below).
Measurement of maximum metabolic rate and aerobic metabolic scope
\( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and MS were determined in all species at 18 °C (N = 10–11) (B. medius: body mass 2.32 g ± 0.37, F. lapillum: body mass 1.98 g ± 0.15, F. varium: body mass 6.1 g ± 0.85, F. malcolmi: body mass 6.97 g ± 0.88). \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) was determined following exhaustive exercise, where fish were continually chased (5 min) in a 30 L tank. An exhaustive exercise protocol has previously been used to elicit \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) values in triplefins (Khan et al. 2014a; McArley et al. 2017) and is best suited for obtaining \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) in benthic species which will not continuously swim in a flume (Clark et al. 2013; Norin and Clark 2016). Following exhaustive exercise, the fish was transferred to a respirometry chamber within 30 s of the conclusion of chasing and repeating 4 min \( \dot{M}{\text{O}}_{2} \) measurement cycles were initiated. \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) was taken as the highest \( \dot{M}{\text{O}}_{2} \) value recorded in any measurement cycle, which in almost all cases was obtained from the first measurement cycle following exhaustive exercise. When the metabolic rate of fish was clearly declining, the measurement period was extended to 8 min and \( \dot{M}{\text{O}}_{2} \) was measured repeatedly for > 16 h. During the overnight measurement period, fish were left undisturbed in the respirometers and \( \dot{M}{\text{O}}_{2} \) typically recovered to levels around SMR within ~ 6–8 h. SMR was estimated from the mean of the lowest 10% of \( \dot{M}{\text{O}}_{2} \) over this time (Khan et al. 2014b; McArley et al. 2017; Norin et al. 2014). Due to the benthic habit of these triplefin species, they remain relatively inactive and perch in a stationary position on the bottom of the respirometers. This behavioural characteristic means that the lowest \( \dot{M}{\text{O}}_{2} \) values recorded were probably related to periods when the fish was in an inactive state and are therefore likely to be a fair representation of SMR. MS was defined as the difference between the mass corrected \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and SMR (see below) for each individual.
Measurement of critical oxygen tension (P crit)
Pcrit was measured in B. medius (body mass 3.22 g ± 0.41), F. lapillum (body mass 1.72 g ± 0.12), F. varium (body mass 6.6 g ± 0.98) and F. malcolmi (body mass 4.95 g ± 1.16) at 18 °C following a similar protocol to Cumming and Herbert (2016) (N = 9–10). In this protocol, SMR was once again measured under normoxic conditions, then a progressive hypoxic exposure was used to identify the water O2 tension at which \( \dot{M}{\text{O}}_{2} \) was no longer maintained above SMR. SMR was determined using automated intermittent flow respirometry (see respirometry methods above), where fish were left undisturbed in respirometers overnight for ~ 16 h (~ 1700–0900 hours). During overnight respirometry, \( \dot{M}{\text{O}}_{2} \) was repeatedly measured over 7–9 min cycles and SMR was taken as the mean of the lowest 10% of \( \dot{M}{\text{O}}_{2} \) values. \( \dot{M}{\text{O}}_{2} \) measurements were then made at decreasing levels of water O2 content (75%, 55%, 40%, 30%, 25%, 20%, 15%, 10%, and 6% of air saturation) and the required water O2 levels were achieved by bubbling nitrogen into the seawater reservoir supplying respirometers. Three 7–9 min \( \dot{M}{\text{O}}_{2} \) measurements were made at 75%, 55%, 40%, 30%, 25% and 20% of air saturation, and one 7–9 min measurement at 15%, 10% and 6% of air saturation. The progressive decline in O2 tension to determine Pcrit was completed in ~ 3.5 h and the time length of exposure to each O2 level was the same for each species. To determine Pcrit, SMR and \( \dot{M}{\text{O}}_{2} \) during progressive hypoxic exposure were first mass corrected (see below), then plotted against water O2 tension. A linear regression (forced through zero) was then established on \( \dot{M}{\text{O}}_{2} \) values that fell below SMR and Pcrit was calculated by dividing SMR by the slope of this regression line (i.e., the point where \( \dot{M}{\text{O}}_{2} \) under progressive hypoxia could no longer be maintained above SMR; see Fig. 2) (as per the method of Behrens and Steffensen 2007; Cook et al. 2013; Cumming and Herbert 2016; Schurmann and Steffensen 1997).

Critical oxygen tension (Pcrit) and mass-specific oxygen consumption (\( \dot{M}{\text{O}}_{2} \)) under progressive hypoxia exposure in B. medius (A), F. lapillum (B), F. varium (C) and F. malcolmi (D) at 18 °C. All values are mean ± SEM. and \( \dot{M}{\text{O}}_{2} \) values are corrected to a body mass of 4 g (N = 9–10). Triangles show standard metabolic rate (SMR) under normoxia, circles show \( \dot{M}{\text{O}}_{2} \) under progressive hypoxia exposure, vertical line shows Pcrit and vertical dotted line shows Pcrit SEM. The dashed horizontal line shows the break point where \( \dot{M}{\text{O}}_{2} \) falls below SMR (i.e., Pcrit) and is for illustrative purposes only. Different lower case letters adjacent to the vertical line representing Pcrit show significant differences in Pcrit between species (P < 0.05). There were no significant differences in SMR among species (see “Results”)
Scaling of metabolic measurements
To account for body mass differences between species, the \( \dot{M}{\text{O}}_{2} \) values were standardized to the mean body mass of all fish (4 g) using the formula:
where \( \dot{M}{\text{O}}_{{2(4{\text{g}})}} \) is the \( \dot{M}{\text{O}}_{2} \) for a fish with the standardized (corrected) new weight of 4 g, \( \dot{M}{\text{O}}_{{2({\text{meas}})}} \) is the measured \( \dot{M}{\text{O}}_{2} \), w is the weight of the fish, w(4g) is the standardized body mass of fish set to 4 g and A is the weight exponent describing the relationship between metabolic rate and body weight (Schurmann and Steffensen 1997). The mass exponent (A) used to correct \( \dot{M}{\text{O}}_{2} \) to body mass was 0.8 (Clarke and Johnston 1999).
