
Abstract
Since the onset of glaciation following the Oligocene (30–28 Ma), the prevalence of increasingly cold conditions has shaped the evolution of the Antarctic biota. Two hypotheses, postglacial recruitment from extra-regional locations and in situ persistence, have been proposed to explain the biogeography of the contemporary species-poor terrestrial Antarctic biota. Bryophytes, which form a major group of the Antarctic flora, exhibit a strong, inherent ability to survive cold conditions but also have high long-distance dispersal capacities, which are compatible with both hypotheses. Here, we test these hypotheses by means of population genetic and phylogeographic analyses of the cosmopolitan moss Bryum argenteum. We find evidence for at least three independent colonisation events of the species in Antarctica. Ancestral area reconstruction coupled with molecular dating suggests colonisation times of the different Antarctic clades ranging from four million years for the oldest lineage to half a million years for the youngest lineage. This suggests multiple colonisation events of Antarctica by this species during several glacial cycles within the Pleistocene, Pliocene and possibly late Miocene. This is the first study to demonstrate in situ persistence of bryophytes in Antarctica throughout previous glaciations.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The prevalence of increasingly cold conditions has shaped the evolution of the Antarctic biota since its separation from the other Southern Hemisphere continents during the final stages of the breakup of the supercontinent Gondwana (Convey and Stevens 2007; Convey et al. 2009; Fraser et al. 2012). Climate cooling was accompanied by a decrease in diversity of the angiosperm-dominated vegetation that inhabited the Antarctic Peninsula and parts of the continent during the Eocene. A mosaic of southern beech and conifer-dominated woodlands and tundra, comparable with that in parts of southern South America (Patagonia) today, characterised the region throughout the Oligocene (approximately 34–23 Ma). By the middle Miocene (approximately 16–11.6 Ma), localised pockets of tundra persisted until at least 12.8 Ma (Anderson et al. 2011). The transition to cold-based, alpine glacial regimes characterised by perennially dry and frozen conditions from 13.85 Ma has not subsequently been reversed (Lewis et al. 2008). Since the late Miocene, the Antarctic ice sheets have repeatedly thickened and advanced beyond their current limits onto the continental shelf. Ice sheets are believed to have overrun most currently ice-free areas during glaciations in the Pliocene (5–2.6 Ma) and the Pleistocene (2.6 Ma–10 Ka), including the Last Glacial Maximum (LGM; around 22–17 Ka) (Convey et al. 2009, and references therein; Mackintosh et al. 2013). The terrestrial biota remaining in Antarctica today is depauperate in terms of species diversity and is often cryptic. The major groups of organisms represented include micro-invertebrates, cryptogams and microbial groups (Convey 2013).
Two main hypotheses have been proposed to explain how the contemporary terrestrial Antarctic biota endured the glacial events of the Pleistocene (Convey and Stevens 2007; Convey et al. 2008). The first, and long-held but generally untested, view is that the large majority of the Antarctic terrestrial biota was eradicated from both the Antarctic continent and the associated offshore islands and archipelagos of the Scotia Arc (including sub-Antarctic South Georgia). The considerably expanded and thickened ice sheets would have caused a complete loss of terrestrial exposures and habitats during the glacial maxima, as is consistent with current glaciological model reconstructions. As a consequence, most or all current Antarctic terrestrial biota would have had to (re-)colonise the continent after the LGM, either from disjunct populations or from refugia. However, while the long-distance dispersal (LDD) capacity of bryophytes would potentially have facilitated (re)colonisation on such a timescale, the ability to disperse over long distances is apparently more limited for many other groups of Antarctic organisms. An additional weakness of the hypothesis is that many elements of the contemporary Antarctic biota show high levels of endemism (Øvstedal and Lewis Smith 2001; Pugh and Convey 2008), which could not have arisen after the LGM. The persistence of such an endemic biota would require refugia beyond the current confines of Antarctica (for instance, in the more distant sub-Antarctic islands or the other Southern Hemisphere continents), which would have been situated beyond the current distributions of many species. Therefore, this hypothesis would imply multiple colonisation events out of Antarctica when refugia were required, and subsequent extinction from them once Antarctica was recolonised. The alternative hypothesis is that species have survived in situ, in multiple refugia that must have been present in different regions across Antarctica. This hypothesis has received increasingly strong support in recent years from both molecular and classic biogeographical studies, as well as from geological and geomorphological evidence demonstrating the diachrony of ice-sheet expansions around Antarctica, and refining both the thickness and timing of previous episodes of maximum ice-sheet extent (Convey et al. 2008, 2009; Pugh and Convey 2008; Vyverman et al. 2010; Fraser et al. 2012).
