
Abstract
Dietary interventions such as caloric restriction (CR) extend lifespan and health span. Recent data from animal and human studies indicate that CR slows down the aging process, benefits general health, and improves memory performance. Caloric restriction also retards and slows down the progression of different age-related diseases, such as Alzheimer’s disease. However, the specific molecular basis of these effects remains unclear. A better understanding of the pathways underlying these effects could pave the way to novel preventive or therapeutic strategies. In this review, we will discuss the mechanisms and effects of CR on aging and Alzheimer’s disease. A potential alternative to CR as a lifestyle modification is the use of CR mimetics. These compounds mimic the biochemical and functional effects of CR without the need to reduce energy intake. We discuss the effect of two of the most investigated mimetics, resveratrol and rapamycin, on aging and their potential as Alzheimer’s disease therapeutics. However, additional research will be needed to determine the safety, efficacy, and usability of CR and its mimetics before a general recommendation can be proposed to implement them.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Modifying lifestyle factors such as basic dietary regimes and limitation of food intake have an undeniable effect on lifespan and health span (Witte et al. 2009). Among these dietary interventions, caloric restriction (CR), defined here as reduction of energy intake that does not cause undernutrition combined with normal intake of vitamins, minerals, and essential biomolecules, is believed to retard the age-related health decline and to extend lifespan in different organisms. This research field has gained considerable attention since the first study published by McCay and colleagues, who demonstrated that CR increased the lifespan of rats in comparison to rats fed ad libitum (AL) (McCay et al. 1989). Since then, many investigators have confirmed this observation in different model systems. In mammals, the effect of CR is associated with diverse health benefits: reduction of abdominal fat mass, lowered body temperature and metabolic rate, increased insulin sensitivity, and reduction of levels of pro-inflammatory cytokines, reactive oxygen species (ROS) and atherosclerotic lipids in the blood (Murphy et al. 2014; Redman and Ravussin 2011; Weiss and Fontana 2011). However, the mechanism by which CR counteracts the deleterious effects of aging remains controversial. Numerous hypotheses have been proposed to explain the molecular basis of how CR prolongs lifespan. These hypotheses include attenuation of immunologic and hormonal changes, enhanced damage-repair capacity, changes in gene expression, enhanced autophagy and apoptosis, alterations in insulin-like growth factor (IGF) and insulin signaling, mammalian target of rapamycin (mTOR) signaling, hormesis, activation of sirtuins, decreased body temperature and metabolic rate, and attenuation of ROS generation and oxidative stress as summarized in Table 1 (Sohal and Forster 2014). Accumulating data indicate that not only CR but also specific diet components such as proteins, amino acids, and lipids may affect age-associated intracellular signaling pathways (Hulbert 2005; Martin-Montalvo et al. 2013; Pamplona et al. 2002; Perrone et al. 2013; Sanchez-Roman and Barja 2013). Furthermore, CR affects not only lifespan but—most importantly—also health span, which refers to the number of years of life without pathology or disease. CR can delay different age-associated diseases, such as cancer, diabetes, atherosclerosis, cardiovascular diseases and neurodegenerative diseases (Holloszy and Fontana 2007; Li et al. 2011). Interestingly, young Alzheimer’s disease (AD) transgenic mice models subjected to short-term CR displayed reduced accumulation of amyloid β (Aβ) plaques and activation of astrocytes (Patel et al. 2005). Long-term CR led to similar results (Mouton et al. 2009; Wang et al. 2005) and was associated with up‐regulation of genes associated with neurogenesis and neuroplasticity, and down-regulation of inflammatory genes (Wu et al. 2008).
The proportion of people reaching the age of 65 years has increased and so has their life expectancy. It is crucial to gain a deeper understanding of the mechanisms of aging and the mechanisms by which limitation of energy intake can influence aging and the age-related decline in health. Such research may pave the way towards novel strategies to maintain cognitive brain functions throughout life and interventions to delay aging and age-related degenerative diseases in humans.
In this review, we shed light on the effects of CR on aging and age-related neurodegenerative brain diseases, especially AD, in animal models as well as humans. In addition, we discuss the epigenetic involvement in dietary interventions and CR mimetics (CRMs), which provide interesting opportunities for novel preventive and therapeutic strategies in aging and age-related diseases.
Caloric restriction’s effects on aging (lifespan) and neurodegeneration (health span)
Caloric restriction regimes
The influence of different dietary interventions, including CR, intermittent fasting (IF), and specific diet components on lifespan and health span, has been examined in diverse model organisms. CR regimes usually comprise a consistent 20–40 % reduction in caloric intake below that of AL energy intake. In most CR rodent models, carbohydrates, lipids, and proteins are reduced while intake of all essential nutrients such as vitamins and minerals is preserved. This energy intake reduction is known to increase longevity by about 30 % and to delay aging by improving insulin sensitivity and metabolic function (Li et al. 2011; Masoro 2005; Pugh et al. 1999; Wood et al. 2015). The effects of CR have been evaluated in animals of different ages and for different durations. Future studies might define the ideal CR diet to optimize cognition and longevity (Patel et al. 2005).
A second intensively investigated dietary intervention is IF, which involves alternating between periods of AL caloric intake and partial or complete restriction of caloric intake while maintaining intake of all essential nutrients (Mattson et al. 2003; Murphy et al. 2014). Overall, IF results in a 20–30 % reduction of caloric intake (Martin et al. 2006; Ribaric 2012). Animal studies showed a positive effect of IF on age-related diseases, for example an increased resistance to oxidative and metabolic insults and an enhancement of neuronal plasticity including neurotrophic factors such as brain-derived neurotrophic factor (BDNF), protein chaperones such as heat-shock proteins, and mitochondrial uncoupling proteins (Duan et al. 2001; Lee et al. 2000; Mattson 2015; Mattson et al. 2003).
Accumulating data indicate that not only CR but also specific diet components such as proteins, sulfur-containing amino acids (methionine and cysteine), and lipids may act as regulators of age-associated intracellular signaling pathways (Hine et al. 2015; Hine and Mitchell 2015; Hulbert 2005; Martin-Montalvo et al. 2013; Pamplona et al. 2002; Perrone et al. 2013; Sanchez-Roman and Barja 2013). In the yeast Saccharomyces cerevisiae (S. cerevisiae), ammonium was shown to have a negative effect on extending the chronological lifespan (i.e., the survival time of populations of non-dividing cells), especially in amino acid restricted cells. Indeed, lifespan shortening under amino acid restriction could be completely reverted by removing ammonium from the culture medium (Santos et al. 2015). Protein restriction (PR) and methionine restriction (MetR) have been shown to increase the maximum lifespan in rodent models of CR. Interestingly, only 7 weeks of 40 % PR or 40 % MetR resulted in a decrease of mitochondrial ROS production and DNA damage (Sanchez-Roman and Barja 2013). MetR has even been suggested to be responsible for the positive effects observed in PR animals, because in rodent models MetR resulted in reduced adipose tissue mass, improved insulin sensitivity, and decreased oxidative damage and inflammation (Martin-Montalvo et al. 2013; Perrone et al. 2013). In mammals, however, the combination of methionine and cysteine restriction is required in order to observe health benefits such as stress resistance and extended longevity (Elshorbagy et al. 2013), and therefore can better be referred to as sulfur amino acid (SAA) restriction. Recent research suggests that the increased activity of the transsulfuration pathway (TSP) is an evolutionary conserved response to multiple CR regimes from yeast to mammals and the resulting increase in hydrogen sulfide (H2S) production represents a common molecular mechanism underlying several CR benefits (Hine et al. 2015; Hine and Mitchell 2015). However, much remains to be determined on the extent by which the beneficial effects of CR depend on H2S, as well as on the molecular mechanisms underlying these effects (Hine and Mitchell 2015).
Furthermore, research has shown that the fatty acid composition of membranes, via its influence on lipid peroxidation, is an important determinant of lifespan (Hulbert 2005) and may prevent lipoxidation-derived damage to other macromolecules (Pamplona et al. 2002).
In the following section, we will briefly discuss the key mechanisms underlying the beneficial effects during CR; we will focus further on the effects of CR on brain aging and AD disease models and discuss the findings from animal models and human studies.
Mechanisms of CR
The exact mechanisms through which CR promotes health and lifespan are complex and still not fully understood. Nevertheless, numerous studies were designed to unravel the responses to dietary restriction. Different connections between longevity and caloric intake have been hypothesized including mTOR, IGF and insulin signaling, activation of sirtuins, adenosine monophosphate-activated protein kinase (AMPK) signaling and enhanced autophagy and apoptosis, ROS generation, oxidative stress and epigenetics (Table 1).
The mTOR signaling controls growth, metabolism, and energy homeostasis in a cell autonomous manner. mTOR is a serine-threonine kinase and functions as an intracellular energy sensor (Finkel 2015). mTOR becomes activated after growth factor stimulation or increasing levels of amino acids and regulates the overall metabolism (Albert and Hall 2015). CR seems to lead to the genetic inhibition of mTOR signaling resulting in lifespan extension in multiple organisms ranging from Caenorhabditis elegans (C. elegans) (Vellai et al. 2003), Drosophila (Kapahi et al. 2004), to mammals (Wu et al. 2013). The exact mechanism however remains unclear and it seems likely that CR operates by many mechanisms in addition to decreased mTOR signaling (Goldberg et al. 2015). This is strengthened by the observed different metabolic outcomes in aged rapamycin-treated mice and diet-restricted mice (Miller et al. 2014).
Insulin and insulin receptors regulate multiple metabolic functions including glucose transport, oxidative stress response, neurotransmission, and lifespan (Mattson 2002; Murphy et al. 2003; Schulingkamp et al. 2000; Srivastava and Haigis 2011). Reduction of insulin/IGF-1 signaling has been shown to extend lifespan in C. elegans (Kenyon et al. 1993; Kimura et al. 1997; Murphy et al. 2003), Drosophila models (Tatar et al. 2001), and mammals (Anisimov and Bartke 2013). The mammalian forkhead box (FOXO) family of transcription factors act as key regulators of longevity downstream of insulin and IGF signaling during CR and regulate cell survival mechanisms (Martins et al. 2016).
Sirtuins have been reported to be involved in regulating aging and CR-related lifespan extension (Leibiger and Berggren 2006). They have a function in the nicotinamide adenine dinucleotide (NAD)-dependent deacetylation of both histone and non-histone proteins. Sirtuin 1 (SIRT1) was initially discovered from its role in chromatin remodeling associated with gene silencing and the prolongation of lifespan in yeast (Brachmann et al. 1995). The CR-induced SIRT1 expression, which promotes lifespan extension, has been demonstrated in different model systems like C. elegans (Wang et al. 2006), Drosophila (Rogina and Helfand 2004), mammals (Cohen et al. 2004), and in humans (Bordone et al. 2007; Cohen et al. 2004; Guarente and Picard 2005; Haigis and Guarente 2006; Lin et al. 2000; Wakeling et al. 2009). Lin and colleagues suggested that the increased longevity induced by CR requires the activation of SIRT1 by NAD in S. cerevisiae (Lin et al. 2000). More recently, other reports challenged the involvement of sirtuins in the CR-mediated effects observed in yeast (Finkel 2015). SIRT1 is able to deacetylate different molecules, including key transcriptional factors and regulatory proteins, including cellular tumor antigen p53, FOXO proteins, and peroxisome proliferator-activated receptor-γ coactivator (PGC) 1α (Guarente and Picard 2005; Wakeling et al. 2009). SIRT1-mediated deacetylation and consequent downregulation of p53 and FOXO protein may affect lifespan by negatively regulating cellular apoptosis and replicative senescence processes downstream of these transcription factors (Langley et al. 2002; Li et al. 2011; Luo et al. 2001; Vaziri et al. 2001). Furthermore, SIRT1 influences metabolic pathways by activating PGC-1α, thereby promoting gluconeogenic genes and improving mitochondrial function by inducing mitochondrial biogenesis (Leibiger and Berggren 2006; Wakeling et al. 2009).
