
Abstract
Chromium (Cr) contamination of agricultural soils is a major threat to human and plant health worldwide and causes reductions in plant growth and crop yields. 24-epibrassinolide (EBL) and nitric oxide (NO) have been shown to ameliorate the reductions in growth caused by the stresses induced by heavy metals; however, the interactions between EBL and NO on the alleviation of Cr-induced phytotoxicity have been poorly studied. Hence, this study was undertaken to examine any beneficial effects of EBL (0.01 µM) and NO (100 µM), applied alone or in combination, on the mitigation of stress induced by Cr (100 µM) in soybean seedlings. Although EBL and NO applied alone reduced the toxic effects of Cr, the combined treatment had the greatest effect. Mitigation of Cr intoxication occurred via reduced Cr uptake and translocation and by ameliorating reductions in water contents, light-harvesting pigments, and other photosynthetic parameters. In addition, the two hormones increased the activity of enzymatic and non-enzymatic defense mechanisms increasing the scavenging of reactive oxygen species, thereby reducing membrane damage and electrolyte leakage. Furthermore, the hormones reduced the accumulation of the toxic compound, methylglyoxal, by amplifying activities of glyoxalase I and glyoxalase II. Thus, applications of NO and EBL can significantly mitigate Cr-phytotoxicity when cultivating soybean plants in Cr-contaminated soils. However, further more-in depth studies including field investigations parallel with calculations of cost to profit ratios and yield losses are requested to validate the effectiveness of NO and/or EBL for remediation agents in Cr-contaminated soils with using key biomarkers (i.e., oxidative stress, antioxidant defense, and osmoprotectants) involved in the uptake, accumulation, and attenuation of Cr toxicity tested in our study.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Heavy metals (HM) cause severe damage to crops (Basit et al. 2022a, e; Kushwaha et al. 2019) and become a source of human intoxication if they enter the food chain (Singh et al. 2016). Chromium (Cr) is considered the most common toxic pollutant, and anthropogenic activities are the major reason for Cr contamination of agricultural lands (Basit et al. 2022b; Naz et al. 2021). The Cr-induced toxicity causes numerous changes in plant physiology, morphology, and biochemistry (Husain et al. 2022). Besides, Cr suppresses crop growth, reduces photosynthetic attributes, and disrupts ionic homeostasis by inducing cellular oxidative stress because of the higher production of reactive oxygen species (ROS), compounds that lead to the degradation of macromolecules and membranes (Basit et al. 2022c; Hussain et al. 2022; Prakash et al. 2022). In addition, Cr stimulates the accumulation of a toxic compound, methylglyoxal (MG), which also accumulates due to other environmental stresses (Jin et al. 2015; Kharbech et al. 2020a, b). The increased generation and accumulation of ROS and MG cause lipid peroxidation and membrane damage and the production of cytotoxic products such as malondialdehyde (MDA) and 4-hydroxy-2-nonenal (4-HNE). In addition, both MG and ROS perturb membrane-bound enzymes like H+-ATPase causing electrolyte leakage and disturbing the osmotic balance and the production of soluble sugars and proteins (Wang et al. 2019; Zhang et al. 2017). To overcome the negative effects of Cr, plants activate various defense mechanisms, including elimination of Cr by binding it to cell walls, by cytosolic chelation, and by vacuolar compartmentalization (Singh et al. 2013; Kharbech et al. 2017). Under a Cr-stressed environment, plants also increase their enzymatic and non-enzymatic defensive activities including superoxide dismutase (SOD), ascorbate peroxidase (APX), catalase (CAT), glutathione reductase (GR), glutathione (GSH) (Kharbech et al. 2017, 2020a) as well as signaling cascades associated with defense gene expression (Adhikari et al. 2018). Furthermore, plants also have a MG detoxification mechanism composed of glyoxalase I (GlyI) and glyoxalase II (GlyII) (Hossain et al. 2012). GSH is used by GlyI to convert MG into a thioester; following which, GlyII further hydrolyzes the thioester to regenerate GSH (Hossain et al. 2012; Kharbech et al. 2020a). These antioxidant defense mechanisms have the potential for the development of Cr-tolerance; however, the processes behind these antioxidant defense systems in relation to Cr stress need to be further explored.
Nitric oxide (NO) can minimize the negative consequences of various environmental stresses such those caused by salinity (Ahmad et al. 2016), iron deficiency (Shanmugam et al. 2015), drought (Cechin et al. 2015), and Cr (Basit et al. 2022a, e; Singh et al. 2022). Currently, the role of signaling molecules such as NO to ameliorate the negative impacts of HM stress by suppressing both ROS and MG accumulation and regulating antioxidant activities is under investigation. NO acts as a multifunctional signaling molecule inside plants that activates the defensive mechanism and other metabolic pathways under various environmental stressed conditions (Shivaraj et al. 2020). Additionally, NO can modulate the production of GSH which reduces MG over-production and accumulation (Mostofa et al. 2018).
Brassinosteroids (BRs) are a group of naturally occurring phytohormones involved in plant development and numerous physiological processes (Basit et al. 2021b; Yokota and Mori 2018). They are involved in the activation of transcriptional responses that are responsible for cell division (Hu et al. 2000) as well as cell and hypocotyl elongation (Szekeres et al. 1996); thus, they enhance the plant growth. BRs alleviate HM toxicity in plants by encouraging growth, the accumulation of osmolytes, the uptake of mineral nutrients, and through antioxidative and glyoxalase defense systems (Jan et al. 2018) and have been shown to mitigate the effects of chromium (Basit et al. 2021a), cadmium (Santos et al. 2018), aluminum (Basit et al. 2022d), zinc (Ramakrishna and Rao 2015), and lead (Kohli et al. 2018). However, the effect of combined applications of NO and 24-epibrassinolide (EBL) to counteract Cr toxicity in soybeans at the physio-biochemical and metabolic levels has been little studied.
Soybean (Glycine max L. (Merr.)) is an important cash crop worldwide that provides oils and proteins that are essential for human and animal nutrition (Ning et al. 2019). Yields of this crop can be markedly reduced due to different HM stresses including Cr (Basit et al. 2022e). EBL has been shown to mitigate losses in biomass and growth in pepper plants subjected to water stress, a phenomenon that was reversed by the combined use of a NO scavenger (Kaya et al. 2019). Given this interaction between NO and EBL, this study aimed to understand the crosstalk and interplays between these compounds in the alleviation of Cr toxicity in soybean plants.
Materials and methods
Plant materials
The soybean cultivar used in this study was Huaidou-19 (HD-19), which was purchased from the Zhejiang Nongke Seeds CO., Ltd. Hangzhou, Zhejiang Province, China. 24-Epibrassinolide (EBL) and sodium nitroprusside (as nitric oxide (NO) donor) were purchased from Shanghai, Yuanye Biotechnology Co., Ltd., China.
