
Abstract
The risk of hydrocarbon contamination in marine polar areas is constantly increasing. Autochthonous bacteria, due to their ability to cope and survive under extreme environmental conditions, can play a fundamental role in the hydrocarbon degradation. The degradation process is often enhanced by the production of biosurfactant molecules. The present study reports for the first time on the isolation of biosurfactant-producing bacteria from Arctic and Antarctic shoreline sediments. A total of 199 psychrotolerant bacterial isolates were obtained from hydrocarbon-amended (with crude or diesel oil) microcosms. A total of 18 isolates were selected for their ability to grow in the presence of crude oil and produce biosurfactants, as it was revealed by the production of good E24 values (≥50 %) and/or reduction in the surface tension (under 30 mN/m). The positive response of the isolates to both tests suggests a possible production of biosurfactants with emulsifying and interfacial activities. Biosurfactant-producing isolates were mainly affiliated to the genera Rhodococcus (14 isolates), followed by Pseudomonas (two isolates), Pseudoalteromonas (one isolate) and Idiomarina (one isolate). Thin-layer chromatography of biosurfactant crude extracts revealed that the majority of the selected isolates were able to produce glycolipidic surfactants. Our results enlarge the knowledge, which is still poor and fragmentary, on biosurfactant producers from polar areas and indicate marine polar sediments as a source of bacteria with potential applications in the remediation of hydrocarbon-contaminated cold environments.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Polar environments are increasingly subject to anthropogenic pollution. The exploration and exploitation of oil in Arctic offshore stations and the growing research interest about extreme environments, such as Antarctica, are constantly increasing the risk of hydrocarbon contamination. Hydrocarbons that reach the marine environment are naturally modified by physical, chemical and biological processes. Biodegradation is an essential part of the process, contributing greatly to the natural attenuation of hydrocarbons. To enhance the availability of hydrophobic pollutants to be used as carbon and energy source, microorganisms can employ several strategies such as the solubilization mediated by polymers with surfactant activity or biosurfactants (Cameotra et al. 2010). Among the characteristics that are assignable to such molecules are, for example, the increase in the contact area of substrates insoluble in water through emulsification, the involvement in the process of pathogenesis, antimicrobial activity or also accession control and detachment of microorganisms to/from the external surfaces (Desai and Banat 1997).
Chemical and microbial surfactants are amphiphilic compounds, meaning that they contain both hydrophobic and hydrophilic groups. The hydrophilic groups consist of mono-, oligo- or polysaccharides, peptides or proteins, while the hydrophobic moiety usually contains saturated, unsaturated and hydroxylated fatty acids or fatty alcohols (Lang 2002). Compared to their chemically synthesized equivalents, biological surfactants have many advantages. For example, they are environmentally friendly, biodegradable, less toxic and non-hazardous. In addition, they have better foaming properties and higher selectivity. Finally, they are active at extreme temperatures, pH and salinity as well and can be produced from industrial wastes and by-products (Desai and Banat 1997).
Hydrocarbon-oxidizing bacteria are ubiquitous, even in environments with no evident contamination, and inhabit also cold polar habitats. A wide variety of autochthonous microorganisms have been proven to possess the ability to degrade aliphatic and aromatic hydrocarbons at low temperatures in the marine environment, therefore playing a significant role in the in situ biodegradation of hydrocarbons (De Domenico et al. 2004; Michaud et al. 2004; Deppe et al. 2005; Gerdes et al. 2005). However, to date, among them only few marine cold-adapted biosurfactant producers have been characterized and they have been mainly isolated from seawater (Yakimov et al. 1999; Pepi et al. 2005; Pini et al. 2007). Only recently, Cai et al. (2014) reported on the isolation of biosurfactant-producing bacteria from petroleum hydrocarbon-contaminated sediment (and water) from North Atlantic, Canada.
In this context, the present research was aimed at investigating the ability of some Arctic and Antarctic cold-adapted culturable bacteria from shoreline sediment in producing biosurfactants.
Materials and methods
Sample collection
Shoreline marine sediment (first 10 cm) and coastal seawater samples were aseptically collected from two polar sites, i.e. the Byers Peninsula, South Shetlands Islands (Antarctica) (MicroBy; 62°40′09,9″S–61°09′04″W; Austral Summer 2008–2009), and the Kongsfjorden, Svalbard Islands (Norwegian Arctic) (MicroSval; 78°55,606′N–11°56,377′E; July 2009). In particular, the Arctic sampling site was next to the Ny-Ålesund research village. Sampling was performed using pre-sterilized polycarbonate bottles. Samples were stored at 4 °C until processing.
At sampling time, water temperature was 6 and 7.9 °C, while salinity was 30 and 22.8 for the Antarctic and Arctic site, respectively.
