
Abstract
The relationship between farming communities in south-eastern Europe and wild plant resources, fruit and nut trees in particular, is explored in this paper, based on charred plant remains from House 1 at late Neolithic Dikili Tash in eastern Macedonia, northern Greece, retrieved between 2010 and 2012. Within the rubble of a burnt destruction level dated to the second half of the 5th millennium cal BC, a wide range of cultivated crops like cereals, pulses and flax were stored together with a variety of fruit and nuts, such as acorns, wild pears, grapes, including grape pips and grape pressings and possibly figs, too. These finds provide a rare opportunity to investigate the use of fruit as well as the origins and context of wine making and consumption in the Neolithic of south-eastern Europe. Human interference with natural vegetation in relation to use of wild trees is discussed in light of the archaeobotanical, palynological and charcoal evidence from the wider area of the site. It is suggested that the remains from Dikili Tash may be pointing towards some early form of arboriculture in the region. The interplay of wild and domesticated plant resources encountered at the site is discussed within the framework of established oppositions between ‘wild’ and ‘domesticated’ in archaeological discourse. It is suggested that fruit and nut use at Dikili Tash might correspond to old traditions dating back to the hunter-gatherers of south-eastern Europe while wine, for which there is evidence at the site, might have acted as a mediator between human communities, cultivated landscapes and wild vegetation, inducing altered states of consciousness and cultivated/wild boundary transitions.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Archaeological investigation of Neolithic subsistence among village communities which flourished in south-eastern Europe from the 7th millennium bc onwards has focused heavily on the cultivation of domesticated crops (Marinova et al. 2013). Wild plant use, however, has received little attention, partly due to scarce finds of wild plants in the archaeobotanical records of these early farming communities. Moreover, an ‘ideological’ dichotomy between “agrios” (wild) and “domus” (domestic/ated) (Hodder 1990), has been considered as the underlying factor for the near absence of wild plants from early agricultural settlements of Greece (Perlès 2001). This dichotomous perception of the realm of the ‘wild’ (plants and animals) and the ‘domesticated’ in a process of domesticating nature and society (Hodder 1990) characterises archaeological discourse and is rooted in 19th century perceptions of culture versus nature (Ingold 1996, McNeely 2004). The antithetic couple between wild and domesticated forms of plants appears well established in botanical and cultural classifications since ancient Greek times, continuing into modern western thought: for ancient Greeks, civilisation was equivalent to cultivation and consumption of cooked food, while barbarism was defined by gathering from the wild and eating raw food (Vidal-Naquet 1983). Ample ethnographic evidence, however, suggests that this view was not universal or a constant in time and space (Ingold 1996), while a continuum has been proposed rather than a clear cut dichotomy between gathering wild plants and cultivating domesticated ones (Harris 1996). Ethnographic accounts are rich in examples that underline the problematic nature of such a dichotomous classification of nature and plants.
Archaeobotanical, ethnohistorical and ethnographic evidence shows that wild tree resources, nuts and acorns in particular, have constituted food staples for populations in different parts of the world such as in East Asia (Hosoya 2011), Latin America (Rival 1998) and among the Californian Indians (Anderson 2006). Recent hunter gatherers manage wild arboreal vegetation, encouraging the development and propagation of wild trees that they use (Rival 1998). Many ethnographic accounts underline the significant contribution of wild plants in recent farming communities (Ertuğ 2009; Forbes 1996). The management of patches or selected individual trees among wild vegetation might have increased yields for prehistoric hunter-gatherers of south-eastern Europe and the Near East, while the fig tree may represent the first domesticated plant (Kislev et al. 2006). In the Neolithic, a combined use of domesticated crops and harvested wild plants, including trees, is attested among early farming communities in south-western Asia and Europe (Fairbairn et al. 2007; Zapata 2000; Antolín Tutusaus 2013; Jacomet 2006, 2009; Borojevic 2009; Cunningham 2010, Marinova et al. 2013; Tolar et al. 2011). Despite the ideological dichotomy between wild and domesticated encountered in the archaeological literature (Hodder 1990), rather than being ostracised from early farming subsistence practices, gathering in the wild may have made a significant contribution to diet and symbolic/classificatory systems of early farming communities.
The view that wild plant resources were intentionally absent from early farming settlements of Greece has been questioned on the grounds of (a) inadequate archaeobotanical data and (b) cultural variability in plant classifications as wild or cultivated (Valamoti and Kotsakis 2007). For late Neolithic Greece, wild fruits and nuts, found in relative abundance, have been seen as a diversification strategy, increasing variety in diet and subsistence resources (Halstead 1994). However, the evidence from early and middle Neolithic contexts on wild plant use from northern Greek sites is extremely scarce (Valamoti and Kotsakis 2007), while biases affecting fruit recovery among charred assemblages render generalisations rather premature or even unfounded, regarding the deliberate, symbolic exclusion of wild plants from early sites (Perlès 2001) or the broadening of the subsistence base during the late and final Neolithic (Halstead 1994). Recent unpublished data from the 7th millennium bc site of Mavropigi-Fyllotsairi suggest that Cornus mas (Cornelian cherry) fruits were systematically harvested (Valamoti 2011). Studies of archaeobotanical assemblages with waterlogged preservation have clearly demonstrated that wild resources, including fruit, are underrepresented in the charred archaeobotanical record, so extra caution is needed in generalisations about the contribution of wild plant food resources in the prehistoric past (for a recent review, see Jacomet 2013). The bias introduced by preservation to what actually survives in terms of plant macro-remains is nicely illustrated by a case study whereby harvesting of wild garlic was demonstrated only by pollen analysis (Hadorn 1994) and this point has been raised in the literature since the early days of archaeobotanical inquiry (Willerding 1971; Dennell 1976).