Plant growth conditions and hydroponic culture
The sterilization of seeds was produced by dipping them into 5% (w/v) sodium hypochlorite solution for 20 min; after which, they were washed with double-distilled water (ddH2O). The seeds were then germinated in a growth chamber at 25 °C with an 8/16 h light/dark cycle (Zheng et al. 2006). Seeds producing a radicle after 4 days of germination were selected and transferred to 2-L black plastic containers filled with Hoagland nutrient solution. The nutrient solution was made of the following ingredients: 0.5-µM potassium nitrate (KNO3); 0.5-µM calcium nitrate (Ca(NO3)2); 0.5-µM magnesium sulfate MgSO4; 2.5-µM monopotassium phosphate (KH2PO4); 2.5-µM ammonium chloride (NH4Cl); 100-µM ferric EDTA (Fe–K–EDTA); 30-µM boric acid (H3BO3); 5-µM manganese monosulfate (MnSO4); 1-µM copper sulfate (CuSO4); 1-µM zinc sulfate (ZnSO4); and 1-µM ammonium heptamolybdate ((NH4)6Mo7O24) per 1000-mL ddH2O. The pH of the nutrient solution was adjusted to 5.7 ± 0.1 daily with hydrochloric acid (HCl) or sodium hydroxide (NaOH) and maintained throughout the experiment.
Treatments, sampling, and growth analysis
The experiment was carried out with complete random block with 4 replicates per treatment. After another 10 days’ growth in the Hoagland nutrient solution, seedlings of uniform growth were selected and divided into two groups: one group was treated with Cr for 7 days (Cr treatment, the final concentration of Cr in nutrient solution was 100 µM), and another group was without Cr. Meanwhile, four treatments containing EBL (0.01 µM), NO (100 µM), EBL + NO (0.01 µM + 100 µM), and double-distilled water (ddH2O) were, respectively, added to the nutrient solution for each group. Therefore, there were totally 8 treatments, which were H2O, EBL, NO, EBL + NO, H2O + Cr, EBL + Cr, NO + Cr, and EBL + NO + Cr. Sampling was carried out at the 7th day under Cr stress treatment, and then all seedlings were taken out and washed with ddH2O to remove the Cr deposits. The seedlings’ heights and fresh weights were assessed. To determine the dry masses, seedling shoots, and roots were separately oven dried at 80 °C for 24 h.
Determination of photosynthetic pigments, gas exchange parameters, and relative water contents
Fresh leaf samples (0.2 g) were placed in 3 mL of 95% (v/v) ethanol, the mixture homogenized then centrifuged at 5000 × g for 10 min for pigment extraction. Afterward, 9 mL of ethanol was added into 1-mL aliquots in test tubes and left for overnight. Chlorophyll contents were assessed at 645-, 663-, and 470-nm UV–VIS spectrophotometer (Ultrospecific 3000 Biochrom Ltd. Cambridge, England) (Lichtenthaler 1987). The measurement of total chlorophyll was done by using the formula:
The quantities of total chlorophyll content were presented as mg g−1 of plant extract.
Gas exchange parameters (net photosynthetic (Pn), CO2 concentration (Ci), stomatal conductance (gn), and photochemical efficacy of PS II (Fv/Fm)) were estimated with a LiCor-6400 portable photosynthesis system (Li-Cor Inc., Lincoln, NE, USA) by the methodology of Zhou and Leul (1999).
The relative water contents (RWC) of leaves were estimated according to the method of Mostofa and Fujita (2013) using the formula:
RWC were determined on the bases of fresh weight (FW), dry weight (DW), and turgid weight (TW).
Determination of Cr accumulation and micronutrients level
Aliquots (0.2 g) of ground roots and shoots were used for the estimation of Cr contents. The samples were heated on hot plate at 500 °C for 24 h, and then digested in HNO3–HClO4 (3:1, v/v) for 48 h at room temperature. Afterward, the digested samples were diluted with 15 mL of ddH2O. The final filtrate was used to quantify the Cr content measurements using an atomic absorption spectrometer (Aziz et al. 2015). Translocation factors (TFs) were calculated using the formula
Determination of total soluble sugar, protein, proline, and relative water contents
To determine total soluble sugars (TSS), fresh leaf samples (0.5 g) were ground using mortars and pestles along with extraction buffer [phosphate buffer (50 mM, pH 7), glycerol (10%, v/v), ascorbate (1 mM), potassium chloride (100 mM), and β-mercaptoethanol (5 mM)]. After, the homogenate was centrifuged for 15 min at 12,000 × g. The supernatant was used for the quantification of TSS using the phenol–sulfuric acid assay of Dubois et al. (1951). The supernatant was also used to analyze the total soluble protein (TSP) (Bradford 1976). To estimate the proline contents, 0.5 g of fresh samples was homogenized in sulfosalicylic acid (3%, w/v), then centrifuged at 10,000 × g for 15 min. Acid-ninhydrin, glacial acetic acid, and supernatant were mixed in 1:1:1 ratio, heated at 100 °C for 1 h and then cooled instantly on ice to avoid any further reaction. The colored-chromophore produced was extracted in toluene, and its absorbance at 520 nm was measured (Bates et al. 1973). Proline contents were determined using proline standards.
Estimation of electrolyte leakage (EL), malondialdehyde (MDA), and methylglyoxal contents
To estimate the electrolyte leakage (EL) (dSm−1), 1 g of seedlings was immersed in 25 mL of ddH2O and incubated at 25 °C for 24 h. The samples were transferred to another blank beaker, and dd H2O was added up to 25-mL volume; EL was quantified in dSm−1 (Ista et al. 2004). For the measurement of MDA,1.5 mL of tissue was homogenized in 2.5 mL of 5% 2-thiobarbituric acid (TBA) and diluted in 5% trichloroacetic acid at 100 °C for 20 min, cooled on ice and then centrifuged, and the absorbance at 532 and 600 nm was measured using UV–Vis spectrophotometer (Hitachi U-2910) (Heath and Packer 1968). Methylglyoxal (MG) contents were estimated according to the description of Yadav et al. (2005).
Determination of hydrogen peroxide (H2O2) and superoxide content (O2 .−)
The hydrogen peroxide (H2O2) was estimated by homogenizing the seedling in a phosphate buffer and centrifuged at 6000 × g. The 0.1% titanium sulfate containing 20% (v/v) H2SO4 was added to the supernatant and intermixed. The intensity of the yellow color was measured at 410 nm, and then the H2O2 contents were calculated according to the method of Kwasniewski et al. (2013). The O2.− production rate was monitored following the method described by Jiang and Zhang (2001). The 250 mL of 7.2 mM 1,2-diaminobenzene, 650 mL of sample extract, and 100 mL of 5-M perchloric acid were used to prepare the assay mixture of 1 mL. The absorbance of the mixture was monitored at 336 nm, and a standard curve of pure MG was used to deliberate the final concentration of MG.
Enzyme extraction and determination activities
The samples of uppermost expended leaves and roots were used for each antioxidant. To estimate the SOD activity, the absorbance of a reaction mixture containing 50-mM KH2PO4 buffer, 2.24-mM nitro-blue tetrazolium (NBT), and 2.36-mM xanthine was measured for 2 min according to the described protocol of El-Shabrawi et al. (2010). The required amount of enzyme was used to restrict NBT reduction (50%), and the unit of SOD activity was evaluated as min−1 mg−1 protein. Catalase (CAT) activity was determined by the methodology of Hossain et al. (2010) and used an extinction coefficient of 39.4 M−1 cm−1. APX activity was determined as the protocol of Nakano and Asada (1981). GR was assayed based on the method of Schaedle and Bassham (1977) with a slight modification; reduced glutathione (GSH) and oxidized glutathione (GSSG) were measured according to the protocol of Law et al. (1983).
