
Abstract
The combination of an archaeobotanical analysis from two different sites in the city of Huelva (Spain) and the identification of vine fields in the same area suggests that different fruit tree species (grapevine, olive, almond and pomegranate) and vegetables such as melon were introduced into the Iberian peninsula in the transition from the 9th to the 8th century cal bc. These dates represent the earliest chronology for arboriculture within the Iberian Peninsula. The material has been preserved by waterlogging allowing the preservation of a wide variety of species which indicate the development of fruit tree cultivation. The archaeological context provides information on the connections between this innovation and the Phoenician communities that established in the region in search of metal resources. Fruit tree cultivation, and particularly wine production, had a great impact on the local agriculture which was traditionally based on the production of annual crops. The new crops soon became an essential agricultural element of the communities that lived in the south and east of the Iberian Peninsula. From the 8th century cal bc onwards, agricultural production would be mostly market oriented.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Archaeobotanical research into plant domestication has traditionally focused on cereals and legumes while comparatively much less is known about fruit domestication (Bouby and Ruas 2014), yet fruit tree cultivation is an important component of food production. Understanding the beginnings of fruit crop cultivation across the Mediterranean is not an easy task as, differently from the cereals, fruit domestication rarely displays diagnostic morphological characters that allow distinction between wild and domesticated specimens (Weiss 2015). Domestication of the main Mediterranean species, such as the olive tree (Olea europaea), grapevine (Vitis vinifera), date palm (Phoenix dactylifera) and fig (Ficus carica), occurred in the Fertile Crescent (Zohary and Spiegel-Roy 1975) from where most of the evidence derives (Zohary et al. 2012). A very early domestication of the fig tree was proposed between 11,400−11,200 cal bp at the site of Gilgal, Israel (Kislev et al. 2006) but this was later contested (Lev-Yadun et al. 2006).
According to the latest review of fruit growing in the Old World (Weiss 2015), the four species considered here, Olea, Vitis, Phoenix and Ficus, were already cultivated during the late Neolithic (seventh millennium bp) rather than in the Chalcolithic or the Early Bronze Age, but tree cultivation across the Mediterranean is detected much later, between the fourth and the third millennium cal bc. Some of these species, such as Vitis, were already being collected by Palaeolithic and Mesolithic hunter-gatherers around the Mediterranean (Weiss et al. 2004; Aura et al. 2005; Lev et al. 2005; Vaquer and Ruas 2009) and also played a significant role not only amongst the first farmer communities in these areas (Antolín and Jacomet 2015) but also in later societies. What is more, different types of evidence indicate that wine may have been produced in some areas, such as Iran, where the occurrence of tartaric acid, related to the presence of winemaking, has been identified during the sixth millennium cal bc (McGovern et al. 1996). In addition, significant evidence of pressing grapes for obtaining juice, presumably for some kind of wine, has been recorded in northern Greece as already occuring by the fifth millennium cal bc (Valamoti 2015; Valamoti et al. 2007).
However, despite the evidence, the systematic use of these species and their domestication are not yet entirely clear. Based on the presence of seeds only, several authors have proposed the domestication of certain fruit species (Rivera and Walker 1989; Buxó 1997). Moreover, different morphological criteria for distinguishing between wild and cultivated seeds have been also put forward relating to Olea (Terral et al. 2004), Vitis (Stummer 1911; Mangafa and Kotsakis 1996; Bouby et al. 2013; Ucchesu et al. 2016) and Phoenix (Gros-Balthazard et al. 2013, 2016). Likewise, substantial work on the genetics of Vitis and Olea has been carried out (Arroyo-García et al. 2006; This et al. 2006; Cappellini et al. 2010; Besnard et al. 2011; Myles et al. 2011; Wales et al. 2016). Lastly, the presence of structures related to grape processing and wine and oil production supporting the cultivation of both species has been emphasised (Gómez Bellard et al. 1993; Brun 2004; Salavert 2008; Sarpaki 2012).
What remains clear, however, is that the annual cycle that had governed the lives of farmers for the previous four millennia, changed with the introduction of fruit cultivation. Changes occurred at various levels; on the one hand, farmers’ relationship with the land was transformed by the need to exert control of and access to the land property for much longer periods than in the case of annual crops; on the other, the rhythmic pace of agrarian tasks was also disrupted as the new crops required new ways of management. In annual crops such as cereals and legumes, yields are obtained several months after sowing, while fruit trees live through a number of growing seasons producing fruit several years after planting, with optimal production only being reached much later. This protracted cycle of production favoured stronger ties to the land as communities needed to care for and protect the fields throughout more lengthy periods.
In terms of nutrition, cultivated fruits provided a wide variety of sugar, vitamin, mineral and fibre supplements to the diet, and contributed a great diversity of other important economic products (firewood, fodder, medicines, fuel etc.) and services (shade, fencing etc.) of interest to past communities (Bouby and Ruas 2014; Braadbaart et al. 2016). Furthermore, in some cases fruit cultivation allowed the putting into cultivation of areas unsuited for annual crops.
Some authors (Renfrew 1972; Gilman 1981) have related the development of fruit cultivation to social complexity based on the rather substantial investment put into its production, only achievable by the most privileged classes. Within the context of hierarchical social relations, the proponents of this model see only ruling classes as capable of sustaining not only delayed economic returns, but also guaranteeing distribution, since many fruit products, particularly wine and oil, have been frequently commercialized (Leonard 1997; Zamora 2000).
The spread of fruit products to the western Mediterranean has often been connected to the inter-regional exchange networks existing in the area. Recent work in Sardinia has shown that arboriculture had already emerged in the middle of the second millennium cal bc (Sabato et al. 2015) while further west the evidence of this appears to be more recent (Buxó 2008; Bouby et al. 2014).
In this paper, archaeobotanical material from the city of Huelva (southern Spain) is presented. These new data allow the pushing back of the beginnings of fruit cultivation in the Iberian Peninsula to the transition between the 9th and the 8th centuries bc.