Impressive finds from recent excavations at the late Neolithic site of Dikili Tash in northern Greece allow a detailed glimpse into harvests from beyond the fields: acorns, wild pears, grapes, blackberries and figs have been found within a Neolithic house. They provide the incentive for (a) a discussion of the status of these ‘wild’ fruit and nut harvests within the context of landscape use by subsistence farmers of south-eastern Europe and (b) a reconsideration of the wild/cultivated dichotomy characterising archaeobotanical discourse for the Neolithic.
The site investigated
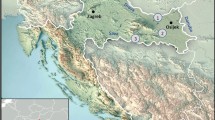
Dikili Tash is a multiperiod tell site, one of the largest in the Balkans, situated in the south-east part of the plain of Drama in eastern Macedonia, northern Greece, near the ancient city of Philippi (Fig. 1). Its habitation began in the 7th millennium bc, the early Neolithic, based on recent coring (Lespez et al. 2013), spanned the Neolithic and the Bronze Age and continued during Classical/Hellenistic, Roman and Byzantine times (Darcque et al. 2007). During the late Neolithic habitation of the site (approximately 4,500–4,000 cal BC), houses made of daub, usually comprising several compartments, were each equipped with a cooking facility, storage facilities, tools and vessels for storage, food serving and consumption. The latest group of houses was destroyed by fire during the second half of the 5th millennium bc, in the period called locally Late Neolithic II (or Chalcolithic or Final Neolithic). The Neolithic houses yielded a rich inventory of stored crops and harvests from the wild, in a very good state of charred preservation. There were stored concentrations of Triticum monococcum (einkorn) spikelets, Hordeum vulgare var. nudum (naked barley), Lens culinaris (lentil), Vicia ervilia (bitter vetch), Lathyrus sativus (grass pea), possibly Ficus carica (fig), Pyrus amygdaliformis (wild pear) and Quercus sp. (oak, acorns) (Table 1; Valamoti 2004).

Map showing location of the multi-period tell site of Dikili Tash in northern Greece
Materials and methods
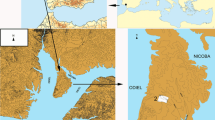
The archaeobotanical material discussed in this paper originates from one of these late Neolithic houses, House 1, excavated during the 2010 and 2012 seasons (Figs. 2, 3); the deposits have been radiocarbon dated to 4,350–4,250 cal BC (Maniatis et al. 2014). The focus on this house stems from the finds of large quantities of grape pips (2,460) and grape pressings (more than 300), consisting of empty, pressed grape skins that were excavated in 1989 (Valamoti et al. 2007). Lack of contextual information on these intriguing finds, retrieved from patches among the unexcavated collapsed house rubble, led to the systematic excavation and detailed archaeobotanical sampling of House 1 in recent years.

House 1 in relation to the other late Neolithic houses destroyed by fire at Dikili Tash

House 1 grid plan indicating location of studied samples and main features (brown ovens, red large circle pear concentration in pot; red small circles grape pip concentrations). (Color figure online)
Sampling strategy within House 1 and its surrounding area corresponds to near total sampling, as all undisturbed contexts have been sampled, while the burnt destruction layer has been sampled in its entirety. Sampling of the house interior was directed by the contexts that were being unearthed as excavation progressed, rather than an arbitrary grid. The aim was to obtain as detailed a picture as possible of the distribution of plant remains. Thus special features such as bins, ovens and pots are represented by individual samples comprising all the soil associated with them (Fig. 4a). When no obvious specific context was visible, all the excavated burnt destruction fill was taken as one or several soil samples, depending on the extent and depth of the deposit corresponding to each excavation unit. Excavation units were kept relatively small in size and each sample did not exceed 30 l, as it was considered preferable to have small individual samples rather than a single big sample from each excavation unit. When concentrations of seeds of a certain taxon were visible during excavation, special care was taken to observe the extent of each concentration and to sample it separately (Fig. 4b). When such concentrations extended over a large area or in depth, different individual samples were taken. The actual burnt and collapsed clay rubble blocks were not targeted for sampling for plant remains, as they have been kept separately for a specialised analysis currently under way. During 2010 and 2012, a total of 566 samples and 4,606 l of soil from the late Neolithic levels of House 1, covering approximately an area of 30 m2, were processed by flotation using a variant of the Ankara machine (French 1971) with a smallest sieve aperture of 300μ. Sorting of the flots for plant remains was carried out with the aid of a stereomicroscope and magnification 8×; for identification, magnifications of 8×–80× were used. This paper is based on preliminary results from these Neolithic samples collected between 2010 and 2012 which were scanned in order to evaluate the archaeobotanical contents of each. As the full study of all the Dikili Tash plant remains is currently in progress, here we present the remains from a selection of 23 samples, selected as representative of the range of taxa present and the concentrations indicated by the initial scanning of all samples and preliminary identifications of 153 samples which were rich in charred plant remains (Figs. 3, 5, ESM). Identification and quantification procedures follow Valamoti (2004), while plant names are given according to Flora Europaea (Tutin et al. 1964–1980).

House 1 at Dikili Tash; a collapsed wall rubble and pots in situ; acorns were found adjacent to the square pot; b Pyrus concentration overlapping a Lathyrus sativus concentration within the burnt destruction layer

Bar chart indicating sample composition; only taxa represented by at least 50 items are indicated. Pear fragments smaller than halves have not been included in quantification
Results
The archaeobotanical remains identified from House 1 (Figs. 3, 5) reveal the presence of pure, dense concentrations of hundreds or thousands of seeds of the following field crops with densities ranging from 1.7 to 1,317 identified items per litre (ESM): T. monococcum L. (einkorn), V. ervilia (L.) Willd. (bitter vetch), L. sativus L./L. cicera L. (grass pea) and Linum usitatissimum L. (flax). T. monococcum is probably represented by spikelets, as the einkorn samples contained grain, spikelet forks and glume bases. Hordeum sp. (barley) dominated one sample, albeit in relatively small numbers while Lens sp. (lentil) has been found in small quantities in some of the samples. Since more areas within House 1 were sampled in 2013, it is possible that these taxa were also present in large numbers inside this house. Indeed this seems to be the case as a dense, rich concentration of lentils has been identified in the 2013 samples. Some samples of mixed composition probably represent areas between the crop concentrations. The crops, in their vast majority were largely clean of weed seeds. Cladium mariscus (L.) Pohl occurred in some of the samples.
