
Abstract
Gibberellic acid (GA) and sulfur (S) have been known to modulate physiological processes of plants in normal and stressful conditions. Cultivars of mungbean (Vigna radiata L.), Pusa Vishal, PusaRatna, Pusa 9531, Ganga, and MH 318 were screened for photosynthetic S-use efficiency (p-SUE) and growth in presence of 200 mg Cd kg−1 soil. Pusa 9531 showed maximum p-SUE, growth, and Cd tolerance. The mechanism of 10 µM GA-induced alleviation of Cd stress in Cd tolerant cultivar Pusa 9531 grown with 100 mg S kg−1 soil (100S) or 200 mg S kg−1 soil (200S) was investigated. Plants receiving GA in presence of 100S maximally utilized available S and improved photosynthetic characteristics of plants through improvement in SUE and antioxidant metabolism and alleviated Cd stress; the addition of 200S was of no additional benefit. Such effects of GA and 100S in protection of photosynthetic performance and growth involved nitric oxide (NO), which was evidenced on the use of NO modulators: 100 µM SNP (sodium nitropruside; NO promoter) or 100 µM c-PTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide; NO scavenger). The application of SNP improved the mechanisms of N- and S-assimilation and antioxidant metabolism which helped in tolerance of plants to Cd stress. Moreover, c-PTIO supplementation to GA plus S reversed the positive effects of GA and S on photosynthesis and growth, signifying the involvement of NO in mechanisms induced by GA + S under Cd stress. The study provides the evidence that GA and NO interact and mechanisms induced by GA in presence of S were mediated by NO. The inter-relation of GA and NO may be used for augmenting photosynthesis and growth through utilization of S in mungbean grown under Cd stress.
Similar content being viewed by others

Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Cadmium (Cd) is one of the most toxic heavy metals with much longer biological half-life (more than 20 years) and is highly toxic even at low concentration (Wagner 1993). The concentration of cadmium has been increasing at an extremely faster rate (Bolan et al. 2000), which is alarming in many countries. Its presence in soil has invoked the unprecedented rise in agricultural land deterioration and toxicity in plants (Kavamura and Esposito 2010; Nazar et al. 2012; Asgher et al. 2014). It is accumulated in living tissues, thus perturbing the food chain (Choppala et al. 2014; Wahid and Khaliq 2015). The accumulation of Cd in plant cells induces physiological, biochemical, and molecular mechanisms (Emamverdian et al. 2015; Singh et al. 2016), with regulatory networks with other signaling molecules (Dalcarso et al. 2010; Asgher et al. 2014; Masood et al. 2012; 2016). The inhibition of growth and photosynthesis with Cd accumulation has been reported in Lepidium sativum (Gill et al. 2012) and Brassica juncea (Mobin and Khan 2007; Nazar et al. 2012; Masood et al. 2012; Asgher et al. 2014). Accumulation of high levels of cadmium disrupts photosynthetic machinery and structure of chloroplast, affects nutrient uptake and their assimilation, and negatively influences biosynthesis of chlorophyll (Mobin and Khan 2007; Masood et al. 2012; Choppala et al. 2014). The physiological performance of plants is highly adversely affected due to the buildup of reactive oxygen species (ROS) generated in high Cd levels. Excessive ROS generation due to high Cd levels disturbs the redox state, disrupts the cellular and membrane integrity, and limits the functionality of several proteins and plant homeostasis (Noriega et al. 2007; Singh et al. 2016). Plant growth reduction was related to inhibition of carbon assimilation (Masood et al. 2012; Nazar et al. 2012; Khan et al. 2015) and ribulose 1,5-bisphosphate carboxylase/oxygenase (Rubisco) activity (Asgher et al. 2014). Cd restricts the acquisition of various essential mineral ions such as magnesium (Mg), potassium (K), phosphorus (P), calcium (Ca), and iron (Fe), thus developing their deficiency in the affected plants (Metwally et al. 2005; Nazar et al. 2012) and also obstructs the metabolism and interaction among carbon (C), sulfur (S) and nitrogen (N) (Harada et al. 2002; Gill et al. 2012; Nazar et al. 2012; Asgher et al. 2014; Khan et al. 2015). Plants tend to develop various protective strategies to dissipate the toxic effects of Cd-induced oxidative stress by potentiating the defense mechanisms (Anjum et al. 2016; Singh et al. 2016). The supplementation of phytohormones and maintaining plant nutritional status are some of the best sustainable stress alleviation strategies adopted. Sulfur is a macronutrient that stands fourth after N, P, and K for its importance in regulation of plant growth and developmental processes. It plays a critical role in optimization of photosynthetic process under normal as well as stress conditions (Nazar et al. 2012; Khan et al. 2015) being an integral component in the electron transport chain and iron–sulfur clusters (Marschner 1995). The deficiency of S in plants hampers N-assimilation and metabolism and thereby negatively affects growth, photosynthesis, and dry matter production (Tandon 1995; Kopriva et al. 2002; Nazar et al. 2012) and reduces content of chlorophyll and Rubisco (Lunde et al. 2008). Sulfur is an important part of amino acids, cysteine (Cys), and methionine (Met), non-enzymatic antioxidants and reduced glutathione (GSH), glucosinolates, and Fe–S clusters which played significant role in mitigation of Cd-induced stress (Anjum et al. 2008; Khan et al. 2016). Previous report by Masood et al. (2016) showed that S supplementation accelerated the activity of antioxidant metabolism leading to improved photosynthetic performance in mustard under Cd stress. It has been shown that accumulation of thiol compounds (Cys, GSH), activity of enzymes involved in ascorbate–glutathione, and interaction with S and N and S-use efficiency in plant cells imparted tolerance to the toxic Cd concentration (Nocito et al. 2002; Sun et al. 2007; Khan et al. 2009; Hossain et al. 2012; Lancilli et al. 2014; Masood et al. 2016).