The sites
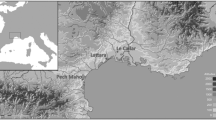
The city of Huelva is located to the west of the Gibraltar Strait, on a small peninsula surrounded by the estuaries of the rivers Tinto and Odiel which flow into the Atlantic sea. The last 4,000 years have witnessed a rapid sedimentary filling of the estuary giving way to a marshy environment around this small peninsula, quite different from the milieu characterizing the prehistoric and protohistoric occupation of the area (Fig. 1) (Vera et al. 2010) in both the city (Fernández Jurado 1990) and its surroundings (Gómez Toscano et al. 2014).

Map with the location of the site showing the area excavated (1) and the vineyards (2), from a theoretical reconstruction of the coastline during the Flandrian marine transgression
Recent archaeological work in two urban plots in the city centre of Huelva has uncovered the earliest archaeological levels of the Iberian Peninsula related to the colonization period. These contain abundant Phoenician, Greek and Sardinian materials together with local products (González de Canales et al. 2006; García Fernández et al. 2015). The archaeobotanical material comes from these contexts.
The various occupations and activities that gave rise to these deposits are connected to the presence in this area of an important protohistoric port (Gómez Toscano 2009) where during the transition from the 9th to the 8th centuries bc, and perhaps earlier, the presence of oriental groups related to the exploitation of the rich mining deposits of the Sierra of Huelva (Campos and Alvar 2013) was significant. Contemporary with this occupation, various technical innovations were progressively adopted. Of them, the development of arboriculture is certainly one of the most relevant (Vera and Echevarría 2013; Echevarría and Vera 2015; Pérez Jordà 2015).
Materials and methods
The archaeobotanical material under study was recovered from two different urban plots in the city of Huelva. In 1998, samples were collected from a plot in Calle Méndez Núñez 7–13 (from now on Méndez Núñez) (González de Canales et al. 2004, 2006), while in 2009 and 2010 samples were obtained from Calle Concepción 3 (from now on Concepción). In both cases, the archaeological layers were excavated down to the water table. In Méndez Núñez, water was pumped out so the layers below the water table, including a 1 m thick dark layer that contained archaeobotanical remains, could be mechanically removed. The samples studied here were taken from this removed soil.
Distinction between the upper and lower layers at Méndez Núñez was based on the sediment colour being the dark earth characteristic of the layers below the water table. Since within the dark sediment a proper stratigraphic sequence was not detected the archaeobotanical remains were all collected as one single sample. The samples were deposited in the Huelva city museum were they were kept for several years.
In Concepción, the level below the water table was ca. 80 cm thick and it consisted of different alternating layers of sandy matrix and levels rich in plant material. Most samples were collected by hand from this layer except A04 and A05 which were mechanically extracted. Samples were then sieved out by the excavators. The residues, together with the samples from Méndez Núñez, were later sent to the Archaeobotany Lab of the Instituto de Historia (CSIC) in Madrid, where sorting and identification were carried out. Samples have been fully examined although there is no information on the original sample sizes. As far as conservation is concerned, the archaeobotanical material from Concepción was intentionally dried out by the excavators so deformation and breaking were detected, and it is likely that some material got destroyed. In Méndez Núñez, sampling and processing were also carried out by excavators without archaeobotanical training, which to some extent limits the results of this study. There are 11 samples from Concepción while from Méndez Núñez the only available sample was analysed. The identification of remains was done using the reference collection of the CSIC. Botanical names of cultivated plants follow the traditional binomial classification used by Zohary et al. (2012), while for the wild specimens the Flora Ibérica (Castroviejo 1986–2012) has been followed.
Results
The material under study (Table 1) was preserved by both charring and waterlogging, the latter remains being much more numerous. The charred remains, even if present in all samples, never exceeded 5%.
Charred remains
Both cereal grains and chaff have been recovered from both sites (Fig. 2). The commonest species is hulled barley (Hordeum vulgare ssp. vulgare) from which only one caryopsis has been identified as belonging to a “dense eared type” following van Zeist (1968). The free-threshing wheat group is the second most frequent, being represented only by caryopses, which impedes distinction between both species. Spikelets of possible emmer (Triticum cf. dicoccum) appear only occasionally.

Charred seeds and fruits. 1 Hordeum vulgare ssp. vulgare, 2 Hordeum vulgare rachis, 3 Triticum aestivum-durum, 4 Triticum cf. dicoccum spikelet fork, 5 Triticum cf. dicoccum spikelet fork, 6 Lens culinaris, 7 Chamaerops humilis, 8 Lolium temulentum. Scale bar 1 mm
Legumes are only present in Concepción although in rather sparse numbers. The only species documented are lens (Lens culinaris), pea (Pisum sativum) and broad bean (Vicia faba). Wild plants have been also identified. Stones of the dwarf palm (Chamaerops humilis), several grasses (Lolium sp., Lolium temulentum and Phalaris sp.), one Malvaceae (Malva sylvestris/nicaensis) and one Polygonaceae (Polygonum sp.) are the species represented. Grasses, and in particular L. temulentum, are the dominant taxa.
Waterlogged remains
A large part of the plant material (Fig. 3) was preserved by waterlogging. It consists mostly of remains of F. carica and V. vinifera, the two most abundant species, which have been identified in both sites. In the case of Vitis, both seeds and pedicels are present. There are also seeds from melon (Cucumis melo), pomegranate (Punica granatum) and, more rarely, olives (O. europaea) and almonds (Prunus dulcis) were also recovered. As far as cereals and legumes are concerned, waterlogging does not favour their preservation apart from glumes. In fact, the waterlogged remains include only a single millet (Panicum miliaceum) glume.