Bryophytes (mosses and liverworts) are the dominant land plant flora in Antarctica, reaching their greatest diversity and extent in the Antarctic Peninsula and Scotia Arc (Ochyra et al. 2008; Convey 2013). As a group, they are generally regarded as possessing strong LDD capacities, supported by both direct (van Zanten 1978, 1981; Lönnel et al. 2012; Sundberg 2013; Lewis et al. 2014) and indirect (see Szövényi et al. 2012 for review) evidence. These characteristics would, in principle, equip them well for recolonisation of Antarctica following any episode of regional extinction. Elsewhere, recent evidence points to the major role of oceanic islands as glacial refugia for the subsequent (re-)colonisation of continents (Laenen et al. 2011; Hutsemékers et al. 2011). However, the geographic scale of Antarctic isolation from other land masses, along with protection from direct north–south transfer by atmospheric and ocean currents, gives the continent considerable geographic isolation (Barnes et al. 2006). Alternatively, a feature common among most bryophytes is their ability to grow at low (sub-optimal) temperatures. More than half of the 40 temperate species investigated by Furness and Grime (1982) showed a growth reduction of less than 50 % at 5 °C compared with growth at their optimal temperature, and this feature has also been described in Arctic and Antarctic bryophytes (Longton 1988). Indeed, many species, including some from the tropics, seem to be physiologically preadapted to cold and can survive temperatures ranging from −10 to −27 °C (Glime 2007). Recently, La Farge et al. (2013) have provided evidence for totipotent capacity (the ability of a cell to dedifferentiate into a meristematic state, and subsequently regrow) in Arctic bryophyte tissue buried by ice for 400 years, and Roads et al. (2014) have reported regrowth from gametophytes in moss banks preserved in permafrost for over 1.5 Ka. Furthermore, population genetic data for the temperate moss Homalothecium sericeum (Hedw.) Schimp. support persistence of the species in micro-refugia within the extensively glaciated northern Europe during the LGM (Désamoré et al. 2012). These examples suggest that bryophytes may be viable candidates to have survived Antarctic glacial cycles in situ.
Using the cosmopolitan moss Bryum argenteum Hedw. as a model, Hills et al. (2010) interpreted the lower genetic diversity observed in Antarctic vs non Antarctic samples as a consequence of a lower rate of DNA substitution and isolation in refugia within Victoria Land since the Pleistocene. However, in the absence of (1) evidence for heterogeneity of DNA substitution rate among lineages, (2) an explicit time frame, (3) representative sampling across the entire range of the species and (4) explicit analyses of population genetic structure, the hypothesis of survival in extra-Antarctic areas with subsequent (re-)colonisation cannot be excluded.
Here, we present a reconstruction of the phylogeography of B. argenteum at the global scale, and use molecular dating techniques to determine whether its presence in Antarctica is the result of recent (re-)colonisation from Pleistocene refugia outside Antarctica (H1), or whether it survived the Quaternary and Tertiary glaciations in situ (H2). If H1 holds true, we would expect Antarctic populations to be of recent, postglacial origin and therefore to show relatively little genetic differentiation from populations from other regions. Under that hypothesis, colonisation events might occur more frequently than under a scenario of long-term in situ persistence. Therefore, we would also expect under H1 a high gene flow from populations outside Antarctica, which could lead to a decrease in the signature of any founder effect. Conversely, if H2 holds true, we would predict that extant Antarctic populations derive from ancestors distributed on this continent before the LGM. We would further expect, provided that gene flow with the sub-Antarctic islands and other Southern Hemisphere areas has been limited, Antarctic populations to be genetically isolated from other regions and exhibit a clear phylogeographic signal (sensu Pons and Petit 1996).
Materials and methods
Specimen sampling and molecular protocols
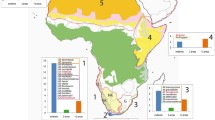
B. argenteum is a cosmopolitan, weedy moss species that thrives in disturbed environments. Its natural occurrence in Antarctica is indicated by its presence in the earliest botanical records for the continent and its widespread distribution within the regions where it occurs (Ochyra et al. 2008; Cannone et al. 2013). A total of 154 accessions of B. argenteum were sampled from Africa, America, Asia, Antarctica, the sub-Antarctic islands, Australasia and Europe (Appendix 1 in Supplementary material 1). From these accessions, 28 samples were taken from previously published papers (Hills et al. 2010; Pisa et al. 2013; Skotnicki et al. 2005) available in GenBank. The remaining samples were sequenced for this study and included 47 specimens collected by the authors and colleagues; all retained at the herbarium of the Universidad de Murcia, Spain (MUB), and 70 specimens held at the institutional herbaria of the British Antarctic Survey, UK (AAS); California Academy of Sciences, USA (CAS); Eszterházy Károly College, Hungary (EGR); Institute of Terrestrial Ecology, UK (ACHE); Main Botanical Garden of the Russian Academy of Sciences, Russia (MHA); Moscow State University, Russia (MW); New York Botanical Garden, USA (NY); Royal Botanic Garden Edinburgh, UK (E); Swedish Museum of Natural History, Sweden (S), University of Connecticut, USA (CONN), and the private herbaria of D.T. Holyoak and B. Goffinet. Four of the closely related species to B. argenteum (Wang and Zhao 2009) were selected as outgroup species (Appendix 1 in Supplementary material 1). The geographic location of the accessions is detailed in Fig. 1.

