
Abstract
The first open-sea (non-coastal) Marine Protected Area (MPA) in Argentina was created in 2013 (and named “Namuncurá”), at Burdwood Bank, an undersea plateau located about 200 km south from Malvinas/Falkland Islands, SW Atlantic Ocean. This contribution represents the most recent and complete checklist of benthic organisms (mainly mega and macrofauna) that summarized 240 taxa collected at three sites in the mentioned area. The benthic richness here documented largely exceeds the values coming from other productive areas in Argentinean waters. Faunistic composition was different among the sites, although several species were shared. Bryozoans presented the highest richness compared to the other groups, followed by mollusks and sponges. These data represent valuable information and a very helpful starting point for incoming research, conservation efforts and management. It is essential to establish the spatial distribution of indicator taxa, like corals, sponges and bryozoans, to better attempt at the protection of benthic biodiversity in this rich Argentinean marine system. This is a key issue considering the notable extension of the MPA compared to other areas in Argentina, the bathymetric variations, particular oceanographic conditions and different fishing circumstances within the area.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
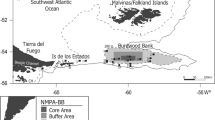
The Burdwood Bank (BB) is an undersea plateau located about 200 km south from Malvinas/Falkland Islands and 150 km east from Isla de los Estados (Tierra del Fuego Province). It comprises nearly 34.000 km2 circumscribed by the 200 m isobath, between 54°–55°S and 56°–62°W, with a slight slope extended nearly 370 km east–west. The north–south width of the area varies between 50 and 100 km (Fig. 1). The depth of the plateau ranges between 50 and 200 m, where the bottom abruptly breaks into a wall reaching 1100 to more than 3000 m depth in some regions (Zunino and Ichazo 1979). This area is characterized by the influence of subantarctic waters of 4–5 °C on the bottom and part of the shelf break during the whole year. No thermoclines have been detected in the plateau area, and mean salinity values reached 34 (Zunino and Ichazo 1979; Piola and Gordon 1989; Guerrero et al. 1999). This plateau was first described at the beginning of an Antarctic Exploration Cruise in 1842, commanded by Captain Sir James Ross (Findlay 1867).

Location of the three sub-areas comprised in the Namuncurá MPA, Burdwood Bank (Argentina). Solid circles show the sampling sites during the research cruise onboard the OV “Puerto Deseado,” April 2013. Image courtesy of Valeria Falabella
Burdwood Bank is an intermediate area between South America and Antarctica and part of the islands and shelf that constitute the Scotia Arc. All these were formerly connected in the supercontinent of Gondwana, but separated since Antarctica’s geographic and thermal isolation through the Cenozoic. Despite geographic distance, depths and oceanographic conditions, the Scotia Arc linked Patagonia and the Antarctic Peninsula, playing a connection role for many marine species (Arntz 2005). Thus, the Patagonia-Scotia Arc-Antarctic Peninsula has become a crucial region of the globe for biological and climatological research (Barnes 2005).
Probably, the first mention (or one of the oldest) regarding the high species richness found at this benthic community at Burdwood Bank was reported by the “Scotia” Expedition, in 1903. One trawl at station no 346 (54°55′S and 57°28′W) reached half a ton (“the largest and richest of the whole Antarctic expedition”), with a preliminary list of 70 species, dominated by sponges and bryozoans (Scotish National Antarctic Expedition 1908). More recently, during the LAMPOS expedition performed by the RV Polarstern (April 2002), a rich benthic community was also found in the east and west slopes of the Burdwood Bank. It was mainly constituted by Porifera and Cnidaria (Arntz and Brey 2003). Currently, BB is formally recognized as a subantarctic area of ecological importance, where biological processes that deserve conservation occurred, when areas of fisheries management and protection were implemented years ago. In 2004, a prohibited area of bottom trawling and fishery of the Patagonian toothfish Dissostichus eleginoides was established at the western portion of the Bank when catches of juveniles exceeded 15 % of the total catch (Resolution of the Consejo Federal Pesquero No 3/2004, Argentina). Thereafter, in 2008, another area of permanent prohibition of fisheries was implemented within the central portion of the Bank (currently the “core” of the Marine Protected Area), in order to conserve the marine bottoms (Resolution of the Consejo Federal Pesquero No 18/2008, Argentina) (Fig. 1). These regulations were the first steps toward the creation (in 2013) of the first open-sea (non-coastal) Argentine Marine Protected Area (Law 26875, Argentina).