Blood haematology, tissue glycogen and gill morphometry
Haematology, tissue glycogen, and gill morphometry were assessed in B. medius (body mass 4.25 g ± 0.24), F. lapillum (body mass 2.16 g ± 0.09), F. varium (body mass 10.55 g ± 0.88) and F. malcolmi (body mass 8.74 g ± 0.81) acclimated to 18 °C for a period of 4 weeks (N = 8–10). 24 h prior to sampling, fish were transferred to individual 14 L flow-through seawater tanks so that they were not disturbed by the repeated removal of fish from the same common holding tanks. First, the fish was netted from its individual holding tank and immediately euthanized by pithing of the brain. The caudal peduncle was then severed and blood was collected by holding the tail of the fish into the base of a heparinised 0.5 mL Eppendorf tube. The brain, liver and a sample of white skeletal muscle were then immediately frozen under liquid N2 and stored at − 80 °C. The right gill basket was removed and fixed in Bouin’s solution for 48 h and then stored in 70% ethanol. Haemoglobin (Hb) concentration of fresh blood was quantified spectrophotometrically at 540 nm using modified Drabkin’s reagent (Wells et al. 2007). Haematocrit (Hct) was determined in 75 mm capillary tubes spun for a period of 10 min in a haemofuge (Haemocentaur, MSE, London, UK). Tissue glycogen content was determined in thawed brain, liver and white muscle which was homogenised in ice cold 0.6 M perchloric acid. Glycogen (in glycosol units) was determined by measuring glucose content of tissue extract with and without prior incubation with amyloglucosidase as per the method of Keppler and Decker (1974). Glucose content of the tissue extract was determined using a commercially available assay kit (d-Glucose HK Assay Kit, Megazyme, Ireland).
Gill morphometry was assessed by light microscopy on intact whole gill arches (WGA) and longitudinally sectioned gill arches (SGA) from the right gill basket. All analyses of gill images were performed using Image J software (U. S. National Institutes of Health, Bethesda, MD, USA). Several gill morphometric parameters were measured including: (1) filament number (WGA), (2) filament length (WGA), (3) secondary lamellae (SL) density (WGA), (4) SL basal length (WGA), (5) SL protruding height and thickness (SGA), (6) SL bilateral surface area, and (7) mass-specific gill surface area. First, the four gill arches of the right gill basket were separated and photographed under a stereomicroscope. The number of filaments on each gill arch was counted and the length of all filaments in a single row on each arch was measured. The filament length in a single row was then multiplied by two to estimate the total length of filaments on each gill arch. To estimate the total length of gill filaments for the entire gill system (i.e., the right and left gill basket), the total length of filaments measured on each gill arch was summed and then multiplied by two. SL density was estimated by measuring the distance occupied by at least ten SL on four filaments on each gill arch. The bilateral surface area of SL (mm2) was calculated as the area of a half ellipse based on measurements of the basal length, protruding height and thickness of SL according to the formula outlined in Matey et al. (2008). SL basal length was measured on different filaments than SL height and SL thickness due to the requirement for sectioning. As such, the mean value of each of these parameters for individual fish was used to calculate a global average of SL bilateral SA for each fish. To estimate the basal length of SL, individual filaments were removed from the third gill arch and photographed under a stereomicroscope. The width of filaments, where the base of the SL attaches, was then measured at ten evenly spaced distances along their length to approximate the basal length of the SL. To measure the thickness and protruding height of SL, the whole second gill arch was embedded in paraffin, sectioned longitudinally (5 µm) using a microtome, mounted on slides and stained with haematoxylin and eosin. The height and width of 30 SL was then measured from sections where the central venous sinuous of the filament was visible. To estimate the total surface area of the gills, the total length of gill filaments was multiplied by SL density to provide an estimate of the total number of SL in the entire gill system. The total number of SL was then multiplied by the SL bilateral surface area to produce an estimate of total gill surface area in mm2. The total gill surface area was then standardized to body mass and is presented as mm2 per g of body mass. To take into account the differences in the thickness of the SL between species, the maximum O2 uptake capacity of the gills was determined according to Fick’s law:
where VO2 is the maximum O2 uptake rate for the gills obtained by morphometric measurements, K is a diffusion constant, A is the gill area, dPO2 is the difference between the partial pressure of O2 on either side of the membrane and t is the thickness of the water–blood barrier (Kunzmann 1990). The thickness of the water–blood barrier (t) was taken as half the thickness of the SL and the difference in partial pressure of O2 between seawater and blood was assumed to be 110 mmHg. K was taken as the diffusion coefficient for O2 in water at 18 °C.