Microcosm set-up
For each site, microcosms were created by filling 250-ml pre-sterilized glass beakers with 150 g of sediment and 150 ml of filter-sterilized seawater. Microcosms were amended in duplicate with crude oil (Arabian light) or commercial diesel oil (Q8 Italy) (1.5 %, v/v).
Hereinafter, each microcosm is identified by the site name (i.e. MicroSval or MicroBy) and the amendments with crude oil (C) or diesel oil (D) (e.g. MicroSval-D is the sample from the Kongsfjorden that was amended with diesel oil). Microcosms were incubated in the dark at 4 °C. Every day, microcosms were manually stirred under sterile conditions to provide aeration.
Bacterial isolation
Aliquots (1 ml) of the upper sediment layer (up to 2 cm under the surface) were aseptically collected from each microcosm. Cells were detached from sediment particles by adding 9 ml of phosphate-buffered saline (PBS: 120 mM NaCl, 2.7 mM KCl in 10 mM phosphate buffer, pH 7.6), 0.1 % sodium pyrophosphate and 10 µl of sterile Tween 80 and then shacked vigorously for 20 min.
Serial dilutions of detached solution from sediment samples were prepared (1:10 and 1:100, using filter-sterilized seawater), and 100 µl of each dilution was spread plated in two replicates on the mineral medium Bushnell Haas Agar (BH; Difco) plus 3 % (w/v) of sodium chloride, with the addition of crude oil (Arabian light; BH-C) or diesel oil (Q8 Italy; BH-D) as the sole source of carbon for the growth of hydrocarbon-oxidizing bacteria. The carbon sources were spotted on a portion of sterile filter paper (pads) and then placed on the lid of the Petri dishes (Lo Giudice et al. 2010). Such method mainly allows more volatile compounds of crude oil and diesel oil to be used by bacteria for growth. Plates were incubated at 4 °C for 1 month. For bacterial isolation, colonies were randomly selected from agar plates, picked and subcultured three times under the same conditions.
Screening of the isolates for growth on crude oil
Isolates were screened for their ability to growth aerobically in BH medium (plus 3 % NaCl; 10 ml) with crude oil (2 %, v/v) as sole supplied carbon and energy source. Bacterial growth was assessed by comparison between inoculated tube plus added substrate and inoculated tube with no carbon source. Uninoculated cultures were used as negative controls. Bacterial growth was observed after 1 month of incubation at 4 °C.
Selection of biosurfactant producers
Isolates that were able to grow in the presence of crude oil were further screened for biosurfactant production. For a preliminary screening, aliquots (100 µl) of bacterial suspensions in BH were used to inoculate 100 ml of BH supplemented with tetradecane or sunflower oil (a good low-cost alternative to hydrocarbons) at a final concentration of 2 % (v/v) as a sole carbon source. All tests were performed in duplicate in 500-ml Erlenmeyer flaks. Bacterial cell cultures were grown under shaking (120–130 rpm) at 15 and 4 °C for 7 or 21 days, respectively.
Surface tension of each cell culture was measured by using a digital tensiometer (TD1, Lauda) with du Noüy platinum ring. Cultures were preliminary centrifuged at 4700 rpm for 20 min at 4 °C, and only the cell-free supernatants were used. A surface tension lower than 40 mN m−1 was considered as an expression of biosurfactant production (Cooper et al. 1979; Bodour and Maier 1998).
The capability of isolates to produce stable emulsions was determined by the E24 test (Batista et al. 2006). A 2-ml portion of the sample from each culture was added to 2 ml of kerosene (Petroleum ether, Panreac) as test oil, and the mixture was vigorously vortexed for 1 min. After 24 h, the emulsification index (E24) was calculated by dividing the measured height of the emulsion layer by the total height of the mixture and multiplying by 100 (Satpute et al. 2008). Values that were higher than 40 % were considered as a sign of biosurfactant production. Sodium dodecyl sulphate (SDS), a chemical anionic surfactant used in many cleaning and hygiene products, was used as positive control.
Biosurfactant haemolytic activity
The haemolytic activity of produced biosurfactants was evaluated after spotting 5 µl of each culture on a blood agar (BA) plate and incubation at 15 °C for 48 h. Plates were then observed for the presence of clearing halos around the spots, as an indication of biosurfactant production (Carrillo et al. 1996).
Thin-layer chromatography
Biosurfactants were extracted at the time when emulsification and/or surface tension reduction were recorded. Carbon sources (sunflower oil or kerosene) were first extracted by adding hexane (1:1, v/v) to each culture. After mixing, the aqueous phase, the hydrophobic phase and the biomass were separated by centrifugation (4700 rpm at 4 °C for 10 min). The aqueous phase was then transferred into a new flask, acidified with 1 % (v/v) of 85 % H3PO4, and the biosurfactant was extracted two times by centrifugation with 1.25 volume of ethyl acetate.
