
Abstract
Freeze tolerance and freeze avoidance are typically described as mutually exclusive strategies for overwintering in animals. Here we show an insect species that combines both strategies. Individual fungus gnats, collected in Fairbanks, Alaska, display two freezing events when experimentally cooled and different rates of survival after each event (mean ± SEM: −31.5 ± 0.2°C, 70% survival and −50.7 ± 0.4°C, 0% survival). To determine which body compartments froze at each event, we dissected the abdomen from the head/thorax and cooled each part separately. There was a significant difference between temperature levels of abdominal freezing (−30.1 ± 1.1°C) and head/thorax freezing (−48.7 ± 1.3°C). We suggest that freezing is initially restricted to one body compartment by regional dehydration in the head/thorax that prevents inoculative freezing between the freeze-tolerant abdomen (71.0 ± 0.8% water) and the supercooled, freeze-sensitive head/thorax (46.6 ± 0.8% water).
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The fungus gnat Exechia nugatoria Johannsen (1912) (Diptera: Mycetophilidae) found in Fairbanks, Alaska, survives the winter as an adult above the snow under the bark of dead trees, a microhabitat that does not significantly insulate individuals from fluctuating ambient conditions, including low temperatures that can decline below −50°C. Fungus gnats are a diverse and abundant group of insects that can be recognized by their hump-backed appearance, stout and elongate coxae, and well-developed tibial spurs. They often attain large population sizes and play an important role in the food web within the forest environments where they reside. Mycetophilidae are the most diverse in the northern Palearctic Region (Jaklovlev and Siitonen 2004), and in cold climates many members are known to overwinter as adults, particularly those of the tribe Exechiini. Their overwintering sites include caves (Kjaerandsen 1993; Kurina 1996; Hedmark 2000), hollow stems of umbelliferous plants (Väïsänen 1981), and under the bark of dead trees (Hedmark 2000). In these studies, microhabitat temperatures were found to be relatively moderate (approximately −6°C).
In extreme cold climates such as the interior of Alaska, insect species either avoid or tolerate freezing to survive winter. Insects may behaviorally seek microhabitats that ameliorate low subzero temperature. Freeze-avoiding species cease feeding and eliminate the contents of their gut to promote supercooling of body fluids to temperatures well below their equilibrium freezing point (Zachariassen 1985). In addition, these insects must remove ice-nucleating factors, prevent inoculative freezing by production of noncolligative antifreeze proteins (AFPs) (Duman 2001), and/or synthesize colligative antifreezes such as glycerol and other polyols (Storey and Storey 1991). In contrast, freeze-tolerant insects withstand freezing of extracellular water, often by inducing freezing at high subzero temperatures, typically −5 to −12°C, via ice-nucleating factors (Zachariassen 1985; Duman 2001); furthermore, in Alaska some freeze-tolerant insects can be found in microhabitats such as on vegetation that lies above the snow where exposure to low ambient conditions is relatively unimpeded (Miller 1982). Consequently, these individuals are often exposed to temperatures that can cause ice nucleation. In studies that included fungus gnats in Alaska, Miller (1978, 1982) showed that Mycetophila sp. and Exechia sp. in Fairbanks are freeze tolerant but supercool to −30°C, a low temperature that is more typically associated with freeze-avoiding insects. He did not note dual freezing events in these gnats. He did describe two nucleation events occurring in a Lepidopteran (Martyrhilda ciniflonella, at −25 and −36°C) and a Neuropteran (Hemerobius simulans, at −15 and −36°C), although he did not report on differential survival of individuals between the two freezing events.
We present evidence that freeze tolerance and freeze avoidance in individual E. nugatoria are not mutually exclusive. We report on adults that survive initial freezing near −30°C but do not survive a second freezing event near −50°C. Understanding how these flies survive low temperature conditions may help explain how they have become wide spread in cold regions.
Materials and methods
Insect collection and microhabitat characteristics
We collected adult E. nugatoria November 2007–December 2007 from large aggregates found under bark of dead and dry standing Poplar spp. in Fairbanks, Alaska, near the Fairbanks International Airport. We estimate that some trees housed greater than 1,000 individuals. All insects were located above the snow. Insects were transported in cold insulated containers and tested in the laboratory for freezing responses either within 1 h of collection or after 10 days of acclimation at 4°C in 100% relative humidity (RH).
On the north side of a tree at the collecting site, temperatures of the air (2.2 m above ground) and microhabitat (under the bark at 1 m above ground) were monitored using Hobo Pro Series data loggers and downloaded with BoxCar Pro 4 software (Onset Computer Corporation, Bourne, MA, USA).