Statistics
In all statistical tests, significance was accepted at P < 0.05. Pcrit, SMR, \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \), MS, gill morphometric parameters, haematological parameters and tissue glycogen were each compared between species using one-way analysis of variance (ANOVA) with Holm–Sidak post hoc comparisons. Analysis of covariance (ANCOVA) was used to determine whether there was an influence of body mass on Pcrit and mass-specific gill surface area. Since the covariate was not found to be significant in both instances (P > 0.05), a standard one-way ANOVA was used to compare Pcrit and mass-specific gill surface area among the four species. Where assumptions of normality and equality of variances were violated, data were either log transformed for the analysis or a non-parametric Kruskal–Wallis ANOVA on rank test was used. All statistical analysis was carried out using Sigma Plot 13.0 software package.
Results
P crit and standard metabolic rate in hypoxia tolerance assessed fish
The four species examined showed a similar aerobic response to progressive hypoxia exposure with \( \dot{M}{\text{O}}_{2} \) remaining constant and slightly elevated relative to SMR at higher PO2, before abruptly declining and falling below SMR at lower PO2 (Fig. 2). A difference among species in the ability to maintain \( \dot{M}{\text{O}}_{2} \) above SMR under hypoxia was confirmed by significant differences in Pcrit (ANOVA, df = 3, F = 12.83, P < 0.001). Post hoc comparisons showed that B. medius had a lower Pcrit than all other species and F. lapillum and F. varium had a lower Pcrit than F. malcolmi (Fig. 2). In the fish assessed for hypoxia tolerance (Pcrit), the SMR of B. medius (0.079 mg O2 g−1 h−1 ± 0.003) was slightly lower than the SMR of F. lapillum (0.10 mg O2 g−1 h−1 ± 0.008), F. varium (0.096 mg O2 g−1 h−1 ± 0.008) and F. malcolmi (0.11 mg O2 g−1 h−1 ± 0.005) (Fig. 2). However, although there appeared to be a difference in SMR among species (ANOVA, df = 3, F = 2.95, P = 0.046), none of the pairwise post hoc comparisons were significant (P > 0.05).
Maximum metabolic rate and aerobic metabolic scope
In the fish assessed for \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and MS, there were significant differences in SMR among species (Kruskall–Wallis ANOVA, df = 3, H = 11.59, P = 0.009). While B. medius appeared to have the lowest SMR of the four species, post hoc comparisons showed the only difference in SMR was between B. medius and F. varium (Fig. 3A). There were also significant differences in both \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) (ANOVA, df = 3, F = 20.69, P < 0.001) and MS (ANOVA, df = 3, F = 27.39, P < 0.001) between species. B. medius had significantly higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and MS than all other species and F. lapillum had significantly higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and MS than F. varium and F. malcolmi (Fig. 3B, C).

A Standard metabolic rate (SMR), B maximum metabolic rate (\( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \)), and C aerobic metabolic scope (MS) in B. medius, F. lapillum, F. varium and F. malcolmi at 18 °C. All values are mean ± SEM and corrected to a body mass of 4 g (N = 10–11). Lower case letters not shared between bars show significant differences between species (P < 0.05)
Gill morphometric parameters
There were significant differences in the number of filaments per gill arch among species (ANOVA, df = 3, F = 15.31, P < 0.001) with F. lapillum having fewer filaments than B. medius, F. varium and F. malcolmi (Table 2). The length of gill filament per unit of body mass was greater in B. medius and F. lapillum than in F. varium and F. malcolmi (ANOVA, df = 3, F = 70.38, P < 0.001, Table 2) and secondary lamellae density was also greater in B. medius and F. lapillum than in F. varium and F. malcomi (ANOVA, df = 3, F = 123.84, P < 0.001, Table 1). The basal length (Kruskall–Wallis ANOVA, df = 3, H = 24.01, P < 0.001), height (ANOVA, df = 3, F = 32.98, P < 0.001) and thickness (ANOVA, df = 3, F = 46.81, P < 0.001) of secondary lamellae were smaller in B. medius and F. lapillum than in F. varium and F. malcolmi (Table 2) and, as a result, the estimated bilateral surface area of the secondary lamellae was also smaller in B. medius and F. lapillum than in F. varium and F. malcolmi (Table 2). There was a significant difference in mass-specific gill surface area between species (ANOVA, df = 3, F = 10.84, P < 0.001) with F. lapillum having greater mass-specific gill surface than B. medius, F. varium and F. malcolmi (Table 2). There was some evidence that mass-specific gill surface area decreased with increasing body mass (ANCOVA: covariate, df = 1, F = 3.99, P = 0.057). Therefore, the high mass-specific gill surface area of F. lapillum could be due to the comparatively smaller body mass of this species. The maximum rate of O2 uptake across the gills (gill VO2) calculated according to Fick’s law was significantly higher in both B. medius and F. lapillum than in F. varium and F. malcolmi (ANOVA, df = 3, F = 29.19, P < 0.001, Table 2).