In order to provide the necessary baseline knowledge for the establishment of the Namuncurá Marine Protected Area (N MPA), general and historical information on oceanography, biodiversity and fisheries exploitation in the BB was compiled by Falabella et al. (2013). Some reviews (i.e., Zamponi 2008a, b) pointed out the importance of the region based on the presence of vulnerable sessile benthic species like the reported rich coral fauna. Other literature also mentioned the occurrence of several species regarding vulnerable indicators of marine ecosystems (Jones and Lockhart 2011), such as bryozoans, sponges and tunicates. All these organisms have already been recorded at BB (López Gappa 2000; Tatián et al. 2005; Schejter et al. 2012). Criteria of vulnerability of these ecosystems are mainly based on the existence of a necessary high biomass of sessile taxa. The latter are composed by long-living species very sensitive to bottom fishing, high percentages of endemism and very slow recovery times of the community after a perturbation event (FAO 2009; Auster et al. 2011; Portela et al. 2012). Based on these criteria and as a precautionary measure to preserve this ecosystem, a MPA was established.
The N MPA comprises three different sub-areas according to the required protection level. The central one, called “core,” has been defined as the area that contains a representative fraction of the benthic marine biodiversity that requires a strict protection due to the vulnerable characteristics presented by the ecosystem components. No activities are allowed in the core, except for control and monitoring. The core area is surrounded by a “buffer” area, in which activities are allowed after previous authorization (e.g., scientific research, explorations dealing with natural resources and biodiversity, sustainable resources management, restoration and monitoring of global change). Finally, an external “transition” area was defined, in which productive and extractive activities considered in the management plan are allowed. Beyond the transition area, the shelf-break and nearby areas have no protection regulations.
Embedded into the Argentinean “Pampa Azul” Marine National Science Project, the N MPA is a conservation aim that represents a challenge regarding management and planning. It joins the 43 existing Coastal-Marine Protected Areas in Argentina (see Caille et al. 2013). It largely contributes to nearly 4 % of marine areas under some kind of protection in the country, supporting the global objective of the Convention on Biological Diversity (CBD) to protect and preserve the biota of the world’s marine and coastal regions. As a consequence, the acquisition of further knowledge of the region in the near future (species composition, abundance, biomass and distribution of benthic species in the bank and the shelf-break area) is expected.
It is well known that bottom fishing may damage or destroy the seafloor habitat and the associated benthic communities (National Research Council 2002). After drastic fishing activities, the ecosystem would experience severe changes in structure and composition (Hinz et al. 2009). Epibenthic and sessile fauna are vulnerable to fishing disturbance, though the level of disturbance is related to the habitat type and may or may not recover after fishing closures or interrupted activities (Queirós et al. 2006; Asch and Collie 2008). Several countries that created MPAs developed action plans and management strategies in order to preserve the biodiversity of vulnerable and sensitive ecosystems, such as coral habitats (Wattage et al. 2011). Protection and conservation issues also apply for the international waters and vulnerable marine ecosystems (Durán Muñoz and Sayago-Gil 2011; Jones and Lockhart 2011; Durán Muñoz et al. 2012; Portela et al. 2012, 2015). Cold-water coral areas remained largely unappreciated and overlooked until 1990s, although their existence was known since the time of Linnaeus (Cairns 2007; Roberts and Cairns 2014). It is also known that more than 65 % of all the extant corals (Anthipatharia + Scleractinia + Octocorallia + Stylasteridae) are cold-water species, occurring at depths >200 meters (Cairns 2007). These habitats face considerable threat from ocean acidification (Guinotte et al. 2006), and the future distribution pattern of the benthic communities might be influenced by climate change (Fautin et al. 2009). It is expected that the vulnerable benthic communities recognized near N MPA (BB), as those recently described along the Argentine continental margin (i.e., coldwater coral reefs, coral gardens, sponge beds and rocky environments) (Portela et al. 2012, 2015) and South Georgia Islands (Gaitán et al. 2013a, b) should be protected, studied and monitored from now on.
In this contribution, we present current data on benthic richness of the N MPA based on results from a sampling carried out in April 2013. We provide an updated inventory of benthic taxa collected at the core, buffer and shelf-break areas in order to describe the benthic community in the MPA and to recommend future studies that should be developed for its proper management.
Materials and methods
Four sampling sites were supplementary added to the original campaign planning onboard the Oceanographic Vessel “Puerto Deseado” (CONICET, Argentina) at the return of the Argentinean Antarctic Expedition in April 2013. The objective of the sampling was to perform a preliminary assessment of the Burdwood Bank.