Besides the cultivated crop concentrations identified within House 1, a variety of harvests of fruit and nuts has emerged. These include the remains of Quercus sp. (acorns), V. vinifera L. (grapes), cf. P. amygdaliformis (wild pear), F. carica L. (fig) and Rubus fruticosus L. agg. (blackberry). Pyrus was identified on the basis of the rounded cellular pattern of the seed surface which distinguishes it from Malus (apple), this pattern, however, was not observed on all seeds. The seeds were pointed on top, unlike Sorbus seeds which are curved (René Cappers personal communication). Moreover, Sorbus seeds are reported as having elongated cell structures (Lucy Kubiak-Martens personal communication), thus on these grounds they appear as less likely candidates for the archaeobotanical finds, although more reference material needs to be examined in light of the variability shown on seed surface patterns among the different Sorbus species. Of these taxa, Quercus, Vitis and Pyrus have been identified in rich, dense concentrations. Most impressive are the Vitis and Pyrus finds. The Vitis finds consist of pips and skins, either loose or attached to the pips (Fig. 6a, b). Approximately 2,500 pips have been retrieved from the 2010 to 2012 excavation seasons and this number is expected to rise when all samples from House 1 have been fully studied. A dense concentration of pips has been associated with large pottery fragments of a coarse jar (Fig. 7a, c). Yet, grape pips and skins were spread in the form of a ‘carpet’ over a larger area to the northwest of the clay jar, covering an area of approximately 2–3 m2. Not all samples rich in grape pips contained grape skins, while the pips in some of the samples were characterised by a very peculiar morphology, the pip having a ‘sucked out’ appearance (Fig. 6b). Possible depositional factors that might have affected grape-pip preservation, for example, charring inside a liquid, possibly must or wine, could have caused such malformation. This issue is currently under investigation, combining experimental work, SEM micrographs of the archaeobotanical and experimentally treated modern pips together with contextual information and spatial distribution of the grape pips within House 1. Another rich concentration of grape pips, again contained in a pot, was found further north in 2013 and is currently under study (Fig. 3).

a Grape pressings; b malformed pips from Dikili Tash, House 1

House 1 at Dikili Tash, a location of grape pip concentration within and around a coarse ware pot; tartaric acid was detected in this pot; b detail of pip layer; c pot with hole near its base containing wild pears
Pyrus has been found in at least two, well defined, rich concentrations (Figs. 3, 5). One of them, to the north of the L. sativus finds, covered a relatively large area of approximately 3–4 m2 and Pyrus was spread in a horizontal fashion, overlapping a L. sativus concentration in parts (Fig. 4b). A fragment of clay with a fine mat impression found in association with these Pyrus finds might correspond to the remains of a container. The second well defined Pyrus concentration was associated with a jar made of coarse ware with a hole near its base (Fig. 7c). At least 630 whole pears, 75 of which had the stems still attached, were found charred between the smashed sherds of the pot (Fig. 8a–c). In both cases, some of the pears had a wrinkly outer surface.

Charred wild pears contained in pot; a whole pears; b whole with stalks and c seeds
Acorns were found associated with an area of pots lying underneath a collapsed clay wall fragment, to the southeast of a concentration of pots (Fig. 4a). R. fruticosus has been found in a few samples, represented by drupes and stones, in relatively small numbers. Likewise, whole figs have not been found as a pure concentration as yet, but were dispersed mainly in samples to the north part of House 1. Ficus seeds were also found in the samples, albeit in small numbers.
Discussion
Harvests from the fields and beyond: stored products inside House 1
A variety of cultivated plants and harvests from wild plants growing beyond the fields were stored inside House 1, in a fully processed state, ready for consumption or for further processing for consumption.
At least T. monococcum, in the form of clean spikelets and probably Hordeum were the cereals stored inside House 1 at the time of destruction. Dehusking of einkorn spikelets could have been carried out piecemeal, depending on the regular needs of the household. Storage in the spikelet, a practice encountered in northern Greece, both in the Neolithic and the Bronze Age (Jones 1987; Valamoti 2004), reduces stored quantity of grain but is beneficial in other respects, such as the protection of the grain from rodents, insects and mould (Sigaut 1988). L. sativus and V. ervilia, stored inside House 1, were largely clean of weed seeds with the exception in some cases of a few Polygonum and Galium/Asperula seeds. Thus, besides T. monococcum, pulses, too, were fully processed prior to consumption and they would have required only some hand cleaning. V. ervilia and L. sativus would have probably needed detoxification, with the aid of water and/or testa removal prior to consumption (Butler et al. 1999; Valamoti et al. 2011). Lens was also stored inside the house adjacent to a second rich grape pip concentration to the north of the oven, both revealed in 2013 (6618-002, Fig. 3). Some of these crop concentrations have been associated with ceramic containers as is the case with bitter vetch and grape. Storage is also implied by the purity and density of the finds and those crops not contained inside pots were probably stored in perishable containers, for example wooden chests or cloth bags.
Acorns have been found as cotyledons, again stored in some perishable container. It is not possible to tell whether they had been further processed, for example by roasting prior to storage, a process that seems to improve storage (Cunningham 2010). Tannin removal from acorns would have been necessary if they were to be consumed as human food, unless selected trees bearing sweet acorns were being harvested (Mason 1995; Psilakis and Psilakis 2001; Mason and Nesbitt 2009; Antolín-Tutusaus 2013).