Waterlogged cultivated seeds and fruits. 1 Panicum miliaceum, 2 Cucumis melo, 3 Ficus carica, 4 Prunus dulcis, 5 Olea europaea, 6 Linum usitatissimum, 7 Punica granatum, 8 Vitis vinifera, 9 Papaver somniferum. Scale bar 1 mm
A further group is represented by the wild fruits (Fig. 4) including edible species such as white crowberry (Corema album), myrtle (Myrtus comunis), bracts and seeds from stone and Aleppo pines (Pinus pinea, P. halepensis/maritima) and blackberries (Rubus fruticosus). Additionally, seeds from juniper (Juniperus phoenicea) and the mastic tree (Pistacia lentiscus), also acorn (Quercus sp.) hulls and cupules, were recovered, but in lower proportions.

Waterlogged wild plants. 1 Corema album, 2 Juniperus phoenicea, 3 Myrtus communis, 4 Pinus pinea, 5 Quercus sp., 6 Rubus fruticosus, 7 Indet., 8 Asteraceae, 9 Emex spinosa, 10 Chenopodium sp., 11 Fumaria sp., 12 Medicago cf. marina, 13 Medicago sp., 14 Plantago sp., 15 Portulaca oleracea, 16 Raphanus raphanistrum, 17 Reseda phyteuma, 18 Apiaceae, 19 Ranunculus sp., 20 Calendula officinalis. Scale bar 1 mm
Albeit less abundant, other wild species thriving in the typical marshy sandy soils of the area were also identified. These are represented by Raphanus raphanistrum, Rosmarinus officinalis, Silene sp., Thymelaea sp., Anagallis arvensis, Ajuga sp., Corrigiola litoralis, Emex spinosa, Hyoscyamus niger, Medicago cf. marina, Medicago cf. littoralis and Reseda phyteuma. Other taxa from clayish soils such as Hedysarum coronarium and Calendula officinalis were also found.
As far as dating is concerned, four AMS dates carried out on seeds and fruits were obtained (Table 2). A charred caryopsis of barley from Méndez Núñez was dated to ca. the 10th century cal bc which matches previous dates on animal bones from the site (Nijboer and van der Plicht 2006). Also a seed of V. vinifera from the same site was dated to the transition between the 9th and the 8th century cal bc.
In Concepción, a charred barley caryopsis and a grapevine seed were also dated to the transition between the 9th and the 8th century cal bc, providing very similar dates to those from Méndez Núñez. Since in Concepción there is no evidence of different phases of occupation, it is likely that the archaeobotanical material from this site is contemporary with that from the youngest phase of Méndez Núñez.
Discussion
Context and taphonomic problems
A major question that emerged during the study of the material from Méndez Núñez concerned the chronology of the dark sediments below the water table from where the samples studied here come. Archaeologists in charge of the excavation described a single phase represented by a 1 m thick homogeneous layer composed of grey silt rich in organic matter with no divisions. Accordingly, the material found, particularly the pottery, was also attributed to the same chronological period (González de Canales et al. 2006, 2008). However, based on the pottery, other authors claim that this dark earth deposit contained different chronological phases that could be recognized (Gilboa et al. 2008).
One element that may contribute to the chronological discussion of the Méndez Núñez site is provided by the AMS dates (Table 2), obtained on short lived elements (seeds), which suggest the existence of two different chronological periods. One dated to the 10th century cal bc, consistent with the dates obtained on faunal remains from the same level (Nijboer and van der Plicht 2006) and also coinciding with the chronology of a bronze deposit found in the Huelva estuary dated to the 10th century (Ruiz Gálvez 1995). The second period dated to the transition from the 9th to the 8th century cal bc. According to these dates, we suggest that during the 10th century bc, the area was occupied by an indigenous settlement and that the faunal and botanical remains from this period may have resulted from events of rubbish disposal at the site. In fact, the archaeobotanical assemblage from this period, preserved by charring, is composed of plant categories (cereals, legumes and wild plants, Table 1) commonly found in indigenous sites dated to the transition between the second and the first millennium bc when fruit cultivation was not yet attested (Pérez Jordà 2013).
A second assemblage from this site, more abundant and preserved by waterlogging, has been dated to the transition from the 9th to the 8th century bc, which is in agreement with the chronology suggested by the pottery from this layer (González de Canales et al. 2006, 2008; Gilboa 2013). This phase is more recent and it seems contemporaneous with the layers excavated in Concepción (García Fernández et al. 2015). If the AMS dates are taken to represent two different phases of occupation, the earliest one would be that of the charred plant remains (cereals and legumes) and the animal bones, interpreted as rubbish leftovers from human activities. There is no evidence of waterlogged material from this layer which suggests that the area was above the surface during this early phase.
The plant assemblage from the transition from the 9th to the 8th century cal bc is different. Charred remains are less and most of the plant material (cultivated fruits and wild plants) is waterlogged due to the flooding of the site, probably related to changes in marine levels (Goy et al. 1996).
Marshlands and field crops
Plant remains include both cultivated and wild species. Some of the taxa are interpreted as refuse from human activities carried out in the area while some others appear to represent the natural vegetation typical of marshy areas. This mixture of species from human and natural origins is also found in lake sites (Brombacher 1997; Jacomet and Brombacher 2005; Antolín 2016) and in areas under marine influence (Peña-Chocarro and Zapata Peña 1996, 2005; Teira 2010). In the samples studied, wild plants are sparsely represented but this may reflect sampling and processing limitations as well as problems related to the treatment of the material after its collection by non-specialists.
A large part of the botanical taxa identified, such as Anagallis arvensis, Corema album, Corrigiola litoralis, Emex spinosa, Hyoscyamus niger, Juniperus phoenicea, Myrtus communis, Raphanus raphanistrum (perhaps the ssp. landra), and Reseda phyteuma, thrive well in waterlogged sandy soils. There are also species more adapted to waterlogged clay soils such as Calendula officinalis and Hedysarum coronarium, although the latter does not thrive in saline soils. Moreover, some taxa just develop in waterlogged soils without showing soil type preferences as is the case with Ajuga sp., which may well be A. reptans.