Gly I (EC 4.4.1.5) and Gly II (EC 3.1.2.6) were determined according to Mostofa et al. (2014). MG was used as a substrate to determine the GlyI in a reaction mixture that comprised magnesium sulfate (15 mM), potassium phosphate buffer (100 mM, pH 7.0), GSH (1.7 mM). The absorbance at 240 nm was measured and an extinction coefficient of 3.37 mM−1 cm−1. The activity of GlyII was measured by analyzing GSH at 412 nm and an extinction coefficient of 13.6 mM−1 cm−1. Furthermore, final volume of 1 mL was prepared by using DTNB (0.2 mM), S-D-lactoylglutathione (1 mM), and Tris–HCl buffer (100 mM, pH 7.2).
Statistical analysis
All experimental data were analyzed by applying analysis of variance with the least significant differences (LSD) determined at p < 0.05 level between mean values using Statistix (8.1) software. All the experimental values were mean of four replicates ± standard deviation (SD).
Results
Effect of exogenous supplementation of EBL, NO, and EBL + NO on the plant growth and biomass under Cr stress
The plants treated with H2O, EBL, and NO without Cr exposure were considered as the control group. In this group, EBL and NO alone had a slightly greater effect on plant growth than H2O (Fig. S1). However, compared with the control group of soybean cultivar, plant length under Cr stress was significantly inhibited, leaf size was relatively small, and leaf color was more yellow (Fig. S1). Compared with H2O treatment (H2O + Cr), single application of EBL and NO could significantly improve plant height and reduce the poisoning symptoms of plants under Cr stress. Whereas, the combined supplementation of EBL + NO further increased the plant height with greenish leaf texture in comparison to their sole application under Cr-induced toxicity (Fig. S1).
Relative to the control, the changing at the microphysical level was recorded inside soybean plants under Cr exposure such as shoot length (SL), root length (RL), fresh weight (FW), and dry weight (DW) of the plant by 52.4%, 40.7%, 49.3%, and 61.6%, respectively, while EBL and NO alone enhanced the SL, RL, FW, as well as DW by 31.1/33.62%, 24.76/27.98%, 35.72/39.14%, 37.65/35.72%, respectively, as compared to H2O + Cr. Additionally, EBL + NO treatment further improved the above-mentioned indexes by an average of 26.43%, 29.72%, 27.65%, and 24.39% individually than EBL + Cr and NO + Cr (Fig. 1). In the control group without Cr, EBL + NO improved the SL, RL, DW, and FW of plants obviously in comparison to the alone treatment of EBL and NO (Fig. 1). However, the above indexes were slightly increased in the plants treated with EBL and NO alone compared with those treated with H2O. In general, the results showed that both EBL and NO could alleviate the Cr damage on plant growth attributes, but under Cr stress, EBL + NO promoted plant growth and biomass more significantly than EBL and NO alone.

Effect of nitric oxide (NO) and 24-epibrassinolide (EBL) on plant biomass of soybean seedlings exposed to chromium (Cr) (100 µM). a Shoot length, b root length, c dry weight, d fresh weight. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Effect of EBL, NO, and EBL + NO on the plant photosynthetic attributes under Cr stress
Under Cr stress, H2O treatment significantly inhibited the photosynthetic characteristics of plants compared with no Cr stress. Compared with H2O, EBL and NO alone increased the total chlorophyll content by 28.7% and 31.4%, carotenoids by 21.7% and 24.6%, Pn by 29.2% and 33.7%, Ci by 26.4% and 29.2%, gs by 34.4% and 37.7%, and Fv/m value by 39.6% and 44.1%, respectively. Moreover, EBL + NO could even remarkably provoke the level of photosynthetic pigments as compared to EBL and NO treatment under Cr stress alone (Fig. 2a–f). In the absence of Cr stress (control group), EBL + NO treatment significantly promoted the above photosynthetic indexes compared with H2O treatment, while EBL and NO alone did not significantly increase the chlorophyll content of soybean seedlings.

Interactive effect of nitric oxide (NO) and 24-epibrassinolide (EBL) on the photosynthetic attributes of soybeans exposed to chromium (Cr) (100 µM). a Total chlorophyll, b carotenoids, c net photosynthetic rate (Pn), d CO2 concentration (Ci), e stomatal conductance (gs), f Fv/m values. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Modulation of uptake and accumulation of endogenous Cr contents by EBL, NO, and EBL + NO application under Cr exposure
Relative to the control treatments, data exhibited the great uptake and accumulation of Cr in roots and its translocation toward shoots as compared to the control plants. Under Cr stress, the accumulation of Cr in soybean roots and shoots was significantly higher than that in Cr-untreated plants (Fig. 3a, b). However, compared with the H2O + Cr, the inhibition rates of EBL and NO alone on Cr accumulation in roots/shoots were 36.2/28.9% and 37.7/33.2%, respectively, and Cr uptake and transport were reduced by 70.3% and 57.4%, respectively, in EBL + NO + Cr treatment (Fig. 3c).

Effect of nitric oxide (NO) and 24-epibrassinolide (EBL) on Cr contents and translocation in soybean tissues under chromium (Cr) (100 µM) stress. a Cr contents in shoots, b Cr contents in roots, c translocation factor. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Effect of EBL, NO, and EBL + NO on the plant proline, relative water, total soluble sugar, and protein contents under Cr stress
In the control group without Cr, no significant alternations were perceived among the H2O, NO, and EBL-alone treatments, as well as EBL + NO, in the case of proline and TSS (Fig. 4a, d), although a minor stimulation was noticed inside EBL + NO-treated plants as compared to H2O, EBL- and NO-alone application in the values of RWC and TSP (Fig. 4b, c). Relative to the control group, the Cr-treated plants showed a remarkable increase in the values of proline (53.6%), although the supplementation of EBL and NO minimized the proline level by 32.7% and 36.2%, respectively, under Cr-induced toxicity. Moreover, the EBL + NO application further lowered the proline level by 25.8% in contrast to the EBL and NO alone under Cr exposure (Fig. 4a). The RWC, TSS, and TSP of H2O + Cr were significantly decreased by 58.1%, 51.7%, and 44.3% as compared to H2O treatment without Cr stress, respectively. However, compared with H2O + Cr, EBL + Cr, and NO + Cr improved the above three parameters by 28.8/30.7%, 23.2/26.8%, 36.9/38.1%, respectively, and EBL + NO + Cr significantly increased RWC (49.6%), TSS (45.3%), and TSP (57.3%).