Epibenthic organisms were collected using a bottom otter trawl (14 min, mean trawling time), while infaunal small mollusks were sampled using a small box dredge (45 × 50 × 12 cm). Due to constraints of the sampling methods, only qualitative data have been taken, though visual inspections of the total catch permitted a general assumption of the main taxonomic groups collected in each site.
Due to weather conditions, only three sites (out of the four originally planned) were sampled. These were located at the core, buffer and shelf-break areas of the MPA (Table 1). Epibenthic taxa were sorted, photographed and preserved onboard (frozen, 70 % ethanol and/or formalin–seawater 4 %, depending on the taxa). Taxa were identified at laboratory by different specialists included as co-authors in this paper. Species richness of trawl samples was compared using a similarity index (SI = S/N, where S = taxa in common between samples A and B, and N = cumulative number of taxa in samples A and B), following a similar analysis performed by Lovell and Trego (2003) at Deception Island, Antarctica.
Results
A total of 240 benthic taxa were collected at three different sites at the Namuncurá MPA, Burdwood Bank (Table 2).
One hundred and forty-eight taxa were identified at the core of the MPA (sample volume <90 l). Although not quantitatively assessed, the visual inspection of the general catch revealed that this area was characterized by a relatively high biomass of sponges that corresponded to 16 morphospecies (14 Demospongiae and 2 Calcarea). Few fragments of corals belonging to the stylasterid E. antarctica were also found, with some individuals of the lepadomorph crustacean Ornatoscalpellum gibberum as epibionts. Ten common species of echinoderms were recorded and 12 crustaceans, including 6 decapods comprising the commercial king crabs Lithodes confundens and Paralomis granulosa, 5 peracarids and the above-mentioned lepadomorph. Bryozoans and mollusks presented the highest richness (regarding major groups) in the area: 47 and 25 taxa, respectively. Hydrozoans were represented by 11 taxa, brachiopods by 2 species, ascidians by 8 taxa and pycnogonids by a single taxon, while several polychaetes (13 taxa) were also recorded, although belonging to the infaunal fraction. Ascidians were represented in this station by eight taxa. Most of them were colonial organisms belonging to the family Polyclinidae. Members of this particular Family were rather difficult to identify to the lowest taxonomic level. The reproductive state of the samples collected (asexual phase) was characterized by a regression of the anterior region of the zooids, and the sole presence of phagocytic cells heavily laden with reserves in the postabdomen did not allow us to reach the species or genus level. Four fish species were also caught by the otter trawl.
The sample collected at the station in the buffer area (sample volume <40 l) was mainly composed of brachiopods (living organisms and dead valves) and calcareous polychaete tubes (Serpulidae). A total of 109 taxa were identified. Fifteen echinoderm species (asteroids, sea urchins) were recorded, 48 bryozoans including some big colonies (comprising the lace coral Reteporella magellensis), 8 sponges, 1 hydrocoral (E. antarctica), 5 decapods and 1 amphipod among crustaceans, 8 mollusks, 7 polychaetes, 10 hydrozoans, 1 ascidian and 2 fishes.
A remarkable presence of corals and ophiuroids was detected visually in the shelf break of the bank, at the end of the transition area of the MPA. These coral aggregations at the slope of the shelf break probably caused the breakage of the otter trawl net, hence producing the poorest sample in terms of volume and consequently in taxa richness (sample volume <30 l). A total of 92 taxa were identified, being primnoid corals (Thouarella spp.) the most conspicuous organisms in this site. Two individuals of Flabellum cf. thouarsi, several E. antarctica pieces and 2 Alcyonium sp. colonies were recorded. Ophiura lymani was the most abundant ophiuroid collected, and Gorgonocephalus chilensis was also important in terms of the relative observed biomass in the catch. Other 12 echinoderm taxa, 12 sponges, 15 mollusks, 9 polychaetes, 1 hydrozoan colony, 1 ascidian, 3 decapods and 3 peracarid crustaceans (isopods), 1 pycnogonid, 3 sea anemones, 1 brachiopod and 2 fish species were recorded.
Similarity index (SI) obtained for core and buffer areas was 0.3066; for buffer and shelf break, it was 0.1994; and for core and shelf break, it was 0.1881.