The obtained biosurfactant crude extract was then preliminary characterized by the thin-layer chromatography (TLC) according to Syldatk et al. (1985). As a reference material, rhamnolipids were used. The rhamnolipids were prepared from Jeneil JBR 425 (Jeneil Biosurfactants Company, Saukville, USA), containing the rhamnolipids 1 to 4 (RL1 to RL4) of Pseudomonas aeruginosa. For the reference, 85 μl of JBR 425 was suspended in 1 ml of 0.1 M sodium phosphate buffer, pH 7, and acidified with 10 μl of concentrated phosphoric acid. This mixture was extracted with 1.333 ml of ethyl acetate, and the obtained ethyl acetate phase, containing the crude rhamnolipid extract, was applied as TLC standard. The same procedure was applied to the samples. Silica60-coated plates were used for TLC (Machery and Nagel 20 × 20 mm. Matrix: 0.20 mm Silicagel 60 ALUGRAM SIL G for TLC REF 818 033), and the solvent system was chloroform:methanol:acetic acid (65:15:2). The sugar moieties were stained with anisaldehyde (anisaldehyde:sulphuric acid:glacial acetic acid 0.5:1:50); the fatty acid moieties were stained with ammonium molybdate/cerium sulphate (0.42 % w/v ammonium molybdate and 0.2 % w/v cerium(IV) sulphate in 6.2 % sulphuric acid). The colour was developed by heating with a fan at 200 °C.
Biomolecular characterization of BS-producing bacterial isolates
Biomolecular characterization was done for those strains that produced low surface tension or/and interesting spots on TLC plates and/or emulsification. Single colonies grown on marine agar (MA) plates were lysed by thermal shock to extract genomic DNA. Cell lysates were directly used for 16S rRNA gene amplification by PCR (Rizzo et al. 2013).
The amplification of the 16S rRNA gene was performed with a thermocycler (Mastercycler, GeneAmp PCR-System 9700, Applied Biosystem, USA) using the domain bacteria-specific primers 27F (5′–AGA GTT TGA TCM TGG CTC AG–3′) and 1385R (5′–CGG TGT GTR CAA GGC CC–3′) as follows: one activation step for the polymerase (HotStar TaqTM, Qiagen, Hilden, Germany) for 15 min at 95 °C, 30 cycles of denaturation steps for 1 min at 94 °C, annealing for 1 min at 55 °C, and elongation for 1 min and 30’ at 72 °C, followed by a final elongation step for 10 min at 72 °C. The PCR mix contained 5 μl of 1:10 diluted DNA template, 1.25 U Taq polymerase, 0.7 μl deoxyribonucleotide triphosphates (dNTPs; 0.2 mM final concentration of each dNTP; GE Healthcare, Buckinghamshire, UK), 1 μl of each primer (20 pM final concentration per vial) and 3.5 μl of a reaction buffer (10 × , Qiagen, Hilden, Germany), the total volume being 35 μl.
The results of the amplification reactions were analysed by agarose gel electrophoresis (0.8 %, w/v) in TAE buffer (0.04 M Tris-acetate, 0.02 M acetic acid, 0.001 M EDTA), containing 1 µg/ml of ethidium bromide (Sigma-Aldrich).
Sequencing of PCR products was performed using the reverse primer 1385R (GATC Biotech AG, Konstanz, Germany). The resulting sequences were compared to 16S rRNA gene sequences in the NCBI GenBank database (http://www.ncbi.nlm.nih.gov) using the Basic Local Alignment Search Tool (BLASTN) algorithm.
Morphological, biochemical and physiological characterization of bacterial isolates
Selected isolates were characterized by means of a combination of phenotypic tests as previously reported by Lo Giudice et al. (2012). Colony morphology and pigmentation was recorded from growth on MA at 4 °C. The growth of isolated bacteria at different temperatures was tested in marine broth (MB) incubated at 4, 10, 15, 20, 25 and 30 °C for up to 4 weeks. The pH range for optimal growth was determined in MB with pH values of separate batches of medium adjusted to 4, 5, 6, 7, 8 and 9 by the addition of HCl and NaOH. Salt tolerance test was performed in modified MB prepared with artificial seawater by adjusting the NaCl concentration in the range 0–9 % (w/v). In addition, API 20E and API 20NE tests (BioMerieux) were performed according to manufacturer’s instruction.
Nucleotide sequence accession numbers
Nucleotide sequences obtained from 18 isolates have been deposited in the NCBI GenBank database under the accession numbers KF683118-KF683135.
Results
Bacterial isolation and characterization
A total of 199 bacterial strains were achieved after culturing aliquots of microcosm sediment on BH amended with crude oil or diesel oil (116 and 83 isolates, respectively). In particular, they were mainly isolated from the microcosm MicroBy-C (85 isolates), followed by MicroBy-D (62 isolates), MicroSval-C (31 isolates) and MicroSval-D (21 isolates).