Phytohormones are recognized as important signaling molecules in abiotic stress management (Wani et al. 2016). Gibberellins (GA) are plant hormones involved in diverse functions in plants, such as improving growth and photosynthesis and maintaining source–sink relationship under optimal and stressful environments (Iqbal et al. 2011). GA signaling is involved in adjustment of plants through interaction with nutrients or other hormones under limiting environmental conditions (Masood et al. 2012, 2016; Masood and Khan 2013). Our earlier studies have shown that GA application improved leaf area index, carbonic anhydrase activity, plant dry mass (Khan 1996; Khan et al. 1996), nutrients uptake, and yield efficiency in mustard (Khan et al. 1998, 2002, 2005). The ameliorative role of GA in reducing toxicity of other metal stress such as chromium in pea seedlings (Gangwar et al. 2010) and nickel in mungbean (Ali et al. 2015) has been reported. The reports on the ameliorative role of GA in plants under Cd stress are also available (Rubio et al. 1994; Picazo et al. 2007; Meng et al. 2009; Masood and Khan 2013; Masood et al. 2016). Recently, nitric oxide (NO) has become a subject of extensive research regarding its role in the management of biotic as well as abiotic stresses in plants (Singh et al. 2008; Xiong et al. 2009; Asgher et al. 2016). NO regulates cellular functions both positively and negatively which is dependent on its concentration and cellular location (Leitner et al. 2009; Gill et al. 2013). Asgher et al. (2016) reported that NO counteracts oxidative stress by interacting with several other phytohormones and signaling molecules. It has been reported that NO level increased endogenously under Cd stress, while some reported a decrease in NO level (Asgher et al. 2016). Previous reports have shown the role of NO in counteracting Cd-induced reduction of growth and photosynthesis through increase in the enhancement of efficiency of antioxidant system and sequestration of uncontrolled ROS generated (Hsu and Kao 2004; Per et al. 2017).
The present study was conducted to connect our findings that GA and S ameliorates Cd stress through enhanced S-assimilation and S-metabolites production (Masood and Khan 2013; Khan et al. 2015; Masood et al. 2016), and NO has role in N- and S-assimilation in plants grown under salt stress (Fatma et al. 2016; Sehar et al. 2019; Jahan et al. 2020) and Cd stress (Per et al. 2017). As there is crosstalk between NO and other phytohormones (Asgher et al. 2016), there could be an interaction between NO and GA in regulating use efficiency of S, antioxidant system, and Cd toxicity. It was expected that NO participated in the effects of GA and S in enhancing the production of thiols, antioxidant defense system, minimizing the toxicity generated by ROS, and restoration of growth and photosynthetic potential in mungbean.
Materials and Methods
Plant Material and Growth Conditions
Seeds of five cultivars of mungbean (Vigna radiata L.), Pusa Vishal, PusaRatna, Pusa 9531, Ganga, and MH 318, were obtained from Indian Agricultural Research Institute, New Delhi, India. The seeds were surface sterilized with HgCl2 (0.01%) and washed repeatedly with double-distilled water following sterilization. The seeds were then sown in earthen pots of 23 cm diameter containing 5 kg of sandy loam soil.
The five cultivars of mungbean were tested for Cd tolerance by raising plants with 200 mg Cd kg−1 soil applied as CdCl2 at the time of sowing on the basis of photosynthetic and growth parameters. On the basis of screening experiment, cultivar Pusa 9531 (selected as most tolerant cultivar) was grown individually with 10 µM GA or 100 mg S kg−1 soil (100S) and 200 mg S kg−1 soil (200S) or in combination in presence or absence of 200 mg Cd kg−1 soil to assess the effects of GA and S in the maximum Cd stress alleviation. Further, in another experiment, the role of NO was assessed in GA and 100S (S)-mediated alleviation of Cd stress using promoter or inhibitor of NO. Plants were individually treated with 100 µM sodium nitroprusside (SNP; NO source or 100 µM 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide (c-PTIO; NO scavenger). SNP was given with Cd or Cd + S. GA was given with Cd plus S, and c-PTIO was supplemented with the combined dose of GA plus 100S. The mechanisms of S-assimilation and antioxidants were evaluated for providing Cd stress tolerance and restoration of growth and photosynthesis. The elemental sulfur was used as source of S applied basally 15 d prior to sowing. Spray treatments of GA, NO, or c-PTIO included 0.5% teepol as surfactant. The pots were irrigated with 300 ml of distilled water every day. The treatments (each with three replicates) were arranged in randomized block design and pots were kept in the natural conditions of day/night in net house of the Department of Botany, Aligarh Muslim University Aligarh (India) with average day/night temperatures of 35/22 ± 3 °C, photosynthetically active radiation (PAR) of 600 ± 35 µmol m−2 s−1, and relative humidity of 64–75%. Samples were collected for various growth, physiological, and biochemical studies at 30 d after sowing (DAS).