Discussion
This contribution represents the most recent and complete checklist of benthic organisms (mainly mega and macrofauna) that summarized 240 taxa collected at three sites in the Namuncurá Marine Protected Area, at Burdwood Bank (Table 2). Only partial information by taxonomic group (e.g., fishes, mollusks, crustaceans) is available from previous studies performed in the current N MPA (see a general compilation at Falabella et al. 2013). The Ocean Biogeographic Information System (OBIS) Web site recorded no information for the three sites here studied and reported only 90 taxa in the transect sampling line that contains the stations that were carried out in the present study (checked at http://www.iobis.org/ in December 2015). Previous studies performed in the slope area of the BB (Arntz and Brey 2003; Schejter et al. 2012) also reported a rich and diverse benthic community. We are well aware that one sample per site is not enough to fully characterize the benthic assemblage, but still, in the present scenario, these data represent a valuable source of information and a very helpful starting point for incoming research, conservation efforts and management. To generate complete inventories at species level of big areas is a very expensive and time-consuming venture. Studies that used taxa higher than species level (e.g., genus and family) as surrogates for predicting species level diversity in marine areas proved to be useful, and taxa assemblages together with the information of endemic, vulnerable and threatened taxa help to evaluate areas that merit conservation efforts (Vanderklift et al. 1998). The National Marine Scientific Policy and Strategy in Argentina includes the scheduling of research cruises in this MPA (among other regions) in the years to come in order to improve data and management of ecologically important areas in this country.
Faunistic composition of the three sampled sites at BB was different, although some species were shared among them (Table 2). The benthic assemblage collected from the buffer area shared more than 50 % of the recorded taxa with the core area (mainly bryozoans, hydrozoans and decapod crustaceans). Only 30 % of the taxa were shared among shelf-break and buffer areas and also 30 % among core and shelf-break regions (mainly bryozoans in both cases). Similarity indexes also reflected these differences among stations, being core and buffer regions more related among them than the shelf-break area. These differences highlighted the importance of a better characterization of the benthic communities that inhabit the Namuncurá MPA and the nearby shelf break. This huge extension presents not only bathymetric variations and particular oceanographic conditions (Zunino and Ichazo 1979; Piola and Gordon 1989; Guerrero et al. 1999) but also a different disturbance history in the whole area related to fishing restrictions (see Introduction) with the consequent variations on recovery times of the target and nontarget species involved in the fishing operations. Affinities with other benthic communities of the Argentinean shelf are evident. At least at the core region in areas of about 100 m depth, faunal species recorded were frequent taxa at Patagonian scallop benthic community distributed along the Argentinean shelf break front at similar depths (Bremec and Lasta 2002; Genzano et al. 2009; López Gappa and Landoni 2009; Sánchez et al. 2011; Schejter et al. 2012). On the other hand, it is possible that shelf-break fauna of BB caught at more than 200 m depth is similar to regions sampled at similar depths outside the Argentinean continental shelf in northern latitudes (see Portela et al. 2012, 2015).
Considering the phyla here recorded, bryozoans presented the highest richness compared to the other groups, although among the three areas, the lowest richness was reported for the shelf-break area. This fact could be due to the smaller size of the benthic sample compared to the others. Bryozoa is one of the best studied phyla from the given list; this group has been revised from samples collected from more than 30 years ago to the present by the same taxonomist (López Gappa and Lichtschein 1990; López Gappa 2000). Another important contribution was carried out by Hayward (1980). Also, additional records from outside the MPA coming from the LAMPOS expedition mentioned in the Introduction section can be found in Moyano (2005).
High abundances of recent and fossil corals from Burdwood Bank slope and other nearby areas are known from the literature (e.g., Zamponi 2008a, b; Margolin et al. 2014). In accordance with this, a high diversity of anthozoans (octocorals, scleractinians and sea anemones) was previously found outside the MPA, in the eastern side of the bank (López-González et al. 2003), as well as in the west (Margolin et al. 2014). However, during the present study, anthozoans were only registered in the shelf-break area of the BB, a site where relatively high abundance and richness of these organisms were recorded. Corals, coral gardens and reefs are also present and frequent beyond the shelf break of Argentina (Muñoz et al. 2012; Cairns and Polonio 2013; Portela et al. 2012, 2015), even though they could also be found in lower abundances at the Argentinean shelf, below 200 m depth (Zamponi 2008b).
Sponges were not shared among areas, except for one species (Mycale sp. 1). Although not quantified, this phylum was responsible for the main biomass in the core area, probably the less known area regarding spongofauna. Other previous sponge reports from BB came from sampling sites located in the transition area or beyond the shelf break (López Gappa and Landoni 2005; Schejter et al. 2012).
Peracarid’s species richness collected with the otter trawl underestimated the real values since these organisms, as well as other small invertebrates, require a different sampling method. During the same research cruise, a Rauschert sledge equipped with a 1 mm mesh size took three additional samples in the same sites (material not available for this study). A preliminary revision showed a remarkable high peracarid species richness. However, these results were not included in this study. At least 90 species (including amphipods, isopods, cumaceans and tanaidaceans) were identified and many of them represented new distributional range records (Doti et al. 2014; Chiesa et al. 2015).