A total of 63 out of the 199 isolates were able to utilize crude oil for their growth as sole energy and carbon source. In particular, 49 of them were from microcosms amended with crude oil (31 and 18 from MicroBy and MicroSval, respectively) and 14 from microcosms amended with diesel oil (8 and 6 from MicroBy and MicroSval, respectively). Among them, 18 isolates were selected as biosurfactant producers (see below), identified by the 16S rRNA gene sequencing and phenotypically characterized. Isolates were mainly affiliated to the genus Rhodococcus (14 out of 18 isolates), followed by few members of the genera Pseudomonas (two isolates), Pseudoalteromonas (one isolate) and Idiomarina (one isolate) (Table 1).
Results on the phenotypic characterization of the BS-producing isolates are reported in Table 2. All isolates were psychrotolerant being able to grow between 4 and 30 °C, but not at 37 °C, according to the definition given by Morita (1975). Lysine and ornithine decarboxylases were only detected in Rhodococcus spp., whereas acetoin was only produced by Pseudomonas sp. 280. None of the isolates were able to produce indole, H2S or amygdaline. Nitrate reduction was observed for all except Rhodococcus sp. 192, and among Rhodococcus isolates, this is the only one that did not produce urease. None of Rhodococcus and Pseudomonas strains showed hydrolysis of protease. Isolates generally hydrolysed at least one of the macromolecules supplied, except for Idiomarina sp. 185.
The majority of isolates grew well at pH values between 4/5 and 9. Only Rhodococcus sp. 176 and Pseudoalteromonas sp. 93 showed a narrower pH range (6–9 and 7–9, respectively). NaCl concentration for growth was generally between 0 and 3 %. Higher NaCl concentrations were tolerated by Pseudoalteromonas sp. 93 and Idiomarina sp. 185 (up to 8 and 9 %, respectively). No isolate was able to grow at the highest tested NaCl concentration (15 %). Based on the phenotypic characterization (including results on biosurfactant productions, see the following section), Rhodococcus isolates that showed high similarity (99–100 %) with the sequence of Rhodococcus sp. EG2 (Accession Number KF5711869) cannot be considered clonal.
Selection and identification of biosurfactant producers
Table 3 summarizes the strain ability for biosurfactant production on tetradecane and sunflower oil as a carbon source. Nine isolates (i.e. Pseudoalteromonas sp. 93 and Rhodococcus spp. 174, 176, 179, 181, 187, 188, 190 and 224) gave positive results in the presence of both substrates, even if not always at the same temperature. All the rest of isolates were able to produce biosurfactants when grown on a single substrate (tetradecane or sunflower oil) and incubated at a single incubation temperature (4 or 15 °C).
Overall, differences in biosurfactants production, in terms of emulsification activity or surface tension reduction, were frequently observed depending on the carbon source used for the cultivation. Eight Rhodococcus isolates (i.e. 174, 176, 179, 180, 181, 188, 225 and 231) showed better results during incubation with sunflower oil. The same was true for Pseudomonas sp. 235. On the contrary, Rhodococcus spp. 187, 190, 191, 227 and Idiomarina sp. 185 appeared to be more efficient in the presence of tetradecane. Among rhodococcus isolates, only Rhodococcus sp. 224 gave similar results in the presence of both substrata. The higher E24 index percentages were achieved by Rhodococcus sp. 176, when growing in presence of sunflower oil, with value of 67 and 60 % at 4 and 15 °C, respectively. The same strain was one of most able emulsifier also when incubated with tetradecane, by achieving an E24 index of 58 %, and a surface tension value of 28.3 mN/m.
The β-haemolysis on blood agar plate was observed for ten isolates (i.e. Pseudoalteromonas sp. 93, Idiomarina sp. 185, and Rhodococcus spp. 174, 176, 180, 181, 187, 190, 224 and 225). Among these, seven isolates (i.e. Pseudoalteromonas sp. 93 and Rhodococcus spp. 174, 176, 181, 187, 190 and 224) produced halos on both substrates. A single isolate (i.e. Pseudomonas sp. 280) showed α-haemolysis (Table 3).
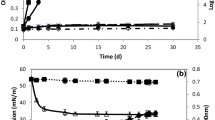
The organic crude extracts from the 18 selected strains were separated on TLC plates according to their hydrophobicity. Yellow–orange spots after staining with anisaldehyde, which revealed the possible presence of carbohydrates in the biosurfactants, were observed for all but three isolates (i.e. Pseudoalteromonas sp. 93, Pseudomonas sp. 280 and Rhodococcus sp. 192). All examined extracts showed a similar profile on TLC plates, with the highest spot at nearly the same height of RL3 or RL4 (rhamnolipid 3 and rhamnolipid 4, respectively) of the standard. The retention factor (Rf) values of spots ranged from 0.23 to 0.35 (Fig. 1a). TLC plates stained with molybdenum for the fatty acid determination showed spots corresponding to those highlighted with anisaldehyde staining, with the same Rf values, by revealing the probable presence of fatty acids (Fig. 1b).