Other species found correspond to crop weeds or ruderal species. From these some have been preserved waterlogged, such as Portulaca oleracea, Malva sp. and Medicago littoralis, although the larger part is charred (Lolium sp., L. temulentum, Polygonum sp.).
It is clear that an important part of the plant remains seems to have accumulated naturally. Among them, some of the taxa could have been used as food or for other purposes, as is the case with Chamaerops humilis, Pinus pinea and P. halepensis/maritima, Quercus sp., Corema album, Rubus fruticosus, Myrtus communis and Pistacia lentiscus. All of these have edible fruits and their use has been documented in other sites from the southern part of the Iberian Peninsula. For instance, the Palaeolithic levels of Cueva de Nerja (Málaga) contained pine nuts (Badal 1998; Aura et al. 2002) while they have been also identified from within the first millennium bc along the Andalousian Atlantic coast (Chamorro 1994). Equally, acorns like blackberries are traditional resources that have been exploited until the present day.
An interesting case is that of C. album, a typical species of the Atlantic fringe of the Iberian Peninsula. Its edible fruits are characterized by their refreshing quality and ability to quench thirst and have been traditionally used in western Andalucía (Gil-López 2011). None of the seeds is charred so it is unclear whether their presence may just reflect a natural accumulation of seeds without human intervention as this species thrives well around waterlogged areas or to human use.
Cultivated plants are clearly related to human activies. While cereals and legumes are charred, other cultivated species such as the fruits have been preserved by waterlogging. Seeds of C. melo, Ficus, Olea, P. dulcis, Punica and Vitis, some present in large numbers, can be safely interpreted as food residues from human activities carried out in this area.
The archaeological record has not provided information on the range of domestic structures that may have existed in this area between the 9th and the 8th century cal bc. However, some of the identified elements suggest the presence of a harbour where domestic residues, including seeds and fruits, may have accumulated. Such accumulations are often found in port contexts very similar to the context analysed here. In the Iberian Peninsula, the site of the Roman port of Irun (Peña-Chocarro and Zapata Peña 2005) is significant while further examples come from the Mediterranean area (Šoštarić and Küster 2001; Bertacchi et al. 2008; Jennifer 2010; Allevato et al. 2016).
First fruit trees
The main issue related to this interesting assemblage concerns the possibility of establishing the beginnings of tree fruit production in the Iberian Peninsula. According to some researchers (Gilman 1981; Stika and Jurich 1999; Rovira 2007), with reference to O. europaea, F. carica and V. vinifera, fruit culture may already have been practiced between the third and the second millennium bc. All of the species quoted grow wild in the region and archaeobotanical data seems to suggest they, particularly the wild olive, may have been intensively used during the third millennium cal bc in the southeastern part of the Iberian Peninsula. Surprisingly, archaeobotanical data indicates that this practice was not continued during the following millennium (Alonso et al. 2016).
Beyond the use of taxa growing naturally in the area, the archaeobotanical record has yielded other species, originally from the Near East, that are absent from the wild flora of the Iberia Peninsula. This is the case for Prunus dulcis, Punica granatum and Cucumis melo that arrived already domesticated to the Iberian Peninsula. Their presence in western Mediterranean sites is clearly dated to later that in the eastern part. In Sardinia, the archaeobotanical record does not provide evidence of their cultivation until the last third of the second millennium bc at the site of Sa Osa (Ucchesu et al. 2015; Sabato et al. 2015). Here, the botanical material comes from waterlogged contexts, in particular from three wells, and the assemblage is very similar to that presented in this paper; Ficus and Vitis concentrations are abundant while melon seeds and olives are also present. The main difference between the Iberian and the Sardinian sites is the presence of black mulberry (Morus nigra) at Sa Osa and the absence of almonds and pomegranates, which are correspondingly present in Huelva.
Notwithstanding the chronological gap between data from Sardinia (1300–1200 bc) and from Huelva (800 bc), both areas exerted a powerful attraction on oriental populations who established in both regions in search of metals, probably silver (Kassianidou 2006; Murillo-Barroso et al. 2016). Fruit tree production was likely introduced in these areas through colonial encounters between Sardinians and Cypriots on the island, while people from Huelva region would have had contacts with Phoenicians. In both cases, fruit culture and the production of derivatives such as olive oil and wine were part of the knowledge brought by colonisers.
Up to the most recent discovery from Huelva, the earliest plant assemblages including domesticated fruits came from the late 8th century bc levels from Castillo de Doña Blanca (Puerto de Santa María, Cádiz) (Chamorro 1994). Later finds come from the Mediterranean coast and the Ebro Valley at sites like Alt de Benimaquía (Alicante), Cabezo de la Cruz (Zaragoza) or Font de la Canya (Barcelona) among others (Pérez Jordà 2009, 2013; López et al. 2013).
Based on the available data what remains clear is that colonisers from the eastern Mediterranean had a significant influence on the beginnings of fruit tree cultivation (Buxó 2008; Iriarte et al. 2016). This new practice was later expanded among the local population. Within this context, Huelva is of great interest not only because of its archaeobotanical record, but also because the nearby territory has provided evidence of a prehistoric vineyard which testifies to the cultivation of V. vinifera. In fact, at the site of Orden-Seminario (Huelva) it has been possible to document a plot with the first evidences of grape cultivation dated back to the transition from the 9th to the 8th century cal bc, a date that matches well the one provided by the grape pips. The chronology of the most ancient traces of cultivation has been established from the pottery found within the filling of the planting pits and based on the stratigraphic relationships of cuts and re-cuts that occurred with most recent habitat structures and other planting trenches or pits. These two criteria allow the proposal of phases of use defined in ante quem and post quem chronological terms (Vera and Echevarría 2013). This small vineyard, ca. 0.35 ha., dated to this period was subsequently followed by a larger one dated to the transition between the 7th and the 6th century cal bc. The data collected suggest that fruit tree production, and in particular viticulture, was already established in the area since earlier times. The possibility that part of the fruits was arriving through commercial networks with other Mediterranean areas is open, but in the case of V. vinifera local cultivation seems proven, while for the remaining fruit species documented at the site this may also well be the case.