AMPK, a detector of low energy and cell de-energization, has been described as a molecular transducer of beneficial starvation signals in model organisms (Mair et al. 2011; Martin-Montalvo et al. 2013; Pani 2015). In worms, flies, and mammals, dietary restriction requires the activation of AMPK in order to achieve longevity (Canto and Auwerx 2011; Greer et al. 2007; Stenesen et al. 2013), but this activation is dependent on the CR protocol (Canto and Auwerx 2011). Interestingly, AMPK can inhibit the mTOR cascade through the phosphorylation and activation of the mTOR inhibitory tuberous sclerosis complex (TSC), TSC1/TSC2 (Inoki et al. 2003). In addition in mammals, AMPK is known to regulate autophagy through the phosphorylation of one of its many targets, UNC-51-like kinase 1 (ULK1).
Autophagy is the process whereby damaged organelles or macromolecules form autophagosomes that can fuse with lysosomes allowing enzymatic digestion of the cargo (Mizushima and Komatsu 2011), and this process can be described as a cell rejuvenating mechanism (Pani 2015). Autophagy is tightly regulated by nutrient availability and monitored by a number of nutrient sensors and cascades including the AMPK-mTOR circuitry (Pani 2015) and has a double purpose namely the recycling of cellular waste that can be used for either novel macromolecules or as energy source (Mizushima and Komatsu 2011). Autophagy is transcriptionally down-regulated during normal aging in the human brain (Lipinski et al. 2010), and the experimental impairment of autophagy decreased lifespan and earlier features of cellular aging in worms, flies and mice (Cuervo 2008; Juhasz et al. 2007; Pyo et al. 2013; Simonsen et al. 2008; Toth et al. 2008).
Increased generation of ROS has been postulated as one of the major hallmarks of aging. Restriction of caloric intake lowered levels of oxidative stress and damage in mammals (Sohal and Weindruch 1996). Similarly, rodents subjected to CR showed attenuation of age-associated increases in rates of mitochondrial and hydrogen peroxide (H2O2) generation and a reduction in oxidative damage (Sohal et al. 1994; Sohal and Weindruch 1996).
Recently, the role of epigenetics has also been recognized as a major player in controlling aging and in the influence of CR on lifespan. Epigenetics refers to any change in gene regulation that is caused by modifications to the DNA’s packaging proteins or by changes in the DNA molecules that do not affect its sequence. The two principal epigenetic changes, DNA methylation and histone modification, are thought to be a way by which environmental factors such as nutrition can influence genetics and different biological processes (Lord and Cruchaga 2014). DNA methylation, which occurs mainly on cytosine residues of CpG dinucleotides, can dynamically regulate gene expression and maintain DNA integrity and stability in numerous biological processes, including aging (Chan et al. 2000; Li et al. 1993, 2011). Aging has been reported to be tightly regulated by DNA methylation. A decrease in global DNA methylation has been reported, but the promoter regions often become methylated during aging, which decreases gene expression. Interestingly, CR likely reverses these local aberrant DNA methylation patterns by locus specific control in order to maintain chromatin function and increase genomic stability (Li et al. 2011; Munoz-Najar and Sedivy 2011). Moreover, acetylation and deacetylation at the lysine residues in histones are the most prevalent histone modifications determining the level of gene activity. Acetylation, leading to a weaker binding of nucleosomal components so more transcription factors are able to bind, is known to increase the expression of genes through transcriptional activation. In contrast, deacetylation of histones leads to decreased levels of gene expression or gene silencing. In this process, histone deacetylases (HDAC) interact with different transcription factors (de Ruijter et al. 2003; Kadonaga 1998), and these interactions are important in the regulation of gene expression and in different cellular functions. The activity of HDACs is increased during CR, indicating that deacetylation might be a protective mechanism during CR promoting longevity (Kanaya et al. 1998; Li et al. 2010; Meyerson et al. 1997). An interesting class III NAD-dependent HDAC is SIRT1 as discussed above. The findings on epigenetic changes during CR were corroborated by a recent transcriptome analysis in which RNA sequencing of rat cerebral cortex identified overexpression of neuroprotective genes in response to CR. This gene expression profile pointed to a role for miRNA (miR-98-3p) in maintaining the homeostasis of HDAC and histone acetyltransferase (HAT) (Wood et al. 2015).
The health benefits of CR on brain aging
Aging is characterized by numerous functional and structural alterations, including improper functioning of the CNS barriers (Gorlé et al. 2016), that together impair brain plasticity. Dietary interventions such as CR seem to improve the resilience of synapses to metabolic and oxidative damage and are associated with greater synaptic activity, leading to higher synaptic plasticity and stimulation of neuroprotective pathways in the brain (Mattson 2012; Murphy et al. 2014; Park and Prolla 2005). In addition, generation of neurotrophic factors in particular BDNF in response to CR contributes to the neuroprotective effect of reduced energy intake (Bekinschtein et al. 2011; Mattson 2002; Murphy et al. 2014; Rothman et al. 2012; Vivar et al. 2013). Neurotrophic factors promote synaptic function and plasticity and are crucial for neuronal survival, morphology, and differentiation in the adult central nervous system (CNS) (Reichardt 2006). Furthermore, CR leads to a major reduction in body fat mass and exerts beneficial effects on age-related processes, including insulin resistance, inflammation, and oxidative stress in diverse organisms ranging from yeast to higher mammals (Colman et al. 2009; Fontana et al. 2010; Redman and Ravussin 2011).
Animal studies
The influence of CR on aging, neuroplasticity, neuroprotection and cognitive performance has been evaluated in different animal models. The first studies on dietary restriction indicated that it promotes longevity and cognition: CR was demonstrated to enhance spatial memory performance in rats (Stewart et al. 1989) and further food restriction over 4 months reduced age-related impairment of motor and learning tasks in mice (Ingram et al. 1987). More recent studies reported similar beneficial effects on learning and memory in rodents. These benefits could be gained by reducing caloric intake throughout life, starting in mid-life (Means et al. 1993), and by short CR later in life (Goto et al. 2007; Kaur et al. 2008; Sharma et al. 2010). In a rat model, in addition to cognitive decline, the decrease in key synaptic proteins like synaptophysin (SPH), N-methyl-d-aspartate (NMDA)-type, and alpha-amino-3-hydroxy-5-methyl-4-isoxazole propionic acid (AMPA)-type ionotropic glutamate receptors in the CA3 region of the hippocampus that occur across lifespan is stabilized by CR. This age-related decrease and CR-induced stabilization are likely to affect CA3 synaptic plasticity and, as a result, hippocampal function (Adams et al. 2008). Additionally, protein expression profiling of synapse-related proteins in rats revealed stabilization of the levels of three presynaptic proteins (SPH, growth-associated protein 43 (GAP-43), and α-synuclein (α-Syn)) in the hippocampus in response to long-term CR. This indicates the beneficial effect of CR on age-related decline in the capacity for synaptic plasticity (Mladenovic Djordjevic et al. 2010). Experimental evidence has shown that CR mediates neurogenesis in the dentate gyrus of the hippocampus in rodent brains. This neurogenesis is associated with the increased expression of BDNF (Lee et al. 2000, 2002). The study provided the first evidence that diet can affect neurotrophic factor production. Furthermore, BDNF was shown to promote survival of newly generated neurons and thereby contributes to the enhancement of neurogenesis in response to CR (Lee et al. 2002).
DNA microarray analysis of brain tissue in mice revealed that CR was associated with transcriptional reprogramming in the brain. CR prevented the induction of stress- and immune-response-related genes (Prolla 2002). The results are in agreement with studies suggesting reduction in autoimmunity and oxidative damage (Sohal et al. 1994) in the brain of CR mice (Prolla 2002). Furthermore, transcriptional analysis revealed the induction of growth and neurotrophic factors upon CR (Prolla 2002).
Since the beneficial effects of CR have been observed in diverse species, it became important to investigate them in primates. The initial studies exploring the beneficial consequences of reduced energy intake in rhesus monkeys yielded inconsistent results. The discrepancies in survival and disease between the studies might be explained by differences in diet composition and study design, including different methods of calculating the nutritional demands (Cava and Fontana 2013; Colman et al. 2008, 2009). Nevertheless, there is mounting evidence for the positive effects of CR in primates (Colman et al. 2009). Additionally, in a recent study, a long-term 30 % restricted diet implemented from young adulthood significantly improved age-related and all-cause survival in rhesus monkeys (Colman et al. 2014).
Human clinical trials
The beneficial effects of CR observed in diverse animal models, including primates, paved the way to human trials. A multicenter study on CR, the CALERIE study (Comprehensive Assessment of Long-Term Effects of Reducing Intake of Energy), is a randomized controlled trial that examined the effects on cognitive functioning of 6 months of CR (25 % energy intake reduction) combined with physical exercise in overweight individuals (25 ≤ BMI < 30). The study results indicate that CR was not associated with a consistent pattern of cognitive impairment (Martin et al. 2007; Murphy et al. 2014). However, the limitations of the study have to be taken into account when interpreting these results, namely, the small sample size, limited CR duration and statistical power (Martin et al. 2007).
The ENCORE study (Exercise and Nutrition Interventions for Cardiovascular Health) is a randomized trial that examined the effects of exercise combined with dietary modification on neurocognitive functioning in individuals with high blood pressure. Among sedentary and overweight individuals with prehypertension or hypertension, the combination of exercise and CR improved neurocognitive function (Smith et al. 2010b). The neurocognitive improvement might have been due to the combination of CR, exercise, and weight loss.
In a prospective intervention trial, Witte and colleagues demonstrated the beneficial effects of CR (30 % reduction) over 3 months. This study showed significant improvement in memory performance in a cohort of healthy elderly individuals participating in a CR regime compared to a test group with increased unsaturated fatty acids (UFA) intake and an AL control group (Witte et al. 2009). The authors proposed that improved insulin sensitivity and reduced inflammatory activity could have mediated the beneficial effects of CR on memory functions. The study supports the experimental animal studies and epidemiological studies in humans that point to the beneficial effects of CR on the aging brain, which might lead to higher synaptic plasticity and stimulation of neuroprotective pathways in the brain (Witte et al. 2009).
A recent two-year randomized controlled trial tested the feasibility, safety, and effects of CR in non-obese humans (Ravussin et al. 2015). This study provides the first evidence from a randomized controlled trial that sustained CR is feasible and has no adverse effects on the quality of life of non-obese individuals. Furthermore, some of the potential modulators of longevity observed in this trial resemble those reported in previous CR studies on laboratory animals, including weight loss, lower body temperature, lowered thyroid function and reduction in inflammatory markers (tumor necrosis factor (TNF)-α and high-sensitivity C-reactive protein (CRP)) (Ravussin et al. 2015).
Clinical trial data suggest that reduction of caloric intake in humans is generally associated with several health benefits, namely, reduction of abdominal fat mass, increased insulin sensitivity, reduced levels of pro-inflammatory cytokines, ROS and atherosclerotic lipids in the blood, and lowered body temperature and metabolic rate (Murphy et al. 2014; Redman and Ravussin 2011; Weiss and Fontana 2011).
The effect of CR on neurodegenerative diseases
Animal studies have provided evidence that dietary regimes such as CR affect not only the normal aging process, but also the onset and progression of different age-related diseases such as cancer and neurodegenerative brain diseases. A significant proportion of the rodents subjected to CR reached a very old age without any sign of disease (Omodei and Fontana 2011). Due to the rising global prevalence of neurodegenerative brain diseases such as AD and the lack of an effective therapy, these diseases have become a growing public health challenge with enormous economic and social implications (Gillette-Guyonnet et al. 2013). AD is clinically characterized by progressive deterioration of memory and cognitive functions, eventually leading to loss of autonomy requiring full-time medical care. In addition to severe neuronal loss, the neuropathological features of AD include the aggregation of Aβ in extracellular senile plaques and the formation of intraneuronal neurofibrillary tangles consisting of hyperphosphorylated tau protein (Selkoe 2011).
Animal studies
Numerous studies were conducted to investigate the effect of dietary regimens such as CR on cognition and on the pathological characteristics of AD in transgenic mouse models. Interestingly, young transgenic AD mice subjected to short-term CR displayed reduced accumulation of Aβ plaques and activation of astrocytes (Patel et al. 2005). Long-term reduced caloric intake provided similar results: attenuation of Aβ peptide accumulation and neuritic plaque deposition in the brain of Tg2576 and APP/PS1 mouse models of AD neuropathology after a CR dietary regimen (Mouton et al. 2009; Schafer et al. 2015; Wang et al. 2005). The Tg2576 mouse carries the human amyloid precursor protein (APP) double KM670/671NL mutation and the APP/PS1 model contains the human APP KM670/671NL mutations and presenilin 1 (PS1) L166P mutation. The observed beneficial effects in Tg2576 mice might be due to promotion of anti-amyloidogenic α-secretase activity (Wang et al. 2005) or by the reduced expression of presenilin enhancer 2 and PS1, components of the γ-secretase complex (Schafer et al. 2015).