Geographic location of Bryum argenteum and outgroup accessions used
Total genomic DNA was extracted following the protocol described in Werner et al. (2002), or using the DNeasy Plant Mini Kit (Qiagen GmbH, Hilden, Germany). PCR amplification and sequencing of the nuclear internal transcribed spacer (ITS) region were performed using the protocol described in Pisa et al. (2013). Most of the amplifications were carried out using universal primers AB101 and AB102 (Douzery et al. 1999), with some nucleotide modifications to adapt these primers to B. argenteum. In some cases, sequences were generated using universal primers ITS-A and ITS-B as described in Blattner (1999), employing a similar PCR step as in Pisa et al. (2013), with exceptions being the use of the Taq PCR Core Kit (Qiagen GmbH, Hilden, Germany) and an annealing temperature of 50 °C. Forward and reverse sequence fragments for both ITS1 and ITS2 were edited and assembled using BioEdit 7.05 (Hall 1999), and every polymorphism was checked from the chromatograms. The sequences were aligned by eye, adding gaps where necessary to conserve homology between sequences (Appendix 2 in Supplementary material 2). Gaps were counted with SeqState (Müller 2005) using complex indel-coding. The number of polymorphic sites was calculated with DnaSP (Librado and Rozas 2009).
ITS remains the most widely used source of information on genetic variation at the species level in plants and fungi (reviewed by Nagy et al. 2012), although its use in phylogenetics has been questioned due to the potential presence of paralogs and pseudogenes (see Nieto Feliner and Rosselló 2007 for review). In mosses in particular, evidence for ITS paralogy was recently reported (Košnar et al. 2012). We consider that the use of ITS in B. argenteum was justified in the current study as no conflicting base calls during direct sequencing were observed, suggesting that the presence of intragenomic paralogous copies is unlikely.
Population genetic analyses
The sequences were grouped into haplotypes using DnaSP. For each of the seven geographic regions defined above, we calculated haploid diversity (h), unbiased haploid diversity by population (uh) and frequency of endemic haplotypes (x) using GENALEX 6.5 (Peakall and Smouse 2006) and Tajima’s D using Arlequin 3.5 (Excoffier et al. 2005). Genetic differentiation among geographic regions and presence of phylogeographic signal in the data were assessed by means of comparative analyses employing fixation index (F ST) and N ST. N ST is a measure of genetic differentiation among populations; it is analogous to F ST but takes the genetic distances among genotypes (here, the pairwise distance among them) into account (Pons and Petit 1996). When N ST > F ST, it means that mutation rates are higher than dispersal rates between geographical regions, generating a phylogeographic pattern. The significance of F ST and N ST were tested by constructing the distribution of the null hypothesis by means of 9,999 random permutations of individuals among geographic regions, as implemented by SPAGeDI 1.3 (Hardy and Vekemans 2002). The existence of a phylogeographic signal was tested by assessing the significance of the observed difference between N ST and F ST values by means of 9,999 random permutations of the allele distance matrix. Global F and N statistics among the seven geographic regions were computed, as well as pairwise statistics among regions. The correction of Benjamini and Yekutieli (2001) for multiple tests was applied to determine the significance of the pairwise statistics.
Phylogeny, molecular dating and ancestral area reconstructions
The phylogeny and divergence time among ITS genotypes within B. argenteum were investigated using BEAST v1.7.5 (Drummond et al. 2012). In the absence of fossil evidence, a prior on the absolute rate of molecular evolution was used, following the procedure described in Huttunen et al. (2008) and Aigoin et al. (2009). In the absence of a substitution rate for ITS in bryophytes, we used a normal distribution with a mean and standard deviation of 4.125 and 1.807e−3 substitutions per site per million years, respectively, which corresponds to the average absolute substitution rates of ITS across a wide range of annual herbaceous species (Kay et al. 2006). However, we consider that this rate is likely to overestimate substitution rates in mosses, which are longer lived and rely for a large part on asexual reproduction. This is particularly the case in B. argenteum, which is thought to be sterile in Antarctica (Ochyra et al. 2008). On average, the substitution rate of 18S rDNA, the neighbouring region of ITS, in mosses is suggested to be less than half that of vascular plants (Stenøien 2008). It is likely that the substitution rate used here may therefore be an overestimate of the true rate for this species and therefore that divergence times derived from this substitution rate may be significantly underestimated.