Interactive effect of nitric oxide (NO) and 24-epibrassinolide (EBL) on a proline, b relative water contents, c total soluble sugars, and d total soluble proteins under chromium (Cr) (100 µM) stress. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Effect of EBL, NO, and EBL + NO on ROS generation, MDA level, electrolyte leakage, and methylglyoxal contents under Cr stress
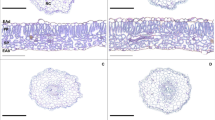
Relative to the plants in control group, data demonstrated that Cr-contaminated condition prominently accumulated the MDA (60.9%), EL (64.8%), H2O2 (78.4%), O2.− (69.3%), and MG (71.2%) contents in soybean plants (Fig. 5a–e). The individual applications of EBL and NO inside Cr-contaminated media noticeably lowered the accumulation of MDA (40.9/41.7%), EL (32.3/34.6%), H2O2 (38.2/41.4%), O2.− (45.4/48.2%), and MG (50.1/52.6%) contents, though EBL + NO + Cr more notably diminishes the upsurge of MDA by 23.2%, EL by 27.9%, H2O2 by 30.1%, O2• ‒ by 26.5%, and MG by 33.5% than t EBL + Cr and NO + Cr (Fig. 5). To further validate the accumulation of H2O2 and O2•‒, the 3, 3-diaminobenzidine (DAB) and nitro blue tetrazolium (NBT) staining was used with EBL, NO, and EBL + NO under Cr disclosure (Fig. 5a, b). The leaves of soybean plants were exposed to 100-µM Cr toxicity, and soybean control plants signified dark brown and dark blue staining for H2O2 and O2•‒, exclusively. Particularly, EBL and NO-alone applications avert the staining intensity of DAB and NBT inside soybean leaves, but a more prominent diminishing effect was noted in EBL + NO as compared to the H2O treatment under Cr-induced toxicity (Fig. 6).

Interactive effect of nitric oxide (NO) and 24-epibrassinolide (EBL) on malondialdehyde contents, electrolyte leakage, and reactive oxygen species generation under chromium (Cr) (100 µM) stress. a MDA contents, b electrolyte leakage, c H2O2, d O2•‒, and e MG contents. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)

Effect of nitric oxide (NO) and 24-epibrassinolide (EBL) alone or in combination on the histochemical staining of soybean leaves under chromium (Cr) (100 µM) stress to identify the accumulation of hydrogen peroxide (H2O2) and superoxide (O2•‒) by 3,3-diaminobenzidine (DAB) and nitro blue tetrazolium (NBT), respectively
Effect of EBL, NO, and EBL + NO on MG detoxification mechanism
Data revealed that the combined treatment of EBL + NO slightly upsurged the level of GlyI, and GlyII values under control group without Cr in comparison with the EBL and NO alone. However, a non-significant difference was perceived among EBL, NO, and H2O treatments (Fig. 7a, b). In contrast, H2O + Cr treatment increased the values of GlyI and GlyII by 61.5% and 54.7%, respectively, than Cr-untreated plants of H2O treatment. Interestingly, the alone applications of EBL or NO under Cr toxicity boosted the GlyI and GlyII values by 21.8/22.9% and 27.4/29.8%, correspondingly, as compared to the H2O + Cr-treated plants. Moreover, EBL + NO + Cr increased the amplification of GlyI and GlyII by 19.9% and 22.6%, respectively, compared with EBL and NO alone under Cr toxicity (Fig. 7).

Effect of nitric oxide (NO) and 24-epibrassinolide (EBL) alone or in combination on a glyoxalase I (GlyI) and b glyoxalase II (GlyII) activities of soybean plants under chromium (Cr) (100 µM) stress. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Effect of EBL, NO, and EBL + NO on antioxidant activities
In comparison with H2O treatment without Cr., the alone application of Cr considerably enhanced the activities of SOD by 65.6%, CAT by 75.3%, APX by 67.5%, GSH by 50.9%, and GSSG by 62.7% (Fig. 7a, b, c, e, f). As compared to the H2O + Cr, the EBL + Cr, and NO + Cr treatments stimulated the SOD by 20.5/24.1%, CAT by 23.2/26.7%, APX by 19.1/22.6%, GSH by 23.9/24.5%, and GSSG by 27.3/29.1%, respectively (Fig. 8). It is worth noting that EBL + NO + Cr could increase the above parameters by 17.2%, 24.7%, 19.7%, 26.9%, and 22.3% compared with EBL + Cr and NO + Cr, respectively. In contrast, the alone Cr disclosure in nutrient media solution caused inhibition in the GR values by 50.9% in comparison with the H2O treatment without Cr. Although relative to the H2O + Cr, the EBL or NO application alone improved the GR content level by 32.6% and 33.3%, separately under Cr-induced stress (Fig. 8d). In addition, the EBL + NO + Cr treatment further enhanced the GR values by 21.9% as compared to their alone applications under Cr exposure (Fig. 8d). However, a non-significant difference was noticed among the control group while EBL + NO represented a minor upsurge in the values of CAT, APX, and GR.

Effect of nitric oxide (NO) and 24-epibrassinolide (EBL) applied alone or in combination on antioxidant enzymes and non-enzymatic activities in soybean plants subjected to chromium (Cr) (100 µM) stress: a SOD, b CAT, c APX, d GR, e GSH, f GSSG. Bars annotated with different letters indicate significant differences at p < 0.05. Data are the means and standard deviations of four replicates (n = 4)
Discussion
Once Cr is accumulated inside plant tissues, it hinders numerous morpho-physiological, metabolic, biochemical, as well as molecular attributes, which cause diminishing of embryonic growth. Herein, the aim of the current study was to scrutinize the potential mechanism adopted by EBL and NO solely and/or combined application to cope with the Cr-induced phytotoxicity by underlining plant growth, biomass, photosynthetic, and osmolyte attributes, Cr accumulation, and antioxidant activities. There is a gap in the literature related to the combined supplementation of EBL and NO against Cr exposure in soybean seedlings. Our outcomes clearly showed the Cr-tempted phytotoxicity on soybean seedlings and significantly hindered their biomass as well as growth (Fig. 1). This decline may be occurred due to high Cr accumulation and inhibition in EBL and/or NO accretion inside soybean plants. Similarly, the inhibition in growth and biomass indices of soybean plants was observed in earlier studies as well (Basit et al. 2022b; Wen et al. 2022). It probably happened due to the damage of chlorophyll and the reduction in protein synthesis. A higher accumulation of Cr contents may cause a clear decrease in root elongation. Possibly, the Cr greater accumulation suppression of cell growth, elongation, and division at the root apex further extended the cell-cycle duration (Singh et al. 2013). Nonetheless, the exogenous supplementation of EBL and/or NO noticeably enhanced the plant growth attributes, accumulation of biomass (Fig. 1) by restricting the Cr uptake, and accumulation in plant tissues (Fig. 3). Probably, EBL and/or NO improved the cell elongation, division, and chlorophyll pigments synthesis, which may increase the soybean root and shoot growth as well as biomass accumulation. The plant growth and biomass accumulation were enhanced by the alone application of EBL (Choudhary et al. 2012; Sharma et al. 2022a) and NO (Basit et al. 2022b; Huang et al. 2018). However, the EBL and NO interplayed to restore the biomass and growth traits in pepper plants (Kaya et al. 2019).