To evaluate the dietary effects of CR on different AD pathological brain changes (brain atrophy, tau hyperphosphorylation, synaptic dysfunction, and cognitive deficit), a double-knockout mouse model of PS1 and PS2 was tested. A four-month CR regime improved memory, attenuated ventricle enlargement, caspase-3 activation and astrogliosis, and reduced tau hyperphosphorylation. Furthermore, CR was associated with up‐regulation of genes associated with neurogenesis and neuroplasticity and down-regulation of inflammatory genes (Wu et al. 2008). However, the molecular basis of these effects remains unclear.
The beneficial consequences of dietary regimes have also been investigated in other degenerative diseases, such as Parkinson’s disease (PD), Huntington disease (HD), amyotrophic lateral sclerosis (ALS), and models of tau deposition. Dietary restriction ameliorated the loss of dopaminergic neurons and deficits in motor function in a 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP)-induced rodent model of PD (Duan and Mattson 1999). The administration of 2-deoxy-d-glucose (2-DG), a non-metabolizable analogue of glucose to mimic the beneficial effects of CR, reduced damage of dopaminergic neurons in the substantia nigra and improved behavior (Duan and Mattson 1999). Surprisingly, the beneficial effects of dietary restrictions were not observed in a 6-hydroxydopamine (6-OHDA) model of PD. In this model, CR was not able to prevent nigrostriatal degeneration (Armentero et al. 2008). A primate model of PD reported that a CR diet can reduce the severity of neurochemical deficits and motor dysfunction. After MPTP treatment, CR monkeys exhibited significantly higher levels of locomotor activity compared with controls as well as higher levels of dopamine and dopamine metabolites in the striatal region of the brain (Maswood et al. 2004).
HD is caused by a polyglutamine repeat expansion in the huntingtin (HTT) protein. In a huntingtin mutant mice model (N171-82 Q), the formation of huntingtin inclusions and apoptotic protease activation, motor dysfunction, and metabolic glucose intolerance were delayed by a dietary restriction regime, which also slows down the disease progression and extended their lifespan (Duan et al. 2003). Further study showed that the overexpression of SIRT1 attenuates neurodegeneration and improves motor function in this N171-82Q HD mice model (Jiang et al. 2012). Transgenic mice overexpressing the mutant human superoxide dismutase 1 (SOD1) gene (G93A mice) are used to model amyotrophic lateral sclerosis (ALS). Transgenic ALS SOD1 G93A mice show reduced motor neuron survival in spinal cord and hind limb muscle, reduced lifespan, and increased oxidative stress. Dietary restriction transiently improves motor performance but accelerates the clinical onset of disease in G93A mice in comparison to the AL-fed counterparts (Hamadeh et al. 2005). The results suggest that CR diet is contraindicative for ALS (Hamadeh et al. 2005). A recent study however showed the increase in AMPK activity and the downregulation of heat shock protein (Hsp) 70 expression in comparison to mice fed AL. Due to the negative role of AMPK in motor neuron survival, the inhibition of AMPK may provide a therapeutic strategy for ALS (Zhao et al. 2015c). In a Tg4510 mouse model of tau deposition, CR partially rescued the memory deficits (Brownlow et al. 2014). The CR mice showed an improved short-term memory and performed better in contextual fear conditioning than AL-fed mice. Despite an apparent rescue of associative memory, CR had no consistent effects on pathological outcomes of the mouse model of tau deposition, since there were no changes in the activation of both astrocytes and microglia nor tau or phospho-tau levels (Brownlow et al. 2014).
Human clinical trials
Epidemiologic studies on AD reported an association of high caloric diets with increased risk for developing the disease. In addition, genetic studies suggested the importance of cholesterol metabolism and increased AD risk. Several clinical trials are ongoing on the effects of nutrition on neurodegenerative brain diseases (clinicaltrials.gov). The pathological process of AD and Aβ deposition in the brain proceeds silently for decades before cognitive failure becomes evident. Therefore, trials are focusing on individuals with mild cognitive impairment (MCI). These people present with mild cognitive defects as a transition state between healthy cognition and serious memory problems. A recent clinical trial evaluated the cognitive effect of intentional weight loss in obese elderly MCI patients. In these patients, CR-induced weight loss was associated with cognitive improvement (Horie et al. 2016).
Caloric restriction mimetics
Due to its ability to neutralize some of the detrimental effects of aging, CR is a promising approach to delay the onset or possibly also treat age-related neurodegenerative diseases such as AD. However, it is unlikely that humans will have the willpower to commit themselves to such a stringent diet. Therefore, research has focused on finding compounds named CRMs that mimic the biochemical and functional effects of CR. As proposed by Ingram et al., a CRM should mimic the metabolic, hormonal and physiological effects of CR without significantly reducing food intake. Besides, it should activate the stress response pathways observed in CR and provide protection against a variety of stressors. Finally, it should produce CR-like effects on longevity, reduction of age-related disease and maintenance of function (Ingram et al. 2006). Here, we will focus on two of the most studied CRM molecules, resveratrol and rapamycin, with special attention to AD.
Resveratrol
One of the important mechanisms by which CR is proposed to affect aging and extend lifespan is induction of the NAD-dependent HDAC, SIRT1, as discussed above. An in vitro screen for molecules that activate SIRT1 discovered resveratrol as being such a molecule (Howitz et al. 2003). Therefore, the potential beneficial effect of resveratrol in aging and age-related diseases has been extensively studied.
Resveratrol (Table 2) is found mainly in the skin of red grapes and therefore red wine (Nikolai et al. 2015) and has been reported to affect several pathways involved in aging and to promote longer life. In aged mice, resveratrol treatment starting at 12 months of age delayed functional decline by improving bone health, attenuating cataract formation and improving balance and motor coordination. Moreover, it improved vascular function and attenuated the expression of several inflammatory markers. Clearly, resveratrol slows the general age-related decline. However, the treatment did not increase lifespan (Pearson et al. 2008). In accordance with these data, a later study in which a two- to eightfold higher dose of resveratrol was used did not show a survival advantage of the treatment in mice (Miller et al. 2011). In old Fisher rats, resveratrol treatment improved spatial working memory, which indicates that resveratrol can reverse some of the deleterious effects of aging on cognitive performance. The spatial memory improvement correlated with the resveratrol concentration in the hippocampus (Joseph et al. 2008). Long-term treatment with resveratrol significantly improved cognitive abilities in elderly patients (Witte et al. 2014). The memory improvement correlated with an increase in the resting-state functional connectivity of the hippocampus in the resveratrol-treated group, which might reflect an improvement in the integrity and functionality of the hippocampus. This enhancement was also linked with a reduction in glycated hemoglobin A1c (HbA1c), a marker of glucose control, which indicates that resveratrol improves glucose metabolism. Since the hippocampus is thought to be vulnerable to disturbances in glucose supply, the authors suggest that resveratrol might, by this mechanism, have protective effects on neuronal functioning and consequently on cognition (Witte et al. 2014). Oxidative stress too plays a central role in aging (Beckman and Ames 1998). Tung et al. showed an increase in oxidative damage and a reduction in antioxidant activities in the liver of aging mice. Resveratrol treatment was able to attenuate this oxidative damage while the activity of several antioxidant enzymes increased. Importantly, resveratrol treatment was more effective in mature and aged mice, indicating that it is effective even when started at a later age (Tung et al. 2014). More recently, it was shown that resveratrol also increases the antioxidant activity in the heart and muscle of aged mice (Tung et al. 2015). Resveratrol has also been proposed as an anti-aging molecule due to its antioxidant, neuroprotective and anti-inflammatory activity in hippocampal astrocyte cultures from aged and adult Wistar rats (Bellaver et al. 2014). Moreover, resveratrol also seems to have an anti-apoptosis effect because pretreatment with resveratrol attenuated isoflurane-induced cognitive impairment, at least in part by its anti-neuroinflammation and anti-apoptosis effect (Li et al. 2014).
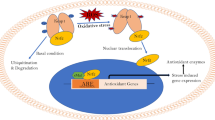
Next to its beneficial influence on the general age-related decline, resveratrol can ameliorate the pathology of age-related AD. Long-term treatment with resveratrol reduced the number and intensity of amyloid plaques in the hippocampus and the medial cortex of APP/PS1 mice, while also preventing memory loss (Porquet et al. 2014). Surprisingly, SIRT1 protein levels were decreased, which contradicts the postulated effect of resveratrol, namely, the activation of SIRT1. On the other hand, the levels of acetyl-p53, a substrate of SIRT1, were decreased. The explanation proposed by the authors is that resveratrol decreased the synthesis of SIRT1 but increased its activity (Porquet et al. 2014). In agreement with this suggestion, Park et al. have shown that resveratrol indirectly activates SIRT1 due to its effect on cyclic adenosine 3′,5′-monophosphate (cAMP) signaling. Resveratrol elevates cAMP levels by competitive inhibition of cAMP-degrading phosphodiesterases, leading to activation of the AMPK pathway, which ultimately increases the activity of SIRT1 (Park et al. 2012). In accordance with these data, Porquet et al. reported upregulation of AMPK (Porquet et al. 2014). Also in Tg19959 AD mice, containing a mutant human APP transgene, resveratrol diminished plaque formation in the brain without affecting SIRT1 levels (Karuppagounder et al. 2009).
Although it has been shown several times that resveratrol can reduce the amyloid burden (Karuppagounder et al. 2009; Porquet et al. 2014; Zhao et al. 2015a), the mechanism remains unclear. Some authors state that resveratrol treatment does not seem to affect APP processing because no changes in the levels of APP cleavage products were observed (Karuppagounder et al. 2009; Porquet et al. 2014). In contrast, the resveratrol trimer Miyabenol C reduced the amyloidogenic APPβ levels by directly inhibiting the activity of the APPβ generating β-site APP-cleaving enzyme 1 (BACE1), while the protein levels of BACE1 were unaltered. Although the levels of the non-amyloidogenic APPα increased, the activity of the APPα-generating α-secretase, a disintegrin and metalloproteinase 17 (ADAM17), also known as tumor necrosis factor alpha converting enzyme (TACE), did not increase (Hu et al. 2015). In agreement with these results, CR treatment had no effect on the ADAM17 activity in AD transgenic mice, but the activity of another α-secretase (ADAM10) surprisingly increased (Wang et al. 2005). It is also known that SIRT1 upregulates ADAM10 in neurons (Theendakara et al. 2013). Another possible reason for the reduction of Aβ levels by resveratrol was provided by Zhao et al. They showed that expression of the receptor of advanced glycation end products (RAGE) was reduced in AD rats, which suggests reduction of the influx of Aβ1-42 through the blood–brain barrier (BBB). Resveratrol even protected the integrity of the BBB by decreasing the expression of matrix metalloprotein-9 (MMP9) and increasing the expression of the tight junction protein Claudin-5 (Zhao et al. 2015a).
All these findings indicate that resveratrol might be useful in the treatment of age-related diseases, and particularly AD, because it can protect against amyloid plaque formation and to prevent cognitive decline. A recent clinical trial in individuals with mild to moderate AD showed that resveratrol is safe and well tolerated, and that it penetrates the blood–brain barrier (Turner et al. 2015). However, larger studies are needed to determine the potential effects of resveratrol treatment in humans (Patel et al. 2011; Subramanian et al. 2010) .