This rapid spread of Vitis cultivation, and possibly of winemaking has been already detected in southern France after the foundation of Massalia (Bouby et al. 2014). In both areas, southern Spain and southern France, these products acquired a significant role linked to the economic development of their territories.
Conclusions
The analysis of two assemblages of plant remains from Huelva, together with the evidence provided by the excavation of a prehistoric vineyard at the site of La Orden-Seminario (Vera and Echevarría 2013), permits the suggestion of the cultivation of different fruit species, and in particular grapes, in the transition between the 9th and the 8th century cal bc. Furthermore, the data obtained allow the connection of this agricultural development with trade networks established by eastern Mediterranean seafarers in search of metals.
Fruit cultivation was not a local phenomenon but a foreign innovation adopted by indigenous communities, and a good example of this intense relationship between cultivation processes and trade activities with oriental colonisers is represented by the archaeological record from both Sardinia and Huelva. In these regions, prior to the arrival of eastern Mediterranean traders, species such as Olea or Vitis, growing wild in the area, appeared only occasionally in the archaeobotanical record, whereas both would systematically and abundantly turn up in the plant remain assemblages as soon as commercial contacts were established. Other species such as almond, pomegranate or melon would also be present and may be considered as introduced crops.
Indigenous populations must have adopted the new arrivals relatively quickly, although there is still a chronological gap between the remains from Huelva and the earliest examples of Vitis cultivation and winemaking in indigenous contexts such as that of Alt de Benimaquia dated to the end of the 7th century cal bc (Gómez Bellard et al. 1993; Pérez Jordà 2013). Much research is still needed to further clarify the pace and spread of this process as well as to provide detailed explanations of its development.
The new crops, and their derivatives, not only added an important supplement to diet but they also enlarged the possibilities of agriculture. Poor soils, inadequate for cereal and legume cultivation were instead suitable for fruit trees. In addition, the significant impact of a technology represented by the first iron tools transformed agricultural production and gave greater stability to local communities. This, in turn, originated population growth and spread. Vitis cultivation was intimately related to the development of wine making. Moreover, wine acquired a prominent role in social relations from this period onwards (Dietler 1990, 2010). In fact, in certain periods, the intense wine production determined an agrarian model that was clearly market oriented (Pérez Jordà 2015).
Defining the way this process occurred is one of the key issues in current research. The arrival of a series of fruit varieties originally from the eastern Mediterranean has been traditionally assumed, but morphological studies of grape pips (Bouby et al. 2013, 2014), together with genetic research (Arroyo-García et al. 2006), point to the important role of local varieties in the process. New research is essential to explore whether grape varieties entered into the colonised areas or local vine stocks were put into cultivation, or perhaps if both modalities were practiced in a more complex process. Nevertheless, whatever the process was, it was prompted by contacts with knowledgeable groups familiar with the techniques, who introduced this innovation into the local agriculture. From this point onwards, different groups living in the same territory sharing technology and culture will develop giving way to a new scenario where fruit cultivation, basically grape, became one of the key elements.
References
Allevato E, Saracino A, Fici S, Di Pasquale G (2016) The contribution of archaeological plant remains in tracing the cultural history of Mediterranean trees: the example of the Roman harbour of Neapolis. Holocene 4:603–613
Alonso N, Pérez Jordà G, Rovira N, López Reyes D (2016) Gathering and consumption of wild fruits in the east of the Iberian Peninsula from the 3rd to the 1st millennium bc. Q Int 404(Part A):69–85
Antolín F (2016) Local, intensive and diverse? Early farmers and plant economy in the North-East of the Iberian Peninsula (5500–2300 cal bc). Advances in Archaeobotany 2. Barkhuis, Groningen
Antolín F, Jacomet S (2015) Wild fruit use among early farmers in the Neolithic (5400–2300 cal bc) in the north-east of the Iberian Peninsula: an intensive practice? Veget Hist Archaeobot 24:19–33
Arroyo-García R, Ruiz-García L, Bolling L et al. (2006) Multiple origins of cultivated grapevine (Vitis vinifera L. ssp. sativa) based on chloroplast DNA polymorphisms. Mol Ecol 15:3,707–3,714
Aura JE, Jordá Pardo JF, Pérez Ripoll M, Rodrigo García MJ, Badal García E, Guillem Calatayud P (2002) The far south: the Pleistocene–Holocene transition in Nerja Cave (Andalucía, Spain). Q Int 93–94:19–30
Aura JE, Carrión Y, Estrelles E, Pérez Jordà G (2005) Plant economy of hunter-gatherer groups at the end of the last Ice Age: plant macroremains from the cave of Santa Maira (Alacant, Spain) ca. 12000–9000 B.P. Veget Hist Archaeobot 14:542–550