The benthic richness here documented largely exceeds the values coming from other productive areas in Argentinean waters where different trawling fisheries are conducted, even considering the limitations of this inventory. In this sense, this area should be considered a hot spot of benthic biodiversity. Other important but less biodiverse areas of the SW Atlantic Ocean are located on the continental shelf between 38°S and 45°S, at the Patagonian scallop fishing grounds. In these areas, species richness was not higher than 70 species per site, considering a maximum of 38 megafaunal taxa, 15 bryozoa and about 17 sponges and other epibiotic taxa (see Bremec et al. 2000, 2008; Schejter and Bremec 2007, 2009; Schejter et al. 2011a, b, 2013; Escolar et al. 2009; López Gappa and Landoni 2009). Also, the Argentinean–Uruguayan Common Fishing Zone (35°S–39°S) and the Patagonian shelf (44°S–47°S) (common hake fishery, see Giberto et al. 2014) showed lower values of benthic richness. In addition to the shelf areas mentioned above, 86 taxa of mega and macroinvertebrates were found using a dredge of 10 mm mesh size in a submarine canyon at 43°35′S and 59°33′W, 325 m depth (Bremec and Schejter 2010). The faunal composition of the latter area was similar to that referred to in adjacent external shelf areas, mainly between 90 and 130 m depth (Bremec and Lasta 2002). Moreover, after studying epibiotic relationships in the mentioned canyon, species richness was increased to 127 taxa (Schejter et al. 2014), a value still lower than the richness here reported for BB. Research on benthic biodiversity using samples collected at the Scotia Arc and also referred that BB contributed much to diversity in the study area, such as the case of gorgonians, gastropods and bivalves (Arntz and Brey 2003). Roux et al. (2002) found a richness of a maximum of 27 taxa per site at South Georgia Islands, using demersal trawls. Antarctic studies at Deception Island also revealed a similar richness of 35 taxa per sample (Lovell and Trego 2003). Although benthic richness in the different regions of Argentina and Antarctica has been estimated using different sampling methods, we consider that the present results on mega and macrofauna are useful as a general reference regarding the benthic richness of the marine realm.
In the case of the N MPA, an extensive review of scattered benthic information, future sampling cruises, possible ROV images and sediment sampling studies will help to update species inventories. It will also help detect indicator taxa vulnerable to trawling, recognize distribution patterns of the benthic organisms and support management strategies. As epibiotic relationships greatly increase benthic richness in other southern benthic systems, similar studies should also be conducted. It is also essential to establish the spatial distribution of indicator taxa, like corals, sponges and bryozoans, to better attempt to the protection of benthic biodiversity in the rich Argentinean marine system herein studied. Furthermore, this information will provide a stronger idea about the biogeographic importance of the Burdwood Bank, either as center of endemisms and/or as connection for marine benthic species between South America and the Antarctic Peninsula.
References
Arntz WE (2005) The Magellan-Antarctic connection: links and frontiers at southern high latitudes. Summ Rev Sci 62:359–365
Arntz W, Brey T (eds) (2003) Expedition ANTARKTIS XIX/5 (LAMPOS) of RV “Polarstern” in 2002. Ber Polarforsch Meeresforsch 462, 124 pp
Asch RG, Collie JS (2008) Changes in a benthic megafaunal community due to disturbance from bottom fishing and the establishment of a fishery closure. Fish Bull 106:438–456
Auster PJ, Gjerde K, Heupel E, Watling L, Grehan A, Rogers AD (2011) Definition and detection of vulnerable marine ecosystems on the high seas: problems with the “move-on” rule. ICES J Mar Sci 68:254–264
Barnes DKA (2005) Changing chain: past, present and future of the Scotia Arc’s and Antarctica’s shallow benthic communities. Sci Mar 62:65–89