Tissue glycogen stores and haematological parameters
There were differences in brain (ANOVA, df = 3, F = 7.38, P < 0.001) and white muscle (ANOVA, df = 3, F = 17.04, P < 0.001) glycogen stores among species. B. medius had a greater amount of glycogen in brain and white muscle than F. lapillum, F. varium and F. malcolmi and F. lapillum had greater white muscle glycogen than F. malcolmi (Table 2). There were no differences in the amount of glycogen per gram of liver tissue among species (ANOVA, df = 3, F = 0.35, P = 0.79), but liver size (hepatosomatic index, HSI) was also different among species (ANOVA, df = 3, F = 6.85, P < 0.001, Table 2). Thus, when liver glycogen was expressed per gram of body mass, there were differences among species (ANOVA, df = 3, F = 3.62, P = 0.022). While B. medius had higher liver glycogen per gram of body mass than all other species, only the post hoc comparison with F. lapillum was significant (Table 2). Haemoglobin concentration was different among species (ANOVA, df = 3, F = 6.34, P = 0.002) with B. medius and F. lapillum having a higher haemoglobin concentration than F. malcolmi (Table 2). There was no difference in haematocrit among species (ANOVA, df = 3, F = 0.55, P = 0.65).
Discussion
The pattern of P crit between intertidal and subtidal triplefin species
Bellapiscis medius, which appears to be exclusively intertidal and occupies rock pools high on the shoreline (Hilton et al. 2008), has a Pcrit which is 53%, 77% and 128% lower than F. lapillum, F. varium, and F. malcolmi, respectively. Lower Pcrit in intertidal fish species has also been demonstrated among sculpins (Mandic et al. 2009) and the observed pattern mirrors the abrupt change in the prevalence of environmental hypoxia that exists between intertidal and subtidal habitats. F. lapillum was also observed in moderately hypoxic rock pools (Table 1), so the intermediate Pcrit of this species is consistent with the balance of residing in both intertidal and relatively shallow subtidal habitats. Comparing the two exclusively subtidal species, F. varium had a lower Pcrit than F. malcolmi, but this is unlikely to reflect a difference in exposure to hypoxia as these species overlap across depth (Table 2) and utilise almost identical habitats (Wellenreuther et al. 2007). An organism with a low Pcrit can meet O2 demand under more severe levels of hypoxia than can an organism with a high Pcrit and the advantages are twofold. First, it is possible a low Pcrit delays the initiation of anaerobic ATP production to maintain energy balance during a hypoxic event (Mandic et al. 2009; Richards 2011). Second, among intertidal and subtidal sculpins, a correlation has been demonstrated between Pcrit and the ability to maintain an upright position (i.e., equilibrium, a proxy for survival) under severe hypoxia (Mandic et al. 2012). This suggests a lower Pcrit plays a role in allowing intertidal fishes to survive severe hypoxia for longer periods of time than their subtidal counterparts. Although loss of equilibrium was not directly assessed in the current study, it was frequently observed in F. varium and F. malcolmi at the lowest O2 levels during Pcrit determination. Furthermore, in a parallel investigation, we have observed a strong correlation between Pcrit and time to loss of equilibrium under severe hypoxia among the four species included in the current study (Devaux et al. unpublished data). Thus, for intertidal fishes that reside in hypoxia-prone rock pools, a lower Pcrit appears to make an important contribution to an enhanced ability to survive periods of severe hypoxia exposure.
Previous investigations by Hilton (2010) and Hilton et al. (2010) also found B. medius has a lower Pcrit than subtidal triplefin species. However, Pcrit in the current study at 18 °C was 58–157% lower for all species compared to the data of Hilton et al. at 15 °C. This likely results from the longer recovery time used in our study (16 h versus 2.5 h) and the use of intermittent flow respirometry for the determination of Pcrit (Snyder et al. 2016). Estimates of resting \( \dot{M}{\text{O}}_{2} \) in Hilton (2010) and Hilton et al. (2010) were also at least 50% higher compared to SMR in the current study and in the case of F. varium were 232% higher. Therefore, when hypoxia exposure was initiated only 2.5 h following entry to respirometers and closed respirometry was used, it is likely the fish under observation by Hilton et al. were still recovering from initial handling stress and that this resulted in relatively high estimates of Pcrit because the fish were not at SMR. However, despite these discrepancies and differences in technique, all studies on triplefins to date show that intertidal species have a lower Pcrit than their related subtidal counterparts.
Physiological characteristics associated with P crit in triplefin fishes
Organisms with a high O2 extractive capacity will be able to meet the O2 demand of a given aerobic metabolic rate at lower O2 tensions (Richards 2011). Therefore, among organisms which have a similar SMR, those with a high extractive capacity for O2 should be able meet the O2 demands of resting metabolism at lower O2 tensions, and hence have lower Pcrit (Cook et al. 2011). In the current study, \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) following exhaustive exercise was assessed to provide an index of the maximum capacity of the cardiorespiratory system of each species to remove O2 from the environment (i.e., the maximal extractive capacity for O2). The two intertidal species B. medius and F. lapillum had higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) than the exclusively subtidal species demonstrating that the cardiorespiratory system of the intertidal species has a higher capacity to take up and deliver O2 to tissues. As there were only small differences in SMR among species, the high extractive capacity for O2 of the intertidal species allows resting O2 demand to be met at a lower O2 tension and, therefore, their Pcrit is also lower (e.g., see Cook et al. 2011). A high extractive capacity for O2 would also lower the limiting PO2 for other O2 demanding activities over and above SMR (e.g., digestion, feeding, activity, etc.). This could benefit intertidal fishes as it may allow them to avoid limitation of aerobic performance for longer periods of time during hypoxia exposure (i.e., to lower O2 tensions). While the Pcrit of an animal is likely a function of maximal O2 extraction capacity, it could also depend on resting O2 demand (assessed as SMR in the current study). The intertidal specialist B. medius had a lower Pcrit and higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) than the other species, but there was little evidence of it also having a lower SMR (Figs. 2, 3). It was also clear that F. lapillum, F. varium and F. malcolmi had an almost identical SMR, but different Pcrit. As a result, there was no trend between Pcrit and SMR in the four species examined. Among sculpins, variation in routine O2 demand (an estimate of resting metabolic rate) also explained a comparatively smaller part of the variation in Pcrit among species than did variation in characteristics associated with extractive capacity for O2 (HbO2-binding affinity and mass-specific gill surface area) (Mandic et al. 2009; Richards 2011). Thus, the findings of the current study suggest that, like sculpins, the extractive capacity for O2 plays a more important role than resting demand for O2 in setting Pcrit among triplefin fishes.