Badal E (1998) El interés económico del pino piñonero para los habitantes de la Cueva de Nerja. In: Sanchidrián JL, Simón Vallejo MD (eds) Las culturas del Pleistoceno Superior en Andalucía. Patronato de la Cueva de Nerja, Málaga, pp 287–300
Bertacchi A, Lombardi T, Sani A, Tomei PE (2008) Plant macroremains from the Roman harbour of Pisa (Italy). Environ Archaeol 13:181–188
Besnard G, Hernandez P, Khadari B, Dorado G, Savolainen V (2011) Genomic profiling of plastid DNA variation in the Mediterranean olive tree. BMC Plant Biol 11:80
Bouby L, Ruas MP (2014) Introduction. In: Bouby L, Ruas M-P (eds) Chapter 4: Adding diversity. Between occasional food and speculative productions: diversity of fruit uses, diversity of practices regarding fruit tree cultivation. In: Chevalier A, Marinova E, Peña-Chocarro L (eds) Plants and people. Choices and diversity through time. Oxbow Books, Oxford, pp 141–149
Bouby L, Figueiral I, Bouchette A, Rovira N, Ivorra S, Lacombe T, Pastor T, Picq S, Marinval P, Terral JF (2013) Bioarchaeological insights into the process of domestication of grapevine (Vitis vinifera L.) during Roman times in southern France. PLoS One 8:e63195
Bouby L, Marinval P, Terral JF (2014) From secondary to speculative production? The protohistory history of viticulture in southern France. In: Bouby L, Ruas M-P (eds) Chapter 4: Adding diversity. Between occasional food and speculative productions: diversity of fruit uses, diversity of practices regarding fruit tree cultivation. In: Chevalier A, Marinova E, Peña-Chocarro L (eds) Plants and people. Choices and diversity through time. Oxbow, Oxford, pp 175–181
Braadbaart F, Marinova E, Sarpaki A (2016) Charred olive stones: experimental and archaeological evidence for recognizing olive processing residues used as fuel. Veget Hist Archaeobot 25:415–430
Brombacher C (1997) Archaeobotanical investigations of Late Neolithic lakeshore settlements (Lake Biel, Switzerland). Veget Hist Archaeobot 6:167–186
Brun JP (2004) Archéologie du vin et de l’huile. De la préhistoire à l’époque hellénistique. Editions Errance, Paris
Buxó R (1997) Presence of Olea europaea and Vitis vinifera in archaeological sites from the Iberian Peninsula. Lagascalia 19:271–282
Buxó R (2008) The agricultural consequences of colonial contacts on the Iberian Peninsula in the first millennium. Veget Hist Archaeobot 17:145–154
Campos JM, Alvar J (eds) (2013) Tarteso. El Emporio del metal. Colección Historia. Serie Huellas del Pasado. Editorial Almuzara, Córdoba
Cappellini E, Gilbert MTP, Geuna F et al (2010) A multidisciplinary study of archaeological grape seeds. Naturwissenschaften 97:205–217
Castroviejo S (1986–2012) Flora Ibérica vol 1–8, 10–15, 17–18, 21. Real Jardín Botánico. CSIC, Madrid
Chamorro J (1994) Flotation startegy: method and sampling plant dietary resources of Tartessian times at Doña Blanca. In: Roselló E, Morales A (eds) Castillo de Doña Blanca. Archaeo-environmental investigations in the Bay of Cádiz (750–500 bc). BAR Int Ser 593. Tempus Reparatum, Oxford, pp 21–36
Dietler M (1990) Driven by drink: the role of drinking in the political economy and the case of early Iron Age France. J Anthropol Archaeol 9:352–406
Dietler M (2010) Archaeologies of colonialism: consumption, entanglement, and violence in ancient Mediterranean France. University of California Press, Berkeley
Echevarría A, Vera JC (2015) Los inicios de la viticultura en la Península Ibérica a partir de las huellas de cultivo. In: Francia Verde R (ed) Historia y arqueología en la cultura del vino. Instituto de Estudios Riojanos, Logroño, pp 57–68
Fernández Jurado J (1990) Tartessos y Huelva. Huelva Arqueológica X-XI. Diputación Provincial de Huelva, Huelva
García Fernández M, González de Canales Cerisola F, Serrano Pichardo L, Llompart Gómez J, Ramón Torres J, Domínguez Monedero A (2015) New data about the beginning of the Phoenician colonization in Huelva. Archaeological excavation in 3, Concepción Street. Forum Romanum Belgicum, Roma
Gilboa A (2013) À propos Huelva: a reassessment of ‘Early’ Phoenicians in the West. In: Campos JM, Alvar J (eds) Tarteso. El Emporio del metal. Colección Historia. Serie Huellas del Pasado. Editorial Almuzara, pp 311–342
Gilboa A, Sharon I, Boaretto E (2008) Tel Dor and the chronology of phoenician “pre-colonization” stages. In: Sagona C (ed) Beyond the homeland: markers in Phoenician chronology. Monograph Series of Ancient Near Eastern Studies. Louvain, Dudley, pp 113–204
Gil-López MJ (2011) Etnobotánica de la camarina (Corema album, Empetraceae) en Cádiz. Acta Bot Malacit 36:137–144
Gilman A (1981) The development of social stratification in Bronze Age Europe. Curr Anthropol 1:1–23
Gómez Bellard C, Guérin P, Pérez Jordà G (1993) Témoignage d’une production de vin dans l’ Espagne préromaine. In: Amouretti MC, Brun JP (eds) La production du vin et l’ huile en Mediterranée. Bulletin de Correspondance Héllénique, No Suppl 26, pp 379–395
Gómez Toscano F (2009) Huelva en el año 1000 a.c., un puerto cosmopolita entre el Atlántico y el Mediterráneo. Gerión 27:33–65
Gómez Toscano F, Beltrán JM, González Batanero D, Vera JC (2014) El Bronce Final en Huelva. Una visión preliminar del poblamiento en su ruedo agrícola a partir del registro arqueológico de La Orden-Seminario. Complutum 25:139–158
González de Canales F, Serrano L, Llompart J (2004) El emporio fenicio precolonial de Huelva: ca 900–700 a.c. Biblioteca Nueva, Madrid
González de Canales F, Serrano L, Llompart J (2006) The pre-colonial Phoenician emporium of Huelva ca 900–770 bc. Bull Antieke Beschav 81:13–29