Wild pears could have been dried and then stored in the pot with the hole which contained them, the hole perhaps facilitating ventilation and preventing mould development or related to a former use of the pot, unrelated to the pears. The wrinkly surface observed in several of the pears may be indicative of dried fruit. Alternatively, the pears might have been preserved by lactic acid fermentation in brine (water, salt and/or vinegar), stored therefore as ‘pickled’ fruit, something which might have affected the structure of the fruit and which might be detectable in the archaeobotanical remains. We plan to pursue this in the future through experimental and SEM examination. Pickled fruits of P. amygdaliformis are recorded in Balikesir and the island of Bozcaada/Tenedos in north-western Turkey and the north-east Aegean respectively, used as food and medicine for lowering blood sugar levels (Emre-Bulut and Tuzlacı 2008; Arıtuluk and Ezer 2012). The other Pyrus concentration may correspond to fruit in the process of being dried or stored dry in some form of shallow, flat container of perishable materials, again preventing mould development or, already dry they might have been stored inside a bag hanging from the roof, the contents of which might have spread on to the floor upon destruction.
In contrast to the plant storage evidence from House 3 at Dikili Tash (Table 1; Fig. 2), House 1 stands out by its wealth of finds, partly at least reflecting the detailed and systematic sampling followed in the recent excavation. Although finds of grape pips and skins are reported from House 3, House 1 has yielded the greatest variety especially as regards wild plant food resources.
Besides the storage of clean grain, fruit and nuts, the storage of some grape product in House 1 is indicated by the numerous grape pips, grape skins and grape pressings.
Neolithic wine
The extraction of grape juice and the preparation of some form of grape juice product, perhaps wine, had been suggested nearly a decade ago, when a concentration of grape pips and grape skins were identified from House 1 in Dikili Tash, albeit devoid of any contextual information, other than that it originated from within the rubble of yet another burnt Neolithic House on the site (Valamoti et al. 2007). In the recent excavation seasons of 2010–2013 the context of this grape find has been excavated and several of its aspects form part of an ongoing investigation. The grape pip remains were numerous and spread over a relatively large area. Within House 1, a number of jars and cups have been excavated (Fig. 9). Moreover, the grape remains were associated with a coarse-ware clay pot from which two sherds were analysed by N. Garnier (SAS Laboratoire Nicolas Garnier, Vic le Comte, France). The results showed the presence of tartaric acid (N. Garnier unpublished report and personal communication). This suggests that the grape pips and skins were contained in at least one coarse-ware pot together with the grape juice; upon destruction by fire and collapse of the house, the pot was broken and its contents spread over a large area due to the liquid that was presumably contained within. Although tartaric acid is indicative of a wide range of grape products, for example vinegar, wine, must or grape syrup, the co-occurrence of grape juice and grape pressings in the same container make a very strong case for fermentation, and thus wine in the process of being made, which was interrupted by the destruction of the house by fire.

Clay cup extracted from House 1 at Dikili Tash, northern Greece
The grape pips from the same context, excavated in 1989, have been identified as morphologically wild (Mangafa and Kotsakis 1996). The recent finds are currently being studied using morphometric analysis (Terral et al. 2010) by Clémence Pagnoux in order to attempt to further explore the status of these Dikili Tash grape vines in relation to cultivation and domestication (Pagnoux et al. 2014). Yet, as has previously been argued, these finds of morphologically wild grape pips do not necessarily imply that they were gathered from grapevines growing in the wild (Valamoti 1998, 2009; Valamoti et al. 2007). Grape vines around Dikili Tash could have been tended, managed, husbanded, or even cultivated, despite their morphological resemblance to wild ones. Indeed, modern, cultivated V. vinifera varieties exist with morphologically wild pips, using Stummer’s (1911) breadth/length indices, such as Pinot Noir (Rivera-Nuñez and Walker 1989). Of course, it is equally plausible that wild grapes were gathered for wine making. The presence of Vitis wood, however, in the charcoal assemblage from the site (Stéphanie Thiébault personal communication) may be a further indication of management and cultivation, following Miller’s suggestion that propagating selected plants yielding sweet grapes was the incentive towards cultivation and domestication, with pruning being one of the means of tending these grapevines (Miller 2008). Thus, House 1 at Dikili Tash, besides the rich harvests from fields and beyond, has provided the earliest evidence for wine making in the eastern Mediterranean, the Aegean and Europe so far.
Vitis vinifera is not particular to Dikili Tash as it has been found elsewhere in Neolithic northern Greece, especially in the late Neolithic, though in Neolithic Bulgaria only sporadic finds of single pips are reported (Marinova and Valamoti 2014). Yet, the richness of the Dikili Tash find, together with the so far unique indication for wine-making in south-eastern Europe and the eastern Mediterranean, call for a closer examination of the particular circumstances generating this find and its wider regional context. The 5th millennium finds from Dikili Tash are later than those identified at Hadji Firuz tepe in Iran dated to the 6th millennium bc (McGovern et al. 1996), but roughly contemporary with those from Areni cave in Armenia (Barnard et al. 2011). Yet, the signs of wine making at Dikili Tash come from several lines of evidence and not mainly pottery residue analysis. In addition to the identification of tartaric acid at Dikili Tash, grape juice is also indicated by the rich archaeobotanical finds of grape pips and grape pressings, while fermentation, serving and consumption of a liquid are implied by the association of these finds with a large clay container and ceramic serving and drinking vessels. These finds suggest that wine making could have started in different areas within the wider geographical range of the wild grape vine, rather than in a single region (Valamoti et al. 2007). Moreover, although viticulture is not a prerequisite for wine making, the Dikili Tash grape finds could have resulted from some form of incipient cultivation or management of wild grapevines or cultivated ones still bearing morphologically wild-type pips. Of course one cannot entirely rule out the possibility that knowledge of wine making or grapevine tending or cultivation spread via contact networks of the Neolithic.
Harvesting the wild at Neolithic Dikili Tash?