a Example of interesting yellow spots on TLC plate after staining with anisaldehyde (a) or molybdate/cerium (b) RL1 to RL4, standard rhamnolipids from P. aeruginosa
Discussion
In the present study, 18 (out of 199) psychrotolerant bacterial isolates, which were obtained during a microcosm experiment on hydrocarbons, were selected for their ability to grow in the presence of crude oil and produce biosurfactants, mainly by meaning of the production of good E24 values (≥50 %) and/or reduction in the surface tension (under 30 mN/m). Isolates belonged to bacterial genera that have been previously detected in hydrocarbon-contaminated environments, both natural and artificial. In particular, all strains affiliated to Rhodococcus genus were from the same sampling site and carbon source (MicroBy-C). Among them, 13 out of 14 isolates showed the high match with the same relative of the GenBank (KF571869, Rhodococcus sp. EG2). The occurrence of Actinobacteria members in cold habitats has been often reported as being independent of contamination, as they are generally present to a similar degree in pristine and contaminated sites (Whyte et al. 2002; Labbè et al. 2007). Cold-adapted hydrocarbon-degrading members in this genus have been previously isolated from marine Antarctic areas, and their biodegradative potential has often been established (Yakimov et al. 1999, 2004; De Domenico et al. 2004, Michaud et al. 2004). According to the results of phylogenetic determination, it is possible to assume that only two Rhodococcus species were isolated. On the other hand, after a careful observation of biochemical results and by considering the different responses to all the tests performed, we cannot exclude the affiliation to more than two species, thus explaining the differences at specie-specific level.
Among isolates, Pseudomonas and Rhodococcus are well-known biosurfactant producers (Syldatk et al. 1985; Desai and Banat 1997; Husain et al. 1997; Coelho et al. 2003; Maier 2003), but such capability was rarely reported for cold-adapted isolates (Yakimov et al. 1999; Pini et al. 2007; Cai et al. 2014). Our Antarctic isolate Rhodococcus sp. 176 was particularly efficient in biosurfactant production as it reduced the surface tension below 30 nM/n when grown in the presence of sunflower oil and produced an E24 value higher than 50 % growing in the presence of tetradecane at 4 °C. This confirms a previous suggestion by Lo Giudice et al. (2010) who observed a rapid emulsification of crude oil by of a Rhodococcus strain. Due to their enormous catabolic versatility, rhodococci appear to be an interesting target for the development of bioremediation techniques in cold environments (Bej et al. 2000; Saul et al. 2005; Ruberto et al. 2005).
To the best of our knowledge, biosurfactant production by Pseudoalteromonas and Idiomarina from cold environments is here reported for the first time. Strains within the former genus can produce extracellular polymeric substance (EPS), also in polar environments (Bozal et al. 1994; Mancuso et al. 2004, 2005). However, a unique study exists on the production of a polymeric emulsifying agent by a pseudoalteromonad under mesophilic conditions (Gutierrez et al. 2008). Interestingly, we recently reported on the production of biosurfactants by a Idiomarina isolate (strain A19) that was isolated from the Mediterranean Polychaete Megalomma claparedei (Rizzo et al. 2013). Further studies are in progress in order to analyse and compare biosurfactant activity of Idiomarina spp. A19 and 185 under different cultural conditions.
There are two primary pathways for bacterial BS biosynthesis: the way of hydrocarbons and the way of carbohydrates (Desai and Banat 1997). Choosing the carbon source is fundamental for the production of tensioactive molecules: the most commonly used includes carbohydrates, hydrocarbons and vegetable oils, and different studies have shown the influence of the carbon source on the chemical structure of biosurfactants (Putheti and Patil 2009; Kiran et al. 2010). Our results suggest that the analysed cold-adapted bacteria produced different types of biosurfactants also depending on the utilized substrata (tetradecane or sunflower oil) and incubation temperature. A simple hydrocarburic substrate (tetradecane) was used to select bacterial strains with good potentiality for study and application in the bioremediation field. On the other hand, the use of sunflower oil was introduced to evaluate the possible use of a low-cost alternative for BS production.