Bremec CS, Lasta ML (2002) Epibenthic assemblage associated with scallop (Zygochlamys patagonica) beds in the Argentine shelf. Bull Mar Sci 70:89–105
Bremec C, Schejter L (2010) Benthic diversity in a submarine canyon in the Argentine sea. Rev Chil Hist Nat 83:453–457
Bremec C, Brey T, Lasta M, Valero J, Lucifora L (2000) Zygochlamys patagonica beds on the Argentinean shelf. Part I: energy flow through the scallop bed community. Arch Fish Mar Res 48:295–303
Bremec C, Marecos A, Schejter L, Escolar M, Souto V (2008) Riqueza específica y asociaciones faunísticas en los bancos comerciales de vieira patagónica (Zygochlamys patagonica) a lo largo del frente de talud. Período 2007. INIDEP Tech Rep 53:29
Caille G, Musmeci JM, Harris G, Delfino Schenke R (2013) Sistema Inter-Jurisdiccional de Áreas Protegidas Costero Marinas -SIAPCM - Argentina (Proyecto ARG/10/G47 GEF -PNUD). Frente Maritimo 23:55–64
Cairns SD (2007) Deep-water corals: an overview with special reference to diversity and distribution of deep-water scleractinian corals. Bull Mar Sci 81:311–322
Cairns SD, Polonio V (2013) New records of deep-water Scleractinia off Argentina and the Falkland Islands. Zootaxa 3691:58–86
Chiesa IL, Urteaga D, Martinez AI, Doti BL, Roccatagliata D (2015) Biodiversidad de anfípodos bentónicos del AMP Namuncurá – Banco Burdwood. Abstracts Book, IX Jornadas Nacionales de Ciencias del Mar, Ushuaia p263
Doti B, Chiesa IL, Alberico N, Sganga D, Giachetti C, Pereira E, Roccatagliata D (2014) Biodiversidad de crustáceos Peracarida del Banco Burdwood/Namuncurá: resultados preliminares. Abstracts Book, III Congreso Uruguayo de Zoología, Montevideo, pp 291–292
Durán Muñoz P, Sayago-Gil M (2011) An overview of cold-water coral protection on the high seas: the Hatton bank (NE Atlantic)—a case study. Mar Policy 35:615–622
Durán Muñoz P, Sayago-Gil M, Murillo FJ, Del Río JL, López-Abellán LJ, Sacau M, Sarralde R (2012) Actions taken by fishing Nations towards identification and protection of vulnerable marine ecosystems in the high seas: the Spanish case (Atlantic Ocean). Mar Policy 36:536–543
Escolar M, Diez M, Hernández D, Marecos A, Campodónico S, Bremec C (2009) Invertebrate bycatch in Patagonian scallop fishing grounds: a study case with data obtained by the on board observers programm. Rev Biol Mar Oceanogr 44:369–377
Falabella V, Campagna C, Caille G, Krapovickas S, Moreno D, Michelson A, Piola A, Schejter L, Zelaya D (2013) Banco Burdwood: Contribuciones para el establecimiento de una línea de base y plan de manejo de la futura Área Marina Protegida. Preliminar report, 51 pp
FAO (2009) Report of the technical consultation on international guidelines for the management of deep-sea fisheries in the high seas. Rome, 4–8 February and 25–29 August 2008. FAO fish aquac rep 881
Fautin DG, Guinotte JM, Orr JC (2009) Comparative depth distribution of corallimorpharians and scleractinians (Cnidaria: Anthozoa). Mar Ecol Prog Ser 397:63–70
Findlay AG (1867) A sailing directory for the Ethiopic or South Atlantic Ocean including the coasts of South America and South Africa. R.H. Laurie, London, 691 p
Gaitán E, Schejter L, Giberto D, Escolar M, Bremec C (2013a) Report of vulnerable marine ecosystems in South Georgia Islands (CCAMLR Subarea 48.3) through research dredge sampling. CCMLAR, Working group-fish stock assessment -13/58
Gaitán E, Schejter L, Giberto D, Escolar M, BremecC (2013b) Registro de invertebrados bentónicos en las Islas Georgias del Sur (sub-área CCRVMA 48.3) como potenciales taxones indicadores de Ecosistemas Marinos Vulnerables. INIDEP research rep 77, 7 pp
Genzano PN, Giberto D, Schejter L, Bremec C, Meretta P (2009) Hydroid assemblages from Southwestern Atlantic Ocean (34–42°S). Mar Ecol 30:33–46
Giberto DA, Romero MV, Souto V, Escolar M, Bremec CS, Machinandiarena L (2014). Fauna bentónica asociada a prerreclutas de merluza en la Zona Común de Pesca Argentino-Uruguaya y en la plataforma patagónica entre 44° y 47°S. INIDEP research rep 49, 18 pp
Guerrero RA, Baldoni A, Benavides H (1999) Oceanographic conditions at the southern end of the argentine continental slope. INIDEP Sci Doc 5:7–22