A house full of field crops harvested in the summer months with fruit and nuts harvested during the autumn months, together with strong indications of fermentation being underway when fire broke out, suggests that the destruction of House 1 at Dikili Tash occurred some time in late autumn or winter, depending on the time of harvest and duration of fermentation of the grape juice together with the grape pressings. House 1 may stand out for its wealth and variety of wild fruits and nuts, while finds of jewellery and golden beads (Chronique de fouilles en ligne 2012), in association with the indications of wine making may indicate some special function or position of this particular house within the settlement. A series of questions concerning socio-economic organisation and emerging hierarchy among the family groups occupying the Neolithic houses is currently under investigation. What is certain from the plant remains is the intensive use of the ‘wild’ non-arable vegetation surrounding the site. Do the finds from Dikili Tash suggest that trees interspersed with fields belonged to the realm of the cultivated land? Was there a visual and conceptual continuum from sown fields to tended thickets of trees including grape vines? Or, by contrast, were there particular patches of managed wild vegetation where these trees grew, perceived as ‘margins’ marking boundaries and a threshold to woodland proper?
Whatever the people–plant relationship in these early periods, the Dikili Tash finds suggest intensive use of the ‘wild’ or even some form of early fruit and nut tending or incipient cultivation, during the second half of the 5th millennium bc.
Dense mixed oak woodland appears to have dominated the Drama plain in the Neolithic (Bottema and Woldring 1990; Greig and Turner 1974). Riparian environments in the vicinity of the settlement are also indicated, and Vitis yielding the thousands of pips would have probably grown there. P. amygdaliformis, a wild fruit tree, would probably have grown in patches of managed open woodland vegetation or clearings or even within crop fields near the settlement, as it requires well drained soils with open or cleared vegetation, with ample light in order to yield fruit. Fig trees also require open, sunny spots, and would have flourished both in riverside environments as well as dry areas with underground water. The oak trees that yielded the stored acorns within House 1 might have been trees selected for their sweet acorns or their size or both.
Based on the archaeobotanical finds from Dikili Tash, natural woodland surrounding the settlement would have been intensively managed and transformed into cleared patches of crop fields, as well as patches where wild fruit trees were encouraged or even cultivated in some cases. Such an intensive interference with woodland vegetation in the vicinity of Dikili Tash, although of a scale undetectable by regional pollen diagrams, seems to be shown in recent environmental proxies from the periphery of the settlement (Glais et al. 2013; Laurent Lespez personal communication), the details of which are currently under investigation.
Among Neolithic farming societies of northern Greece, crop fields would have required constant tending, with soil preparation, manuring, weeding, sowing etc. (Halstead 2000). The relationship with parts of the landscape comprising fruit and nut yielding wild trees, is more obscure. Wild trees might have required no, little or a different form of attention compared to fields and crops. Plants are incorporated in classificatory systems of societies (Levi-Strauss 1962), thus on a symbolic level, areas with wild vegetation might have been associated with particular notions structuring society at late Neolithic Dikili Tash. For Neolithic farmers, fruit and nut trees and the areas where these grew might have corresponded to the realm of a distant, collective hunter-gatherer past. These field crops and harvests from a wild or semi-natural vegetation around the settlement, besides providing diversification in diet and reliable supplies for survival of the Neolithic household, on a symbolic level they may have somehow corresponded to perceptions of nature and culture ambivalent, contradictory but ultimately reconciled once housed under the same roof. House 1 contains harvests from both realms which are brought together in space and time by a ‘co-habitation’ of the wild and cultivated. Wild and cultivated plants might, on a symbolic level, refer to distant ancestors, hunter-gatherers and the first farmers of the region respectively. Of course these plants constituted food but at the same time they were probably involved in classifications and perceptions related to different parts of the landscape, to different relationships between plants and people at Dikili Tash, seasonal modes and tasks, symbolism, food taboos and gender relations. The harvested fields were the focus of late spring/early summer, the ‘wilderness’, perhaps managed, was the focus of late summer and autumn.
Wine, identified in House 1 takes us a step further in this discourse: in this Neolithic house, a fruit harvest from beyond the crop fields was transformed into an intoxicating substance, probably served with specially designed decanters and consumed in small cups. Its consumption would have led to the experiencing of particular states of consciousness and it might have prescribed specific rules of participation and etiquette of consumption. Patches of woodland and the realm of what could be considered as ‘wild’ was the source of a special liquid usually associated with feasting, solidarity, hierarchy and ritual. Symbolic aspects of life, in particular in relation to this liquid, remain largely unknown for Neolithic Dikili Tash and its region as we lack the rich literary and pictorial evidence of later periods: for ancient Greeks, wild vegetation, fruit trees and wine were closely associated with uncooked food including cannibalism, ecstatic behaviour, rage and sexuality, all elements of an ambivalent god, Dionysos, breaching the wild with culture through wine, his gift to mankind, a produce strongly related to civilisation, social status and institutionalised celebrations (Schlesier 2011). At Dikili Tash, in the cold days of winter, a celebration perhaps of communal character might have revolved around the village’s new wine, served in the cups found inside House 1. Such practices, rooted in the Neolithic, might have contributed to the consideration of the wider area where Dikili Tash belonged in antiquity, ancient Thrace, as one of the birth places of Dionysos.
Conclusions
The preliminary results from the rich archaeobotanical finds from inside the burnt rubble of House 1 at Dikili Tash, after its destruction by fire, indicate the storage, under the same roof, of several crops as well as plants gathered from beyond the fields, corresponding either to parts of the wild vegetation growing in the vicinity of the house, or from managed parts of wild vegetation around the settlement. Exceptionally good preservation conditions and systematic sampling at Dikili Tash have shown that trees were an important food resource at the end of the Neolithic in the region and probably in south-eastern Europe as a whole. In this large amount of fruit remains found at the site, we may be facing the evolution of gathering from trees to early fruit growing, including viticulture, in the region. Wine making and viticulture may have been begun independently in various regions, the northern Aegean being one of them.