In a recent study, Cr toxicity caused a momentous decline in the values of chlorophyll contents (total chlorophyll and carotenoids contents), as well as gas exchange parameters (net photosynthetic rate, CO2 concentration, stomatal conductance, and Fv/m level) (Fig. 2). As well, the visible indications of soybean leave chlorosis (Fig. S1) under alone exposure to Cr also supported our results. Possibly, the Cr accumulation caused the inhibition of specific biosynthetic enzymes (δ-aminolaevulinic acid activities and protochlorophyllide reductase activities), which caused strong disruption inside the biosynthetic pathway responsible for chlorophyll content production. Consequently, increased cellular oxidative damage further instigated chloroplast impairment (Ganesh et al. 2008; Hu et al. 2017; Yang et al. 2021). However, the supply of EBL and/or NO amplified the light-harvesting pigments, which proposed the defensive behavior of EBL and/or NO on photosynthetic pigments by stimulating the chlorophyll pigment production. Prior investigations also proved the positive effects of EBL or NO alone supply on the biosynthesis of chlorophyll contents under Cr-contaminated conditions in radish (Choudhary et al. 2012), lead-stressed in brassica (Soares et al. 2020), and watermelon seedlings (Khan et al. 2021), as well as cadmium stress in mustard (Per et al. 2017). Although, the literature concerning with combined effect of EBL and NO on light-harvesting pigments is still unknown in soybean cultivars under Cr-induced oxidative damage. Relatively, the Cr accumulation was noticed greater in roots than in shoots of soybean seedlings (Fig. 3), which hampered the higher retention ability of Cr and restrained its translocation from the root toward the shoot. Similarly, more accumulation of Cr was noticed inside the root of rice (Basit et al. 2021a) and brassica (Ulhassan et al. 2019) than in shoots. The supplementation of EBL or NO hindered the Cr uptake and translocation (Fig. 3) and stabilized the cell membrane. Most probably, the EBL or NO made complexes with Cr ions by acting as metal-binding agents and therefore lowered the accumulation of Cr inside soybean tissues. Formerly, it is reported that photosynthetic attributes regulated the TSS and TSP under stressed environments (Gao et al. 2020; Mostofa and Fujita 2013; Oumarou Abdoulaye et al. 2019). In these findings, photosynthetic pigments also positively interacted with TSS and TSP under Cr stress. As well, the EBL and NO boosted the chlorophyll contents and gas exchange attributes by modulating the TSS and TSP that might help to counter the Cr toxic effects and assist the energy and metabolite stimulations. As per expectations, an austere water loss and, as a result, proline accumulation were observed in the leaves of soybean seedlings under a Cr-contaminated environment (Fig. 4). Likewise, the accumulation of proline contents and water loss in response to photosynthetic attribute impairment in different plant species is reported under HM-induced toxicity, which leads toward cellular dehydration and robust accumulation of osmoprotectants (i.e., proline contents) (Mostofa and Fujita 2013; Sharma et al. 2020; Singh and Prasad 2019; Singh et al. 2020; Ulhassan et al. 2019). Current findings supported previously testified studies and displayed a significant accumulation in proline contents to overcome the water losses under Cr-mediated stress (Fig. 4a, b). Interestingly, the EBL and/or NO applications regulated the proline metabolism to restrict the water loss and improve the growth as well as photosynthetic traits under a Cr-stressed environment (Figs. 2 and 3). These upshots verified the EBL and/or NO-based mechanistic stance to cope with the ROS over-generation by modulating the proline greater accumulation and improving leaf water status in plants (Singh et al. 2013). Subsequently, shelter the cellular functions of plants by constraining the cellular dehydration under Cr-induced oxidative stress as stated in brassica plants (Ulhassan et al. 2019) and solanum nigrum (UdDin et al. 2015) seedlings.
In this research, Cr exposure caused a predominant increase of reactive oxygen species (ROS), MDA contents, electrolyte leakage as well as methylglyoxal contents in soybean seedlings, which confirmed that the higher Cr accumulation in plants stimulated the Cr-induced oxidative stress, lipid oxidation, and considerable membrane damage as well. Several studies reported a higher generation of the above-described parameters under Cr disclosure in brassica (Sharma et al. 2022b), maize (Malik et al. 2022), wheat (Ashraf et al. 2022), and spinach (Hussain et al. 2022). The exogenous supply of EBL and/or NO substantially scavenged the ROS over-production (Fig. 5), alleviated the negative effect induced by cellular oxidative stress, and improved the performance of photosynthetic attributes. As well, EBL and/or NO improved the membrane stability by lowering the MDA contents as well as EL and MG levels in soybean seedlings under Cr stress. Results documented that the EBL + NO can detoxify the ROS more efficiently as compared to their alone applications (EBL or NO). Perhaps, the EBL and/or NO treatment maintained the cellular metabolism by alleviating the extra-generation of free radicles and, thus, upholding the water status. Former investigations also testified that alone supply of EBL or NO can restrain the ROS extra-production, MDA, EL, and MG level in Ficus concinna plants (Jin et al. 2015), maize (Kharbech et al. 2020b), and tobacco seedlings (Yadav et al. 2005).
To mitigate the cellular oxidative damage caused by ROS over-generation, plants stimulate their antioxidative defense system. Under Cr-induced toxicity, soybean seedlings upsurged the SOD, CAT, APX, GSH, and GSSG and lowered the GR (Fig. 8). However, the EBL and/or NO further boosted the above-mentioned indices, while lowered the GSSG under Cr-contaminated environment. These modulations of antioxidant (enzymatic and non-enzymatic) activities indicated the EBL and/or NO exogenous application has capability to detoxify the ROS accumulation induced by Cr stress. Possibly, the EBL and/or NO treatment sustained the redox balance, which was the major source of ROS excessive generation and MG contents under Cr-induced toxicity inside cellular compartments. The regulation of these antioxidant activities is also documented by individual application of EBL (Kaya et al. 2020; Soares et al. 2020; Vázquez et al. 2013) and NO (Aziz et al. 2015; Basit et al. 2022e; Fancy et al. 2017).
Methylglyoxal (MG) is known as a cytotoxic compound produced through glycolysis pathway against abiotic stresses. Higher production of MG causes disturbance inside antioxidative defense mechanism (Hasanuzzaman et al. 2011). To cope with the greater production of MG contents, plants stimulate the GlyI and GlyII activities, which probably detoxify the MG contents via using glyoxalase system (Kharbech et al. 2020b). Our findings supported the prior investigations by demonstrating the higher MG contents under Cr stress (Fig. 5e), whereas the EBL and/or NO application lowered the MG level by upgrading the GlyI and GlyII activities (Fig. 7a, b), which further detoxified the MG contents by using glyoxalase pathway and converted it into GSH contents (Fig. 8e). Conceivably, the higher level of GSH contents by alone and combined application of EBL and NO under Cr-induced oxidative damage is the key point to mitigate the Cr toxicity in soybean cultivars.
Conclusions
We have evaluated the mitigating role of exogenous applications of EBL and NO, applied alone and in combination, on Cr-induced stress in soybean seedlings (Fig. 9). Both hormones partially ameliorated the effects of Cr; however, the combination of EBL and NO had the greatest effect and reduced both Cr uptake and translocation. This ameliorating effect was seen in the concentrations of light-harvesting pigments, in the antioxidative defense system, and in the accumulation of osmoprotectants and water retention. In addition, the hormones reduced MG contents by stimulating the activities of GlyI and GlyII, which convert the MG to GSH. These effects all resulted in enhancements of plant biomass and growth (Fig. 9). These findings suggested that the interaction of other signaling molecules, such as hydrogen sulfide, methylglyoxal, and hydrogen peroxide, in ameliorating the effects of Cr contamination warrants further investigation.

Schematic diagram to represent the NO and BR-mediated mechanism to mitigate the Cr intoxication in soybean seedlings
Data availability
All data generated or analyzed during this study are included in this published article.