Guinotte J, Orr J, Cairns S, Freiwald A, Morgan L, George R (2006) Will human-induced changes in seawater chemistry alter the distribution of deep-sea scleractinian corals? Front Ecol Environ 4(3):141–146
Hayward PJ (1980) Cheilostomata (Bryozoa) from the South Atlantic. J Nat Hist 14:701–722
Hinz H, Prieto V, Kaiser MJ (2009) Trawl disturbance on benthic communities: chronic effects and experimental predictions. Ecol Appl 19:761–773
Jones CD, Lockhart SJ (2011) Detecting vulnerable marine ecosystems in the Southern Ocean using research trawls and underwater imagery. Mar Pol 35:732–736
López Gappa J (2000) Species richness of marine Bryozoa in the continental shelf and slope off Argentina (south-west Atlantic). Div Distrib 6:15–27
López Gappa J, Landoni NA (2005) Biodiversity of Porifera in the Southwest Atlantic between 35 S and 56 S. Rev Mus Arg Cs Nat 7:191–219
López Gappa J, Lichtschein V (1990) Los Briozoos Coleccionados por el B/I Shinkai Maru en la Plataforma Continental Argentina. Parte I. Servicio de Hidrografia Naval, Buenos Aires
López Gappa J, Landoni NA (2009) Space utilisation patterns of bryozoans on the Patagonian scallop Psychrochlamys patagonica. Sci Mar 73:161–171
López-González P, Rodríguez E, Vert N (2003) Biogeography and Ecology of Cnidaria. In: Arntz W, Brey T (eds) Expedition ANTARKTIS XIX/5 (LAMPOS) of RV “Polarstern” in 2002. Ber Polarforsch Meeresforsch 462: 13–18
Lovell LL, Trego KD (2003) The epibenthic megafaunal and benthic infaunal invertebrates of Port Foster, Deception Island (South Shetland Islands, Antarctica). Deep-Sea Res II 50:1799–1819
Margolin AR, Robinson LF, Burke A, Waller RG, Scanlon KM, Roberts ML, Auro ME, van de Flierdt T (2014) Temporal and spatial distributions of cold-water corals in the Drake Passage: insights from the last 35,000 years. Deep-Sea Res II 99:237–248
Moyano HI (2005) Scotia Arc bryozoans from the LAMPOS expedition: a narrow bridge between two different faunas. Sci Mar 69(Suppl.2):103–112
Muñoz A, Cristobo J, Ríos P, Druet M, Polonio V, Uchupi E, Acosta J, Group A (2012) Sediment drifts and cold-water coral reefs in the Patagonian upper and middle continental slope. Mar Petrol Geol 36:70–82
National Research Council (2002) Effects of Trawling and Dredging on Seafloor Habitat. Committee on Ecosystem Effects of Fishing: Phase 1- Effects of Bottom Trawling on Seafloor Habitats, Ocean Studies Board, Division on Earth and Life Studies. Washington DC, National Academy, p 126
Piola AR, Gordon AL (1989) Intermediate waters in the southwest South Atlantic. Deep-Sea Res 36(1):1–16
Portela J, Acosta J, Cristobo J, Muñoz A, Parra S, Ibarrola T, Del Río JL, Vilela R, Ríos P, Blanco R, Almón B, Tel E, Besada V, Viñas L, Polonio V, Barba M, Marín P (2012) Management Strategies to Limit the Impact of Bottom Trawling on VMEs in the High Seas of the SW Atlantic, In: Cruzado A (ed) Marine Ecosystems. in tech, pp 199–228
Portela J, Cristobo J, Ríos P, Acosta J, Parra S, Del Río JL, Tel E, Polonio V, Muñoz A, Patrocinio T, Vilela R, Barba M, Marín P (2015) A first approach to asses the impact of bottom trawling over vulnerable marine ecosistems on the high seas of the Southwest Atlantic. In: Blanco JA (ed) Biodiversity in ecosystems - linking structure and function. InTech, p 28. http://www.intechopen.com/books/biodiversity-inecosystems-linking-structure-and-function/a-first-approach-to-assess-the-impact-of-bottom-trawling-overvulnerable-marine-ecosystems-on-the-hi
Queirós AM, Hiddink JG, Kaiser MJ, Hinz H (2006) Effects of chronic bottom trawling disturbance on benthic biomass, production and size spectra in different habitats. J Exp Mar Biol Ecol 335:91–103
Roberts JM, Cairns SD (2014) Cold-water corals in a changing ocean. Curr Op Environ Sust 7:118–126
Roux A, Calcagno JA, Bremec C (2002) Macrobenthic assemblages of demersal fishing grounds off South Georgia Islands. “R/V Eduardo Holmberg” survey, February–March, 1994. Arch Fish Mar Res 49:231–241
Sánchez MA, Giberto D, Schejter L, Bremec C (2011) The Patagonian scallop fishing grounds in shelf break frontal areas: the non assessed benthic fraction. Lat Am J Aquat Res 39:167–171