The Hasegawa, Kishino and Yano (HKY) model with gamma distribution and invariant sites had the best Bayesian information criterion (BIC) score for the ITS dataset using jModelTest 2.1.4 (Darriba et al. 2012). A relaxed clock with lognormal distribution was employed for the analysis. Before running the final dating analysis, the performance of five tree models (i.e. coalescent with constant size population, coalescent under an extended Bayesian skyline including the two linear and stepwise models, speciation under a birth–death process and speciation under Yule process) were compared using a model selection procedure based on Bayesian factors calculated in TRACER v1.5 (Rambaut and Drummond 2009). Overall, the model using the coalescent under a stepwise extended Bayesian skyline model (Heled and Drummond 2008) performed best (data not shown). Four independent Markov chain Monte Carlo (MCMC) analyses were each run for 100,000,000 generations for every model. Parameter values were sampled every 10,000 generations, and convergence and acceptable mixing of the samples were checked using the program TRACER v1.5. After discarding the burn-in steps (2,000 trees), the runs were combined to obtain an estimate of the posterior probability distributions of the dates of divergence.
To estimate ancestral areas, we used the maximum likelihood dispersal–extinction–cladogenesis (DEC) method (Ree et al. 2005; Ree and Smith 2008a) as implemented in the software Lagrange build 20091004 (Ree and Smith 2008b) on the BEAST chronogram. Each genotype was assigned to one or more of the seven geographic regions defined above. We conducted the analysis in Lagrange based on an unconstrained model permitting an equal probability of dispersal between all areas at any time.
Results
The complete alignment had a total length of 928 bp after the exclusion of the 5.8S rRNA gene. No further region was excluded from the alignment. There were 328 sites with gaps corresponding to 106 indels and 111 polymorphic sites. The alignment excluding outgroup sequences had a total length of 844 bp. There were 173 sites with gaps corresponding to 78 indels and 81 polymorphic sites allowing for the identification of 77 haplotypes (Appendix 1 in Supplementary material 1). Haploid diversity unbiased by population size was lowest in Antarctica (uh = 0.62) as compared to other regions (0.79–0.94) (Table 1). The frequency of endemic haplotypes exhibited the reverse trend, reaching its highest value (x = 0.90) in Antarctica. None of Tajima’s D statistics differed significantly from 0.
There was a significant difference in genotype frequency among geographic regions (Global F ST = 0.146, P < 0.0001). The global N ST (0.267, P < 0.0001) was significantly higher than F ST (P < 0.0001) providing evidence that, on average, the genotypes from the same region were more closely related than the genotypes from different regions. This geographic structure was largely due to the significant genetic isolation of Antarctica. The phylogeographic signal between Antarctica and any of the other six regions was consistently significant, whereas a significant phylogeographic signal could not be detected among any other pairs of regions (Table 2).
Accessions from Antarctica belonged to three clades (Fig. 2). Clade I was mainly composed of Antarctic genotypes, with the exception of one European genotype and one common genotype shared between Antarctica, Europe, Asia and America. Clade II was composed of Antarctic, sub-Antarctic, American and Australasian genotypes. Clade III was composed of Antarctic and American genotypes. The Lagrange analysis indicated that the most recent common ancestor of clade I, which may have been distributed across Asia, Europe and America, colonised Antarctica 4.36 Ma (Highest Posterior Density, HPD, 1.79–14.72). In clades II and III, the earliest colonisation of Antarctica dates back to 1.43 Ma (HPD 0.42–4.97) and 0.55 Ma (HPD 0.13–1.97), respectively, from an ancestor most closely related to American populations. Potential dispersal events from Antarctica to other regions are not excluded as genotypes in clade II located in Australasia and the sub-Antarctic islands come from ancestors distributed across Antarctica and America.

Spatial and temporal dimensions of Bryum argenteum evolution. Chronogram of the fifty per cent majority rule consensus of the trees sampled from the posterior probability distribution generated by the BEAST analysis of ITS sequences of the B. argenteum genotypes sampled worldwide. The maximum likelihood reconstruction of geographical range evolution under the unconstrained dispersal–extinction–cladogenesis (DEC) model (ln L = 214.9) implemented in Lagrange is given in boxes at each node of interest. The geographical areas defined for this study and each haplotype are provided. The two series of reconstructions at each internal node indicate the ML ancestral range estimate for the upper and lower branch connecting that node, respectively. Boxes at terminal nodes indicate the geographic areas occupied by the accessions belonging to each haplotype. The vertical bars (I, II and III) indicate the clades with Antarctic genotypes. Thicker branches indicate the support for the Antarctic clades (PP > 0.9). * indicates the support for clades with PP > 0.9)
Discussion
Evidence for at least three distinct origins of B. argenteum in Antarctica was found, with colonisation times ranging between approximately 4.4 Ma (clade I), 1.4 Ma (clade II) and 0.6 Ma (clade III). However, as mentioned above, the substitution rate of Kay et al. (2006) is based on studies of annual herbaceous species and is likely to be much higher than in mosses. A study on relative substitution rates amongst major plant groups showed that, on average, the substitution rate of 18 s rDNA, the neighbouring region of ITS, is more than twice as high in vascular plants compared with mosses (Stenøien 2008). This suggests that the presence of B. argenteum in Antarctica may be considerably more ancient than estimated here. The current study therefore provides the first support for the long-term persistence in situ of a bryophyte species in Antarctica, with timescales in the order of millions of years. No evidence supporting strict postPleistocene (i.e. recent) colonisation (H1) was found in any of the lineages. Our results, however, do not exclude potential dispersal events from Antarctica to other regions and future studies with a larger sample size and obtained from more locations may identify evidence for recent colonisation events.