Supercooling
To determine supercooling points of gnats, a single copper-constantan thermocouple junction (36 g) was rested against an individual. The junction and insect were placed in closed 0.6-ml plastic vials submersed in alcohol–water baths that were cooled from 0°C at 0.2°C/min. Thermocouples leads were attached to a computer controlled multi-channel thermocouple thermometer (Iso-Thermex, Columbus Instruments, Columbus, OH, USA) that recorded temperature every 5-s. The lowest body temperature recorded at the onset of freezing, as evidenced by an exotherm (a transient rise in temperature due to the release of the latent heat of fusion), is the supercooling point (SCP).
Since we found that these insects displayed dual exotherms, we investigated which body compartments were responsible for each freezing event by dissecting with scissors the abdomen from the head/thorax (including wings, legs, and antennae), and cooling each body compartment in separate tubes. If either halter, the reduced second pair of wings used for balance, was accidentally severed, a new individual was used. Immediately after dissection, each body part was placed in a tube so that it rested against a thermocouple using a microscope to ensure the body part was in contact with the thermocouple junction. Since, this visual inspection showed little to no fluid loss, the dissected parts were not sealed with oil.
To ensure that the second exotherm in whole insects was not dependent on the first, a supercooling run was performed in March 2008 in which the bath was programmed to hold at temperatures below the first supercooling point (approximately −34°C) for 48 h. After 48 h, the bath was programmed to cool to −60°C to record a possible second exotherm.
Survival
To assess immediate survival of gnats removed from the field, insects were collected on several occasions in November and December 2007. They were brought to the lab and placed on moist towels at 4°C and 100% RH to assess survival (return of coordinated movements) over a 1-week period. To determine whether flies survived the first freezing event, they were cooled to approximately −35°C on 20 and 30 November and 18 and 21 December 2007 and then warmed to 0°C. Both cooling and warming rates were conducted at 0.2°C/min. Individuals were held overnight on ice and then transferred to a container at 4°C and 100% RH. Similarly, to determine whether flies survived the second freezing event, they were cooled to approximately −58°C on 20 and 30 November 2007 then rewarmed as above.
Water content
Masses of individual larvae (whole body) were determined to the nearest 0.1 mg. Larvae were then dried at 60°C for 5 days to constant mass. Absolute body water was calculated as milligram per milligram dry mass and termed water content (WC) (Hadley 1994). Water contents of dissected body parts (the abdomen vs. the head/thorax) were similarly determined to the nearest 0.01 mg.
Results
Microhabitat temperatures varied widely from transient highs above 0°C to lows of –43°C during this study, and air and under-bark microhabitat temperatures were similar (Fig. 1; mean difference = 0.2°C; n = 8281). From 1 December 2007 to 28 March 2008, mean temperature was −18.1 ± 0.1°C (n = 5710, recorded every 30 min).

Microhabitat (under bark) temperature 1 m above the ground (red) and air temperature 2.2 m above the ground (black) at the fungus gnat collecting site
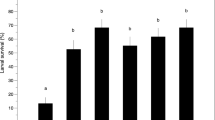
Directly after collection from the field, gnats (fresh mass = 1.7 ± 0.4 mg, n = 78) froze at an initial supercooling point (SCP1) of −31.5 ± 0.2°C (Fig. 2a); 70.2% of these were alive after thawing, a survival rate similar to that in field-collected insects that were assessed for movement after direct transfer to the lab and warming to 4°C (Table 1; combined average 85%). Gnats cooled below SCP1 showed a second exotherm (SCP2) at −50.7 ± 0.4°C (Fig. 2a) that was 100% lethal (Table 1).