Schejter L, Bremec C (2007) Benthic richness in the Argentine continental shelf: the role of Zygochlamys patagonica (Mollusca, Bivalvia, Pectinidae) as settlement substrate. J Mar Biol Assoc UK 87:917–925
Schejter L, Bremec C (2009) Epibiosis contest at Zygochlamys patagonica fishing grounds: Which is the winner? 17th international pectinid workshop, Santiago de Compostela, Spain, April 22–28 2009. Book of abstracts pp 143–144
Schejter L, Escolar M, Bremec C (2011a) Variability in epizoic colonization on Fusitriton magellanicus shells. J Mar Biol Assoc UK 91(4):897–906
Schejter L, Bertolino M, Calcinai B, Cerrano C, Bremec C (2011b) Epibiotic sponges on the hairy triton Fusitriton magellanicus in the SW Atlantic Ocean, with the description of Myxilla (Styloptilon) canepai sp. nov. Aquat Biol 14:9–20
Schejter L, Bertolino M, Calcinai B,Cerrano C, Pansini M (2012) Banco Burdwood: resultados preliminares sobre composición y riqueza específica de esponjas (Phylum Porifera), a partir de muestras colectadas en la campaña del buque rompehielos estadounidense “Nathaniel B. Palmer”, abril-mayo 2008. INIDEP research rep 122, 6 pp
Schejter L, Escolar M, Marecos A, Bremec C (2013) Riqueza específica en los bancos comerciales de vieira patagónica (Zygochlamys patagonica) a lo largo del frente de talud. Período 2012. INIDEP research rep 18, 13 pp
Schejter L, López Gappa J, Bremec C (2014) Epibiotic relationships on Zygochlamys patagonica (Mollusca, Bivalvia, Pectinidae) increase biodiversity in a submarine canyon in Argentina. Deep Sea Res II 104:252–258
Scotish National Antarctic Expedition 1902–1904 (1908) Report on the scientific results of the voyage of the S.Y. “Scotia” during the years 1902, 1903, and 1904 under the leadership of William S. Bruce, vol 4, Zoology
Tatián M, Antacli JC, Sahade R (2005) Ascidians (Tunicata: Ascidiacea): species distribution along the Scotia Arc. Sci Mar 62:205–214
Vanderklift MA, Ward TJ, Phillips JC (1998) Use of assemblages derived from different taxonomic levels to select areas for conserving marine biodiversity. Biol Conserv 8686:307–315
Wattage P, Glenn H, Mardle S, Van Rensburg T, Grehan A, Foley N (2011) Economic value of conserving deep-sea corals in Irish waters: a choice experiment study on marine protected areas. Fish Res 107:59–67
Zamponi M (2008a) Informe Banco Burdwood. Cnidaria. Mar del Plata, 15 pp
Zamponi M (2008b) La corriente de Malvinas: ¿una vía de dispersión para cnidarios bentónicos de aguas frías? Rev Real Acad Galega Cs XXVII:183–203
Zunino G, Ichazo MM (1979) Los peces demersales del Banco Burdwood: distribución, abundancia de las especies y frecuencia de tallas (según datos de los B/I Walther Herwig y Shinkai Maru, campañas 1978–1979). Final report, Oceanografía Biológica, Universidad de Buenos Aires, 66 pp
Acknowledgments
We would like to thank to the OV “Puerto Deseado” and the crew for the help during sampling procedures, as well as to Pablo Bonuccelli and the Hydrographic Research Vessels Division (Argentinean Navy), for the given permission to add Burdwood Bank sampling sites in the schedule of the survey. We also thank Gustavo Lovrich for their critical suggestions on preliminary versions of this MS and for the help during the identification of the king crab species. We also thank the reviewers for their critical revision of the MS and the suggestions received that improve the final result of this work. Prof. María Cecilia Martin helped in correcting the English grammar. This study was partially supported by PICT 2013-0629 (to LS), PICT-2012-1043 and PIP 11220120100247 (to JLG), PIP 11420110100323 (to DGZ), FACEPE—Fundação de Amparo à Ciência e Tecnologia do Estado de Pernambuco (APQ-0828-2.04/12) (to CDP) EXA 639/13 UNMdP 2013-2014 and PIP 11220110100152 (to GG).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Schejter, L., Rimondino, C., Chiesa, I. et al. Namuncurá Marine Protected Area: an oceanic hot spot of benthic biodiversity at Burdwood Bank, Argentina. Polar Biol 39, 2373–2386 (2016). https://doi.org/10.1007/s00300-016-1913-2
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-016-1913-2