González de Canales F, Serrano L, Llompart J (2008) The emporium of Huelva and Phoenician chronology: present and future possibilities. In: Sagona C (ed) Beyond the homeland: markers in Phoenician chronology. Monograph series of ancient near eastern studies. Louvain. Dudley, Paris, pp 631–655
Goy J, Zazo C, Dabrio CJ, Lario J, Borja F, Sierro FJ, Flores J (1996) Global and regional factors controlling changes of coastlines in Southern Iberia (Spain) during the Holocene. Q Sci Rev 15:773–780
Gros-Balthazard M, Newton C, Ivorra S, Tengberg M, Pintaud J-C, Terral J-F (2013) Origines et domestication du palmier-dattier (Phoenix dactylifera L.): État de l’art et perspectives d’étude. Revue d’Ethnoécologie 4. doi:10.4000/ethnoecologie.1524
Gros-Balthazard M, Newton C, Ivorra S, Pierre MH, Pintaud JC, Terral J-F (2016) The domestication syndrome in Phoenix dactylifera seeds: toward the identification of wild date palm populations. PLoS One 11:e0152394
Iriarte MJ, Ocete-Pérez CA, Hernández Beloqui B, Ocete-Rubio R (2016) Vitis vinifera in the Iberian Peninsula: a review. Plant Biosyst 151:245–257
Jacomet S, Brombacher C (2005) Reconstructing intra-site patterns in Neolithic lakeshore settlements: the state of archaeobotanical research and future prospects. In: Della Casa Ph, Trachsel M (eds) WES’04—Wetland Economies and Societies. Collectio Archaeologica 3. Chronos, Zürich, pp 69–94
Jennifer R (2010) Trade or trash: an examination of the archaeobotanical remains from the Byzantine harbour at Caesarea Maritima, Israel. Int J Naut Archaeol 39:376–382
Kassianidou V (2006) The production, use and trade of metals in Cyprus and Sardinia: so similar and yet so different. Instrumentum 23:9–11
Kislev ME, Hartmann A, Bar-Yosef O (2006) Early domesticated fig in the Jordan Valley. Science 312:1,372–1,374
Leonard AJ (1997) “Canaanite jars” and the Late Bronze Age Aegeo-Levantine wine trade. In: McGovern PE, Fleming SJ, Katz SH (eds) The origins and ancient history of wine. Gordon and Breach Publishers, New York, pp 233–254
Lev E, Kislev ME, Bar-Yosef O (2005) Mousterian vegetal food in Kebara Cave, Mt. Carmel. J Archaeol Sci 32:475–484
Lev-Yadun S, Ne’eman G, Abbo S, Flaishman MA (2006) Comment on “Early domesticated fig in the Jordan Valley”. Science 314:1,683a
López D, Jornet R, Morer J, Asensio D (2013) La viticultura prerromana al Penedès: indicadors arqueològics a Font de la Canya (Avinyonet del Penedès, Alt Penedès, Barcelona). In: Sancho D (ed) El món de la viticultura, els vins, caves i aiguardents al Penedès i al Camp de Tarragona. Actes del V Seminari d’Història del Penedès. Institut d’Estudis Penedesencs, Vilafranca del Penedès, pp 31–52
Mangafa M, Kotsakis K (1996) A new method for the identification of wild and cultivated charred grape seeds. J Archaeol Sci 23:409–418
McGovern PE, Glusker DL, Exner LJ, Voight MM (1996) Neolithic resinated wine. Nature 381:480–481
Murillo-Barroso M, Montero-Ruiz I, Rafel N, Hunt Ortiz MA, Armada XL (2016) The macro-regional scale of silver production in Iberia during the first millennium bc in the context of Mediterranean contacts. Oxford J Archaeol 35:75–100
Myles S, Boyko AR, Owens CL et al. (2011) Genetic structure and domestication history of the grape. Proc Natl Acad Sci USA 108:3,530–3,535
Nijboer AJ, van der Plicht J (2006) An interpretation of the radiocarbon determinations of the oldest indigenous-Phoenician stratum thus far, excavated at Huelva, Tartessos (south-west Spain). Babesch 81:31–36
Peña-Chocarro L, Zapata Peña L (1996) Los recursos vegetales en el mundo romano: estudio de los macrorrestos botánicos del yacimiento calle Santiago de Irún (Guipúzcoa). Arch Espan Arqueol 69:119–134
Peña-Chocarro L, Zapata Peña L (2005) Trade and new plant foods in the western Atlantic coast: the Roman port of Irun (Basque Country). In: Urteaga Artigas MM, Noain Maura MJ (eds) Mar Exterior. El Occidente Atlántico en época romana. EEHAR, Rome, pp 169–177
Pérez Jordà G (2009) Estudio paleocarpológico. In: Picazo Millán JV, Rodanés Vicente JM (eds) Los poblados del Bronce Final y de la Primera Edad del Hierro. Cabezo de la Cruz (La Muela. Zaragoza). Gobierno de Aragón, Zaragoza, pp 170–187
Pérez Jordà G (2013) La agricultura en el País Valenciano entre el VI y el I milenio a.C. PhD Thesis, Universitat de València, València
Pérez Jordà G (2015) El cultivo de la vid y la producción de vino en la Península Ibérica durante el I milenio ANE. In: Francia Verde R (ed) Historia y arqueología en la cultura del vino. Instituto de Estudios Riojanos, Logroño, pp 47–56
Reimer PJ, Bard E, Bayliss A et al. (2013) IntCal13 and Marine13 radiocarbon age calibration curves, 0–50,000 years cal bp. Radiocarbon 55:1,869–1,887
Renfrew C (1972) The emergence of civilisation: the Cyclades and the Aegean in the third millenium bc. Methuen, London
Rivera D, Walker MJ (1989) A review of palaeobotanical findings of early Vitis in the Mediterranean and of the origins of cultivated grapevines, with special reference to new pointers to prehistoric exploitation in the Western Mediterranean. Rev Palaeobot Palynol 61:205–237
Rovira N (2007) Agricultura y gestión de los recursos vegetales en el sureste de la Península Ibérica durante la prehistoria reciente. PhD Thesis, Universitat Pompeu Fabra, Barcelona
Ruiz Gálvez M (ed) (1995) Ritos de paso y puntos de paso. La Ría de Huelva en el mundo del Bronce Final europeo. Complutum Extra 5, UCM, Madrid