The observed trend of higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and lower Pcrit in the intertidal species may be associated with a range of factors influencing the cardiorespiratory cascade (e.g., blood O2 carrying capacity, HbO2-binding affinity, gill morphometric parameters, and cardiac output). In the current study, the intertidal species had a higher Hb concentration than subtidal species; this could be involved as a driver of increased \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and lower Pcrit because it would raise blood O2 carrying capacity. Indeed, Cook et al. (2011) used phenylhydrazine to pharmacologically induce anaemia (i.e., lower blood O2 carrying capacity) of a sea bream, which ultimately reduced \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and increased Pcrit compared to sham controls. Secondarily, the O2 extractive capacity of the gills could also feasibly link high \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and low Pcrit together. For example, in a previous study by Mandic et al. (2009), a high mass-specific gill surface area was associated with a lower Pcrit among 12 species of sculpins. The results of the current study provide some evidence that mass-specific gill surface area also plays a role in setting Pcrit among triplefin fish. The mass-specific gill surface area was significantly higher in the occasionally intertidal species F. lapillum than in the two deeper dwelling exclusively subtidal species F. varium and F. malcolmi. This trend, however, was not observed in the intertidal specialist B. medius as, despite a low Pcrit, its mass-specific gill surface area was not greater than the subtidal Forsterygion species. The secondary lamellae of both B. medius and F. lapillum, however, were thinner than those of the two exclusively subtidal species, suggesting a shorter diffusion distance for O2 to reach the red blood cells during gill ventilation. Assuming that half the thickness of the secondary lamellae is representative of the blood O2 diffusion distance across the secondary lamellae, we estimate that gill VO2 (Table 2) was approximately double in the intertidal specialist B. medius and occasional intertidal occupant F. lapillum in comparison to the two exclusively subtidal Forsterygion species. Thus, the larger gill surface area and thinner secondary lamellae of the intertidal specialist B. medius and occasional intertidal F. lapillum provides a high capacity for gill O2 flux, which likely contributes to both increased \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and lower Pcrit in these species. The relationship between gill VO2 and Pcrit among species, however, like that for gill surface area and Pcrit, was also not straightforward. There was no difference in gill VO2 between B. medius and F. lapillum despite these species having different Pcrit and there was also no difference in gill VO2 between F. varium and F. malcolmi despite differences in Pcrit. These discrepancies in the relationship between gill morphometric parameters and Pcrit are likely due to the fact that Pcrit is a complex whole animal trait influenced by a number of other physiological parameters (e.g., ventilatory capacity, cardiac output, blood oxygen binding affinity) not investigated in the current study.
Predominately, as a result of higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) the intertidal specialist B. medius and the occasionally intertidal and shallow subtidal dwelling F. lapillum had higher MS (the difference between \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and SMR) than the deeper dwelling exclusively subtidal species F. varium and F. malcolmi. The first limitation imposed by hypoxia on aerobic capacity, which occurs at PO2 well above Pcrit, is a constraint on \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) and a reduction in MS (Chabot and Claireaux 2008; Claireaux and Lagardère 1999; Claireaux and Chabot 2016; Claireaux et al. 2000; Cook et al. 2011; Farrell and Richards 2009; Lefrançois and Claireaux 2003). Exposure to increased temperature, as occurs for intertidal fish during the day in rock pools (e.g., Fig. 1), also limits MS because increased resting O2 demand reduces the difference between SMR and \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) (Pörtner and Knust 2007; Schulte 2015). As aerobic activities (e.g., growth, feeding, and activity) must occur within the bounds of MS, it is thought that MS-limiting environmental conditions can cause energy budgeting conflicts which lead to a loss of whole organism performance (e.g., reduced growth in Atlantic cod under hypoxia) (Chabot and Claireaux 2008). Potentially, a high MS provides a reserve of aerobic capacity which intertidal fishes can utilise to avoid severe constraints on performance under exposure to hypoxia or high temperature in rock pools. Further studies examining the effects of hypoxia on other aerobic activities (e.g., specific dynamic action, which is probably forced to operate within the bounds of MS) would therefore provide useful insight into whether rock pool fishes utilise high MS to their advantage.
The pattern of MS observed among species in the current study could also result from different regimes of wave exposure between intertidal, shallow subtidal and deeper subtidal habitats. Hickey and Clements (2003) determined that intertidal triplefins are subjected to substantially higher water velocities than those species occupying subtidal habitats, and that water velocities resulting from wave action decline progressively with increasing habitat depth. Thus, the higher MS of the specialist intertidal species B. medius and the occasionally intertidal and shallow subtidal dwelling F. lapillum could reflect an increased requirement for activity (e.g., station holding) in the high water velocities of intertidal and shallow habitats. The comparatively lower MS of F. varium and F. malcolmi may reflect a lower demand for activity as the water velocities resulting from wave action will not be as high in the deeper subtidal habitats occupied by these species.