In agreement with our finding of long-term persistence of B. argenteum in Antarctica, a significant phylogeographical signal was found in all pairwise comparisons between Antarctica and each of the six other global regions, while no such signal was identified amongst the latter. This indicates that extant patterns of genetic diversity of Antarctic B. argenteum populations are better explained in terms of in situ diversification than recruitment of migrants from other areas, resulting in the highest proportion of endemic haplotypes as compared to other regions of the world. Such an interpretation is consistent with recent developments in biogeographical knowledge of much of the contemporary terrestrial biota in Antarctica. Evidence for long-term history in situ has been reported in all major groups except the bryophytes, with timescales ranging from mid-Pleistocene (e.g. diatoms, rotifers, cladocerans) to Pliocene, Miocene and Gondwana-breakup (e.g. springtails, chironomid midges, mites, copepods, microorganisms) (see Convey et al. 2008, 2009, and references therein; Vyverman et al. 2010). McGaughran et al. (2010), in a comparative phylogeographic study of different springrail (Collembola) species in Victoria Land and along the Scotia Arc and Antarctic Peninsula, identified analogous evidence of intraregional differentiation and hence colonisation patterns on timescales dating back to at least the earliest Pleistocene.
How and where bryophytes and other terrestrial biota could have survived through glaciation events within Antarctica is not yet well understood. The lowest ITS haplotypic diversity within B. argenteum worldwide was observed in Antarctica, as in the cosmopolitan moss Ceratodon purpureus Hedw (Brid.) (Clarke et al. 2008, 2009). Although the hypothesis of lower mutation rates in Antarctic B. argenteum populations cannot be ruled out (Hills et al. 2010), such a low regional genetic diversity can also be interpreted in terms of the sterile condition of B. argenteum in Antarctica (Ochyra et al. 2008), either due to the regional absence of one of the sexes in this dioicous species, and/or inhibition of sex expression due to prevailing cold and dry conditions (Longton 1988). Yet, analyses of patterns of genetic diversity in Antarctic populations of B. argenteum failed to evidence a significant bottleneck. This observation does not support the hypothesis of a substantial past decrease in population size and points to the persistence of sufficiently large and numerous populations of the species through time. In Antarctica, areas of heated ground associated with geothermal activity, where B. argenteum is known to occur (Convey et al. 2000), may be particularly relevant in considering the locations of some potential refuge sites (Convey and Lewis Smith 2006, Fraser et al. 2014). Although individually ephemeral, the presence of geothermal habitats may have extended over considerable time periods, as volcanism has been widespread throughout the Tertiary in parts of the northern Antarctic Peninsula and elsewhere in Antarctica (Baker et al. 1975, Convey et al. 2000; Convey and Lewis Smith 2006; Fraser et al. 2014). Therefore, geothermal habitats might have played a key role in the longer-term regional persistence of species with rapid colonisation capacities such as bryophytes, allowing survival through periods of apparently greater environmental extremes than are currently experienced (Convey and Lewis Smith 2006).
Conclusion
This study demonstrates for the first time in situ persistence of bryophytes in Antarctica throughout previous glacial cycles and contradicts the hypothesis of postglacial recruitment from extra-regional locations. Although B. argenteum, like other moss species, exhibits the lowest levels of genetic diversity worldwide in Antarctica, ITS variation is substantial within and among Antarctic populations, making it a highly suitable model for investigating fine-scale patterns of genetic structure and diversity at continental scale in order to reconstruct its biogeographic history. In particular, detailed phylogeographic information at the Antarctic scale would allow for testing Fraser’s et al. (2014) hypothesis that refugia indeed correspond to areas of high geothermal activity and contribute, together with information from other terrestrial organisms (Terauds et al. 2012), to enhancing the identification of bioregions that are not fully represented in the current Antarctic Specially Protected Area network.