Mean (±SEM) supercooling (a) and water content (b) in fungus gnats brought in from the field or after exposure to 4°C for 10 days. Within parentheses are percent water and/or sampling number. Different letters indicate significant differences in means. In a, SCPs of fungus gnats immediately after they were collected and transferred to the laboratory: Shapiro–Wilk normality test, P < 0.05; Wilcoxon two-sample test, S = 1653.0, P < 0.0001. Abdomen versus head/thorax is significantly different: Shapiro–Wilk test, P > 0.05; t test with unequal variance, t = −17.60, P = 0.0001, df = 35.6. SCP1 (collected vs. dissected): Shapiro–Wilk normality test, P < 0.05; Wilcoxon rank-sum, S = 666.0, P = 0.3301. SCP2 (collected vs. dissected): Shapiro–Wilk normality test, P < 0.05; Wilcoxon rank-sum, S = 355.0, P = 0.1813. In b, whole body versus whole body (acclimated) water content, Shapiro–Wilk normality test, P < 0.05; Wilcoxon rank-sum, S = 1049.0, P < 0.0001. For body compartments, Shapiro–Wilk test, P < 0.05; Wilcoxon rank-sum, S = 465, P < 0.0001
The dissected body compartments of newly collected individuals were cooled as above to determine which body segments were responsible for SCP1 and SCP2. Supercooling point values of dissected gnat abdomens averaged −30.1 ± 1.1°C and were statistically indistinguishable from average SCP1 values from intact gnats (Fig. 2a). Supercooling values of dissected head/thoraces were −48.7 ± 1.3°C and did not differ from the average SCP2 in intact insects (Fig. 2a). These results suggest that freezing of the abdomen is responsible for SCP1, and freezing of the head/thorax complex is responsible for SCP2 in intact gnats. After 10 days of acclimation at 4°C and 100% RH, whole gnats froze at −20.6 ± 0.6°C, and all died (Table 1). A second exotherm was not observed in these acclimated animals when they were cooled to −60°C (Fig. 2a).
Insects in the field are regularly exposed to temperatures below levels that are experimentally associated with SCP1 (Fig. 1). For example, insects were collected after a cold period 3–9 February 2008 (Fig. 1) when mean ambient temperature was −39.2 ± 0.1°C (n = 282), with extremes of −31 and −43°C. Survivorship was 91% (64/70) when insects were brought into the laboratory on 18 February. This suggests that fungus gnats naturally survive the freezing of their abdomens and that, since freezing of the thorax and head is lethal, abdominal ice does not inoculate freezing in anterior body compartments, even when exposed to temperatures lower than SCP1 for several consecutive days.
We also demonstrated that SCP2 is not dependent on SCP1. On 28 March 2008, the bath was programmed to hold at approximately −34.5°C for 48 h. After this hold, the bath was decreased to −60°C to trigger SCP2. Although as early as 23 March gnats appeared to be acclimating to spring conditions and SCP2 increased to −42 ± 1.3°C (n = 16), there was a statistically significant difference (P < 0.05; t test with equal variance, t = 5.50, P = < 0.001, df = 24) in SCP1 versus SCP2 (−33.2 ± 0.6°C, n = 16 vs. −39.2 ± 0.8°C, n = 10, respectively) with 10/16 individuals exhibiting SCP2. We believe that if this experiment had been carried out when gnats were still under winter field conditions, SCP2 would have been lower, and all would have displayed a second exotherm.
Discussion
Exechia nugatoria in winter displays two freezing events when experimentally cooled; they survive the first that occurs at temperatures of approximately −32°C, but they do not survive the second freezing event that occurs at approximately −51°C. Testing body compartments separately suggests that it is the abdomen that freezes first while the head/thorax freezes second. Since fungus gnats overwintering in Alaska routinely survive temperatures between −30 and −40°C, they must in nature become partially frozen and prevent the inoculation of ice from occurring between adjoining body segments.
One question prompted by our findings is why dual freezing events evolved in this species. Lundheim and Zachariassen (1993) and Zachariassen et al. (2004, 2008) suggest that some freeze-tolerant insects that have greater trans-cuticular water permeabilities than related freeze-avoiding species conserve body water and avoid dehydration while frozen because they are in vapor pressure equilibrium with ice in the surrounding environment. Since the overwintering site of E. nugatoria is a cold, dry microhabitat, an advantage of maintaining ice in the abdomen may be to lessen evaporative water loss over the course of a long winter. Not only would abdominal water be conserved but also some head/thoracic water vapor may be directed to the abdomen, rather than being lost to the environment. If the entire body froze, however, water loss would be even less. Freezing of the head and thorax may be avoided due to an increased susceptibility of neural tissue to damage from freezing as has been suggested to occur in other freeze-tolerant insects (Collins et al. 1997; Yi and Lee 2003). The large changes in ion concentrations in the remaining unfrozen fraction that result from extracellular freezing may not be tolerated by certain neurons or other cells of the anterior central nervous system.