Determination of Photosynthetic Parameters and Growth
For photosynthetic parameters, leaf chlorophyll content was recorded by SPAD chlorophyll meter (SPAD 502 DL PLUS, Spectrum Technologies). Gas exchange parameters (stomatal conductance, intercellular CO2 concentration and net photosynthesis) were measured in the leaves of plants that were fully expanded using infrared gas analyzer (CID-340, Photosynthesis System, Bio-Science, USA). The activity of Rubisco was determined by monitoring NADH oxidation at 30 °C at 340 nm when 3-phosphoglycerate is converted into glycerol-3-phosphate after addition of enzyme extract to the reaction mixture. The method adopted and detailed procedure are given in Jahan et al. (2020). Data on net photosynthesis and S content in unit leaf area were used to calculate photosynthetic-SUE as their ratio. Dry mass of plants was noted after drying samples in a hot air oven at 80 °C till constant weight was obtained. The measurement of leaf area was done using a leaf area meter (LA211, Systronics, New Delhi, India).
Determination of Content of Cadmium
Leaf samples were dried in an oven at 80 °C for 48 h and the dried tissue was ground to a fine powder and digested with concentrated HNO3/HClO4 (3:1, v/v). The concentration of Cd was determined by the Atomic Absorption Spectrophotometer (GBC, 932 plus; GBC Scientific Instruments, Braeside, Australia).
Determination of H2O2 Content and Lipid Peroxidation
The H2O2 content was determined by initiating the reaction on addition of peroxidase at 25 °C, and the level of lipid peroxidation in leaves was determined by estimating the content of thiobarbituric acid reactive substances (TBARS). The details of the procedures and method adopted are given in Jahan et al. (2020).
Assay of Activity of Antioxidant Enzymes
Fresh leaf tissues (200 mg) were homogenized with an extraction buffer containing 0.05% (v/v) Triton X-100 and 1% (w/v) polyvinylpyrrolidone (PVP) in potassium phosphate buffer (100 mM of pH 7.0) using chilled mortar and pestle. The homogenized material was centrifuged at 15,000×g for 20 min at 4 °C. The supernatant obtained after centrifugation was used for the enzymes assay. For APX activity, extraction buffer was supplemented with 2 mM ascorbate. Activity of superoxide dismutase (SOD; EC, 1.15.1.1), ascorbate peroxidase (APX; EC, 1.11.1.11), and glutathione reductase (GR; EC, 1.6.4.2) was determined by using the modified method described earlier by Jahan et al. (2020).
Activity of ATP-Sulfurylase and SAT, Content of Sulfur, Cysteine and GSH
The activity of ATP-sulfurylase (ATP-S; EC, 2.7.7.4) and serine acetyltransferase (SAT; EC, 2.3.1.30) and content of sulfur, cysteine, and GSH were determined by using the modified methods as described earlier (Khan et al. 2015; Jahan et al. 2020).
Determination of NO Level
NO generation was determined by adopting the method given in Jahan et al. (2020).
Determination of N Content and NR Activity
N content in leaves was estimated by Kjeldahl digestion method and NR activity was estimated by monitoring nitrite content spectrophotometrically. The detailed procedures are given earlier in Jahan et al. (2020).
Statistical Analysis
Analysis of variance was performed on the data expressed as mean ± SE (n = 3) with the help of SPSS 17.0 for Windows. For significant data, least significant difference (LSD) was calculated at p < 0.05. Bars with the same letter were not significantly different by LSD test at p < 0.05.
Results
Screening of Cultivars for Cd Tolerance
Mungbean cultivars, Pusa Vishal, PusaRatna, Pusa 9531, Ganga, and MH 318, tested for cadmium sensitivity showed remarkable gradient in their Cd tolerance. Plants grown with 200 mg Cd kg−1 soil exhibited decrease in photosynthetic sulfur use efficiency (p-SUE) by 42.0%, 47.8%, 35.5%, 57.3%, and 77.1% and plant dry mass by 30.0%, 38.2%, 38.4%, 32.7%, and 28.2%, respectively, as compared to their respective controls. Pusa 9531 showed minimum decrease in p-SUE and plant dry mass under Cd stress and ,therefore, withstood maximum tolerance, while MH 318 showed minimum Cd tolerance due to maximum decrease in p-SUE and plant dry mass (Table 1).
Effect of GA and S Levels in Presence or Absence of Cd
Plants treated with individual or as combined dose of GA and 100S or 200S improved leaf area, plant dry mass, net photosynthesis, p-SUE, GSH content, and dry mass in Cd-exposed plants. The combined treatment of GA and 100S maximally enhanced leaf area, plant dry mass, net photosynthesis, p-SUE, GSH content, and plant dry mass in Cd-exposed plants by 21.0%, 38.4%, 36.3%, 32.9%, 113.9%, and 72.7%, compared to the control plants, while 200S in combination with GA showed no additional significant improvement in increasing these parameters. Hence, 100S was regarded as a sufficient dose for mitigation of Cd stress and further increase in S dose was found of no additional benefit (Table 2).