Although a low Pcrit is beneficial for fish exposed to hypoxia, it does not preclude exposure to O2 tensions capable of threatening survival. Indeed, intertidal fish are more at risk of exposure to O2 tensions less than Pcrit than their subtidal counterparts, because environmental hypoxia does not regularly occur in the habitat of the latter. At O2 tensions below Pcrit, organisms rely at least partly on O2 independent ATP production to maintain energy balance. This is evidenced by the accumulation of lactate in the tissues and plasma of fish exposed to O2 tensions below Pcrit (Herbert and Steffensen 2005; Scott et al. 2008; Speers-Roesch et al. 2013). Thus, fish species that reside in hypoxia-prone habitats should have large stores of endogenous fermentable fuels such as glycogen (Richards 2011). The intertidal specialist B. medius maintained a higher resting concentration of glycogen in white skeletal muscle and brain tissue than the occasional intertidal occupant F. lapillum, and the two exclusively subtidal Forstyregion species. Although there was no difference in liver glycogen concentration between species, B. medius did have a larger liver relative to body mass. Thus, there was some evidence of a trend for increased liver glycogen stores relative to body mass in B. medius, which fits with the theory outlined above that intertidal fishes may store higher levels of endogenous fermentable fuels to power anaerobic metabolism under hypoxia exposure. Higher tissue glycogen stores in intertidal fishes could also be a response to a more variable food supply, as a limitation in food availability may be a common occurrence in rock pool habitats (Silberschneider and Booth 2001).
A limitation of the current study is that the intertidal specialist B. medius is placed in a different genus than the subtidal species examined. Thus, we cannot rule out the possibility that the observed differences in physiological traits between B. medius and the subtidal species are a result of phylogeny rather than adaptation to rock pool conditions. It is noteworthy, however, that F. lapillum, a species which also occurs in the intertidal had a lower Pcrit, and higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \)¸ MS, blood Hb concentration, and gill VO2 than exclusively subtidal species F. varium and F. malcolmi. Thus, the physiological characteristics of B. medius that appear suited to a rock pool existence also differentiated F. lapillum from two closely related subtidal species within its own genus. Future studies that include a greater number of species and take phylogeny into account would be beneficial in further elucidating the adaptive value of the physiological traits examined in the current study.
Conclusions
The present investigation found that the specialist intertidal triplefin species B. medius possesses a lower Pcrit than the occasional rock pool inhabitant F. lapillum, and the two exclusively subtidal species F. varium and F. malcolmi. F. lapillum also had a lower Pcrit than two closely related exclusively subtidal species (F. varium and F. malcolmi). B. medius also had a higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) than all three Forsterygion species and F. lapillum had a higher \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) than F. varium and F. malcolmi. These findings demonstrate that an enhanced capacity of the cardiorespiratory system to extract O2 from water is likely to be an important determinant of Pcrit in triplefin species which inhabit intertidal rock pools. There was, however, virtually no association between SMR and Pcrit among the species examined. This finding suggests that among triplefin fishes, a high extractive capacity for O2 is a comparatively more important determinant of Pcrit than a low resting demand for O2. Overall, triplefin species which can extract more O2 from their environment have a lower Pcrit and therefore, have the ability to meet aerobic metabolic demands at lower O2 tensions. A lower Pcrit is advantageous for an exclusively intertidal species such as B. medius, because environmental hypoxia is a routine occurrence in its rock pool habitat. High \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) also endows intertidal species with a high MS, and therefore a greater capacity to perform aerobic activities over and above maintenance requirements. Whether high MS in intertidal fish can be utilised to mitigate constraints on aerobic performance under hypoxia and high temperature, would be an interesting avenue for future research.