References
Aigoin DA, Devos N, Huttunen S, Ignatov MS, González-Mancebo JM, Vanderpoorten A (2009) And if Engler was not completely wrong? Evidence for multiple evolutionary origins in the moss flora of Macaronesia. Evolution 63:3248–3257
Anderson JB, Warnyb S, Askinc RA, Wellnerd JS, Bohatye SM, Kirshnera AE, Livseyf DN, Simmsf AR, Smitha TR, Ehrmanng W, Lawverh LA, Barbeaui D, Wisej SW, Kulhenekj DK, Weavera FM, Majewskik W (2011) Progressive Cenozoic cooling and the demise of Antarctica’s last refugium. Proc Natl Acad Sci USA 108:11356–11360
Baker PE, McReath I, Harvey MR, Roobol MJ, Davies TG (1975) The geology of the South Shetland Islands: V. Volcanic evolution of Deception Island. Br Antarct Surv Sci Rep 78:1–8
Barnes DKA, Hodgson DA, Convey P, Allen C, Clarke A (2006) Incursion and excursion of Antarctic biota: past, present and future. Global Ecol Biogeogr 15:121–142
Benjamini Y, Yekutieli D (2001) The control of false discovery rate under dependency. Ann Stat 29:1165–1188
Blattner FR (1999) Direct amplification of the entire ITS region from poorly preserved plant material using recombinant PCR. BioTech 27:1180–1185
Cannone N, Convey P, Guglielmin M (2013) Diversity trends of bryophytes in continental Antarctica. Polar Biol 36:259–271
Clarke LJ, Ayre DJ, Robinson SA (2008) Somatic mutation and the Antarctic ozone hole. J Ecol 96:378–385
Clarke LJ, Ayre DJ, Robinson SA (2009) Genetic structure of East Antarctic populations of the moss Ceratodon purpureus. Antarct Sci 21:51–58
Convey P (2013) Antarctic ecosystems. In: Levin SA (ed) Encyclopedia of biodiversity, vol 1, 2nd edn. Elsevier, San Diego, pp 179–188
Convey P, Lewis Smith RI (2006) Geothermal bryophyte habitats in the South Sandwich Islands, maritime Antarctic. J Veg Sci 17:529–538
Convey P, Stevens MI (2007) Antarctic biodiversity. Science 317:1877–1878
Convey P, Lewis Smith RI, Hodgson DA, Peat HJ (2000) The flora of the South Sandwich Islands, with particular reference to the influence of geothermal heating. J Biogeog 27:1279–1295
Convey P, Gibson J, Hillenbrand C-D, Hodgson DA, Pugh PJA, Smellie JL, Stevens MI (2008) Antarctic terrestrial life—challenging the history of the frozen continent? Biol Rev 83:103–117
Convey P, Stevens MI, Hodgson DA, Smellie JL, Hillenbrand C-D, Barnes DKA, Clarke A, Pugh PJA, Linse K, Cary SC (2009) Exploring biological constraints on the glacial history of Antarctica. Quat Sci Rev 28:3035–3048
Darriba D, Taboada GL, Doallo R, Posada D (2012) jModelTest 2: more models, new heuristics and parallel computing. Nat Methods 9:772
Désamoré A, Laenen B, Stech M, Papp B, Hedenäs L, Mateo RG, Vanderpoorten A (2012) How do temperate bryophytes face the challenge of a changing environment? Lessons from the past and predictions for the future. Glob Change Biol 18:2915–2924
Douzery EJP, Pridgeon AM, Kores P, Linder HP, Kurzweil H, Chase MW (1999) Molecular phylogenetics of Disease (Orchidaceae): a contribution from nuclear ribosomal ITS sequences. Am J Bot 86:887–899
Drummond AJ, Suchard MA, Xie D, Rambaut A (2012) Bayesian phylogenetics with BEAUti and the BEAST 1.7. Mol Biol Evol 29:1969–1973
Excoffier L, Laval G, Schneider S (2005) Arlequin (version 3.0): an integrated software package for population genetics data analysis. Evol Bioinform Online 1:47–50
Fraser CI, Nikula R, Ruzzante DE, Waters JM (2012) Poleward bound: biological impacts of Southern Hemisphere glaciation. Trends Ecol Evol 27:462–471
Fraser CI, Terauds A, Smellie J, Convey P, Chown SL (2014) Geothermal activity helps life survive ice ages. Proc Natl Acad Sci USA 111:5634–5639
Furness SB, Grime JP (1982) Growth rate and temperature responses in bryophytes II. A comparative study of species of contrasted ecology. J Ecol 70:525–536
Glime JM (2007) Bryophyte ecology. Volume 1. Physiological ecology. Ebook sponsored by Michigan Technological University and the International Association of Bryologists. http://www.bryoecol.mtu.edu/. Accessed 6 Feb 2014
Hall TA (1999) BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser 41:95–98
Hardy OJ, Vekemans X (2002) SPAGeDI: a versatile computer program to analyse spatial genetic structure at the individual or population levels. Mol Ecol Notes 2:618–620
Heled J, Drummond A (2008) Bayesian inference of population size history from multiple loci. BMC Evol Biol 8:289