Influence of NO Modulators and GA + S on Photosynthetic Characteristics and Growth Under Cd Stress
The involvement of NO was studied using NO modulators in GA plus 100S-mediated alleviation of Cd stress and protection of photosynthesis and growth. Cd stress decreased chlorophyll content by 64.9%, net photosynthesis by 42%, intercellular CO2 concentration by 34.4%, stomatal conductance by 29%, Rubisco activity by 39%, leaf area by 30.2%, and plant dry mass by 38.5%, respectively, compared to the control. The application of SNP resulted in increased photosynthetic and growth parameters of plant in comparison the control, but c-PTIO inhibited these characteristics. SNP in absence of S could not alleviate Cd stress and the values for these observed characteristics were found lesser than the respective control. However, when 100S was supplemented with SNP, it promoted photosynthesis and growth under Cd stress. The maximal effects of GA plus 100S and Cd in increasing chlorophyll content, net photosynthesis, stomatal conductance, intercellular CO2 concentration, Rubisco activity, leaf area, and plant dry mass by 16.8%, 23.1%, 15.9%, 24.3%, 36.4%, 25.04% , and 38.5% in comparison to the respective control were ineffective when c-PTIO was used along with GA plus 100S and Cd, and the values of these characteristics were at par with the control (Table 3).
Influence of NO Modulators and GA + S on Cd Content, Oxidative Stress, and Antioxidant Enzymes
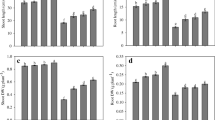
Leaf Cd content was analyzed to quantify the effect of NO modulators and GA plus 100S-mediated Cd stress alleviation. Leaf Cd content increased 33.9 fold in response to Cd exposure as compared to the control plants. Leaf Cd content was lesser in NO modulator-treated plants compared to the Cd grown plants. The application of SNP along with 100S to Cd grown plants decreased leaf Cd content by 96.4%, compared to the Cd treatment, whereas GA plus 100S in presence of Cd could decrease Cd content in leaf by 95.9% compared to the Cd treatment. The use of c-PTIO reversed the effects of GA plus 100S in presence of Cd on leaf Cd content which was 97%, compared to the Cd-treated plant (Fig. 1). The content of H2O2 and TBARS increased by 117.1% and 124.1% in Cd grown plants compared to the control (Fig. 1). The effect of NO modulators was in line with their effects on Cd content (Fig. 1).

Leaf Cd content (a), H2O2 content (b), and TBARS content (c) in mungbean (Vigna radiata L.) cultivar Pusa 9531 at 30 DAS. Plants were grown individually or in combination with 0, 10 µM GA, 100 µM NO (as SNP; sodium nitropruside), 100 mg S kg−1 soil (S), and 100 µM c-PTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide) grown with/without 200 mg Cd kg−1 soil. Data are presented as treatments mean ± SE (n = 3). Bars with the same letter were not significantly different by LSD test at p < 0.05
The activity of antioxidant enzymes was modulated with the use NO modulator and GA plus 100S in presence of Cd. The activity of SOD, APX, and GR increased in Cd grown plants. Application of SNP increased activity of SOD, APX, and GR by 198.5%, 171.4%, and 176.2%, in comparison to the control. The individual use of c-PTIO decreased activity of antioxidant enzymes in comparison to the control. Supplementation of SNP increased the activity of these antioxidant enzymes of Cd grown plants by 250.8%, 180.9%, and 247.6% in comparison to the control. However, the effect of SNP in Cd grown plants was more profound when 100S was supplemented. Supplementation of c-PTIO to GA plus 100S in presence of Cd reduced the effects GA plus 100S under Cd stress. This reflected that NO-induced activity of antioxidant enzymes was one possible mechanism of GA plus 100S-mediated increase in activity of antioxidant enzymes to counter Cd toxicity (Fig. 2).

Activity of superoxide dismutase (SOD) (a), ascorbate peroxidase (APX) (b), and glutathione reductase (GR) (c) in mungbean (Vigna radiata L.) cultivar Pusa 9531at 30 DAS. Plants were grown individually or in combination with 0, 10 µM GA, 100 µM NO (as SNP; sodium nitropruside), 100 mg S kg−1 soil (S), and 100 µM c-PTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide) grown with/without 200 mg Cd kg−1 soil. A unit of SOD was defined as the amount of enzyme that inhibited the NBT reduction by 50% at 560 nm. One unit of the APX was defined as the amount necessary to decompose 1 µmol of substrate per minute at 25 °C. One unit of GR was the amount necessary to decompose 1 µmol of NADPH min−1 at 25 °C. Bars with the same letter were not significantly different by LSD test at p < 0.05
Effect of NO Modulators and GA + S in S-Assimilation Under Cd Stress
Cd stress significantly decreased the S content by 24.3% and increased the content of Cys, GSH, and activity of ATP-S and SAT by 60.6%, 26.0%, 50.3%, and 51.4%, respectively, as compared to the control plants (Table 4). Application of SNP improved S-assimilation, and in absence of NO in c-PTIO-treated plants the S-assimilation reduced as observed by the determined parameters. The SNP or GA along with 100S in Cd grown plants was equally effective in increasing S content, ATP-S activity, SAT activity, Cys content, and GSH content by 58.1%, 165.5%, 105.3%, 160.1%, and 145.0%, respectively, in comparison to the control. The Cd grown plants receiving GA plus 100S treatment when supplemented with c-PTIO reversed the beneficial effects of GA plus 100S effects (Table 4).