Abbreviations
- ACD:
-
Above chart datum
- ADP:
-
Adenosine diphosphate
- ANOVA:
-
Analysis of variance
- ANCOVA:
-
Analysis of covariance
- ATP:
-
Adenosine triphosphate
- CO2 :
-
Carbon dioxide
- Hb:
-
Haemoglobin
- Hct:
-
Haematocrit
- \( \dot{M}{\text{O}}_{2} \) :
-
Mass-specific oxygen consumption
- \( \dot{M}{\text{O}}_{{2,{ \hbox{max} }}} \) :
-
Maximum aerobic metabolic rate post-exhaustive exercise
- MS:
-
Aerobic metabolic scope
- O2 :
-
Oxygen
- P crit :
-
Critical oxygen tension
- PO2 :
-
Oxygen tension
- RBC:
-
Red blood cell
- SGA:
-
Sectioned gill arch
- SL:
-
Gill secondary lamellae
- SMR:
-
Standard metabolic rate
- WGA:
-
Whole intact gill arch
References
Behrens JW, Steffensen JF (2007) The effect of hypoxia on behavioural and physiological aspects of lesser sand eel, Ammodytes tobianus (Linnaeus, 1785). Mar Biol 150:1365–1377
Berschick P, Bridges C, Grieshaber M (1987) The influence of hyperoxia, hypoxia and temperature on the respiratory physiology of the intertidal rockpool fish Gobius cobitis Pallas. J Exp Biol 130:368–387
Bridges CR (1988) Respiratory adaptations in intertidal fish. Am Zool 28:79–96
Brix O, Clements K, Wells R (1999) Haemoglobin components and oxygen transport in relation to habitat distribution in triplefin fishes (Tripterygiidae). J Comp Physiol B 169:329–334
Chabot D, Claireaux G (2008) Environmental hypoxia as a metabolic constraint on fish: the case of Atlantic cod, Gadus morhua. Mar Pollut Bull 57:287–294
Claireaux G, Chabot D (2016) Responses by fishes to environmental hypoxia: integration through Fry’s concept of aerobic metabolic scope. J Fish Biol 88:232–251
Claireaux G, Lagardère J (1999) Influence of temperature, oxygen and salinity on the metabolism of the European sea bass. J Sea Res 42:157–168
Claireaux G, Lefrancois C (2007) Linking environmental variability and fish performance: integration through the concept of scope for activity. Philos Trans R Soc Lond B Biol Sci 362:2031–2041
Claireaux G, Webber D, Lagardère J, Kerr S (2000) Influence of water temperature and oxygenation on the aerobic metabolic scope of Atlantic cod (Gadus morhua). J Sea Res 44:257–265
Clark TD, Sandblom E, Jutfelt F (2013) Aerobic scope measurements of fishes in an era of climate change: respirometry, relevance and recommendations. J Exp Biol 216:2771–2782
Clarke A, Johnston NM (1999) Scaling of metabolic rate with body mass and temperature in teleost fish. J Anim Ecol 68:893–905
Cook DG, Wells RM, Herbert NA (2011) Anaemia adjusts the aerobic physiology of snapper (Pagrus auratus) and modulates hypoxia avoidance behaviour during oxygen choice presentations. J Exp Biol 214:2927–2934
Cook DG, Iftikar FI, Baker DW, Hickey AJ, Herbert NA (2013) Low-O2 acclimation shifts the hypoxia avoidance behaviour of snapper (Pagrus auratus) with only subtle changes in aerobic and anaerobic function. J Exp Biol 216:369–378
Cumming H, Herbert N (2016) Gill structural change in response to turbidity has no effect on the oxygen uptake of a juvenile sparid fish. Conserv Physiol 4:cow033
Diaz RJ, Breitburg DL (2009) The hypoxic environment. In: Richards JF, Farrell AP, Brauner CJ (eds) Fish physiology: hypoxia, vol 27. Academic Press, Boston, pp 2–25
Farrell AP, Richards JG (2009) Defining hypoxia: an integrative synthesis of the responses of fish to hypoxia. In: Richards JF, Farrell AP, Brauner CJ (eds) Fish physiology: hypoxia, vol 27. Academic Press, Boston, pp 487–503
Herbert N, Steffensen J (2005) The response of Atlantic cod, Gadus morhua, to progressive hypoxia: fish swimming speed and physiological stress. Mar Biol 147:1403–1412
Hickey AJR, Clements KD (2003) Key metabolic enzymes and muscle structure in triplefin fishes (Tripterygiidae): a phylogenetic comparison. J Comp Physiol B 173:113–123
Hickey AJ, Clements KD (2005) Genome size evolution in New Zealand triplefin fishes. J Hered 96:356–362
Hilton Z (2010) Physiological adaptation in the radiation of New Zealand triplefin fishes (Family Tripterygiidae). Dissertation, The University of Auckland
Hilton Z, Wellenreuther M, Clements K (2008) Physiology underpins habitat partitioning in a sympatric sister-species pair of intertidal fishes. Funct Ecol 22:1108–1117
Hilton Z, Clements KD, Hickey AJ (2010) Temperature sensitivity of cardiac mitochondria in intertidal and subtidal triplefin fishes. J Comp Physiol B 180:979–990
Keppler D, Decker K (1974) Glycogen. Determination with amyloglucosidase. In: Bergmeyer HU (ed) Methods of enzymatic analysis, 2nd edn. Academic Press, New York, pp 1127–1131
Khan J, Iftikar F, Herbert N, Gnaiger E, Hickey A (2014a) Thermal plasticity of skeletal muscle mitochondrial activity and whole animal respiration in a common intertidal triplefin fish, Forsterygion lapillum (Family: Tripterygiidae). J Comp Physiol B 184:991–1001
Khan J, Pether S, Bruce M, Walker S, Herbert N (2014b) Optimum temperatures for growth and feed conversion in cultured hapuku (Polyprion oxygeneios)—is there a link to aerobic metabolic scope and final temperature preference? Aquaculture 430:107–113
Kunzmann A (1990) Gill morphometrics of two Antarctic fish species Pleuragramma antarcticum and Notothenia gibberifrons. Polar Biol 11:9–18
Lefrançois C, Claireaux G (2003) Influence of ambient oxygenation and temperature on metabolic scope and scope for heart rate in the common sole Solea solea. Mar Ecol Prog Ser 259:273–284
Mandic M, Todgham AE, Richards JG (2009) Mechanisms and evolution of hypoxia tolerance in fish. Proc Biol Sci 276:735–744