References
Adhikari S, Ghosh S, Azahar I, Adhikari A, Shaw AK, Konar S, Roy S, Hossain Z (2018) Sulfate improves cadmium tolerance by limiting cadmium accumulation, modulation of sulfur metabolism and antioxidant defense system in maize. Environ Exp Bot 153:143–162
Ahmad P, Abdel Latef AA, Hashem A, Abd_Allah EF, Gucel S, Tran L-SP (2016) Nitric oxide mitigates salt stress by regulating levels of osmolytes and antioxidant enzymes in chickpea. Front Plant Sci 7:347
Ashraf MA, Rasheed R, Hussain I, Iqbal M, Farooq MU, Saleem MH, Ali S (2022) Taurine modulates dynamics of oxidative defense, secondary metabolism, and nutrient relation to mitigate boron and chromium toxicity in Triticum aestivum L. plants. Environ Sci Pollut Res 29(30):45527–45548
Aziz H, Sabir M, Ahmad HR, Aziz T, Zia-ur-Rehman M, Hakeem KR, Ozturk M (2015) Alleviating effect of calcium on nickel toxicity in rice. Clean (Weinh) 43:901–909
Basit F, Chen M, Ahmed T, Shahid M, Noman M, Liu J, An J, Hashem A, Fahad Al-Arjani A-B, Alqarawi AA (2021a) Seed priming with brassinosteroids alleviates chromium stress in rice cultivars via improving ROS metabolism and antioxidant defense response at biochemical and molecular levels. Antioxidants 10:1089
Basit F, Liu J, An J, Chen M, He C, Zhu X, Li Z, Hu J, Guan Y (2021b) Brassinosteroids as a multidimensional regulator of plant physiological and molecular responses under various environmental stresses. Environ Sci Pollut Res 28:44768–44779
Basit F, Bhat JA, Dong Z, Mou Q, Zhu X, Wang Y, Hu J, Jan BL, Shakoor A, Guan Y (2022a) Chromium toxicity induced oxidative damage in two rice cultivars and its mitigation through external supplementation of brassinosteroids and spermine. Chemosphere 302:134423
Basit F, Bhat JA, Guan Y, Jan BL, Tyagi A, Ahmad P (2022b) Nitric oxide and spermine revealed positive defense interplay for the regulation of the chromium toxicity in soybean (Glycine max L.). Environ Pollut 308:119602
Basit F, Bhat JA, Han J, Guan Y, Jan BL, Shakoor A, Alansi S (2022c) Screening of rice cultivars for Cr-stress response by using the parameters of seed germination, morpho-physiological and antioxidant analysis. Saudi J Biol Sci 29:3918–3928
Basit F, Liu J, An J, Chen M, He C, Zhu X, Li Z, Hu J, Guan Y (2022d) Seed priming with brassinosteroids alleviates aluminum toxicity in rice via improving antioxidant defense system and suppressing aluminum uptake. Environ Sci Pollut Res 29:10183–10197
Basit F, Ulhassan Z, Mou Q, Nazir MM, Hu J, Hu W, Song W, Sheteiwy MS, Zhou W, Bhat JA (2022e) Seed priming with nitric oxide and/or spermine mitigate the chromium toxicity in rice (Oryza sativa) seedlings by improving the carbon-assimilation and minimising the oxidative damages. Funct Plant Biol 50(2):121–135
Bates LS, Waldren RP, Teare I (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Bradford MM (1976) A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem 72:248–254
Cechin I, Cardoso GS, Fumis TdF, Corniani N (2015) Nitric oxide reduces oxidative damage induced by water stress in sunflower plants. Bragantia 74:200–206
Choudhary SP, Kanwar M, Bhardwaj R, Yu J-Q, Tran L-SP (2012) Chromium stress mitigation by polyamine-brassinosteroid application involves phytohormonal and physiological strategies in Raphanus sativus L. PLoS One 7:e33210
Dubois M, Gilles K, Hamilton J, Rebers P, Smith F (1951) A colorimetric method for the determination of sugars. Nature 168:167–167
El-Shabrawi H, Kumar B, Kaul T, Reddy MK, Singla-Pareek SL, Sopory SK (2010) Redox homeostasis, antioxidant defense, and methylglyoxal detoxification as markers for salt tolerance in Pokkali rice. Protoplasma 245:85–96
Fancy NN, Bahlmann AK, Loake GJ (2017) Nitric oxide function in plant abiotic stress. Plant Cell Environ 40:462–472
Ganesh KS, Baskaran L, Rajasekaran S, Sumathi K, Chidambaram A, Sundaramoorthy P (2008) Chromium stress induced alterations in biochemical and enzyme metabolism in aquatic and terrestrial plants. Colloids Surf B 63:159–163
Gao C, Sheteiwy MS, Han J, Dong Z, Pan R, Guan Y, Alhaj Hamoud Y, Hu J (2020) Polyamine biosynthetic pathways and their relation with the cold tolerance of maize (Zea mays L.) seedlings. Plant Signal Behav 15:1807722
Hasanuzzaman M, Hossain MA, Fujita M (2011) Nitric oxide modulates antioxidant defense and the methylglyoxal detoxification system and reduces salinity-induced damage of wheat seedlings. Plant Biotechnol Rep 5:353–365
Heath RL, Packer L (1968) Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch Biochem Biophys 125:189–198
Hossain MA, Hasanuzzaman M, Fujita M (2010) Up-regulation of antioxidant and glyoxalase systems by exogenous glycinebetaine and proline in mung bean confer tolerance to cadmium stress. Physiol Mol Biol Plants 16:259–272
Hossain MA, Piyatida P, da Silva JAT, Fujita M (2012) Molecular mechanism of heavy metal toxicity and tolerance in plants: central role of glutathione in detoxification of reactive oxygen species and methylglyoxal and in heavy metal chelation. J Bot 2012:1–37
Hu Q, Lin C, Guan Y, Sheteiwy MS, Hu W, Hu J (2017) Inhibitory effect of eugenol on seed germination and pre-harvest sprouting of hybrid rice (Oryza sativa L.). Sci Rep 7:1–9
Hu Y, Bao F, Li J (2000) Promotive effect of brassinosteroids on cell division involves a distinct CycD3-induction pathway in Arabidopsis. Plant J 24:693–701
Huang M, Ai H, Xu X, Chen K, Niu H, Zhu H, Sun J, Du D, Chen L (2018) Nitric oxide alleviates toxicity of hexavalent chromium on tall fescue and improves performance of photosystem II. Ecotoxicol Environ Saf 164:32–40
Husain T, Suhel M, Prasad S, Singh V (2022) Ethylene and hydrogen sulphide are essential for mitigating hexavalent chromium stress in two pulse crops. Plant Biol 24:652–659
Hussain I, Saleem MH, Mumtaz S, Rasheed R, Ashraf MA, Maqsood F, Rehman M, Yasmin H, Ahmed S, Ishtiaq M (2022) Choline chloride mediates chromium tolerance in spinach (Spinacia oleracea L.) by restricting its uptake in relation to morpho-physio-biochemical attributes. J Plant Growth Regul 41:1594–1614
Ista LK, Callow ME, Finlay JA, Coleman SE, Nolasco AC, Simons RH, Callow JA, Lopez GP (2004) Effect of substratum surface chemistry and surface energy on attachment of marine bacteria and algal spores. Appl Environ Microbiol 70:4151–4157