Rapamycin
As we discussed previously, the inhibition of the mTOR pathway by CR has been shown to extend lifespan in several model organisms (Johnson et al. 2013). The antibiotic rapamycin (Table 3) is an inhibitor of the protein kinase mTOR (Heitman et al. 1991) and has been used after organ transplantations presumably for its immunosuppressive effects (Nikolai et al. 2015). The National Institute on Aging Interventions Testing Program (ITP) showed that rapamycin treatment of male and female mice started at 20 months of age, which is roughly the equivalent of a 60-year-old person, extends median and maximum lifespan. This indicates that rapamycin has potential as an anti-aging intervention that can be initiated later in life (Harrison et al. 2009). Another ITP study in which the treatment was started at 9 months of age led to similar results, suggesting that starting treatment in later life is as effective as starting it early in adult life. The authors state that this might indicate that rapamycin rather modulates illnesses that characteristically occur in late-life than aging itself (Miller et al. 2011). A more recent study examined this question in male C57BL/6J mice. Rapamycin treatment started at the ages of 4, 13, and 20 months extended lifespan compared to control animals. To determine if rapamycin does slow aging itself, the researchers studied several aging phenotypes in the different cohorts of mice. For example, chronic rapamycin treatment enhanced learning and memory in the Morris Water Maze (MWM) cognitive test in aged mice, but this effect was also seen in young mice, which indicates that the improvement of memory by rapamycin in the MWM test occurred in an age-independent manner. In contrast, rapamycin treatment did not ameliorate age-related impairments such as grip strength reduction, cataract formation, vascular function, hepatic fibrosis or aging-associated changes in clinical chemistry parameters. However, rapamycin treatment significantly reduced the proportion of aged mice developing cancer and/or precancerous lesions (Neff et al. 2013). Importantly, also in the previously mentioned studies, cancer was the leading cause of death in both rapamycin-treated and control animals, but control animals died at a younger age (Harrison et al. 2009; Miller et al. 2011). Collectively, these results indicate that rapamycin does not affect aging itself but presumably exerts some aging-independent effects by which it modulates several aging traits. Likely, the observed beneficial effect on longevity is the consequence of delayed onset or slower progression of several lethal cancers (Neff et al. 2013). Also in female mice, rapamycin treatment dramatically decreased spontaneous carcinogenesis and prolonged lifespan only in the tumor-bearing cohort (Anisimov et al. 2011). As reviewed by Law et al., rapamycin has also been used in cancer treatment because of its ability to inhibit tumor growth, induce tumor cell apoptosis, and suppress tumor angiogenesis (Law 2005).
When mTOR is inhibited, autophagy and the ubiquitin proteasome system are stimulated, promoting degradation of proteins to release free amino acids that can be used to synthesize new proteins and to produce energy (Zhao et al. 2015b). Autophagy is essential for survival of neurons because these postmitotic cells are unable to dilute toxic cytoplasmic components by cell division (Peric and Annaert 2015). It has been shown that AD affects mainly the late steps in the autophagic pathway, namely the clearance of autophagic vacuoles (Boland et al. 2008). Consequently, stimulating autophagy by rapamycin can remove damaged, misfolded or aggregated proteins. This has led to the evaluation of rapamycin as a treatment for AD.
Rapamycin treatment reduced Aβ1-42 levels in the brain of 3xTg and hAPP AD mice. The former mice carry three human transgenes (APP, microtubule associated protein tau (MAPT) and PS1) that bear mutations associated with AD, while the latter contain a human mutant APP transgene. This reduction in Aβ1-42 levels appeared to be related to increased induction of autophagy by rapamycin. Importantly, also early learning and memory deficits were rescued, indicating the therapeutic potential of rapamycin (Caccamo et al. 2010; Spilman et al. 2010). Strikingly, even after the onset of AD-like impairments in the hAPP AD mice, rapamycin treatment improved memory to levels indistinguishable from those of wild-type controls (Lin et al. 2013). These results are at odds with the results of a study by Majumder et al., who showed that rapamycin treatment has no effect on cognitive deficits in 15-month-old 3xTg AD mice (Majumder et al. 2011). However, prophylactic treatment of 3xTg AD mice starting at the age of 2 months and continuing throughout their life did prevent the development of cognitive deficits. The authors suggest that once plaques and tangles are formed, induction of autophagy is not sufficient to rescue the AD-like pathology (Majumder et al. 2011).
Rapamycin can also exert a beneficial effect on tau, the other characteristic component of AD, since systemic rapamycin treatment protected against tau-induced neuronal loss, synaptotoxicity, reactive microgliosis, and astrogliosis in a mouse model of early-stage AD-type tau pathology (Siman et al. 2015). Rapamycin treatment also reduced soluble tau levels in the brain of 3xTg AD mice (Caccamo et al. 2010; Majumder et al. 2011). There seems to be a direct link between mTOR signaling and rapamycin because reducing mTOR signaling with rapamycin ameliorates tau pathology and the associated behavioral deficits in a mouse model overexpressing mutant human tau. These observations were linked to an increased autophagy induction (Caccamo et al. 2013).
Based on the studies described above, rapamycin seems to be a promising candidate for the treatment of AD. However, Zhang et al. showed that rapamycin treatment increased the Aβ levels in the brain of Tg2576 mouse model of AD. In accordance with this observation, the APPα levels decreased while the α-secretase ADAM10 was inhibited (Zhang et al. 2010). In human neural cells, inhibition of the mTOR pathway by rapamycin enhanced Aβ-induced cell death, leading to the suggestion that the mTOR pathway is important for the survival of cells exposed to Aβ (Lafay-Chebassier et al. 2006). Also, mTOR inhibition was shown to be detrimental to tau phosphorylation in rabbit organotypic hippocampal slices, while rapamycin significantly increased phosphorylated tau levels (Marwarha et al. 2010).
Further research is needed to understand the potential of rapamycin as a treatment for AD. The advantage of rapamycin is that is has already been approved by the Food and Drug Administration (FDA) for various uses in humans since 1999 (Richardson et al. 2015). However, its use could be limited by its immunosuppressive action, which may open the door to severe bacterial or viral diseases (Nikolai et al. 2015).
Other CRMs
Next to these two well-studied CRMs, researchers are also investigating the effects of numerous other CRMs. One method to discover such promising molecules is by using gene expression analysis. This approach led to the discovery of metformin as a CRM because this molecule was able to reproduce a CR-induced liver gene expression profile in mice (Dhahbi et al. 2005). Metformin is an insulin-sensitizing drug that is used as a first line oral therapy for the treatment of type 2 diabetes (Viollet et al. 2012). Patients suffering from type 2 diabetes, characterized by insulin resistance, have an increased risk to develop AD (Sridhar et al. 2015). Insulin is able to prevent Aβ oligomer formation in a dose-dependent matter (Lee et al. 2009) and it has also been shown that insulin has a beneficial effect on cognitive function (van der Heide et al. 2006). Therefore, insulin resistance which is often accompanied by peripheral hyperinsulinemia that consequently lowers insulin transport across the BBB and reduces insulin levels in the CNS, plays a detrimental role in the pathogenesis of AD (Craft et al. 2013). Chronic metformin exposure in male, middle-aged mice extended their lifespan and health span (Martin-Montalvo et al. 2013). In contrast, metformin supplementation starting from 6 months of age did not have any effect on the lifespan of male F344 rats whereas a 30 % CR diet significantly extended early lifespan in control animals (Smith et al. 2010a). Also in the case of AD, conflicting results were reported. A twelve-year Taiwanese population cohort study revealed that the use of metformin reduces the risk of developing AD in type 2 diabetes patients. Moreover, the risk was even further reduced when metformin treatment was combined with another anti-hyperglycemic agent sulfonylurea (Hsu et al. 2011). In contrast, a United Kingdom case–control study suggested a slightly higher risk of developing AD in long-term metformin-treated type 2 diabetes patients, whereas sulfonylurea monotherapy did not alter the risk (Imfeld et al. 2012). Also other research provided evidence for a detrimental effect of metformin related to AD. It was shown that metformin increases levels of both extracellular and intracellular Aβ in human APP expressing primary neurons and neuroblastoma cells. These findings were linked to an increased protein level and enzymatic activity of BACE1 (Chen et al. 2009a, b). Further research will be needed to clarify the potential of metformin in slowing down aging and altering the development or progression of AD.
Clearly, also the possible underlying mechanisms of CR provide potential to develop CRM. As mentioned earlier, autophagy has been proposed as an important CR-induced effect that contributes to its beneficial activities. Next to rapamycin, numerous other autophagy inducers have been investigated. They are comprehensively reviewed elsewhere (Marino et al. 2014). SIRT1 activation is another proposed mechanism to explain the CR effects. SIRT1 activates PGC-1α (Rodgers et al. 2005) which consequently induces the proliferation of highly efficient mitochondria that produce less ROS (Lopez-Lluch et al. 2006). Another study showed that a CR regimen induces endothelial nitric oxide synthase (eNOS), accompanied by an increased expression of SIRT1 and mitochondrial biogenesis. Importantly, the induction of SIRT1 expression was much smaller and the mitochondrial biogenesis was compromised in eNOS-deficient mice on a CR diet. These findings point to an important role of nitric oxide (NO), generated by eNOS, in response to CR (Nisoli et al. 2005). Therefore, NO generators including artemisinin have been proposed as CRM. Artemisinin treatment induces eNOS expression and thereby increases NO levels, leading to mitochondrial biogenesis. Antioxidant enzymes are activated and this leads to the scavenging and consequently lowering of ROS levels. Moreover, this is associated with lower expression of DNA repair genes, suggesting less DNA damage, and telomeres are protected against detrimental, age-related shortening (Wang et al. 2015). Additional research will be needed to shed further light on these mechanisms.
More recently, bioinformatics has proven to be a valuable tool to enlarge the existing pool of CRMs (Calvert et al. 2016). A recent study looked for similarities between a CR-induced gene expression profile in rat cells and known drug-induced expression profiles in human cells. Several drugs, with rapamycin being one of the top hits, were selected and tested in C. elegans. One of the most remarkable molecules that prolonged lifespan and improved health span in a CR-like manner but through the induction of a differential gene expression profile compared to rapamycin, was allantoin (Calvert et al. 2016). Additional research is needed to determine the potential of this commonly used anti-aging skin care ingredient.
Also population data can reveal new CRMs. These population data are of great importance because unfortunately, long-term outcomes of a CR diet, such as mortality and lifespan, are often missing in human studies. Therefore, the Okinawans, a Japanese population that has the world’s longest life expectancy and a low incidence of age-related diseases, is of special interest to CR researchers. Their good health is epidemiologically and demographically linked to their consumption of a diet that is mildly restricted in calories (10–15 %) as well as their intake of foods with CRM properties, such as turmeric (Curcuma longa) (Willcox and Willcox 2014). The main curcuminoid found in the Curcuma longa family is curcumin, which is responsible for the yellow color of turmeric (Chin et al. 2013). It has been shown that curcumin increases lifespan in several model organisms (Shen et al. 2013). Moreover, several reports describe the beneficial effects of curcumin on AD development. Curcumin treatment can reduce amyloid deposits in the brain of APP/PS1 and Tg2576 AD mice (Garcia-Alloza et al. 2007; Lim et al. 2001; Shytle et al. 2012) and reduce tau phosphorylation in Tg2576 and 3xTg AD mice (Ma et al. 2009; Shytle et al. 2012).
Concluding remarks
There is mounting evidence that dietary regimes such as CR have an important influence on lifespan and health span. Considerable research aimed at fully understanding the processes and mechanisms behind aging and indicates that the mechanisms are complex and multifactorial, but they are still poorly understood (Sohal and Forster 2014). Because populations are aging and the prevalence of neurodegenerative diseases is increasing, a better understanding of how changes in diet and dietary regimes can affect aging and possibly neurodegeneration could identify genes and pathways relevant for human preventive or therapeutic applications, and that would be of major social and economic significance (Wood et al. 2015). Nevertheless, some considerations need to be taken into account. In most of the research, CR animals are compared to AL-fed counterparts, the latter of which might not represent the best control group because they have the tendency to overeat and become overweight. They lack mental and physical activity, whereas CR rodents tend to search actively for food in their environment, which provides mental and physical activity. A second concern is compliance with a CR diet in humans, which is very difficult to achieve over a long time (Mattson et al. 2003). Moreover, malnutrition is a common problem in older individuals, and a balanced diet is extremely important (Murphy et al. 2014; Pasinetti and Eberstein 2008). Due to the demanding nature of a CR diet, CRM drugs are an attractive alternative. These compounds mimic the biochemical and functional effects of CR without the need to reduce energy intake. In this review, we especially focused on two molecules that might delay aging and treat AD, namely, resveratrol and rapamycin. However, it is still unclear how these molecules exert their effects, and whether their effects are only beneficial is controversial. Therefore, large clinical studies are needed to determine their safety and efficacy in humans. Additionally, the bioavailability of these molecules has to be determined and possibly improved. In summary, these CRM provide interesting opportunities for novel preventive and therapeutic strategies. Furthermore, as CR affects several pathways through complex mechanisms, it is logical to assume that several CRMs have to be used simultaneously in order to mimic the complex effects of CR. However, there still is a long way to go before they can be implemented in our lifestyle to delay aging and prevent or slow down the progression of age-related diseases.