Influence of NO Modulators in GA + S-Mediated Effect on N Content, NR Activity, and NO Generation
Plants grown with Cd showed reduced N content by 52.3% and NR activity by 35.9%, but increased NO generation by 371.4%, compared to the control plants. The influence of SNP optimized N content, NR activity, and NO generation, and the increase recorded was 19.3%, 49.2%, and 171.4% in comparison to the control. Application of c-PTIO showed involvement of NO in N-assimilation and NO generation. Application of SNP or GA supplemented with 100S to plants under Cd stress restored N-assimilation and optimized NO generation compared to Cd grown plants. The effects of GA + 100S in the presence of Cd were mediated by NO, which became evident with the use of c-PTIO to GA + 100S + Cd combination (Fig. 3).

N content (a), nitrate reductase activity (b), and NO generation (c) in mungbean (Vigna radiata L.) cultivar Pusa 9531 at 30 DAS. Plants were grown individually or in combination with 0, 10 µM GA, 100 µM NO (as SNP; sodium nitropruside), 100 mg S kg−1 soil (S), and 100 µM c-PTIO (2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide) with/without 200 mg Cd kg−1 soil. Data are presented as treatments mean ± SE (n = 3). Bars with the same letter were not significantly different by LSD test at p < 0.05
Discussion
Anthropogenic activities and unorganized industrial and household waste management has led to a serious threat of alarming increase in Cd concentration in agricultural land worldwide (Asgher et al. 2014; Emamverdian et al. 2015). In the presence of Cd, plants show initial alterations on biochemical and molecular mechanisms that are manifested in altered physiological processes and growth retardation. In the present study, Cd markedly inhibited growth, photosynthesis, and S status of mungbean plants more conspicuously in a sensitive cultivar. The tolerance of Pusa 9531 cultivar to Cd was attributed to its ability to utilize maximum S available and representing highest photosynthetic-SUE under Cd stress. It has been shown that S-assimilation results in synthesis of reduced compounds that help in reduction of oxidative stress through increase in antioxidant metabolism (Anjum et al. 2016; Khan et al. 2016). The cultivar Pusa 9531 experienced least oxidative stress with strong antioxidant metabolism because of higher S-assimilation and its incorporation into photosynthetic machinery and maintained growth. The supplementation of 100S proved better than 200S in mitigation of Cd toxicity, as 100S proved optimal in increasing the growth and photosynthesis in presence or absence of Cd. Further increasing the S level was of no additional benefit. In the earlier reported research maximum SUE was found with optimal S in mustard plants grown without stress (Khan et al. 2005) or in plants under Cd stress (Masood et al. 2012). Phytohormones have been shown to induce the mechanisms responsible for optimizing growth and photosynthesis under heavy metals stress conditions (Asgher et al. 2014, 2016). GA has been known to improvise photosynthetic and growth (Khan et al. 1996, 1997) through efficient utilization of N, P and S (Khan et al. 1997; 2005), and promoting SUE (Khan et al. 1998; Masood et al. 2012). It has been shown GA application improved growth characteristics of Cd affected plants by reducing its accumulation in cells by depressing the Cd transporter gene IRT1 (Zhu et al. 2012). The improved nutrients acquisition on GA application under Cd stress helped in metabolites synthesis for meeting of increased demand of above-ground parts. The supplementation of S to GA sprayed plants showed improved response towards use efficiency of nutrients and plants acquire S and N more efficiently (Khan et al. 2005) and augmented the positive effect of GA on the growth restoration and photosynthetic reversal under Cd stress. GA has been shown to increase photosynthetic enzymes (Khan et al. 1996; Yuan and Xu 2001), and promotes production of S-containing thiols such as GSH, cysteine, and methionine (Masood et al. 2012). The optimal availability of nutrients promoted by GA regulated N and S metabolism for better incorporated into various important amino acids and proteins including Rubisco, as the role of nutrients in alleviation of Cd toxicity has been shown (Nazar et al. 2012). The role of GA in alleviating the Cd toxicity has been shown in earlier studies (Meng et al. 2009; Masood and Khan 2013; Masood et al. 2012, 2016). Plants when experience Cd stress show increased oxidative stress and ROS generation that disrupts membrane integrity and leakage of essential ions occurs and results in nutritional deficiency. In such circumstances supplementation of S together with GA proved advantageous for plants by synthesizing more of the S-containing reduced metabolites. The production of GSH is controlled by its precursor cysteine regulated by N and S metabolism.
The potential of GA plus S in alleviating Cd stress through increased SUE was NO mediated. The supplementation of NO modulators in the present study has shown that SNP (NO promoter) improves growth and photosynthesis through increased mechanisms of N- and S-assimilation and antioxidant system, while c-PTIO (NO scavenger) inhibited the promotory effects of GA plus S on the these mechanisms and reversed the effects of GA + S on growth and photosynthesis. Studies have shown role of NO in plant development, physiological functions, nutrients acquisitions, antioxidant metabolism ,and other mechanisms under optimal and stressful environments (Du et al. 2008; Basalah et al. 2013; Mostofa et al. 2015; Fatma et al. 2016; Per et al. 2017; Ahmad et al. 2018; Kaya et al. 2019; Sehar et al. 2019; Jahan et al. 2020). NO mediates the functional capacity of GSH to minimize Cd-generated ROS (Asgher et al. 2014; Fatma et al. 2016; Zhao et al. 2015; Hasanuzzaman et al. 2017). NO has been shown to safeguard chlorophyll pigment maintaining the ultra-structure of chloroplast and enhancing net photosynthesis and gas exchange through stomatal aperture (Fatma and Khan 2014). The interaction of NO with other phytohormones under abiotic stress has been shown (Asgher et al. 2014, 2016). Masood and Khan (2013) and Masood et al. (2016) have reported that ethylene was involved in GA-induced S-assimilation and Cd tolerance. The inter-relation of NO with other signaling molecules, such as abscisic acid (Garcia-Mata and Lamattina 2002; Zhou et al. 2005; Wang et al. 2015), glucose (Sehar et al. 2019), and H2S (Kaya et al. 2019), to provide abiotic stress factors tolerance has been reported. However, involvement of NO in GA and S-mediated mechanisms for tolerance to Cd stress has not been shown earlier.