Jan S, Alyemeni MN, Wijaya L, Alam P, Siddique KH, Ahmad P (2018) Interactive effect of 24-epibrassinolide and silicon alleviates cadmium stress via the modulation of antioxidant defense and glyoxalase systems and macronutrient content in Pisum sativum L. seedlings. BMC Plant Biol 18:1–18
Jiang M, Zhang J (2001) Effect of abscisic acid on active oxygen species, antioxidative defence system and oxidative damage in leaves of maize seedlings. Plant Cell Physiol 42:1265–1273
Jin SH, Li XQ, Wang GG, Zhu XT (2015) Brassinosteroids alleviate high-temperature injury in Ficus concinna seedlings via maintaining higher antioxidant defence and glyoxalase systems. AoB plants 21:7
Kaya C, Ashraf M, Wijaya L, Ahmad P (2019) The putative role of endogenous nitric oxide in brassinosteroid-induced antioxidant defence system in pepper (Capsicum annuum L.) plants under water stress. Plant Physiol Biochem 143:119–128
Kaya C, Ashraf M, Alyemeni MN, Ahmad P (2020) The role of nitrate reductase in brassinosteroid-induced endogenous nitric oxide generation to improve cadmium stress tolerance of pepper plants by upregulating the ascorbate-glutathione cycle. Ecotoxicol Environ Saf 196:110483
Khan J, Malangisha GK, Ali A, Mahmoud A, Yang J, Zhang M, Hu Z (2021) Nitric oxide alleviates lead toxicity by inhibiting lead translocation and regulating root growth in watermelon seedlings. Hortic Environ Biotech 62:701–714
Kharbech O, Houmani H, Chaoui A, Corpas FJ (2017) Alleviation of Cr (VI)-induced oxidative stress in maize (Zea mays L.) seedlings by NO and H2S donors through differential organ-dependent regulation of ROS and NADPH-recycling metabolisms. J Plant Physiol 219:71–80
Kharbech O, Massoud MB, Sakouhi L, Djebali W, Mur LAJ, Chaoui A (2020a) Exogenous application of hydrogen sulfide reduces chromium toxicity in maize seedlings by suppressing NADPH oxidase activities and methylglyoxal accumulation. Plant Physiol Biochem 154:646–656
Kharbech O, Sakouhi L, Massoud MB, Mur LAJ, Corpas FJ, Djebali W, Chaoui A (2020b) Nitric oxide and hydrogen sulfide protect plasma membrane integrity and mitigate chromium-induced methylglyoxal toxicity in maize seedlings. Plant Physiol Biochem 157:244–255
Kohli SK, Handa N, Bali S, Arora S, Sharma A, Kaur R, Bhardwaj R (2018) Modulation of antioxidative defense expression and osmolyte content by co-application of 24-epibrassinolide and salicylic acid in Pb exposed Indian mustard plants. Ecotoxicol Environ Saf 147:382–393
Kushwaha BK, Singh S, Tripathi DK, Sharma S, Prasad SM, Chauhan DK, Kumar V, Singh VP (2019) New adventitious root formation and primary root biomass accumulation are regulated by nitric oxide and reactive oxygen species in rice seedlings under arsenate stress. J Hazard Mater 361:134–140
Kwasniewski M, Chwialkowska K, Kwasniewska J, Kusak J, Siwinski K, Szarejko I (2013) Accumulation of peroxidase-related reactive oxygen species in trichoblasts correlates with root hair initiation in barley. J Plant Physiol 170:185–195
Law M, Charles SA, Halliwell B (1983) Glutathione and ascorbic acid in spinach (Spinacia oleracea) chloroplasts. The effect of hydrogen peroxide and of paraquat. Biochem J 210:899–903
Malik Z, Afzal S, Dawood M, Abbasi GH, Khan MI, Kamran M, Zhran M, Hayat MT, Aslam MN, Rafay M (2022) Exogenous melatonin mitigates chromium toxicity in maize seedlings by modulating antioxidant system and suppresses chromium uptake and oxidative stress. Environ Geochem Health 44:1451–1469
Mostofa MG, Fujita M (2013) Salicylic acid alleviates copper toxicity in rice (Oryza sativa L.) seedlings by up-regulating antioxidative and glyoxalase systems. Ecotoxicology 22:959–973
Mostofa MG, Seraj ZI, Fujita M (2014) Exogenous sodium nitroprusside and glutathione alleviate copper toxicity by reducing copper uptake and oxidative damage in rice (Oryza sativa L.) seedlings. Protoplasma 251:1373–1386
Mostofa MG, Ghosh A, Li Z-G, Siddiqui MN, Fujita M, Tran L-SP (2018) Methylglyoxal–a signaling molecule in plant abiotic stress responses. Free Radic Biol Med 122:96–109
Nakano Y, Asada K (1981) Hydrogen peroxide is scavenged by ascorbate-specific peroxidase in spinach chloroplasts. Plant Cell Physiol 22:867–880
Naz R, Sarfraz A, Anwar Z, Yasmin H, Nosheen A, Keyani R, Roberts TH (2021) Combined ability of salicylic acid and spermidine to mitigate the individual and interactive effects of drought and chromium stress in maize (Zea mays L.). Plant Physiol Biochem 159:285–300
Ning L-H, Du W-k, Song H-N, Shao H-B, Qi W-C, Sheteiwy MSA, Yu D-y (2019) Identification of responsive miRNAs involved in combination stresses of phosphate starvation and salt stress in soybean root. Environ Exp Bot 167:103823
Oumarou Abdoulaye A, Lu H, Zhu Y, Alhaj Hamoud Y, Sheteiwy M (2019) The global trend of the net irrigation water requirement of maize from 1960 to 2050. Climate 7:124
Per TS, Masood A, Khan NA (2017) Nitric oxide improves S-assimilation and GSH production to prevent inhibitory effects of cadmium stress on photosynthesis in mustard (Brassica juncea L.). Nitric Oxide 68:111–124
Prakash V, Rai P, Sharma NC, Singh VP, Tripathi DK, Sharma S, Sahi S (2022) Application of zinc oxide nanoparticles as fertilizer boosts growth in rice plant and alleviates chromium stress by regulating genes involved in regulating oxidative stress. Chemosphere 303:134554
Ramakrishna B, Rao SSR (2015) Foliar application of brassinosteroids alleviates adverse effects of zinc toxicity in radish (Raphanus sativus L.) plants. Protoplasma 252:665–677
Santos L, Batista B, Lobato A (2018) Brassinosteroids mitigate cadmium toxicity in cowpea plants. Photosynthetica 56:591–605
Schaedle M, Bassham JA (1977) Chloroplast glutathione reductase. Plant Physiol 59:1011–1012
Shanmugam V, Wang YW, Tsednee M, Karunakaran K, Yeh KC (2015) Glutathione plays an essential role in nitric oxide-mediated iron-deficiency signaling and iron-deficiency tolerance in Arabidopsis. Plant J 84:464–477
Sharma A, Kapoor D, Wang J, Shahzad B, Kumar V, Bali AS, Jasrotia S, Zheng B, Yuan H, Yan D (2020) Chromium bioaccumulation and its impacts on plants: an overview. Plants 9:100
Sharma N, Kour S, Kumar D, Kaur R, Khajuria A, Ohri P (2022a) Role of Brassinosteroids (BRs) in modulating antioxidative defense mechanism in plants growing under abiotic and biotic stress conditions, antioxidant defense in plants. Springer, pp 325–367