Irrespective of whether these ‘harvests from the wild’ were perceived as such by the Neolithic inhabitants of Dikili Tash, they were ‘tamed’ by being housed under the same roof, and in the case of grapes, transformed into an intoxicating liquid, the consumption of which might have opened access to the ‘wild’ in a symbolic sense.
The charred plant remains, corroborated by archaeological context and pottery residue analysis indicate that wine making was under way in this Neolithic house when fire broke out, encapsulating this process sometime in late autumn to early winter. The charred seeds from Dikili Tash are eloquent witnesses to daily strife of the past as well as of special days with special foods and certainly drinks consumed, leading to particular states of consciousness being experienced. The details of these Neolithic festive occasions are of course long gone, together with the consumers of the crops and wild harvests.
Questions concerning the organisation of plant food production, the role of wine making and consumption in relation to intra-settlement social differentiation, hierarchy and/or solidarity await further research of the architectural, artefactual and bioarchaeological finds from Dikili Tash. Beyond subsistence practices within Neolithic households in south-eastern Europe, the Dikili Tash archaeobotanical finds provide challenging food for thought on Neolithic landscapes shaped by human activities and notions about plants and people.
References
Anderson Kat M (2006) Tending the wild: Native American knowledge and the management of California’s natural resources. University of California Press, Berkeley
Antolín Tutusaus F (2013) Of cereals, poppy, acorns and hazelnuts. Plant economy among early farmers (5400–2300 cal. bc) in the NE of the Iberian Peninsula. An archaeobotanical approach. Universitat Autònoma de Barcelona, http://hdl.handle.net/10803/128997
Arıtuluk ZC, Ezer N (2012) Halk Arasında Diyabete Karşı Kullanılan Bitkiler (Turkiye)-II (Use of anti-diabetic plants in Turkish folk medicine-II, in Turkish). Hacettepe Üniversitesi Eczacılık Fakültesi Dergisi 32, Ankara, pp 179–208
Barnard H, Dooley AN, Areshian G, Gasparyan B, Faull KF (2011) Chemical evidence for wine production around 4000 BCE in the Late Chalcolithic Near Eastern highlands. J Archaeol Sci 38:977–984
Borojevic K (2009) Water chestnuts (Trapa natans L.) as controversial plants: botanical, ethno-historical and archaeological evidence. In: Fairbairn A, Weiss E (eds) From Foragers to farmers. Oxbow Books, Oxford, pp 86–97
Bottema S, Woldring H (1990) Anthropogenic indicators in the pollen record of the Eastern Mediterranean. In: Bottema S, Entjes-Nieborg G, Van Zeist W (eds) Man’s role in the shaping of the eastern Mediterranean landscape. Balkema, Rotterdam, pp 231–264
Butler A, Tesfay Z, D’Andrea C, Lyons D (1999) The ethnobotany of Lathyrus sativus L. in the highlands of Ethiopia. In: Van der Veen M (ed) The exploitation of plant resources in ancient Africa. Kluwer/Plenum, New York, pp 123–136
Chronique de fouilles en ligne (2012) http://chronique.efa.gr/index.php/fiches/voir/3406/
Cunningham P (2010) Cache or carry: food storage in prehistoric Europe. In: Millson D (ed) Replication and interpretation: the use of experimental archaeology in the study of the past. Oxbow Books, Oxford, pp 7–28
Darcque P, Koukouli-Chryssanthaki H, Malamidou D, Treuil R, Tsirtsoni Z (2007) Recent researches at the Neolithic settlement of Dikili Tash, Eastern Macedonia, Greece: an overview. In: Todorova H, Stefanovich M, Ivanov G (eds) The Struma/Strymon river valley in prehistory. Proceedings of the International Symposium ‘Strymon Praehistoricus’, Kjustendil-Blagoevgrad (Bulgaria) and Serres-Amphipolis (Greece), 2004. Gerda Henkel Stiftung, Düsseldorf, pp 247–256
Dennell RW (1976) The economic importance of plant resources represented on archaeological sites. J Archaeol Sci 3:229–247
Emre-Bulut G, Tuzlacı E (2008) Bozcaada’nın halk ilac olarak kullanılan bitkileri (Uses of medicines on Bozcaada, in Turkish). In: Akdemir A, Demircan O, Yılmaz S, Takaoglu T, Erginal E (eds) Bozcaada Değerleri Sempozyumu 2008. Aynalı Pazar, Çanakkale
Ertuğ F (2009) Wild plant foods: routine dietary supplements or famine foods? In: Fairbairn A, Weiss E (eds) From foragers to farmers. Oxbow Books, Oxford, pp 64–70
Fairbairn A, Martinoli D, Butler A, Hillman G (2007) Wild plant seed storage at Neolithic Çatalhöyük East, Turkey. Veget Hist Archaeobot 16:467–479
Forbes H (1996) The uses of the uncultivated landscape in modern Greece: a pointer to the value of the wilderness in antiquity? In: Salmon J, Shipley G (eds) Human landscapes in classical antiquity: environment and culture. (Leicester–Nottingham studies in ancient society 6). Routledge, London, pp 68–97
French DH (1971) An experiment in water-sieving. Anatol Stud 21:59–64
Glais A, Lespez L, Lopez-Saez JA (2013) The environment of tell of Dikili Tash: anthropogenic and climatic changes. In: 8th international conference on geomorphology (IAG) 2013, abstracts, p 423
Greig JRA, Turner J (1974) Some pollen diagrams from Greece and their archaeological significance. J Archaeol Sci 1:177–194
Hadorn P (1994) Saint-Blaise/Bains des Dames 1: Palynologie d’un site néolithique et histoire de la végétation des derniers 16000 ans. Musée Cantonal d’Archéologie, Neuchâtel