Conclusion
It may be concluded from the present study that Cd tolerance in cultivar Pusa 9531 was associated with higher use efficiency of S enabling its incorporation in photosynthesis and growth. Plants receiving GA and 100 mg S kg−1 soil show more pronounced effect in mitigation of Cd through increased photosynthetic-SUE and GSH production. The involvement of NO in GA and S-mediated alleviation of Cd toxicity was due to induction of N- and S-assimilation and antioxidant system, thus protecting photosynthesis and growth. There exists interplay between GA and NO in modulating Cd tolerance in presence of S through improvement in mechanisms for resisting Cd adverse effects. Thus, these interactions may be exploited as a sustainable strategy in strengthening the photosynthetic performance and growth of crops affected by heavy metal stress.
References
Ahmad P, Ahanger MA, Alyemeni MN, Wijaya L, Alam P (2018) Exogenous application of nitric oxide modulates osmolyte metabolism, antioxidants, enzymes of ascorbate-glutathione cycle and promotes growth under cadmium stress in tomato. Protoplasma 255:79–93
Ali MA, Asghar HN, Khan MY, Saleem M, Naveed M, Niazi NK (2015) Alleviation of nickel-induced stress in mungbean through application of gibberellic acid. Int J Agric Biol. https://doi.org/10.17957/IJAB/15.0001
Anjum NA, Umar S, Ahmad A, Iqbal M, Khan NA (2008) Sulphur protects mustard (Brassica campestris L.) from cadmium toxicity by improving leaf ascorbate and glutathione. J Plant Growth Regul 54:271–279
Anjum NA, Sharma P, Khan NA (2016) Catalase and ascorbate peroxidase-representative H2O2-detoxifying heme enzymes in plants. Environ Sci Poll Res 23:19002–190029
Asgher M, Khan MIR, Anjum NA, Khan NA (2014) Minimizing toxicity of cadmium in plants—role of plant growth regulators. Protoplasma 252:399–413
Asgher M, Per TS, Masood A, Fatma M, Freschi L, Corpas FJ, Khan NA (2016) Nitric oxide signaling and its crosstalk with other plant growth regulators in plant responses to abiotic stress. Environ Sci Pollut Res 24:2273–2285
Basalah MO, Ali HM, Al-Whaibi MH, Siddiqui MH, Sakran AM, Al Sahli AA (2013) Nitric oxide and salicylic acid mitigate cadmium stress in wheat seedlings. J Pure Appl Microbiol 7:139–148
Bolan N, Kunhikrishnan A, Thangarajan R et al (2000) Remediation of heavy metal(loid)s contaminated soil-to mobilize or to immobilize. J Haz Mat 266:141–166
Choppala G, Saifullah Bolan N, Bibi S, Iqbal M, Rengel Z, Ok YS (2014) Cellular mechanisms in higher plants governing tolerance to cadmium toxicity. Crit Rev Plant Sci 33:374–391
Dalcarso G, Farinati S, Furini A (2010) Regulatory networks of cadmium stress in plants. Plant Sig Behav 5:663–667
Du S, Zhang Y, Lin X, Wang Y, Tang C (2008) Regulation of nitrate reductase by nitric oxide in Chinese cabbage pakchoi (Brassica chinensis L.). Plant Cell Environ 31:195–204
Emamverdian A, Ding Y, Mokhberdoran F, Xie Y (2015) Heavy metal stress and some mechanisms of plant defense response. Sci World J. https://doi.org/10.1155/2015/756120
Fatma M, Khan NA (2014) Nitric oxide protects photosynthetic capacity inhibition by salinity in Indian mustard. J Funct Environ Bot 4:106–116
Fatma M, Masood A, Per TS, Rasheed F, Khan NA (2016) Interplay between nitric oxide and sulfur assimilation in salt tolerance in plants. Crop J 4:153–161
Gangwar S, Singh VP, Prasad SM, Maurya JN (2010) Modulation of manganese toxicity in Pisum sativum L. seedlings by kinetin. Sci Hortic 126:467–474
García-Mata C, Lamattina L (2002) Nitric oxide and abscisic acid crosstalk in guard cells. Plant Physiol 128:790–792
Gill SS, Khan NA, Tuteja N (2012) Cadmium at high dose perturbs growth, photosynthesis and nitrogen metabolism while at low dose it up regulates sulfur assimilation and -antioxidant machinery in garden cress (Lepidium sativum L.). Plant Sci 182:112–120
Gill SS, Hasanuzzaman M, Nahar K, Tuteja N (2013) Importance of nitric oxide in cadmium stress tolerance in crop plants. Plant Physiol Biochem 63:254–261
Harada E, Yamaguchi Y, Koizumi N, Sano H (2002) Cadmium stress induces production of thiol compounds and transcripts for enzymes involved in sulfur assimilation pathways in Arabidopsis. J Plant Physiol 159:445–448
Hasanuzzaman M, Nahar K, Hossain MS, Anee TI, Parvin K, Fujita M (2017) Nitric oxide pre treatment enhances antioxidant defense and glyoxalase systems to confer PEG-induced oxidative stress in rape seed. J Plant Interact 12:323–331