Hills SFK, Stevens MI, Gemmill CEC (2010) Molecular support for Pleistocene persistence of the continental Antarctic moss Bryum argenteum. Antarct Sci 22:721–726
Hutsemékers V, Szövényi P, Shaw AJ, González-Mancebo JM, Muñoz J, Vanderpoorten A (2011) Oceanic islands are not sinks of biodiversity in spore-producing plants. Proc Natl Acad Sci USA 108:18989–18994
Huttunen S, Hedenäs L, Ignatov MS, Devos N, Vanderpoorten A (2008) Origin and evolution of the northern hemisphere disjunction in the moss genus Homalothecium (Brachytheciaceae). Am J Bot 95:720–730
Kay KM, Whittall JB, Hodges SA (2006) A survey of nuclear ribosomal internal transcribed spacer substitution rates across angiosperms: an approximate molecular clock with life history effects. BMC Evol Biol 6:36
Košnar J, Herbstová M, Kolář F, Koutecký P, Kučera J (2012) A case study of intragenomic ITS variation in bryophytes: assessment of gene flow and role of polyploidy in the origin of European taxa of the Tortula muralis (Musci: Pottiaceae) complex. Taxon 61:709–720
La Farge C, Krista H, Williams KH, England JH (2013) Regeneration of Little Ice Age bryophytes emerging from a polar glacier with implications of totipotency in extreme environments. Proc Natl Acad Sci USA. doi:10.1073/pnas.1304199110
Laenen B, Désamoré A, Devos N, Shaw AJ, González-Mancebo JM, Carine MA, Vanderpoorten A (2011) Macaronesia: a source of hidden genetic diversity for post-glacial recolonisation of western Europe in the leafy liverwort Radula lindenbergiana. J Biogeogr 38:631–639
Lewis AR, Marchant DR, Ashworth AC, Hedenäs L, Hemming SR, Johnson JV, Lengh MJ, Machlus ML, Newton AE, Raine JJ, Willenbring JK, Williams M, Wolfe AP (2008) Mid-Miocene cooling and the extinction of tundra in continental Antarctica. Proc Natl Acad Sci USA 105:10676–10678
Lewis LR, Behling E, Gousse H, Qian E, Elphick CS, Lamarre J-F, Bêty J, Liebezeit J, Rozzi R, Goffinet B (2014) First evidence of bryophyte diaspores in the plumage of transequatorial migrant birds. Peer J 2:e424. doi:10.7717/peerj.424
Librado P, Rozas J (2009) DnaSP v5: a software for comprehensive analysis of DNA polymorphism data. Bioinformatics 25:1451–1452
Longton R-E (1988) The biology of polar bryophytes and lichens. Cambridge University Press, Cambridge
Lönnel N, Hylander K, Jonsson BG, Sundberg S (2012) The fate of the missing spores—patterns of realized dispersal beyond the closest vicinity of a sporulating moss. PLoS One 7:e41987
Mackintosh AN, Verleyen E, O’Brien PE, White DA, Jones RS, McKay R, Dunbar R, Gore DB, Fink D, Post AL, Miura H, Leventer A, Goodwin I, Hodgson DA, Lilly K, Crosta X, Golledge NR, Wagner B, Berg S, van Ommen T, Zwartz D, Roberts SJ, Vyverman W, Masse G (2013) Retreat history of the East Antarctic Ice Sheet since the last glacial maximum. Quat Sc Rev. doi:10.1016/j.quascirev.2013.07.024
McGaughran A, Toricelli G, Carapelli A, Frati F, Stevens MI, Convey P, Hogg ID (2010) Contrasting phylogeographic patterns for springtails reflect different evolutionary histories between the Antarctic Peninsula and continental Antarctica. J Biogeogr 37:103–119
Müller K (2005) SeqState. Primer design and sequence statistics for phylogenetic DNA datasets. Appl Bioinform 4:65–69
Nagy LG, Kocsubé S, Csanádi Z, Kovács GM, Petkovits T, Vágvölgyi C, Papp T (2012) Re-mind the gap! Insertion—deletion data reveal neglected phylogenetic potential of the nuclear ribosomal internal transcribed spacer (ITS) of Fungi. PLoS One 11:e49794
Nieto Feliner G, Rosselló JA (2007) Better the devil you know? Guidelines for insightful utilization of nrDNA ITS in species-level evolutionary studies in plants. Mol Phylogenet Evol 44:911–919
Ochyra R, Lewis Smith RI, Bednarek-Ochyra H (2008) The illustrated moss flora of Antarctica. Cambridge University Press, Cambridge
Øvstedal DO, Lewis Smith RI (2001) Lichens of Antarctica and South Georgia. A guide to their identification and ecology. Cambridge University Press, Cambridge
Peakall R, Smouse PE (2006) GENALEX 6: genetic analysis in Excel. Population genetic software for teaching and research. Mol Ecol Notes 6:288–295
Pisa S, Werner O, Vanderpoorten A, Magdy M, Ros RM (2013) Elevational patterns of genetic variation in the cosmopolitan moss Bryum argenteum (Bryaceae). Am J Bot 100:2000–2008