References
Adams MM, Shi L, Linville MC, Forbes ME, Long AB, Bennett C, Newton IG, Carter CS, Sonntag WE, Riddle DR, Brunso-Bechtold JK (2008) Caloric restriction and age affect synaptic proteins in hippocampal CA3 and spatial learning ability. Exp Neurol 211(1):141–149
Albert V, Hall MN (2015) mTOR signaling in cellular and organismal energetics. Curr Opin Cell Biol 33:55–66
Anisimov VN, Bartke A (2013) The key role of growth hormone-insulin-IGF-1 signaling in aging and cancer. Crit Rev Oncol Hematol 87:201–223
Anisimov VN, Zabezhinski MA, Popovich IG, Piskunova TS, Semenchenko AV, Tyndyk ML, Yurova MN, Rosenfeld SV, Blagosklonny MV (2011) Rapamycin increases lifespan and inhibits spontaneous tumorigenesis in inbred female mice. Cell Cycle 10:4230–4236
Armentero MT, Levandis G, Bramanti P, Nappi G, Blandini F (2008) Dietary restriction does not prevent nigrostriatal degeneration in the 6-hydroxydopamine model of Parkinson’s disease. Exp Neurol 212:548–551
Barger JL, Kayo T, Vann JM, Arias EB, Wang J, Hacker TA, Wang Y, Raederstorff D, Morrow JD, Leeuwenburgh C, Allison DB, Saupe KW, Cartee GD, Weindruch R, Prolla TA (2008) A low dose of dietary resveratrol partially mimics caloric restriction and retards aging parameters in mice. PLoS One 3:e2264
Beckman KB, Ames BN (1998) The free radical theory of aging matures. Physiol Rev 78:547–581
Bekinschtein P, Oomen CA, Saksida LM, Bussey TJ (2011) Effects of environmental enrichment and voluntary exercise on neurogenesis, learning and memory, and pattern separation: BDNF as a critical variable? Semin Cell Dev Biol 22:536–542
Bellaver B, Souza DG, Souza DO, Quincozes-Santos A (2014) Resveratrol increases antioxidant defenses and decreases proinflammatory cytokines in hippocampal astrocyte cultures from newborn, adult and aged Wistar rats. Toxicol In Vitro 28:479–484
Boland B, Kumar A, Lee S, Platt FM, Wegiel J, Yu WH, Nixon RA (2008) Autophagy induction and autophagosome clearance in neurons: relationship to autophagic pathology in Alzheimer’s disease. J Neurosci 28:6926–6937
Bordone L, Cohen D, Robinson A, Motta MC, van Veen E, Czopik A, Steele AD, Crowe H, Marmor S, Luo J, Gu W, Guarente L (2007) SIRT1 transgenic mice show phenotypes resembling calorie restriction. Aging Cell 6:759–767
Brachmann CB, Sherman JM, Devine SE, Cameron EE, Pillus L, Boeke JD (1995) The SIR2 gene family, conserved from bacteria to humans, functions in silencing, cell cycle progression, and chromosome stability. Genes Dev 9:2888–2902
Brownlow ML, Joly-Amado A, Azam S, Elza M, Selenica ML, Pappas C, Small B, Engelman R, Gordon MN, Morgan D (2014) Partial rescue of memory deficits induced by calorie restriction in a mouse model of tau deposition. Behav Brain Res 271:79–88
Caccamo A, Majumder S, Richardson A, Strong R, Oddo S (2010) Molecular interplay between mammalian target of rapamycin (mTOR), amyloid-beta, and Tau: effects on cognitive impairments. J Biol Chem 285:13107–13120
Caccamo A, Magri A, Medina DX, Wisely EV, Lopez-Aranda MF, Silva AJ, Oddo S (2013) mTOR regulates tau phosphorylation and degradation: implications for Alzheimer’s disease and other tauopathies. Aging Cell 12:370–380
Calvert S, Tacutu R, Sharifi S, Teixeira R, Ghosh P, de Magalhaes JP (2016) A network pharmacology approach reveals new candidate caloric restriction mimetics in C. elegans. Aging Cell 15:256–266
Canto C, Auwerx J (2011) Calorie restriction: is AMPK a key sensor and effector? Physiology (Bethesda) 26:214–224
Cava E, Fontana L (2013) Will calorie restriction work in humans? Aging (Albany NY) 5:507–514
Chan MF, Liang G, Jones PA (2000) Relationship between transcription and DNA methylation. Curr Top Microbiol Immunol 249:75–86
Chen C, Liu Y, Liu Y, Zheng P (2009a) mTOR regulation and therapeutic rejuvenation of aging hematopoietic stem cells. Sci Signal 2:75
Chen Y, Zhou K, Wang R, Liu Y, Kwak YD, Ma T, Thompson RC, Zhao Y, Smith L, Gasparini L, Luo Z, Xu H, Liao FF (2009b) Antidiabetic drug metformin (GlucophageR) increases biogenesis of Alzheimer’s amyloid peptides via up-regulating BACE1 transcription. Proc Natl Acad Sci USA 106:3907–3912
Chin D, Huebbe P, Pallauf K, Rimbach G (2013) Neuroprotective properties of curcumin in Alzheimer’s disease–merits and limitations. Curr Med Chem 20:3955–3985
Cohen HY, Miller C, Bitterman KJ, Wall NR, Hekking B, Kessler B, Howitz KT, Gorospe M, de Cabo R, Sinclair DA (2004) Calorie restriction promotes mammalian cell survival by inducing the SIRT1 deacetylase. Science 305:390–392
Colman RJ, Beasley TM, Allison DB, Weindruch R (2008) Attenuation of sarcopenia by dietary restriction in rhesus monkeys. J Gerontol A Biol Sci Med Sci 63:556–559
Colman RJ, Anderson RM, Johnson SC, Kastman EK, Kosmatka KJ, Beasley TM, Allison DB, Cruzen C, Simmons HA, Kemnitz JW, Weindruch R (2009) Caloric restriction delays disease onset and mortality in rhesus monkeys. Science 325:201–204
Colman RJ, Beasley TM, Kemnitz JW, Johnson SC, Weindruch R, Anderson RM (2014) Caloric restriction reduces age-related and all-cause mortality in rhesus monkeys. Nat Commun 5:3557
Craft S, Cholerton B, Baker LD (2013) Insulin and Alzheimer’s disease: untangling the web. J Alzheimers Dis 33(Suppl 1):S263–S275
Cuervo AM (2008) Autophagy and aging: keeping that old broom working. Trends Genet 24:604–612
da Luz PL, Tanaka L, Brum PC, Dourado PM, Favarato D, Krieger JE, Laurindo FR (2012) Red wine and equivalent oral pharmacological doses of resveratrol delay vascular aging but do not extend life span in rats. Atherosclerosis 224:136–142
de Ruijter AJ, van Gennip AH, Caron HN, Kemp S, van Kuilenburg AB (2003) Histone deacetylases (HDACs): characterization of the classical HDAC family. Biochem J 370:737–749
Dhahbi JM, Mote PL, Fahy GM, Spindler SR (2005) Identification of potential caloric restriction mimetics by microarray profiling. Physiol Genom 23:343–350
Duan W, Mattson MP (1999) Dietary restriction and 2-deoxyglucose administration improve behavioral outcome and reduce degeneration of dopaminergic neurons in models of Parkinson’s disease. J Neurosci Res 57:195–206
Duan W, Lee J, Guo Z, Mattson MP (2001) Dietary restriction stimulates BDNF production in the brain and thereby protects neurons against excitotoxic injury. J Mol Neurosci 16:1–12
Duan W, Guo Z, Jiang H, Ware M, Li XJ, Mattson MP (2003) Dietary restriction normalizes glucose metabolism and BDNF levels, slows disease progression, and increases survival in huntingtin mutant mice. Proc Natl Acad Sci USA 100:2911–2916
Elshorbagy AK, Valdivia-Garcia M, Mattocks DA, Plummer JD, Orentreich DS, Orentreich N, Refsum H, Perrone CE (2013) Effect of taurine and N-acetylcysteine on methionine restriction-mediated adiposity resistance. Metabolism 62:509–517
Feng X, Liang N, Zhu D, Gao Q, Peng L, Dong H, Yue Q, Liu H, Bao L, Zhang J, Hao J, Gao Y, Yu X, Sun J (2013) Resveratrol inhibits beta-amyloid-induced neuronal apoptosis through regulation of SIRT1-ROCK1 signaling pathway. PLoS One 8:e59888
Finkel T (2015) The metabolic regulation of aging. Nat Med 21:1416–1423
Flynn JM, O’Leary MN, Zambataro CA, Academia EC, Presley MP, Garrett BJ, Zykovich A, Mooney SD, Strong R, Rosen CJ, Kapahi P, Nelson MD, Kennedy BK, Melov S (2013) Late-life rapamycin treatment reverses age-related heart dysfunction. Aging Cell 12:851–862
Fok WC, Chen Y, Bokov A, Zhang Y, Salmon AB, Diaz V, Javors M, Wood WH 3rd, Zhang Y, Becker KG, Perez VI, Richardson A (2014) Mice fed rapamycin have an increase in lifespan associated with major changes in the liver transcriptome. PLoS One 9:e83988
Fontana L, Partridge L, Longo VD (2010) Extending healthy life span–from yeast to humans. Science 328:321–326
Garcia-Alloza M, Borrelli LA, Rozkalne A, Hyman BT, Bacskai BJ (2007) Curcumin labels amyloid pathology in vivo, disrupts existing plaques, and partially restores distorted neurites in an Alzheimer mouse model. J Neurochem 102:1095–1104
Gillette-Guyonnet S, Secher M, Vellas B (2013) Nutrition and neurodegeneration: epidemiological evidence and challenges for future research. Br J Clin Pharmacol 75:738–755
Goldberg EL, Romero-Aleshire MJ, Renkema KR, Ventevogel MS, Chew WM, Uhrlaub JL, Smithey MJ, Limesand KH, Sempowski GD, Brooks HL, Nikolich-Zugich J (2015) Lifespan-extending caloric restriction or mTOR inhibition impair adaptive immunity of old mice by distinct mechanisms. Aging Cell 14:130–138
Gorlé N, Van Cauwenberghe C, Libert C, Vandenbroucke RE (2016) The effect of aging on brain barriers and the consequences for Alzheimer's disease development. Mamm Genome. doi:10.1007/s00335-016-9637-8
Goto S, Takahashi R, Radak Z, Sharma R (2007) Beneficial biochemical outcomes of late-onset dietary restriction in rodents. Ann N Y Acad Sci 1100:431–441
Greer EL, Dowlatshahi D, Banko MR, Villen J, Hoang K, Blanchard D, Gygi SP, Brunet A (2007) An AMPK-FOXO pathway mediates longevity induced by a novel method of dietary restriction in C. elegans. Curr Biol 17:1646–1656
Guarente L, Picard F (2005) Calorie restriction–the SIR2 connection. Cell 120:473–482
Haigis MC, Guarente LP (2006) Mammalian sirtuins–emerging roles in physiology, aging, and calorie restriction. Genes Dev 20:2913–2921
Halagappa VK, Guo Z, Pearson M, Matsuoka Y, Cutler RG, Laferla FM, Mattson MP (2007) Intermittent fasting and caloric restriction ameliorate age-related behavioral deficits in the triple-transgenic mouse model of Alzheimer’s disease. Neurobiol Dis 26:212–220
Halloran J, Hussong SA, Burbank R, Podlutskaya N, Fischer KE, Sloane LB, Austad SN, Strong R, Richardson A, Hart MJ, Galvan V (2012) Chronic inhibition of mammalian target of rapamycin by rapamycin modulates cognitive and non-cognitive components of behavior throughout lifespan in mice. Neuroscience 223:102–113
Hamadeh MJ, Rodriguez MC, Kaczor JJ, Tarnopolsky MA (2005) Caloric restriction transiently improves motor performance but hastens clinical onset of disease in the Cu/Zn-superoxide dismutase mutant G93A mouse. Muscle Nerve 31:214–220
Harrison DE, Strong R, Sharp ZD, Nelson JF, Astle CM, Flurkey K, Nadon NL, Wilkinson JE, Frenkel K, Carter CS, Pahor M, Javors MA, Fernandez E, Miller RA (2009) Rapamycin fed late in life extends lifespan in genetically heterogeneous mice. Nature 460:392–395