Hossain MA, Piyatida P, Teixera da Silva JA, Fujita M (2012) Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J Bot Article 16:1169–1192
Hsu YT, Kao CH (2004) Cadmium toxicity is reduced by nitric oxide in rice leaves. Plant Growth Regul 42:227–238
Iqbal N, Nazar R, Khan MIR, Masood A, Khan NA (2011) Role of gibberellins in regulation of source–sink relations under optimal and limiting environmental conditions. Curr Sci 100:998
Jahan B, Rehman MT, Alajmi MF, Khan NA (2020) Treatment of nitric oxide supplemented with nitrogen and sulfur regulates photosynthetic performance and stomatal behavior in mustard under salt stress. Physiol Plant 168:490–510
KavamuraVN EE (2010) Biotechnological strategies applied to the decontamination of soils polluted with heavy metal. Biotechnol Adv 28:6–69
Kaya C, Ashraf M, Nasser Alyemeni MN, Ahmad P (2019) Responses of nitric oxide and hydrogen sulfide in regulating oxidative defence system in wheat plants grown under cadmium stress. Physiol Plant 168:345–360
Khan NA (1996) Effect of gibberellic acid on carbonic anhydrase, photosynthesis, growth and yield of mustard. Biol Plant 38:1–147
Khan NA, Ansari HR, Mobin M (1996) Effect of gibberellic acid and nitrogen on carbonic anhydrase activity and mustard biomass. Biol Plant 38:601–603
Khan NA, Ansari HR, Samiullah HR (1997) Effect of gibberellic acid spray and basal nitrogen and phosphorus on productivity and fatty acid composition of rapeseed mustard. J Agron Crop Sci 179:29–33
Khan NA, Ansari HR, Samiullah HR (1998) Effect of gibberellic acid spray during ontogeny of mustard on growth, nutrient uptake and yield characteristics. J Agr Crop Sci 181:61–63s
Khan NA, Mir R, Khan M, Javid S, Samiullah HR (2002) Effects of gibberellic acid sprays on nitrogen yield efficiency of mustard grown with different nitrogen levels. Plant Growth Regul 38:243–247s
Khan NA, Mobin M, Samiullah HR (2005) The influence of gibberellic acid and sulfur fertilization rate on growth and S-use efficiency of mustard (Brassica juncea). Plant Soil 270:269–274
Khan NA, Nazar R, Anjum NA (2009) Growth, photosynthesis and antioxidant metabolism in mustard (Brassica juncea L.) cultivars differing in ATP-sulfurylase activity under salinity stress. Sci Hort 122:455–460
Khan MIR, Nazir F, Asgher M, Per TS, Khan NA (2015) Selenium and sulfur influence ethylene formation and alleviate cadmium-induced oxidative stress by improving proline and glutathione production in wheat. J Plant Physiol 173:9–18
Khan MIR, Iqbal N, Masood A, Mobin M, Anjum NA, Khan NA (2016) Modulation and significance of nitrogen and sulfur metabolism in cadmium challenged plants. Plant Growth Regul 78:1–11
Kopriva S, Suter M, von Ballmoos P, Hesse H, Krähenbühl U, Rennenberg H, Brunold C (2002) Interaction of sulfate assimilation with carbon and nitrogen metabolism in Lemna minor. Plant Physiol 130:1406–1413
Lancilli C, Giacomini B, Lucchini G, Davidian JC, Cocucci M, Sacchi GA, Nocito FF (2014) Cadmium exposure and sulfate limitation reveal differences in the transcriptional control of three sulfate transporter (Sultr1; 2) genes in Brassica juncea. BMC Plant Biol 14:132
Leitner M, Vandelle E, Gaupels F, Bellin D, Delledonne M (2009) NO signals in the haze: nitric oxide signalling in plant defence. Curr Opin Plant Biol 12:451–458
Lunde C, Zygadlo A, Simonsen HT, Nielsen PL, Blennow A, Haldrup A (2008) Sulfur starvation in rice: the effect on photosynthesis, carbohydrate metabolism, and oxidative stress protective pathways. Physiol Plant 134:508–521
Marschner H (1995) Mineral nutrition of higher plants. Academic Press, London
Masood A, Khan NA (2013) Ethylene and gibberellic acid interplay in regulation of photosynthetic capacity inhibition by cadmium. J Plant Biochem Physiol 1:111
Masood A, Iqbal N, Khan NA (2012) Role of ethylene in alleviation of cadmium-induced photosynthetic capacity inhibition by sulphur in mustard. Plant Cell Environ 35:524–533
Masood A, Khan MIR, Fatma M, Asgher M, Per TS, Khan NA (2016) Involvement of ethylene in gibberellic acid-induced sulfur assimilation, photosynthetic responses, and alleviation of cadmium stress in mustard. Plant Physiol Biochem 104:1–10
Meng HB, Hua SJ, Shamsi IH, Jilani G, Li YL, Jiang LX (2009) Cadmium-induced stress on the seed germination and seedling growth of Brassica napus L., and its alleviation through exogenous plant growth regulators. J Plant Growth Regul 58:47–59
Metwally A, Safronova VI, Belimov AA, Dietz KJ (2005) Genotypic variation of the response to cadmium toxicity in Pisum sativum L. J Exp Bot 56:167–178