Sharma P, Chouhan R, Bakshi P, Gandhi SG, Kaur R, Sharma A, Bhardwaj R (2022b) Amelioration of chromium-induced oxidative stress by combined treatment of selected plant-growth-promoting rhizobacteria and earthworms via modulating the expression of genes related to reactive oxygen species metabolism in Brassica juncea. Front Microbiol 13:802512
Shivaraj SM, Vats S, Bhat JA, Dhakte P, Goyal V, Khatri P, Kumawat S, Singh A, Prasad M, Sonah H (2020) Nitric oxide and hydrogen sulfide crosstalk during heavy metal stress in plants. Physiol Plant 168:437–455
Singh AP, Dixit G, Kumar A, Mishra S, Singh PK, Dwivedi S, Trivedi PK, Chakrabarty D, Mallick S, Pandey V (2016) Nitric oxide alleviated arsenic toxicity by modulation of antioxidants and thiol metabolism in rice (Oryza sativa L.). Front Plant Sci 6:1272
Singh HP, Mahajan P, Kaur S, Batish DR, Kohli RK (2013) Chromium toxicity and tolerance in plants. Environ Chem Lett 11:229–254
Singh S, Prasad SM (2019) Management of chromium (VI) toxicity by calcium and sulfur in tomato and brinjal: implication of nitric oxide. J Hazard Mater 373:212–223
Singh S, Prasad SM, Singh VP (2020) Additional calcium and sulfur manages hexavalent chromium toxicity in Solanum lycopersicum L. and Solanum melongena L. seedlings by involving nitric oxide. J Hazard Mater 398:122607
Singh S, Dubey NK, Singh VP (2022) Nitric oxide and hydrogen peroxide independently act in mitigating chromium stress in Triticum aestivum L. seedlings: regulation of cell death, chromium uptake, antioxidant system, sulfur assimilation and proline metabolism. Plant Physiol Biochem 183:76–84
Soares TFSN, dos Santos Dias DCF, Oliveira AMS, Ribeiro DM, dos Santos Dias LA (2020) Exogenous brassinosteroids increase lead stress tolerance in seed germination and seedling growth of Brassica juncea L. Ecotoxicol Environ Saf 193:110296
Szekeres M, Németh K, Koncz-Kálmán Z, Mathur J, Kauschmann A, Altmann T, Rédei GP, Nagy F, Schell J, Koncz C (1996) Brassinosteroids rescue the deficiency of CYP90, a cytochrome P450, controlling cell elongation and de-etiolation in Arabidopsis. Cell 85:171–182
UdDin I, Bano A, Masood S (2015) Chromium toxicity tolerance of Solanum nigrum L. and Parthenium hysterophorus L. plants with reference to ion pattern, antioxidation activity and root exudation. Ecotoxicol Environ Saf 113:271–278
Ulhassan Z, Gill RA, Huang H, Ali S, Mwamba TM, Ali B, Huang Q, Hamid Y, Khan AR, Wang J (2019) Selenium mitigates the chromium toxicity in Brassicca napus L. by ameliorating nutrients uptake, amino acids metabolism and antioxidant defense system. Plant Physiol Biochem 145:142–152
Vázquez MN, Guerrero YR, González LM, de la Noval WT (2013) Brassinosteroids and plant responses to heavy metal stress. An Overview Open J Metal 3:34–41
Wang H, Ji F, Zhang Y, Hou J, Liu W, Huang J, Liang W (2019) Interactions between hydrogen sulphide and nitric oxide regulate two soybean citrate transporters during the alleviation of aluminium toxicity. Plant Cell Environ 42:2340–2356
Wen K, Li X, Huang R, Nian H (2022) Application of exogenous glutathione decreases chromium translocation and alleviates its toxicity in soybean (Glycine max L.). Ecotoxicol Environ Saf 234:113405
Yadav SK, Singla-Pareek SL, Ray M, Reddy M, Sopory S (2005) Methylglyoxal levels in plants under salinity stress are dependent on glyoxalase I and glutathione. Biochem Biophys Res Commun 337:61–67
Yang S, Ulhassan Z, Shah AM, Khan AR, Azhar W, Hamid Y, Hussain S, Sheteiwy MS, Salam A, Zhou W (2021) Salicylic acid underpins silicon in ameliorating chromium toxicity in rice by modulating antioxidant defense, ion homeostasis and cellular ultrastructure. Plant Physiol Biochem 166:1001–1013
Yokota T, Mori K (2018) Molecular structure and biological activity of brassinolide and related brassinosteroids, molecular structure and biological activity of steroids. CRC Press, pp 317–340
Zhang J, Wei J, Li D, Kong X, Rengel Z, Chen L, Yang Y, Cui X, Chen Q (2017) The role of the plasma membrane H+-ATPase in plant responses to aluminum toxicity. Front Plant Sci 8:1757
Zheng Y, Hu J, Zhang S, Gao C, Song W (2006) Identification of chilling-tolerance in maize inbred lines at germination and seedling growth stages. Journal of Zhejiang University (Agriculture and Life Science) 32:41–45
Zhou W, Leul M (1999) Uniconazole-induced tolerance of rape plants to heat stress in relation to changes in hormonal levels, enzyme activities and lipid peroxidation. Plant Growth Regul 27:99–104
Funding
This research was supported by the Hainan Provincial Joint Project of Sanya Yazhou Bay Science and Technology City (No. 320LH032), the Hainan Provincial Natural Science Foundation of China (No. 322CXTD522), the National Natural Science Foundation of China (Zhejiang Provincial Natural Science Foundation (No. LY21C130006) and Collaborative Innovation Center for Modern Crop Production co-sponsored by Province and Ministry (CIC-MCP).
Author information
Authors and Affiliations
Contributions
Farwa Basit: conceptualization, investigation, and writing—original draft. Yajing Guan, Jin Hu, and Farwa Basit: conceptualization and investigation. Ji Tao and Mohamed Salah Sheteiwy: validation, writing, reviewing, and editing. Izabela Jośko and Yajing Guan: investigation and formal analysis. Mohamed Salah Sheteiwy, Paul Holford, and Jianyu An: investigation. Farwa Basit and Izabela Jośko: writing, reviewing, and editing.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Responsible Editor: Gangrong Shi
Publisher's note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Springer Nature or its licensor (e.g. a society or other partner) holds exclusive rights to this article under a publishing agreement with the author(s) or other rightsholder(s); author self-archiving of the accepted manuscript version of this article is solely governed by the terms of such publishing agreement and applicable law.
About this article

Cite this article
Basit, F., Tao, J., An, J. et al. Nitric oxide and brassinosteroids enhance chromium stress tolerance in Glycine max L. (Merr.) by modulating antioxidative defense and glyoxalase systems. Environ Sci Pollut Res 30, 51638–51653 (2023). https://doi.org/10.1007/s11356-023-25901-0
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11356-023-25901-0