Bacterial isolates more efficiently reduced the surface tension when grown in the presence of tetradecane than sunflower oil. For example, Rhodococcus sp. 179 reported a value of 27.3 and 58.2 mN/m during incubation at 4 °C in presence of tetradecane and sunflower oil, respectively. Similar results were obtained for Rhodococcus spp. 181, 187 and 188 during incubation at 15 °C (Table 3). Conversely, the E24 index value was higher when bacteria grew in the presence of sunflower oil, as shown by Rhodococcus spp. 179 (E24 ≈ 37 % and ≈55 % in presence of tetradecane and sunflower oil, respectively), 181 (E24 ≈ 33 % and ≈57 in presence of tetradecane and sunflower oil, respectively) and 188 (E24 ≈ 3 % and ≈53 in presence of tetradecane and sunflower oil, respectively). This finding confirms the absence of a direct correlation between emulsifying and interfacial activity, as reported by Rizzo et al. (2013) and could be dependent on different types of biosurfactants, by considering the existence of different pathways for their production. Moreover, surface active substances may have different interfacial capabilities: biosurfactant can be divided into low molecular mass molecules, which reduce surface tension at air–water interface, and high molecular mass polymers, which have a stronger emulsifying activity (Rosenberg and Ron 1999). This suggests the possibility of an influence of the carbon source on the chemical nature of biosurfactant(s), so that isolates probably produce a low molecular molecule in presence of the hydrocarburic substrate and an high molecular one in presence of sunflower oil.
Haemolytic activity is often among the preferred methods to screen biosurfactant-producing strains thanks to its cheapness and speediness (Johnson and Boese-Marrazzo 1980; Carrillo et al. 1996; Moran et al. 2002). In this study, a number of strains showed haemolysis on BA plates, indicating the probable production of anionic surfactants. Nevertheless, such method should be considered as a useful integration of more efficient screening procedures. In fact, some biosurfactant producers gave a negative result to the haemolytic activity assay. This finding is in line with results previously obtained by other authors (Schulz et al. 1991; Youssef et al. 2004; Plaza et al. 2006; Rizzo et al. 2013) who reported the poor specificity of the method.
The application of TLC was useful to make the screening procedure more complete, by integrating the screening test array with a more specific analysis. Acid precipitation was preferred, because it is an easy, inexpensive and readily method available to recover crude BS such as surfactin, lipopeptide and glycolipid (Satpute et al. 2010). The TLC assay generally confirmed the results obtained from the screening tests and indicated the possible occurrence of glycolipidic components and fatty acids in BS molecules positively stained with anisaldehyde and molybdate/cerium sulphate staining reagents (Ellaiah et al. 2002; Satpute et al. 2010). In some cases, the absence of interesting spots on TLC plates was recorded for strains that showed emulsifying activity (i.e. Pseudoalteromonas sp. 93 in incubation with both tetradecane and sunflower oil; Pseudomonas sp. 280, and Rhodococcus sp. 192 during incubation with sunflower oil). This could be due to a probable different chemical nature of the tensioactive molecules, such as a peptidic one, or to high molecular weight biosurfactants, which could be not probably highlighted by the staining procedure used in this work. Another explanation could be a too low BS concentration in the bacterial culture.
In conclusion, we report for the first time on biosurfactant-producing bacteria from Arctic and Antarctic sediments, mainly affiliated to the genera Rhodococcus (14 isolates), reinforcing the idea that the microbial communities inhabiting cold environments possibly possess high biotechnological potentialities. Recently, studies aimed at detecting the most promising bacterial genera and the optimal conditions for BS production have assumed great importance, also for achieving consistent reduction in time and cost for industrial applications. In this context, the fact that most of the strains tested was able to produce biosurfactants in the presence sunflower oil, which is considered a good low-cost alternative, merits consideration. Finally, our strains resulted able to produce biosurfactants at two different temperatures, by suggesting the use of cold-adapted bacteria in both bioremediation and industrial applications. Further studies should be addressed to the improvement of BS activity (e.g. in terms of nutrients, pH, salinity), by taking into consideration also their thermal stability and chemical structure.