Pons O, Petit RJ (1996) Measuring and testing genetic differentiation with ordered versus unordered alleles. Genetics 144:1237–1245
Pugh PJA, Convey P (2008) Surviving out in the cold: antarctic endemic invertebrates and their refugia. J Biogeogr 35:2176–2186
Rambaut A, Drummond AJ (2009) Tracer. Version 1.5. Molecular evolution, phylogenetics and epidemiology Web. http://tree.bio.ed.ac.uk/software/tracer/. Accessed 10 Dec 2013
Ree RH, Smith SA (2008a) Maximum likelihood inference of geographic range evolution by dispersal, local extinction, and cladogenesis. Syst Biol 57:4–14
Ree RH, Smith SA (2008b) Lagrange: likelihood analysis of geographic range evolution. Version 2.0. http://code.google.com/p/lagrange/. Accessed 11 Dec 2013
Ree RH, Moore BR, Webb CO, Donoghue MJ (2005) A likelihood framework for inferring the evolution of geographic range on phylogenetic trees. Evolution 59:2299–2311
Roads E, Longton RE, Convey P (2014) Millenial timescale regeneration in a moss from Antarctica. Curr Biol 24:222–223
Skotnicki ML, Mackenzie AM, Clements MA, Selkirk PM (2005) DNA sequencing and genetic diversity of the 18S–26S nuclear ribosomal internal transcribed spacers (ITS) in nine Antarctic moss species. Antarct Sci 17:377–384
Stenøien HK (2008) Slow molecular evolution in 18S rDNA, rbcL and nad5 genes of mosses compared with higher plants. J Evol Biol 21:566–571
Sundberg S (2013) Spore rain in relation to regional sources and beyond. Ecography 36:364–373
Szövényi P, Sundberg S, Shaw AJ (2012) Long-distance dispersal and genetic structure of natural populations: an assessment of the inverse isolation hypothesis in peat mosses. Mol Ecol 21:5461–5472
Terauds A, Chown SL, Morgan F, Peat HJ, Watts DJ, Keys H, Convey P, Bergstrom DM (2012) Conservation biogeography of the Antarctic. Divers Distrib 18:726–741
van Zanten BO (1978) Experimental studies on trans-oceanic long-range dispersal of moss spores in the Southern Hemisphere. J Hattori Bot Lab 44:455–482
van Zanten BO, Pócs T (1981) Distribution and dispersal of bryophytes. Adv Bryol 1:479–562
Vyverman W, Verleyen E, Wilmotte A, Hodgson DA, Willems A, Peeters K, Van de Vijver B, de Wever A, Leliaert F, Sabbe K (2010) Evidence for widespread endemism among Antarctic micro-organisms. Polar Sci 4:103–113
Wang C-Y, Zhao J-C (2009) Phylogeny of Ptychostomum (Bryaceae, Musci) inferred from sequences of nuclear ribosomal DNA internal transcribed spacer (ITS) and chloroplast rps4. J Syst Evol 47:311–320
Werner O, Ros RM, Guerra J (2002) Direct amplification and NaOH extraction: two rapid and simple methods for preparing bryophyte DNA for polymerase chain reaction (PCR). J Bryol 24:127–131
Acknowledgments
The authors thank the curators of the herbaria AAS, ACHE, CAS, CONN, E, EGR, MHA, MW, NY, S and also the private herbaria of D.T. Holyoak and B. Goffinet for the loan of material and to B. Albertos, R. Garilleti, B. Goffinet, J. M. González-Mancebo, F. Lara and M. Stech for collecting material used in this work. We also thank Oliva Martin-Sanchez for producing Fig. 1. This study has been supported financially by the Spanish Ministry of Science and Innovation (Projects CGL2008-00275/BOS and CGL2011-22936/BOS) and by the European Regional Development Funds. E. M. Biersma is supported by a Natural Environment Research Council PhD studentship (ref NE/K50094X/1), and P. Convey by Natural Environment Research Council core funding to the British Antarctic Society programme ‘Environmental Change and Evolution’. This paper also contributes to the Scientific Committee on Antarctic Research ‘State of the Antarctic Ecosystem’ programme. J. Patiño and A. Vanderpoorten gratefully acknowledge financial support from the Belgian Funds for Scientific Research and the University of Liege. We thank three reviewers for helpful comments.
Author information
Authors and Affiliations
Corresponding author
Additional information
S. Pisa and E. M. Biersma have contributed equally to this paper.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article

Cite this article
Pisa, S., Biersma, E.M., Convey, P. et al. The cosmopolitan moss Bryum argenteum in Antarctica: recent colonisation or in situ survival?. Polar Biol 37, 1469–1477 (2014). https://doi.org/10.1007/s00300-014-1537-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-014-1537-3