A second obvious question arising from this study is how is it possible for the head/thorax to remain unfrozen while water in the thorax is apparently in direct contact with the frozen abdomen? Freshly dissected abdomens contained approximately 71% water, and heads/thoraces had 47% water, either due to proportional differences in hard body parts within body compartments or the selective withdrawal of water from the thorax during freezing. Dehydration decreases SCPs in freeze-avoiding insects (Bigg 1953; Zachariassen 1985; Bennett et al. 2005), and resistance to inoculative freezing increases with dehydration in the Antarctic midge Belgica antarctica (Elnitsky et al. 2008); both of these relationships could contribute to the stability of partial freezing in the fungus gnat. While survival of an individual that both tolerates and avoids freezing has not been noted previously in insects, it occurs in plants, although the underlying mechanisms are not satisfactorily understood (Wisniewski 1995; Quamme 1995). Two exotherms are common in the stems of many species of trees and typically consist of a high temperature exotherm (HTE) between −7 and −13°C and a low temperature exotherm (LTE) below −40°C (Quamme et al. 1972; George et al. 1974). In stems of hardwood and softwood trees from temperate regions, the HTE is due to freezing of extracellular water, especially in the xylem, while the LTE represents freezing in freeze-sensitive xylem ray parenchyma. Consequently, avoidance of the LTE in the freeze-sensitive parenchymal tissue permits adaptation to relatively low temperatures in these trees but also imposes limits on their ranges in terms of latitude, low temperature extremes (George et al. 1974), and altitude (Becwar et al. 1981). Recently, freeze-avoiding xylem ray parenchyma of the katsura tree Cercidophyllum japonicum is reported to have flavonol glycosides with anti-ice nucleation activity that promote supercooling (Katsuga et al. 2008). Supercooling of freeze sensitive flower and leaf buds that are surrounded by frozen extracellular water is also common in trees (Quamme 1995). In conifers, after the extracellular water in tolerant tissues freezes, primordial shoot water migrates toward the frozen tissue where it subsequently freezes. Sakai (1979) describes this as ‘extraorgan freezing’ that allows the freeze-intolerant primordium to supercool to lower temperatures due to the removal of water. Cary (1985), working on Prunis (peach and prune) flowers, modeled supercooling in floral tissue with ice present, showing that water vapor flowing toward ice crystals may “cause a discontinuity in the liquid phase” that acts as a barrier to nucleation. The abdominal freezing of the fungus gnat may function as “extraorgan freezing” allowing water to be withdrawn from the thorax to further decrease the supercooling point. In fungus gnats, abdominal freezing (−30°C) and a subsequent movement of water vapor from the head/thorax may create a dry area (discontinuity) as a barrier against inoculative freezing of the remaining supercooled fluid in the thorax. This mechanism is dependent on a freezing event, yet our observations of water content differences were on unfrozen insects. These may have previously frozen and the differences in water content remained after thawing. Even though the gnats display a difference in water content after they have rewarmed, the gnat acclimation increases total body water through rehydration of the head/thorax that also results in high, lethal SCPs and the absence of SCP2. With the slight but significant increase in whole-body water content, there is an increase in the temperature of the first freezing event and elimination of the second and no gnat survives.
This study presents a unique example of a mixed overwintering strategy in a single insect, tolerating freezing in the abdomen while avoiding freezing in the thorax and head. While a similar mixed overwintering strategy is more typical in plants, this is the first arthropod known to overwinter in this manner. At present, the mechanism(s) underlying this strategy and rationale for its evolution remain unknown.
Abbreviations
- SCP1:
-
Supercooling point 1
- SCP2:
-
Supercooling point 2
- RH:
-
Relative humidity
- WC:
-
Water content
References
Becwar MR, Rajashekar C, Hansen-Bristow KJ, Burke MJ (1981) Deep supercooling of tissue water and winter hardiness limitations in timberline flora. Plant Physiol 68:111–114
Bennett VA, Sformo T, Walters K, Toien Ø, Jeannet K, Hochstrasser R, Pan Q, Serianni AS, Barnes BM, Duman JG (2005) Comparative overwintering physiology of Alaska and Indiana populations of the beetle Cucujus clavipes (Fabricius): roles of antifreeze proteins, polyols, dehydration and diapause. J Exp Biol 208:4467–4477