Halstead P (1994) The north-south divide: regional paths to complexity in prehistoric Greece. In: Mathers C, Stoddart S (eds) Development and decline in the Mediterranean Bronze Age. (Sheffield Archaeological Monographs 8), J.R.Collis Publications Sheffield, pp 195–219
Halstead P (2000) Land use in postglacial Greece: cultural causes and environmental effects. In: Halstead P, Frederick C (eds) Landscape and land use in postglacial Greece. (Sheffield Studies in Aegean Archaeology 3). Oxbow, Oxford, pp 110–128
Harris DR (1996) Introduction: themes and concepts in the study of early agriculture. In: Harris DR (ed) The origins and spread of agriculture and pastoralism in Eurasia. University College Press, London, pp 1–9
Hodder I (1990) The domestication of Europe: structure and contingency in Neolithic societies. Blackwell, Oxford
Hosoya LA (2011) Staple or famine food? Ethnographic and archaeological approaches to nut processing in East Asian prehistory. Archaeol Anthropol Sci 3:7–17
Ingold T (1996) Growing plants and raising animals: an anthropological perspective on domestication. In: Harris DR (ed) The origins and spread of agriculture and pastoralism in Eurasia. University College Press, London, pp 12–24
Jacomet S (2006) Plant economy of the northern alpine lake dwelling area: 3500–2400 bc cal. In: Karg S, Baumeister R, Schlichtherle H, Robinson DE (eds) Economic and environmental changes during the 4th and 3rd millennia bc. Environ Archaeol 11: 64–83
Jacomet S (2009) Plant economies and village life in Neolithic lake dwellings at the time of the Alpine Iceman. Veget Hist Archaeobot 18:47–59
Jacomet S (2013) Archaeobotany: analyses of plant remains from waterlogged archaeological sites. In: Menotti F, O’Sullivan A (eds) The Oxford handbook of wetland archaeology. Oxford University Press, Oxford, pp 497–514
Jones G (1987) Agricultural practice in Greek prehistory. Ann Br School Athens 82:115–123
Kislev ME, Hartmann A, Bar-Yosef O (2006) Early domesticated fig in the Jordan valley. Science 312:1,372–1,374
Lespez L, Tsirtsoni Z, Darcque P, Koukouli-Chryssanthaki H, Malamidou D, Treuil R, Kourtessi-Philippakis G, Davidson R, Oberlin C (2013) The lowest levels at Dikili Tash, Northern Greece: a missing link in the Early Neolithic of Europe. Antiquity 87:30–45
Levi-Strauss C (1962) La Pensée Sauvage. Paris, Plon (Greek Translation 1977, Papazisis Publications, Athens)
Mangafa M, Kotsakis K (1996) A new method for the identification of wild and cultivated charred grape seeds. J Archaeol Sci 23:409–418
Maniatis Y, Tsirtsoni Z, Oberlin C, Darcque P, Koukouli-Chryssanthaki H, Malamidou D, Siros T, Miteletsis M, Papadopoulos S, Kromer B (2014) New 14C evidence for the Late Neolithic-Early Bronze Age transition in Southeast Europe. In Tykot RH (ed), Proceedings of the 38th International Symposium on Archaeometry, May 10th–14th 2010, Tampa, Florida, Open Journal of Archaeometry 2: 5,262
Marinova E, Valamoti SM (2014) Crop diversity and choice in prehistoric southeastern Europe: cultural and environmental factors shaping the archaeobotanical record of northern Greece and Bulgaria. In: Chevalier A, Marinova E, Peña-Chocarro L (eds) Plants and people: choices and diversity through time. Oxbow Books, Oxford, pp 46–54 and 92–102
Marinova EM, Filipović D, Obradović D, Allué E (2013) Wild plant resources and land use in the Mesolithic and Early Neolithic South-East Europe: archaeobotanical evidence from the Danube catchment of Bulgaria and Serbia. Offa 69(70):467–478
Mason S (1995) Acornutopia? Determining the role of acorns in past human subsistence. In: Wilkins J, Harvey D, Dobson M (eds) Food in antiquity. University of Exeter Press, Exeter, pp 12–23
Mason S, Nesbitt M (2009) Acorns as food in Southeast Turkey: implications for prehistoric subsistence in Southwest Asia. In: Fairbairn A, Weiss E (eds) From foragers to farmers: papers in honour of Gordon Hillman. Oxbow, Oxford, pp 71–85
McGovern P, Glusker DL, Exner LJ, Voight MM (1996) Neolithic resinated wine. Nature 381:480–481
McNeely JA (2004) Nature versus nurture: managing relationships between forests, agroforestry and wild biodiversity. Agrofor Syst 61–62:155–165
Miller NF (2008) Sweeter than wine? The use of the grape in early western Asia. Antiquity 82:937–946
Pagnoux C, Bouby L, Ivorra S, Petit Ch, Valamoti S-M, Pastor T, Picq S, Terral JF (2014) Inferring the agrobiodiversity of Vitis vinifera L. (grapevine) in ancient Greece by comparative shape analysis of archaeological and modern seeds. Veget Hist Archaeobot. doi10.1007/s00334-014-0482-y
Perlès C (2001) The early Neolithic of Greece. Cambridge University Press, Cambridge
Psilakis N, Psilaki M (2001) Το Ψωμί των Ελλήνων (The bread of the Greeks, in Greek). Karmanor Publications, Herakleion
Rival L (1998) Domestication as a historical and symbolic process: wild gardens and cultivated forests in the Equatorial Amazon. In: Balée WL (ed) Advances in historical ecology. Columbia University Press, New York, pp 232–250
Rivera Nunez D, Walker MJ (1989) A review of palaeobotanical findings of early Vitis in the Mediterranean and of the origins of cultivated grape-vines, with special reference to new pointers to prehistoric exploitation in the western Mediterranean. Rev Palaeobot Palynol 61:205–237
Schlesier H (2011) A different god? Dionysos and ancient polytheism. Walter de Gruyter, Berlin
Sigaut F (1988) A method for identifying grain storage techniques and its application for European agricultural history. Tools Tillage 7:3–32