Heitman J, Movva NR, Hall MN (1991) Targets for cell cycle arrest by the immunosuppressant rapamycin in yeast. Science 253:905–909
Hine C, Mitchell JR (2015) Calorie restriction and methionine restriction in control of endogenous hydrogen sulfide production by the transsulfuration pathway. Exp Gerontol 68:26–32
Hine C, Harputlugil E, Zhang Y, Ruckenstuhl C, Lee BC, Brace L, Longchamp A, Trevino-Villarreal JH, Mejia P, Ozaki CK, Wang R, Gladyshev VN, Madeo F, Mair WB, Mitchell JR (2015) Endogenous hydrogen sulfide production is essential for dietary restriction benefits. Cell 160:132–144
Holloszy JO, Fontana L (2007) Caloric restriction in humans. Exp Gerontol 42:709–712
Horie NC, Serrao VT, Simon SS, Polo Gascon MR, Xavier Dos Santos A, Zambone MA, Bigio de Freitas MM, Cunha-Neto E, Marques EL, Halpern A, Edna de Melo M, Mancini MC, Cercato C (2016) Cognitive effects of intentional weight loss in elderly obese individuals with mild cognitive impairment. J Clin Endocrinol Metab 101:1104–1112
Howitz KT, Bitterman KJ, Cohen HY, Lamming DW, Lavu S, Wood JG, Zipkin RE, Chung P, Kisielewski A, Zhang LL, Scherer B, Sinclair DA (2003) Small molecule activators of sirtuins extend Saccharomyces cerevisiae lifespan. Nature 425:191–196
Hsu CC, Wahlqvist ML, Lee MS, Tsai HN (2011) Incidence of dementia is increased in type 2 diabetes and reduced by the use of sulfonylureas and metformin. J Alzheimers Dis 24:485–493
Hu J, Lin T, Gao Y, Xu J, Jiang C, Wang G, Bu G, Xu H, Chen H, Zhang YW (2015) The resveratrol trimer miyabenol C inhibits beta-secretase activity and beta-amyloid generation. PLoS One 10:e0115973
Huang TC, Lu KT, Wo YY, Wu YJ, Yang YL (2011) Resveratrol protects rats from Abeta-induced neurotoxicity by the reduction of iNOS expression and lipid peroxidation. PLoS One 6:e29102
Hulbert AJ (2005) On the importance of fatty acid composition of membranes for aging. J Theor Biol 234:277–288
Imfeld P, Bodmer M, Jick SS, Meier CR (2012) Metformin, other antidiabetic drugs, and risk of Alzheimer’s disease: a population-based case-control study. J Am Geriatr Soc 60:916–921
Ingram DK, Weindruch R, Spangler EL, Freeman JR, Walford RL (1987) Dietary restriction benefits learning and motor performance of aged mice. J Gerontol 42:78–81
Ingram DK, Zhu M, Mamczarz J, Zou S, Lane MA, Roth GS, deCabo R (2006) Calorie restriction mimetics: an emerging research field. Aging Cell 5:97–108
Inoki K, Zhu T, Guan KL (2003) TSC2 mediates cellular energy response to control cell growth and survival. Cell 115:577–590
Jiang M, Wang J, Fu J, Du L, Jeong H, West T, Xiang L, Peng Q, Hou Z, Cai H, Seredenina T, Arbez N, Zhu S, Sommers K, Qian J, Zhang J, Mori S, Yang XW, Tamashiro KL, Aja S, Moran TH, Luthi-Carter R, Martin B, Maudsley S, Mattson MP, Cichewicz RH, Ross CA, Holtzman DM, Krainc D, Duan W (2012) Neuroprotective role of Sirt1 in mammalian models of Huntington’s disease through activation of multiple Sirt1 targets. Nat Med 18:153–158
Johnson SC, Rabinovitch PS, Kaeberlein M (2013) mTOR is a key modulator of ageing and age-related disease. Nature 493:338–345
Joseph JA, Fisher DR, Cheng V, Rimando AM, Shukitt-Hale B (2008) Cellular and behavioral effects of stilbene resveratrol analogues: implications for reducing the deleterious effects of aging. J Agric Food Chem 56:10544–10551
Juhasz G, Erdi B, Sass M, Neufeld TP (2007) Atg7-dependent autophagy promotes neuronal health, stress tolerance, and longevity but is dispensable for metamorphosis in Drosophila. Genes Dev 21:3061–3066
Kadonaga JT (1998) Eukaryotic transcription: an interlaced network of transcription factors and chromatin-modifying machines. Cell 92:307–313
Kanaya T, Kyo S, Takakura M, Ito H, Namiki M, Inoue M (1998) hTERT is a critical determinant of telomerase activity in renal-cell carcinoma. Int J Cancer 78:539–543
Kapahi P, Zid BM, Harper T, Koslover D, Sapin V, Benzer S (2004) Regulation of lifespan in Drosophila by modulation of genes in the TOR signaling pathway. Curr Biol 14:885–890
Karuppagounder SS, Pinto JT, Xu H, Chen HL, Beal MF, Gibson GE (2009) Dietary supplementation with resveratrol reduces plaque pathology in a transgenic model of Alzheimer’s disease. Neurochem Int 54:111–118
Kaur M, Sharma S, Kaur G (2008) Age-related impairments in neuronal plasticity markers and astrocytic GFAP and their reversal by late-onset short term dietary restriction. Biogerontology 9:441–454
Kenyon C, Chang J, Gensch E, Rudner A, Tabtiang R (1993) A C. elegans mutant that lives twice as long as wild type. Nature 366:461–464
Kim D, Nguyen MD, Dobbin MM, Fischer A, Sananbenesi F, Rodgers JT, Delalle I, Baur JA, Sui G, Armour SM, Puigserver P, Sinclair DA, Tsai LH (2007) SIRT1 deacetylase protects against neurodegeneration in models for Alzheimer’s disease and amyotrophic lateral sclerosis. EMBO J 26:3169–3179
Kimura KD, Tissenbaum HA, Liu Y, Ruvkun G (1997) daf-2, an insulin receptor-like gene that regulates longevity and diapause in Caenorhabditis elegans. Science 277:942–946
Ladiwala AR, Lin JC, Bale SS, Marcelino-Cruz AM, Bhattacharya M, Dordick JS, Tessier PM (2010) Resveratrol selectively remodels soluble oligomers and fibrils of amyloid Abeta into off-pathway conformers. J Biol Chem 285:24228–24237
Lafay-Chebassier C, Perault-Pochat MC, Page G, Rioux Bilan A, Damjanac M, Pain S, Houeto JL, Gil R, Hugon J (2006) The immunosuppressant rapamycin exacerbates neurotoxicity of Abeta peptide. J Neurosci Res 84:1323–1334
Lagouge M, Argmann C, Gerhart-Hines Z, Meziane H, Lerin C, Daussin F, Messadeq N, Milne J, Lambert P, Elliott P, Geny B, Laakso M, Puigserver P, Auwerx J (2006) Resveratrol improves mitochondrial function and protects against metabolic disease by activating SIRT1 and PGC-1alpha. Cell 127:1109–1122
Langley E, Pearson M, Faretta M, Bauer UM, Frye RA, Minucci S, Pelicci PG, Kouzarides T (2002) Human SIR2 deacetylates p53 and antagonizes PML/p53-induced cellular senescence. EMBO J 21:2383–2396
Law BK (2005) Rapamycin: an anti-cancer immunosuppressant? Crit Rev Oncol Hematol 56:47–60
Lee J, Duan W, Long JM, Ingram DK, Mattson MP (2000) Dietary restriction increases the number of newly generated neural cells, and induces BDNF expression, in the dentate gyrus of rats. J Mol Neurosci 15:99–108
Lee J, Duan W, Mattson MP (2002) Evidence that brain-derived neurotrophic factor is required for basal neurogenesis and mediates, in part, the enhancement of neurogenesis by dietary restriction in the hippocampus of adult mice. J Neurochem 82:1367–1375
Lee CC, Kuo YM, Huang CC, Hsu KS (2009) Insulin rescues amyloid beta-induced impairment of hippocampal long-term potentiation. Neurobiol Aging 30:377–387
Leibiger IB, Berggren PO (2006) Sirt1: a metabolic master switch that modulates lifespan. Nat Med 12:34–36 discussion 36
Li E, Beard C, Jaenisch R (1993) Role for DNA methylation in genomic imprinting. Nature 366:362–365
Li Y, Liu L, Tollefsbol TO (2010) Glucose restriction can extend normal cell lifespan and impair precancerous cell growth through epigenetic control of hTERT and p16 expression. FASEB J 24:1442–1453
Li Y, Daniel M, Tollefsbol TO (2011) Epigenetic regulation of caloric restriction in aging. BMC Med 9:98
Li XM, Zhou MT, Wang XM, Ji MH, Zhou ZQ, Yang JJ (2014) Resveratrol pretreatment attenuates the isoflurane-induced cognitive impairment through its anti-inflammation and -apoptosis actions in aged mice. J Mol Neurosci 52:286–293
Lim GP, Chu T, Yang F, Beech W, Frautschy SA, Cole GM (2001) The curry spice curcumin reduces oxidative damage and amyloid pathology in an Alzheimer transgenic mouse. J Neurosci 21:8370–8377
Lin SJ, Defossez PA, Guarente L (2000) Requirement of NAD and SIR2 for life-span extension by calorie restriction in Saccharomyces cerevisiae. Science 289:2126–2128
Lin AL, Zheng W, Halloran JJ, Burbank RR, Hussong SA, Hart MJ, Javors M, Shih YY, Muir E, Solano Fonseca R, Strong R, Richardson AG, Lechleiter JD, Fox PT, Galvan V (2013) Chronic rapamycin restores brain vascular integrity and function through NO synthase activation and improves memory in symptomatic mice modeling Alzheimer’s disease. J Cereb Blood Flow Metab 33:1412–1421
Lipinski MM, Zheng B, Lu T, Yan Z, Py BF, Ng A, Xavier RJ, Li C, Yankner BA, Scherzer CR, Yuan J (2010) Genome-wide analysis reveals mechanisms modulating autophagy in normal brain aging and in Alzheimer’s disease. Proc Natl Acad Sci USA 107:14164–14169
Lopez-Lluch G, Hunt N, Jones B, Zhu M, Jamieson H, Hilmer S, Cascajo MV, Allard J, Ingram DK, Navas P, de Cabo R (2006) Calorie restriction induces mitochondrial biogenesis and bioenergetic efficiency. Proc Natl Acad Sci USA 103:1768–1773
Lord J, Cruchaga C (2014) The epigenetic landscape of Alzheimer’s disease. Nat Neurosci 17:1138–1140
Luo J, Nikolaev AY, Imai S, Chen D, Su F, Shiloh A, Guarente L, Gu W (2001) Negative control of p53 by Sir2alpha promotes cell survival under stress. Cell 107:137–148
Ma QL, Yang F, Rosario ER, Ubeda OJ, Beech W, Gant DJ, Chen PP, Hudspeth B, Chen C, Zhao Y, Vinters HV, Frautschy SA, Cole GM (2009) Beta-amyloid oligomers induce phosphorylation of tau and inactivation of insulin receptor substrate via c-Jun N-terminal kinase signaling: suppression by omega-3 fatty acids and curcumin. J Neurosci 29:9078–9089
Mair W, Morantte I, Rodrigues AP, Manning G, Montminy M, Shaw RJ, Dillin A (2011) Lifespan extension induced by AMPK and calcineurin is mediated by CRTC-1 and CREB. Nature 470:404–408
Majumder S, Richardson A, Strong R, Oddo S (2011) Inducing autophagy by rapamycin before, but not after, the formation of plaques and tangles ameliorates cognitive deficits. PLoS One 6:e25416
Marambaud P, Zhao H, Davies P (2005) Resveratrol promotes clearance of Alzheimer’s disease amyloid-beta peptides. J Biol Chem 280:37377–37382
Marino G, Pietrocola F, Madeo F, Kroemer G (2014) Caloric restriction mimetics: natural/physiological pharmacological autophagy inducers. Autophagy 10:1879–1882
Martin B, Mattson MP, Maudsley S (2006) Caloric restriction and intermittent fasting: two potential diets for successful brain aging. Ageing Res Rev 5:332–353