Sabato D, Masi A, Pepe C, Ucchesu M, Peña-Chocarro L, Usai A, Giachi G, Capreti C, Bacchetta G (2015) Archaeobotanical analysis of a Bronze Age well from Sardinia: a wealth of knowledge. Plant Biosyst 149:205–215
Salavert A (2008) Olive cultivation and oil production in Palestine during the early Bronze Age (3500–2000): the case of Tel Yarmouth, Israel. Veget Hist Archaeobot 17 Suppl 1:S53–S61
Sarpaki A (2012) Re-visiting the visibility of grape, grape products, by-products and some insights of its organization from the Prehistoric Aegean, as guided by new evidence from Monastiraki, Crete. IANSA 3:181–191
Šoštarić R, Küster H (2001) Roman plant remains from Veli Brijun (island of Brioni), Croatia. Veget Hist Archaeobot 10:227–233
Stika H-P, Jurich B (1999) Kupferzeitliche Pflanzenreste aus Almizaraque und Las Pilas, prov. Almería, Südostspanien. Madr Mitt 40:72–79
Stummer A (1911) Zur Urgeschichte der Rebe und des Weinbaues. Mitt Anthropol Ges Wien 41:283–296
Teira A (2010) Wild fruits, domesticated fruits. Archaeobotanical remains from the Roman saltworks at O Areal, Vigo (Galicia, Spain). In: Delhon C, Théry-Parisot I, Thiébault S (eds) Des Hommes et des plantes. Exploitation du milieu et gestion des ressources végétales de la préhistoire à nos jours. APDCA, Antibes, pp 199–207
Terral JF, Alonso N, Buxó R et al (2004) Historical biogeography of olive domestication (Olea europaea L.) as revealed by geometrical morphometry applied to biological and archaeological material. J Biogeogr 31:63–77
This P, Lacombe T, Thomas MR (2006) Historical origins and genetic diversity of wine grapes. Trends Genet 22:511–519
Ucchesu M, Orrù M, Grillo O, Venora G, Usai A, Serreli PF, Bacchetta G (2015) Earliest evidence of a primitive cultivar of Vitis vinifera L. during the Bronze Age in Sardinia (Italy). Veget Hist Archaeobot 24:587–600
Ucchesu M, Orrù M, Grillo O, Venora G, Paglietti G, Ardu A, Bacchetta G (2016) Predictive method for correct identification of archaeological charred grape seeds: support for advances in knowledge of grape domestication process. PLoS One 11:e0149814
Valamoti SM (2015) Harvesting the ‘wild’? Exploring the context of fruit and nut exploitation at Neolithic Dikili Tash, with special reference to wine. Veget Hist Archaeobot 24:35–46
Valamoti SM, Mangafa M, Koukouli-Chrysanthaki C, Malamidou D (2007) Grape-pressings from northern Greece: the earliest wine in the Aegean? Antiquity 81:54–61
Van Zeist W (1968) Prehistoric and early historic food plants in the Netherlands. Palaeohistoria 14:42–173
Vaquer J, Ruas M-P (2009) La grotte de l’Abeurador Félines-Minervois (Hérault): occupations humaines et environnement du Tardiglaciaire à l’Holocène. In: Barbaza M (ed) De Méditerranée et d’ailleurs … Melanges offerts à Jean Guilaine. Archives d’Ecologie Préhistorique, Centre d'Anthropologie des Sociétés Rurales, Toulouse, pp 761–792
Vera JC, Echevarría A (2013) Sistemas agrícolas del I milenio a.C. en el yacimiento de La Orden-Seminario de Huelva. Viticultura protohistórica a partir del análisis arqueológico de las huellas de cultivo. In: Celestino Pérez S, Blánquez Pérez J (eds) Patrimonio cultural de la vid y el vino. Universidad Autónoma de Madrid, Madrid, pp 95–106
Vera JC, Linares JA, Armenteros MJ, González D (2010) Depósitos de ídolos en el poblado de La Orden–Seminario de Huelva: espacios rituales en contexto habitacional. In: Cacho C, Maicas R, Galán E, Martos JA (eds) Los ojos que nunca se cierran. Ídolos en las primeras sociedades campesinas. Ministerio de Cultura, Madrid, pp 199–242
Wales N, Ramos Madrigal J, Cappellini E et al (2016) The limits and potential of paleogenomic techniques for reconstructing grapevine domestication. J Archaeol Sci 72:57–70
Weiss E (2015) “Beginnings of Fruit Growing in the Old World”—two generations later. Israel J Plant Sci 62:75–85
Weiss E, Wetterstrom W, Nadel D, Bar-Yosef O (2004) The broad spectrum revisited: evidence from plant remains. Proc Natl Acad Sci USA 101:9,551–9,555
Zamora JA (2000) La vid y el vino en Ugarit. Banco de datos filológicos semíticos noroccidentales. Monografías 6. CSIC, Madrid
Zohary D, Spiegel-Roy P (1975) Beginnings of fruit growing in the Old World. Science 187:319–327
Zohary D, Hopf M, Weiss E (2012) Domestication of plants in the Old World: the origin and spread of domesticated plants in Southwest Asia, Europe, and the Mediterranean Basin. Oxford University Press, Oxford
Acknowledgements
This paper is an output of the project FRUITFUL: La introducción y el desarrollo de la arboricultura en la Península Ibérica (HAR2015-64953-P) funded by the Spanish Ministerio de Economía y Competitividad within the Programa Estatal de Fomento de la Investigación Científica y Técnica de Excelencia, Subprograma Estatal de Generación del Conocimiento. GPJ work has been carried out within the postdoctoral contract FPDI-2013-16034 and IJCI-2015-25082 funded by the Spanish Ministerio de Economía y Competitividad.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by C. C. Bakels.
Rights and permissions
About this article

Cite this article
Pérez-Jordà, G., Peña-Chocarro, L., García Fernández, M. et al. The beginnings of fruit tree cultivation in the Iberian Peninsula: plant remains from the city of Huelva (southern Spain). Veget Hist Archaeobot 26, 527–538 (2017). https://doi.org/10.1007/s00334-017-0610-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-017-0610-6