References
Batista SB, Mounteer AH, Amorim FR, Tótola MR (2006) Isolation and characterization of biosurfactant/bioemulsifier-producing bacteria from petroleum contaminated sites. Bioresour Technol 97:868–875
Bej AK, Saul D, Aislabie J (2000) Cold-tolerant alkane-degrading Rhodococcus species from Antarctica. Polar Biol 23:100–105
Bodour AA, Maier M (1998) Application of a modified drop-collapse technique for surfactant quantification and screening of biosurfactant-producing microorganisms. J Microbiol Meth 32:273–280
Bozal N, Manresa A, Castellvi J, Guinea J (1994) A new bacterial strain of Antarctica, Alteromonas sp., that produces a heteropolymer slime. Polar Biol 14:561–567
Cai Q, Zhang B, Chen B, Zhu Z, Lin W, Cao T (2014) Screening of biosurfactant producers from petroleum hydrocarbon contaminated sources in cold marine environments. Mar Poll Bull 86:402–410
Cameotra SS, Makkar RS, Kaur J, Mehta SK (2010) Synthesis of biosurfactants and their advantages to microorganisms and mankind. Adv Exp Med Biol 672:261–280
Carrillo PG, Mardaraz C, Pitta-Alvarez SJ, Giulietti AM (1996) Isolation and selection of biosurfactant-producing bacteria. World J Microbiol Biotechnol 12:82–84
Coelho J, Rivonkar CU, Bhavesh NS, Jothi M, Sagodkar UM (2003) Biosurfactant production by the quinoline degrading marine bacterium Pseudomonas sp. strain GU 104, and its effect on the metabolism of green mussel Perna viridis L. Indian J Mar Sci 32:202–207
Cooper DG, Zajic JE, Gerson DF (1979) Production of surface active lipids by Corynebacterium lepus. Appl Environ Microbiol 37:4–10
De Domenico M, Lo Giudice A, Michaud L, Saitta M, Bruni V (2004) Diesel oil and PCB-degrading bacteria isolated from Antarctic seawaters (Terra Nova Bay, Ross Sea). Polar Res 23:141–146
Deppe U, Richnow H-H, Michaelis W, Antranikian G (2005) Degradation of crude oil by an Arctic microbial consortium. Extremophiles 9:461–470
Desai JD, Banat IM (1997) Microbial production of surfactants and their commercial potential. Microbiol Mol Biol R 61:47–64
Ellaiah P, Prabhakar T, Sreekanth M, Taleb AT, Raju PB, Saisha V (2002) Production of glycolipids containing biosurfactant by Pseudomonas species. Indian J Exp Biol 40:1083–1086
Gerdes B, Brinkmeyer R, Dieckmann G, Helmke E (2005) Influence of crude oil on changes of bacterial community in Arctic sea-ice. FEMS Microbiol Ecol 53:129–139
Gutierrez T, Shimmield T, Haidon C, Black K, Green DH (2008) Emulsifying and metal ion binding activity of a glycoprotein exopolymer produced by Pseudoalteromonas sp. strain TG12. Appl Environ Microbiol 74:4867–4876
Husain DR, Goutx M, Acquaviva M, Gilewicz M, Bertrand JC (1997) The effect of temperature on eicosane substrate uptake modes by a marine bacterium Pseudomonas nautica strain 617: relationship with the biochemical content of cells and supernatants. World J Microbiol Biotechnol 13:587–590
Johnson MK, Boese-Marrazzo D (1980) Production and properties of heat stable extracellular hemolysin from Pseudomonas aeruginosa. Infect Immun 29:1028–1033
Kiran SG, Anto T, Selvin J, Sabarathnam B, Lipton AP (2010) Optimization and characterization of a new lipopeptide biosurfactant produced by marine Brevibacterium aureum MSA13 in solid state culture. Bioresour Technol 101:2389–2396
Labbè D, Margesin R, Schinner F, Whyte LG, Greer CW (2007) Comparative phylogenetic analysis of microbial communities in pristine and hydrocarbon-contaminated Alpine soils. FEMS Microbiol Ecol 59:466–475
Lang S (2002) Biological amphiphiles (microbial biosurfactants). Curr Opin Colloid Interface Sci 7:12–20
Lo Giudice A, Casella P, Caruso C, Mangano S, Bruni V, De Domenico M, Michaud L (2010) Occurrence and characterization of psychrotolerant hydrocarbon–oxidizing bacteria from surface seawater along the Victoria Land coast (Antarctica). Polar Biol 33:929–943
Lo Giudice A, Caruso C, Mangano S, Bruni V, De Domenico M, Michaud L (2012) Marine bacterioplankton diversity and community composition in an Antarctic coastal environment. Microb Ecol 63:210–223
Maier RM (2003) Biosurfactants: evolution and diversity in bacteria. Adv Appl Microbiol 52:101–121
Mancuso NCA, Garon S, Bowman JP, Raguenes G, Guezennec J (2004) Production of exopolysaccharides by Antarctic marine bacterial isolates. J Appl Microbiol 96:1057–1066
Mancuso NC, Lardiere SG, Bowman JP, Nichols PD, Gibson JAE, Guezennec J (2005) Chemical characterization of exopolysaccharides from Antarctic marine bacteria. Microb Ecol 49:578–589