Martin CK, Anton SD, Han H, York-Crowe E, Redman LM, Ravussin E, Williamson DA (2007) Examination of cognitive function during six months of calorie restriction: results of a randomized controlled trial. Rejuvenation Res 10:179–190
Martin-Montalvo A, Mercken EM, Mitchell SJ, Palacios HH, Mote PL, Scheibye-Knudsen M, Gomes AP, Ward TM, Minor RK, Blouin MJ, Schwab M, Pollak M, Zhang Y, Yu Y, Becker KG, Bohr VA, Ingram DK, Sinclair DA, Wolf NS, Spindler SR, Bernier M, de Cabo R (2013) Metformin improves healthspan and lifespan in mice. Nat Commun 4:2192
Martins R, Lithgow GJ, Link W (2016) Long live FOXO: unraveling the role of FOXO proteins in aging and longevity. Aging Cell 15:196–207
Marwarha G, Dasari B, Prabhakara JP, Schommer J, Ghribi O (2010) beta-Amyloid regulates leptin expression and tau phosphorylation through the mTORC1 signaling pathway. J Neurochem 115:373–384
Masoro EJ (2005) Overview of caloric restriction and ageing. Mech Ageing Dev 126:913–922
Maswood N, Young J, Tilmont E, Zhang Z, Gash DM, Gerhardt GA, Grondin R, Roth GS, Mattison J, Lane MA, Carson RE, Cohen RM, Mouton PR, Quigley C, Mattson MP, Ingram DK (2004) Caloric restriction increases neurotrophic factor levels and attenuates neurochemical and behavioral deficits in a primate model of Parkinson’s disease. Proc Natl Acad Sci USA 101:18171–18176
Mattson MP (2002) Brain evolution and lifespan regulation: conservation of signal transduction pathways that regulate energy metabolism. Mech Ageing Dev 123:947–953
Mattson MP (2012) Energy intake and exercise as determinants of brain health and vulnerability to injury and disease. Cell Metab 16:706–722
Mattson MP (2015) Lifelong brain health is a lifelong challenge: from evolutionary principles to empirical evidence. Ageing Res Rev 20:37–45
Mattson MP, Duan W, Guo Z (2003) Meal size and frequency affect neuronal plasticity and vulnerability to disease: cellular and molecular mechanisms. J Neurochem 84:417–431
McCay CM, Crowell MF, Maynard LA (1989) The effect of retarded growth upon the length of life span and upon the ultimate body size. 1935. Nutrition 5:155–171 discussion 172
Means LW, Higgins JL, Fernandez TJ (1993) Mid-life onset of dietary restriction extends life and prolongs cognitive functioning. Physiol Behav 54:503–508
Meyerson M, Counter CM, Eaton EN, Ellisen LW, Steiner P, Caddle SD, Ziaugra L, Beijersbergen RL, Davidoff MJ, Liu Q, Bacchetti S, Haber DA, Weinberg RA (1997) hEST2, the putative human telomerase catalytic subunit gene, is up-regulated in tumor cells and during immortalization. Cell 90:785–795
Miller RA, Harrison DE, Astle CM, Baur JA, Boyd AR, de Cabo R, Fernandez E, Flurkey K, Javors MA, Nelson JF, Orihuela CJ, Pletcher S, Sharp ZD, Sinclair D, Starnes JW, Wilkinson JE, Nadon NL, Strong R (2011) Rapamycin, but not resveratrol or simvastatin, extends life span of genetically heterogeneous mice. J Gerontol A Biol Sci Med Sci 66:191–201
Miller RA, Harrison DE, Astle CM, Fernandez E, Flurkey K, Han M, Javors MA, Li X, Nadon NL, Nelson JF, Pletcher S, Salmon AB, Sharp ZD, Van Roekel S, Winkleman L, Strong R (2014) Rapamycin-mediated lifespan increase in mice is dose and sex dependent and metabolically distinct from dietary restriction. Aging Cell 13:468–477
Mizushima N, Komatsu M (2011) Autophagy: renovation of cells and tissues. Cell 147:728–741
Mladenovic Djordjevic A, Perovic M, Tesic V, Tanic N, Rakic L, Ruzdijic S, Kanazir S (2010) Long-term dietary restriction modulates the level of presynaptic proteins in the cortex and hippocampus of the aging rat. Neurochem Int 56:250–255
Mouton PR, Chachich ME, Quigley C, Spangler E, Ingram DK (2009) Caloric restriction attenuates amyloid deposition in middle-aged dtg APP/PS1 mice. Neurosci Lett 464:184–187
Munoz-Najar U, Sedivy JM (2011) Epigenetic control of aging. Antioxid Redox Signal 14:241–259
Murphy CT, McCarroll SA, Bargmann CI, Fraser A, Kamath RS, Ahringer J, Li H, Kenyon C (2003) Genes that act downstream of DAF-16 to influence the lifespan of Caenorhabditis elegans. Nature 424:277–283
Murphy T, Dias GP, Thuret S (2014) Effects of diet on brain plasticity in animal and human studies: mind the gap. Neural Plast 2014:563160
Neff F, Flores-Dominguez D, Ryan DP, Horsch M, Schroder S, Adler T, Afonso LC, Aguilar-Pimentel JA, Becker L, Garrett L, Hans W, Hettich MM, Holtmeier R, Holter SM, Moreth K, Prehn C, Puk O, Racz I, Rathkolb B, Rozman J, Naton B, Ordemann R, Adamski J, Beckers J, Bekeredjian R, Busch DH, Ehninger G, Graw J, Hofler H, Klingenspor M, Klopstock T, Ollert M, Stypmann J, Wolf E, Wurst W, Zimmer A, Fuchs H, Gailus-Durner V, Hrabe de Angelis M, Ehninger D (2013) Rapamycin extends murine lifespan but has limited effects on aging. J Clin Invest 123:3272–3291
Nikolai S, Pallauf K, Huebbe P, Rimbach G (2015) Energy restriction and potential energy restriction mimetics. Nutr Res Rev 28:100–120
Nisoli E, Tonello C, Cardile A, Cozzi V, Bracale R, Tedesco L, Falcone S, Valerio A, Cani O, Clementi E, Carruba MO, Valerio A (2005) Calorie restriction promotes mitochondrial biogenesis by inducing the expression of eNOS. Science 310:314–317
Omodei D, Fontana L (2011) Calorie restriction and prevention of age-associated chronic disease. FEBS Lett 585:1537–1542
Pamplona R, Barja G, Portero-Otin M (2002) Membrane fatty acid unsaturation, protection against oxidative stress, and maximum life span: a homeoviscous-longevity adaptation? Ann N Y Acad Sci 959:475–490
Pani G (2015) Neuroprotective effects of dietary restriction: evidence and mechanisms. Semin Cell Dev Biol 40:106–114
Park SK, Prolla TA (2005) Lessons learned from gene expression profile studies of aging and caloric restriction. Ageing Res Rev 4:55–65
Park SJ, Ahmad F, Philp A, Baar K, Williams T, Luo H, Ke H, Rehmann H, Taussig R, Brown AL, Kim MK, Beaven MA, Burgin AB, Manganiello V, Chung JH (2012) Resveratrol ameliorates aging-related metabolic phenotypes by inhibiting cAMP phosphodiesterases. Cell 148:421–433
Pasinetti GM, Eberstein JA (2008) Metabolic syndrome and the role of dietary lifestyles in Alzheimer’s disease. J Neurochem 106:1503–1514
Patel NV, Gordon MN, Connor KE, Good RA, Engelman RW, Mason J, Morgan DG, Morgan TE, Finch CE (2005) Caloric restriction attenuates Abeta-deposition in Alzheimer transgenic models. Neurobiol Aging 26:995–1000
Patel KR, Scott E, Brown VA, Gescher AJ, Steward WP, Brown K (2011) Clinical trials of resveratrol. Ann N Y Acad Sci 1215:161–169
Pearson KJ, Baur JA, Lewis KN, Peshkin L, Price NL, Labinskyy N, Swindell WR, Kamara D, Minor RK, Perez E, Jamieson HA, Zhang Y, Dunn SR, Sharma K, Pleshko N, Woollett LA, Csiszar A, Ikeno Y, Le Couteur D, Elliott PJ, Becker KG, Navas P, Ingram DK, Wolf NS, Ungvari Z, Sinclair DA, de Cabo R (2008) Resveratrol delays age-related deterioration and mimics transcriptional aspects of dietary restriction without extending life span. Cell Metab 8:157–168
Peric A, Annaert W (2015) Early etiology of Alzheimer’s disease: tipping the balance toward autophagy or endosomal dysfunction? Acta Neuropathol 129:363–381
Perrone CE, Malloy VL, Orentreich DS, Orentreich N (2013) Metabolic adaptations to methionine restriction that benefit health and lifespan in rodents. Exp Gerontol 48:654–660
Porquet D, Casadesus G, Bayod S, Vicente A, Canudas AM, Vilaplana J, Pelegri C, Sanfeliu C, Camins A, Pallas M, del Valle J (2013) Dietary resveratrol prevents Alzheimer’s markers and increases life span in SAMP8. Age (Dordr) 35:1851–1865
Porquet D, Grinan-Ferre C, Ferrer I, Camins A, Sanfeliu C, Del Valle J, Pallas M (2014) Neuroprotective role of trans-resveratrol in a murine model of familial Alzheimer’s disease. J Alzheimers Dis 42:1209–1220
Prolla TA (2002) DNA microarray analysis of the aging brain. Chem Senses 27:299–306
Pugh TD, Oberley TD, Weindruch R (1999) Dietary intervention at middle age: caloric restriction but not dehydroepiandrosterone sulfate increases lifespan and lifetime cancer incidence in mice. Cancer Res 59:1642–1648
Pyo JO, Yoo SM, Ahn HH, Nah J, Hong SH, Kam TI, Jung S, Jung YK (2013) Overexpression of Atg5 in mice activates autophagy and extends lifespan. Nat Commun 4:2300
Ravussin E, Redman LM, Rochon J, Das SK, Fontana L, Kraus WE, Romashkan S, Williamson DA, Meydani SN, Villareal DT, Smith SR, Stein RI, Scott TM, Stewart TM, Saltzman E, Klein S, Bhapkar M, Martin CK, Gilhooly CH, Holloszy JO, Hadley EC, Roberts SB, Group CS (2015) A 2-Year Randomized Controlled Trial of Human Caloric Restriction: feasibility and Effects on Predictors of Health Span and Longevity. J Gerontol A Biol Sci Med Sci 70:1097–1104
Redman LM, Ravussin E (2011) Caloric restriction in humans: impact on physiological, psychological, and behavioral outcomes. Antioxid Redox Signal 14:275–287
Reichardt LF (2006) Neurotrophin-regulated signalling pathways. Philos Trans R Soc Lond B Biol Sci 361:1545–1564
Ribaric S (2012) Diet and aging. Oxid Med Cell Longev 2012:741468
Richardson A, Galvan V, Lin AL, Oddo S (2015) How longevity research can lead to therapies for Alzheimer’s disease: the rapamycin story. Exp Gerontol 68:51–58
Rodgers JT, Lerin C, Haas W, Gygi SP, Spiegelman BM, Puigserver P (2005) Nutrient control of glucose homeostasis through a complex of PGC-1alpha and SIRT1. Nature 434:113–118
Rogina B, Helfand SL (2004) Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci USA 101:15998–16003
Rothman SM, Griffioen KJ, Wan R, Mattson MP (2012) Brain-derived neurotrophic factor as a regulator of systemic and brain energy metabolism and cardiovascular health. Ann N Y Acad Sci 1264:49–63
Sanchez-Roman I, Barja G (2013) Regulation of longevity and oxidative stress by nutritional interventions: role of methionine restriction. Exp Gerontol 48:1030–1042
Santos J, Leitao-Correia F, Sousa MJ, Leao C (2015) Ammonium is a key determinant on the dietary restriction of yeast chronological aging in culture medium. Oncotarget 6:6511–6523
Savaskan E, Olivieri G, Meier F, Seifritz E, Wirz-Justice A, Muller-Spahn F (2003) Red wine ingredient resveratrol protects from beta-amyloid neurotoxicity. Gerontology 49:380–383
Schafer MJ, Alldred MJ, Lee SH, Calhoun ME, Petkova E, Mathews PM, Ginsberg SD (2015) Reduction of beta-amyloid and gamma-secretase by calorie restriction in female Tg2576 mice. Neurobiol Aging 36:1293–1302