Mandic M, Speers-Roesch B, Richards JG (2012) Hypoxia tolerance in sculpins is associated with high anaerobic enzyme activity in brain but not in liver or muscle. Physiol Biochem Zool 86:92–105
Matey V, Richards JG, Wang Y, Wood CM, Rogers J, Davies R, Murray BW, Chen XQ, Du J, Brauner CJ (2008) The effect of hypoxia on gill morphology and ionoregulatory status in the Lake Qinghai scaleless carp, Gymnocypris przewalskii. J Exp Biol 211:1063–1074
McArley TJ, Hickey AJ, Herbert NA (2017) Chronic warm exposure impairs growth performance and reduces thermal safety margins in the common triplefin fish (Forsterygion lapillum). J Exp Biol 220:3527–3535
Nilsson GE, Östlund-Nilsson S (2008) Does size matter for hypoxia tolerance in fish? Biol Rev 83:173–189
Norin T, Clark T (2016) Measurement and relevance of maximum metabolic rate in fishes. J Fish Biol 88:122–151
Norin T, Malte H, Clark TD (2014) Aerobic scope does not predict the performance of a tropical eurythermal fish at elevated temperatures. J Exp Biol 217:244–251
Pörtner HO, Knust R (2007) Climate change affects marine fishes through the oxygen limitation of thermal tolerance. Science 315:95–97
Richards JG (2011) Physiological, behavioral and biochemical adaptations of intertidal fishes to hypoxia. J Exp Biol 214:191–199
Richards J, Wang Y, Brauner C, Gonzalez R, Patrick M, Schulte P, Choppari-Gomes A, Almeida-Val V, Val A (2007) Metabolic and ionoregulatory responses of the Amazonian cichlid, Astronotus ocellatus, to severe hypoxia. J Comp Physiol B 177:361–374
Rogers NJ, Urbina MA, Reardon EE, McKenzie DJ, Wilson RW (2016) A new analysis of hypoxia tolerance in fishes using a database of critical oxygen level (P crit). Conserv Physiol 4:cow012
Schulte PM (2015) The effects of temperature on aerobic metabolism: towards a mechanistic understanding of the responses of ectotherms to a changing environment. J Exp Biol 218:1856–1866
Schurmann H, Steffensen J (1997) Effects of temperature, hypoxia and activity on the metabolism of juvenile Atlantic cod. J Fish Biol 50:1166–1180
Scott GR, Wood CM, Sloman KA, Iftikar FI, De Boeck G, Almeida-Val VM, Val AL (2008) Respiratory responses to progressive hypoxia in the Amazonian oscar, Astronotus ocellatus. Respir Physiol Neurobiol 162:109–116
Silberschneider V, Booth DJ (2001) Resource use by Enneapterygius rufopileus and other rockpool fishes. Environ Biol Fish 61:195–204
Snyder S, Nadler L, Bayley J, Svendsen MBS, Johansen J, Domenici P, Steffensen JF (2016) Effect of closed v. intermittent-flow respirometry on hypoxia tolerance in the shiner perch Cymatogaster aggregata. J Fish Biol 88:252–264
Speers-Roesch B, Mandic M, Groom DJE, Richards JG (2013) Critical oxygen tensions as predictors of hypoxia tolerance and tissue metabolic responses during hypoxia exposure in fishes. J Exp Mar Biol Ecol 449:239–249
Steffensen JF (1989) Some errors in respirometry of aquatic breathers: how to avoid and correct for them. Fish Physiol Biochem 6:49–59
Truchot J, Duhamel-Jouve A (1980) Oxygen and carbon dioxide in the marine intertidal environment: diurnal and tidal changes in rockpools. Respir Physiol 39:241–254
Wellenreuther M (2007) Ecological factors associated with speciation in New Zealand triplefin fishes (Family Tripterygiidae). Dissertation, The University of Auckland
Wellenreuther M, Barrett PT, Clements KD (2007) Ecological diversification in habitat use by subtidal triplefin fishes (Tripterygiidae). Mar Ecol Prog Ser 330:235–246
Wells R, Baldwin J, Seymour R, Christian K, Farrell A (2007) Air breathing minimizes post-exercise lactate load in the tropical Pacific tarpon, Megalops cyprinoides (Broussonet 1782) but oxygen debt is repaid by aquatic breathing. J Fish Biol 71:1649–1661
Acknowledgements
T.J.M. would like to acknowledge support from the University of Auckland Scholarship Office for doctoral funding. Supplementary funds were also provided by a Marsden Grant awarded to A.J.R.H and N.A.H. (UOA1407: How to avoid brain damage during oxygen deprivation? Intertidal fish provide a unique test model). Technical staff Peter Browne and Errol Murray are acknowledged for their assistance with design and fabricating the respirometry setups and John Atkins is acknowledged for design of the respirometry software. Dr. Nicholas Shears and Dr. Caitlin Blain are thanked for allowing us to use of their oxygen and temperature loggers. We also thank three anonymous reviewers for their effort in providing comments and critiques which were helpful in revising an original version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
All procedures performed in studies involving animals were in accordance with the ethical standards of the institution or practice at which the studies were conducted (University of Auckland AEC approval number 001441).
Data availability
The data generated during and/or analysed during the current study are available from the corresponding author on reasonable request.
Additional information
Communicated by H. V. Carey.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
McArley, T.J., Hickey, A.J.R., Wallace, L. et al. Intertidal triplefin fishes have a lower critical oxygen tension (Pcrit), higher maximal aerobic capacity, and higher tissue glycogen stores than their subtidal counterparts. J Comp Physiol B 189, 399–411 (2019). https://doi.org/10.1007/s00360-019-01216-w
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-019-01216-w