Mobin M, Khan NA (2007) Photosynthetic activity, pigment composition and antioxidative response of two mustard (Brassica juncea) cultivars differing in photosynthetic capacity subjected to cadmium stress. J Plant Physiol 164:601–610
Mostofa MG, Fujita M, Tran LSP (2015) Nitric oxide mediates hydrogen peroxide-and salicylic acid induced salt tolerance in rice (Oryza sativa L.) seedlings. J Plant Growth Regul 77:265–277
Nazar R, Iqbal N, Masood A, Khan MIR (2012) Cadmium toxicity in plants and role of mineral nutrients in its alleviation. Am J Plant Sci 3:1476–1489
Nocito FF, Pirovano L, Cocucci M, Sacchi GA (2002) Cadmium-induced sulfate uptake in maize roots. Plant Physiol 129:1872–1879
Noriega GO, Balestrasses KB, Batlle A, Tomaro ML (2007) Cadmium induced oxidative stress in soybean plants also by the accumulation of delta-aminolevulinic acid. Biometals 20:841–851
Per TS, Masood A, Khan NA (2017) Nitric oxide improves S-assimilation and GSH production to prevent inhibitory effects of cadmium stress on photosynthesis in mustard (Brassica juncea L.). Nitric Oxide 68:111–124
Picazo I, Ros R, Moya JL (2007) Heavy metal hormone interactions in rice plants: effect on growth, net photosynthesis and carbohydrate distribution. J Plant Growth Regul 14:61–67
Rubio MI, Escrig I, Martinez-Cortina C, Lopez-Benet FJ, Snaz A (1994) Cadmium and nickle accumulation in rice plants. Effects on mineral nutrition and possible interactions of abscisic and gibberellic acids. Plant Growth Regul 14:151–157
Sehar Z, Masood A, Khan NA (2019) Nitric oxide reverses glucose-mediated photosynthetic repression in wheat (Triticum aestivum L.) under salt stress. Environ Exp Bot 161:277–289
Singh HP, Batish DR, Kaur G, Arora K, Kohli RK (2008) Nitric oxide (as sodium nitroprusside) supplementation ameliorates Cd toxicity in hydroponically grown wheat roots. Environ Exp Bot 63:158–167
Singh S, Parihar P, Singh R, Singh VP, Prasad SM (2016) Heavy metal tolerance in plants: role of transcriptomics, proteomics, metabolomics, and ionomics. Front Plant Sci 6:1143
Sun P, Tian QY, Zhao MG, Dai XY, Huang JH, Li LH, Zhang WH (2007) Aluminum-induced ethylene production is associated with inhibition of root elongation in Lotus japonicus L. Plant Cell Physiol 48:1229–1235
Tandon HLS (1995) Sulphur fertilizers for Indian agriculture. A guide book. New Delhi, Fertilizer Development and Consultation Organisation
Wagner GJ (1993) Accumulation of cadmium in crop plants and its consequences to human health. Adv Agron 51:173–212
Wahid A, Khaliq S (2015) Architectural and biochemical changes in embryonic tissues of maize under cadmium toxicity. Plant Biol 17:1005–1012
Wang P, Du Y, Hou YJ, Zhao Y, Hsu CC, Yuan F, Zhu X, Tao WA, Song CP, Zhu JK (2015) Nitric oxide negatively regulates abscisic acid signaling in guard cells by S-nitrosylation of OST1. Proc Natl Acad Sci USA 112:613–618
Wani SH, Kumar V, Shriram V, Sah SK (2016) Phytohormones and their metabolic engineering for abiotic stress tolerance in crop plants. Crop J 4:162–176
Xiong J, An L, Lu H, Zhu C (2009) Exogenous nitric oxide enhances cadmium tolerance of rice by increasing pectin and hemicellulose contents in root cell wall. Planta 230:755–765
Yuan L, Xu DQ (2001) Stimulation effect of gibberellic acid short-term treatment on leaf photosynthesis related to the increase in Rubisco content in broad bean and soybean. Photo Res 68:39–47
Zhao H, Jin Q, Wang Y, Chu L, Li X, Xu Y (2015) Effects of nitric oxide on alleviating cadmium stress in Typha angustifolia. J Plant Growth Regul 78:243–251
Zhou B, Guo Z, Xing J, Huang B (2005) Nitric oxide is involved in abscisic acid induced antioxidant activities in Stylosanthes guianensis. J Exp Bot 56:3223–3228
Zhu XF, Jiang T, Wang ZW, Lei GJ, Shi YZ, Li GX, Zheng SJ (2012) Gibberellic acid alleviates cadmium toxicity by reducing nitric oxide accumulation and expression of IRT1 in Arabidopsis thaliana. J Hazard Mater 239:302–307
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
There is no conflict of interest.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Hasan, S., Sehar, Z. & Khan, N.A. Gibberellic Acid and Sulfur-Mediated Reversal of Cadmium-Inhibited Photosynthetic Performance in Mungbean (Vigna radiata L.) Involves Nitric Oxide. J Plant Growth Regul 39, 1605–1615 (2020). https://doi.org/10.1007/s00344-020-10175-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00344-020-10175-4