Bigg EK (1953) The supercooling of water. Proc Phys Soc London B66:688–694
Cary JW (1985) Freeze survival in peach and prune flowers. Plant Sci Lett 37:265–271
Collins SD, Allenspach AL, Lee RE (1997) Ultrastructural effects of lethal freezing on brain, muscle and Malpighian tubules from freeze-tolerant larvae of the gall fly, Eurosta solidaginis. J Insect Physio 43:39–45
Duman JG (2001) Antifreeze and ice nucleator proteins in terrestrial arthropods. Ann Rev Physio 63:327–357
Elnitsky MA, Benoit JB, Lee RE, Denlinger DL (2008) Desiccation tolerance and drought acclimation in the Antarctic collembolan Cryptopygus antarcticus. J Insect Physiology 54:1432–1439
George MF, Burke MJ, Pellet HM, Johnson AG (1974) Low temperature exotherms and woody plant distributions. HortScience 9:519–522
Hadley NF (1994) Water relations of terrestrial arthropods. Academic Press, New York
Hedmark K (2000) Svampmyggor i taigan—nya arter för Sverige i ett fennoskandiskt perspectiv (Diptera: Siaroidea exkl Sciaridae). Entomologisk Tidskrift 121:73–89
Jaklovlev J, Siitonen J (2004) Finnish fungus gnats (Diptera Mycetophilidae etc.): faunistics, habitat requirements and threat status. Lammi Notes 30:3–7
Johannsen OA (1912) The Mycetophilidae of North America. Part IV. Maine Agric Exp Station Bull 200:57–146
Katsuga J, Hashidko Y, Nishioka A, Yoshiba M, Arakawa K, Fujikawa S (2008) Deep supercooling xylem parenchyma cells of katsura tree (Cercidiphyllum japonicum) contain flavonol glycosides exhibiting high anti-ice nucleation activity. Plant Cell Environ 31:1335–1348
Kjaerandsen J (1993) Diptera in mines and other cave systems in southern Norway. Entomologica Fennica 4:151–160
Kurina O (1996) Hibernation of fungus gnats (Diptera: Mycetophilidae) in Estonian caves. Studia Dipterologica 3:221–229
Lundheim R, Zachariassen KE (1993) Water balance of over-wintering beetles in relation to strategies for cold tolerance. J Comp Physiol B 163:1–4
Miller LK (1978) Physiological studies of arctic animals. Comp Biochem Physio 59A:327–334
Miller K (1982) Cold hardiness strategies of some adult and immature insects overwintering in interior Alaska. Comp Biochem and Physio 73A:595–604
Quamme HA (1995) Deep supercooling in buds of woody plants. In: Lee RE, Warren GJ, Gusta LV (eds) Biological ice nucleation and its applications. American Phytopathological Society Press, St. Paul, pp 183–199
Quamme HA, Stushnoff C, Weiser CJ (1972) The relationship of exotherms to cold injury in apple stem tissues. J Amer Soc Hort Sci 97:608–613
Sakai A (1979) Freeze avoidance mechanism of the primordial shoots of conifer buds. Plant Cell Physiol 20:1381–1390
Storey KB, Storey JM (1991) Biochemistry of cryoprotectants. In: Denlinger D, Lee RE (eds) Insects at low temperature. Chapman and Hall, New York, pp 64–93
Väïsänen R (1981) Umbelliferous stems as overwintering sites for Mycetophilidae (Diptera) and other invertebrates. Notulae Entomologicae 61:165–170
Wisniewski M (1995) Deep supercooling in woody plants and the roles of cell wall structure. In: Lee RE, Warren GJ, Gusta LV (eds) Biological ice nucleation and its applications. American Phytopathological Society Press, St. Paul, pp 163–181
Yi S-X, Lee RE (2003) Detecting freeze injury and seasonal cold-hardening of cells and tissues in the gall fly larvae, Eurosta solidaginis (Diptera: Tephritidae) using fluorescent vital dyes. J Insect Physio 49:999–1004
Zachariassen KE (1985) Physiology of cold tolerance in insects. Physiol Rev 65:799–832
Zachariassen KE, Perersen SA, Kristiansen E (2004) Advantages and disadvantages of freeze-tolerance and freeze-avoidance overwintering strategies. In: Barnes BM, Carey HV (eds) Life in the cold: evolution mechanisms adaptation, and application. Institute of Arctic Biology, Fairbanks, pp 283–291
Zachariassen KE, Li NG, Laugsand AE, Kristiansen E, Pedersen SA (2008) Is the strategy for cold hardiness in insects determined by their water balance? A study on two closely related families of beetles: Cerambycidae and Chrysomelidae. J Comp Physiol B 178:977–984
Acknowledgments
This study was supported by National Science Foundation grants IOB06-18436 to BMB and IOB06-18342 to JGD. We wish to thank B. Tudor, K. Walters (Notre Dame), D. Wagner (UAF) for use of scales, G. Juday (UAF) and R. Lee (Miami University, Ohio) for critical discussions, and D. Sikes (UAF Museum) for initial insect identification.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by H.V. Carey.
Rights and permissions
About this article
Cite this article
Sformo, T., Kohl, F., McIntyre, J. et al. Simultaneous freeze tolerance and avoidance in individual fungus gnats, Exechia nugatoria . J Comp Physiol B 179, 897–902 (2009). https://doi.org/10.1007/s00360-009-0369-x
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00360-009-0369-x