Schulingkamp RJ, Pagano TC, Hung D, Raffa RB (2000) Insulin receptors and insulin action in the brain: review and clinical implications. Neurosci Biobehav Rev 24:855–872
Selkoe DJ (2011) Alzheimer’s disease. Cold Spring Harb Perspect Biol. doi:10.1101/cshperspect.a004457
Sharma S, Singh R, Kaur M, Kaur G (2010) Late-onset dietary restriction compensates for age-related increase in oxidative stress and alterations of HSP 70 and synapsin 1 protein levels in male Wistar rats. Biogerontology 11:197–209
Shen LR, Parnell LD, Ordovas JM, Lai CQ (2013) Curcumin and aging. BioFactors 39:133–140
Shytle RD, Tan J, Bickford PC, Rezai-Zadeh K, Hou L, Zeng J, Sanberg PR, Sanberg CD, Alberte RS, Fink RC, Roschek B Jr (2012) Optimized turmeric extract reduces beta-Amyloid and phosphorylated Tau protein burden in Alzheimer’s transgenic mice. Curr Alzheimer Res 9:500–506
Siman R, Cocca R, Dong Y (2015) The mTOR Inhibitor Rapamycin Mitigates Perforant Pathway Neurodegeneration and Synapse Loss in a Mouse Model of Early-Stage Alzheimer-Type Tauopathy. PLoS ONE 10:e0142340
Simonsen A, Cumming RC, Brech A, Isakson P, Schubert DR, Finley KD (2008) Promoting basal levels of autophagy in the nervous system enhances longevity and oxidant resistance in adult Drosophila. Autophagy 4:176–184
Smith DL Jr, Elam CF Jr, Mattison JA, Lane MA, Roth GS, Ingram DK, Allison DB (2010a) Metformin supplementation and life span in Fischer-344 rats. J Gerontol A Biol Sci Med Sci 65:468–474
Smith PJ, Blumenthal JA, Babyak MA, Craighead L, Welsh-Bohmer KA, Browndyke JN, Strauman TA, Sherwood A (2010b) Effects of the dietary approaches to stop hypertension diet, exercise, and caloric restriction on neurocognition in overweight adults with high blood pressure. Hypertension 55:1331–1338
Sohal RS, Forster MJ (2014) Caloric restriction and the aging process: a critique. Free Radic Biol Med 73:366–382
Sohal RS, Weindruch R (1996) Oxidative stress, caloric restriction, and aging. Science 273:59–63
Sohal RS, Ku HH, Agarwal S, Forster MJ, Lal H (1994) Oxidative damage, mitochondrial oxidant generation and antioxidant defenses during aging and in response to food restriction in the mouse. Mech Ageing Dev 74:121–133
Spilman P, Podlutskaya N, Hart MJ, Debnath J, Gorostiza O, Bredesen D, Richardson A, Strong R, Galvan V (2010) Inhibition of mTOR by rapamycin abolishes cognitive deficits and reduces amyloid-beta levels in a mouse model of Alzheimer’s disease. PLoS One 5:e9979
Sridhar GR, Lakshmi G, Nagamani G (2015) Emerging links between type 2 diabetes and Alzheimer’s disease. World J Diabetes 6:744–751
Srivastava S, Haigis MC (2011) Role of sirtuins and calorie restriction in neuroprotection: implications in Alzheimer’s and Parkinson’s diseases. Curr Pharm Des 17:3418–3433
Stenesen D, Suh JM, Seo J, Yu K, Lee KS, Kim JS, Min KJ, Graff JM (2013) Adenosine nucleotide biosynthesis and AMPK regulate adult life span and mediate the longevity benefit of caloric restriction in flies. Cell Metab 17:101–112
Stewart J, Mitchell J, Kalant N (1989) The effects of life-long food restriction on spatial memory in young and aged Fischer 344 rats measured in the eight-arm radial and the Morris water mazes. Neurobiol Aging 10:669–675
Strong R, Miller RA, Astle CM, Baur JA, de Cabo R, Fernandez E, Guo W, Javors M, Kirkland JL, Nelson JF, Sinclair DA, Teter B, Williams D, Zaveri N, Nadon NL, Harrison DE (2013) Evaluation of resveratrol, green tea extract, curcumin, oxaloacetic acid, and medium-chain triglyceride oil on life span of genetically heterogeneous mice. J Gerontol A Biol Sci Med Sci 68:6–16
Subramanian L, Youssef S, Bhattacharya S, Kenealey J, Polans AS, van Ginkel PR (2010) Resveratrol: challenges in translation to the clinic–a critical discussion. Clin Cancer Res 16:5942–5948
Tatar M, Kopelman A, Epstein D, Tu MP, Yin CM, Garofalo RS (2001) A mutant Drosophila insulin receptor homolog that extends life-span and impairs neuroendocrine function. Science 292:107–110
Theendakara V, Patent A, Peters Libeu CA, Philpot B, Flores S, Descamps O, Poksay KS, Zhang Q, Cailing G, Hart M, John V, Rao RV, Bredesen DE (2013) Neuroprotective Sirtuin ratio reversed by ApoE4. Proc Natl Acad Sci USA 110:18303–18308
Toth ML, Sigmond T, Borsos E, Barna J, Erdelyi P, Takacs-Vellai K, Orosz L, Kovacs AL, Csikos G, Sass M, Vellai T (2008) Longevity pathways converge on autophagy genes to regulate life span in Caenorhabditis elegans. Autophagy 4:330–338
Tung BT, Rodriguez-Bies E, Ballesteros-Simarro M, Motilva V, Navas P, Lopez-Lluch G (2014) Modulation of endogenous antioxidant activity by resveratrol and exercise in mouse liver is age dependent. J Gerontol A Biol Sci Med Sci 69:398–409
Tung BT, Rodriguez-Bies E, Thanh HN, Le-Thi-Thu H, Navas P, Sanchez VM, Lopez-Lluch G (2015) Organ and tissue-dependent effect of resveratrol and exercise on antioxidant defenses of old mice. Aging Clin Exp Res 27:775–783
Turner RS, Thomas RG, Craft S, van Dyck CH, Mintzer J, Reynolds BA, Brewer JB, Rissman RA, Raman R, Aisen PS, Alzheimer’s Disease Cooperative S (2015) A randomized, double-blind, placebo-controlled trial of resveratrol for Alzheimer disease. Neurology 85:1383–1391
van der Heide LP, Ramakers GM, Smidt MP (2006) Insulin signaling in the central nervous system: learning to survive. Prog Neurobiol 79:205–221
Vaziri H, Dessain SK, Ng Eaton E, Imai SI, Frye RA, Pandita TK, Guarente L, Weinberg RA (2001) hSIR2(SIRT1) functions as an NAD-dependent p53 deacetylase. Cell 107:149–159
Vellai T, Takacs-Vellai K, Zhang Y, Kovacs AL, Orosz L, Muller F (2003) Genetics: influence of TOR kinase on lifespan in C. elegans. Nature 426:620
Viollet B, Guigas B, Sanz Garcia N, Leclerc J, Foretz M, Andreelli F (2012) Cellular and molecular mechanisms of metformin: an overview. Clin Sci (Lond) 122:253–270
Vivar C, Potter MC, van Praag H (2013) All about running: synaptic plasticity, growth factors and adult hippocampal neurogenesis. Curr Top Behav Neurosci 15:189–210
Wakeling LA, Ions LJ, Ford D (2009) Could Sirt1-mediated epigenetic effects contribute to the longevity response to dietary restriction and be mimicked by other dietary interventions? Age (Dordr) 31:327–341
Wang J, Ho L, Qin W, Rocher AB, Seror I, Humala N, Maniar K, Dolios G, Wang R, Hof PR, Pasinetti GM (2005) Caloric restriction attenuates beta-amyloid neuropathology in a mouse model of Alzheimer’s disease. FASEB J 19:659–661
Wang Y, Oh SW, Deplancke B, Luo J, Walhout AJ, Tissenbaum HA (2006) C. elegans 14-3-3 proteins regulate life span and interact with SIR-2.1 and DAF-16/FOXO. Mech Ageing Dev 127:741–747
Wang DT, He J, Wu M, Li SM, Gao Q, Zeng QP (2015) Artemisinin mimics calorie restriction to trigger mitochondrial biogenesis and compromise telomere shortening in mice. PeerJ 3:e822
Weiss EP, Fontana L (2011) Caloric restriction: powerful protection for the aging heart and vasculature. Am J Physiol Heart Circ Physiol 301:H1205–H1219
Wilkinson JE, Burmeister L, Brooks SV, Chan CC, Friedline S, Harrison DE, Hejtmancik JF, Nadon N, Strong R, Wood LK, Woodward MA, Miller RA (2012) Rapamycin slows aging in mice. Aging Cell 11:675–682
Willcox BJ, Willcox DC (2014) Caloric restriction, caloric restriction mimetics, and healthy aging in Okinawa: controversies and clinical implications. Curr Opin Clin Nutr Metab Care 17:51–58
Witte AV, Fobker M, Gellner R, Knecht S, Floel A (2009) Caloric restriction improves memory in elderly humans. Proc Natl Acad Sci USA 106:1255–1260
Witte AV, Kerti L, Margulies DS, Floel A (2014) Effects of resveratrol on memory performance, hippocampal functional connectivity, and glucose metabolism in healthy older adults. J Neurosci 34:7862–7870
Wood SH, van Dam S, Craig T, Tacutu R, O’Toole A, Merry BJ, de Magalhaes JP (2015) Transcriptome analysis in calorie-restricted rats implicates epigenetic and post-translational mechanisms in neuroprotection and aging. Genome Biol 16:285
Wu P, Shen Q, Dong S, Xu Z, Tsien JZ, Hu Y (2008) Calorie restriction ameliorates neurodegenerative phenotypes in forebrain-specific presenilin-1 and presenilin-2 double knockout mice. Neurobiol Aging 29:1502–1511
Wu JJ, Liu J, Chen EB, Wang JJ, Cao L, Narayan N, Fergusson MM, Rovira II, Allen M, Springer DA, Lago CU, Zhang S, DuBois W, Ward T, deCabo R, Gavrilova O, Mock B, Finkel T (2013) Increased mammalian lifespan and a segmental and tissue-specific slowing of aging after genetic reduction of mTOR expression. Cell Rep 4:913–920
Zhang S, Salemi J, Hou H, Zhu Y, Mori T, Giunta B, Obregon D, Tan J (2010) Rapamycin promotes beta-amyloid production via ADAM-10 inhibition. Biochem Biophys Res Commun 398:337–341
Zhang Y, Bokov A, Gelfond J, Soto V, Ikeno Y, Hubbard G, Diaz V, Sloane L, Maslin K, Treaster S, Rendon S, van Remmen H, Ward W, Javors M, Richardson A, Austad SN, Fischer K (2014) Rapamycin extends life and health in C57BL/6 mice. J Gerontol A Biol Sci Med Sci 69:119–130
Zhao HF, Li N, Wang Q, Cheng XJ, Li XM, Liu TT (2015a) Resveratrol decreases the insoluble Abeta1-42 level in hippocampus and protects the integrity of the blood-brain barrier in AD rats. Neuroscience 310:641–649
Zhao J, Zhai B, Gygi SP, Goldberg AL (2015b) mTOR inhibition activates overall protein degradation by the ubiquitin proteasome system as well as by autophagy. Proc Natl Acad Sci USA 112:15790–15797
Zhao Z, Sui Y, Gao W, Cai B, Fan D (2015c) Effects of diet on adenosine monophosphate-activated protein kinase activity and disease progression in an amyotrophic lateral sclerosis model. J Int Med Res 43:67–79
Acknowledgments
This work was supported by the Research Council of Ghent University, the Belgian Foundation of Alzheimer’s Research (SAO), and the Research Foundation Flanders (FWO Vlaanderen). We thank Dr. Amin Bredan for careful editing of the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Caroline Van Cauwenberghe and Charysse Vandendriessche equally contributed to this work.
Rights and permissions
About this article

Cite this article
Van Cauwenberghe, C., Vandendriessche, C., Libert, C. et al. Caloric restriction: beneficial effects on brain aging and Alzheimer’s disease. Mamm Genome 27, 300–319 (2016). https://doi.org/10.1007/s00335-016-9647-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00335-016-9647-6