Michaud L, Lo Giudice A, Saitta M, De Domenico M, Bruni V (2004) The biodegradation efficiency on diesel oil by two psychrotrophic Antarctic marine bacteria during a two-month-long experiment. Mar Poll Bull 49:405–409
Moran M, Martinez A, Sineriz F (2002) Quantification of surfactin in culture supernatant by hemolytic activity. Biotechnol Lett 24:177–180
Morita RY (1975) Psychrophilic bacteria. Bacteriol Rev 39:144–167
Pepi M, Cesarò A, Liut G, Baldi F (2005) An Antarctic psychrotrophic bacterium Halomonas sp ANT-3b, growing on n-hexadecane, produces a new emulsyfying glycolipid. Microb Ecol 53:157–166
Pini F, Grossi C, Nereo S, Michaud L, Lo Giudice A, Bruni V, Baldi F, Fani R (2007) Molecular and physiological characterisation of psychrotrophic hydrocarbon-degrading bacteria isolated from Terra Nova Bay (Antarctica). Eur J Soil Biol 43:368–379
Plaza GA, Ireneusz Z, Banat IM (2006) Use of different methods for detection ofthermophilic biosurfactant producing bacteria from hydrocarbon contaminated and bioremediated soils. J Petrol Sci Eng 50:71–77
Putheti RR, Patil MC (2009) Pharmaceutical formulation development of floating and swellable sustained drug delivery systems: a review. EJ Sci Technol 4:1–12
Rizzo C, Michaud L, Hörmann B, Gerçe B, Syldatk C, Hausmann R, De Domenico E, Lo Giudice A (2013) Bacteria associated with sabellids (Polychaeta: Annelida) as a novel source of surface active compounds. Mar Poll Bull 70:125–133
Rosenberg E, Ron EZ (1999) High and low molecular mass microbial surfactants. Appl Microbiol Biotechnol 52:154–162
Ruberto LAM, Vazquez S, Lobalbo A, Mac Cormack WP (2005) Psychrotolerant hydrocarbon-degrading Rhodococcus strains isolated from polluted Antarctic soils. Antarct Sci 17:47–56
Satpute SK, Bhawsar BD, Dhakephalkar PK, Chopade BA (2008) Assessment of different screening methods for selecting biosurfactant producing marine bacteria. Indian J Mar Sci 37:243–250
Satpute SK, Banpurkar AG, Dhakephalkar PK, Banat IM, Chopade BA (2010) Methods for investigating biosurfactants and bioemulsifiers: a review. Crit Rev Biotechnol 30:127–144
Saul DJ, Aislabie JM, Brown CE, Harris L, Foght JM (2005) Hydrocarbon contamination changes the bacterial diversity of soil around Scott Base, Antarctica. FEMS Microbiol Ecol 53:141–155
Schulz D, Passeri A, Schmidt M, Lang S, Wagner F, Wray V, Gunkel W (1991) Marine biosurfactants, I. Screening for biosurfactants among crude oil degrading marine microorganisms from the North Sea. Z Naturforsch 46:197–203
Syldatk C, Lang S, Wagner F, Wray V, Witte L (1985) Chemical and physical characterization of four interfacial-active rhamnolipids from Pseudomonas spec. DSM 2874 grown on n-alkanes. Z Naturforsch 40:51–60
Whyte LG, Schultz A, van Beilen JB, Luz AP, Pellizzari V, Labbè D, Greer CW (2002) Prevalence on alkane monooxygenase genes in Arctic and Antarctic hydrocarbon-contaminated and pristine soils. FEMS Microbiol Ecol 41:141–150
Yakimov MM, Giuliano L, Bruni V, Scarfì S, Golyshin PN (1999) Characterization of Antarctic hydrocarbon-degrading bacteria capable of producing bioemulsifiers. New Microbiol 22:249–259
Yakimov MM, Gentile G, Bruni V, Cappello S, D’Auria G, Golyshin PN, Giuliano L (2004) Crude oil-induced structural shift of coastal bacterial communities of Rod Bay (Terra Nova Bay, Ross Sea) and characterization of cultured cold-adapted hydrocarbonoclastic bacteria. FEMS Microbiol Ecol 49:419–432
Youssef N, Duncan KE, Nagle D, Savage KN, Knapp R, McInerney MJ (2004) Comparison of methods to detect biosurfactant production by diverse microorganisms. J Microbiol Methods 56:339–347
Acknowledgments
ALG wishes to thank the Antarctic Project “Limnopolar” (POL2006-06635) funded by the Spanish Ministry of Science and Technology, and all colleagues at the Byers camp for their logistic and friendly support to Luigi Michaud during his stay. RM wishes to thank all the colleagues at the Karlsruher Institut für Technologie (KIT), Germany, for assistance and support during her stay in their lab. This research was supported by grants by the project PolyArc (grant number ARCFAC-026129-2008-70) and the National Antarctic Museum (MNA).
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Malavenda, R., Rizzo, C., Michaud, L. et al. Biosurfactant production by Arctic and Antarctic bacteria growing on hydrocarbons. Polar Biol 38, 1565–1574 (2015). https://doi.org/10.1007/s00300-015-1717-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-015-1717-9