Stummer A (1911) Zur Urgeschichte der Rebe und des Weinbaus. Mitt Anthropol Ges Wien 41:283–296
Terral JF, Tabard E, Bouby L, Ivorra S, Pastor T, Figueiral I, Picq S, Chevance JB, Jung C, Fabre L, Tardy C, Compan M, Bacilieri R, Lacombe T, This P (2010) Evolution and history of grapevine (Vitis vinifera) under domestication: new morphometric perspectives to understand seed domestication syndrome and reveal origins of ancient European cultivars. Ann Bot 105:443–455
Tolar T, Jacomet S, Velušček A, Čufar K (2011) Plant economy at a late Neolithic lake dwelling site in Slovenia at the time of the Alpine Iceman. Veget Hist Archaeobot 20:207–222
Tutin TG et al (1964–1980) Flora Europaea vols 1–5. Cambridge University Press, Cambridge
Valamoti SM (1998) Η εκμετάλλευση της αμπέλου στο χώρο της Μακεδονίας και της Θράκης κατά τη Νεολιθική και την Πρώιμη Εποχή του Χαλκού (The exploitation of the vine in Neolithic and early Bronze Age Macedonia and Thrace, in Greek). Αμπελοοινική Ιστορία στο χώρο της Μακεδονίας και της Θράκης. Ε.Τ.Β.Α. Foundation, Athens, pp 137–149
Valamoti S (2004) Plants and people in Late Neolithic and Early Bronze Age northern Greece. An archaeobotanical investigation. (BAR International Series 1258). Archaeopress, Oxford
Valamoti SM (2009) Plant food ingredients and ‘recipes’ from prehistoric Greece: the archaeobotanical evidence. In: Morel JP, Mercuri AM (eds) Plants and culture: Seeds of the cultural heritage of Europe. Centro Europeo per i Beni Culturali Ravello, Edipuglia, Bari, pp 25–38
Valamoti SM (2011) Σπόροι για τους νεκρούς; Αρχαιοβοτανικά δεδομένα από τη Μαυροπηγή Κοζάνης, θέση Φυλλοτσαΐρι (Seed for the dead? Archaeobotanical remains from Mavropigi-Fyllotsairi in Kozani, in Greek). Proceedings of the 1st Archaeological Conference on Archaeological Investigations in Upper Macedonia. Kozani, Archaeological Museum of Aiani, pp 245–256
Valamoti SM, Kotsakis K (2007) Transitions to agriculture in the Aegean: the archaeobotanical evidence. In: Colledge S, Conolly J (eds) The origin and spread of domestic plants in SW Asia and Europe. Left Coast Press, Walnut Creek, University College London Institute of Archaeology Publications, London, pp 75–92
Valamoti SM, Mangafa M, Koukouli-Chrysanthaki H, Malamidou D (2007) Grape-pressings from northern Greece: the earliest wine in the Aegean? Antiquity 81:54–61
Valamoti SM, Moniaki K, Karathanou A (2011) An investigation of processing and consumption of pulses among prehistoric societies: archaeobotanical, experimental and ethnographic evidence from Greece. Veget Hist Archaeobot 20:381–396
Vidal-Naquet P (1983) Le Chasseur Noir (Greek translation). Livanis-Nea Synora Publications, Athens
Willerding U (1971) Methodische Probleme bei der Untersuchung und Auswertung von Pflanzenfunden in vor- und frühgeschichtlichen Siedlungen. Nachr Nieders Urgesch 40:180–198
Zapata L (2000) La recolección de plantas silvestres en la subsistencia mesolítica y neolítica. Datos Arqueobotánicos del País Vasco. Complutum 11:157–169
Acknowledgments
The Dikili Tash excavation program is run under the auspices of the École française d’Athènes (EFA) and the Athens Archaeological Society. I am deeply grateful to the directors of the excavation, Pascal Darcque, Chaido Koukouli-Chrysanthaki, Dimitra Malamidou and Zoï Tsirtsoni for entrusting me with the study of the Dikili Tash material and for generously providing every possible help, in the field and during study and preparation of this paper. Nicolas Garnier kindly provided unpublished information on his residue analysis of the Dikili Tash pottery fragments. Stephanie Thiébault kindly provided unpublished information on the presence of Vitis wood at Dikili Tash. Laurent Lespez generously shared unpublished information on current palynological investigations in the vicinity of Dikili Tash. Special thanks go to a large number of students of the Department of Archaeology of the Aristotle University of Thessaloniki who provided their help in the field and in the laboratory between 2010 and 2012, in particular our postgraduate students Angeliki Karathanou, Martha Kokkidou, Chryssa Petridou, Eleni Telioridou and Aimilia Kokonaki. Chryssa Petridou provided invaluable help in the study of the plant remains presented here. Natalia Alonso, Ferran Antolín-Tutusaus, René Cappers, Lucy Kubiak Martens and Hans-Peter Stika provided help towards Pyrus amygdaliformis identification. Tassos Bekiaris and Nikos Katsikaridis helped with Fig. 8a, b. Hüseyin Çınar Öztürk and Burcin Erdoğu facilitated comprehension of the Turkish literature. I am also grateful to Stephanie Jacomet and an anonymous reviewer for their constructive comments on an earlier draft of this manuscript. I wish to gratefully acknowledge the financial support of the archaeobotanical work at Dikili Tash, largely provided by the National Geographic Society with further financial help from the French Ministry of Foreign Affairs, the Institute for Aegean Prehistory (INSTAP) and the École française d’Athènes.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by: F. Bittmann.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Valamoti, S.M. Harvesting the ‘wild’? Exploring the context of fruit and nut exploitation at Neolithic Dikili Tash, with special reference to wine. Veget Hist Archaeobot 24, 35–46 (2015). https://doi.org/10.1007/s00334-014-0487-6
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-014-0487-6