
Abstract
Antarctica has several apparent advantages for the study of biodiversity change along latitudinal gradients including a relatively pristine environment and simple community structures. Published analyses for lichens and mosses show no apparent gradient in biodiversity along the western Ross Sea coast line, the longest ice-free area in Antarctica spanning 14° latitude. One suggestion is that the area remains poorly surveyed. Here, we combine available species lists from four sites along the coast with new own data from two additional sites [Taylor Valley (77°30′S) and Diamond Hill (79°S)]. We show a decline in total terrestrial biodiversity with latitude from Cape Hallett (72°S) to Diamond Hill. However, the southernmost site, the Queen Maud Mountains (84°S), is exceptional with almost the same diversity as Cape Hallett. A categorization of lichens according to their proposed ecology shows the proportion of tolerant species remains relatively constant. However, the absolute number of conformant species declines with latitude, again with a minimum at Diamond Hill. Similarity indices are low and not very different between sites with Diamond Hill being the exception with very few species. We suggest that terrestrial biodiversity best reflects microhabitat water availability rather than macroclimatic temperature changes and use climate data from Taylor Valley and Diamond Hill to support this suggestion. We propose that the importance of microhabitats and landscape location is one of several possible limitations to the application of bioclimatic modeling along the Ross sea coastline. In the absence of a definitive link between macroclimate and the biota, predicting the effects of climate changes will be more challenging.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Natural distributional patterns of species can serve as a basis for modeling strategies to predict the potential impacts of climate change. Although other factors in addition to climate, such as biotic interactions, evolutionary change, and dispersal ability, can play an important role in determining species distribution, the bioclimate envelope-concept, correlating current species distributions with climate variables (Heikinnen et al. 2006), is a useful approximation for the impacts of climate on biodiversity (Pearson and Dawson 2003). Antarctica would seem to be a suitable place for applying bioclimatic models for several reasons. First, terrestrial systems appear to be predominantly abiotically driven (Hogg et al. 2006) so that there should be a direct linkage between climate and biota. Second, other than two species of higher plants that occur only in the north-western Antarctic Peninsula, the vegetation is composed entirely of cryptogamic organisms (lichens and mosses, Ochyra et al. 2008). Accordingly, community structures are relatively simple making biotic interactions more tractable in comparison with lower latitudes. Predacious species are few (e.g., some tardigrades), and the largest year-round terrestrial animals are the fungivorous springtails (Collembola) and mites (Acari) (Adams et al. 2006).
Two major research programs, Evolution and Biodiversity in the Antarctic (EBA), a Scientific Committee on Antarctic Research (SCAR) initiative, and Latitudinal Gradient Project (LGP) have targeted these features as a basis for examining Antarctic species distribution and predicting the consequences of global climate changes for Antarctica. Specifically, the LGP focused on the latitudinal gradient along the Ross Sea coastline and used it as a proxy for a climate gradient (Howard-Williams et al. 2006, 2010), providing an excellent natural laboratory to investigate potential impacts of climate on terrestrial organisms (De Frenne et al. 2013).
A recent analysis of terrestrial biodiversity and macroclimate has shown sound correlations between the number of lichen and bryophyte species present with mean annual temperature at latitudes north of the Ross Sea, although little correlation within the Ross Sea region (Green et al. 2011a). This lack of correlation between terrestrial biodiversity and latitude along the Ross Sea coast was also noted by Peat et al. (2007), who suggested that the region may still not be sufficiently surveyed for terrestrial biota and that this deficiency could be concealing any overall pattern. Since then, data for terrestrial species within the Ross Sea region have improved considerably with species lists now available for a range of sites including Cape Hallett (72 °C, Green, unpubl. data), Terra Nova Bay (74°S, Castello 2003), Botany Bay (77°S, Seppelt et al. 2010), Lake Wellman (79°S, Magalhães et al. 2012), and the Queen Maud Mountains area (84°S, Green et al. 2011b). Biological soil crusts that are small scale communities mainly composed of lichens, mosses, algae, cyanobacteria, and fungi have also recently been reported from the Diamond Hill region and Garwood Valley (Colesie et al. 2013) and appear to represent a major proportion of the lichen and moss communities in the area.
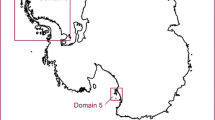
In order to further assess patterns of diversity along the Ross Sea coastline, we combine these existing data with new data from two additional sites intending to identify correlations with climate. The first of the two additional sites is lower Taylor Valley, which lies within the Dry Valleys, and located roughly at the center of the Ross Sea latitudinal gradient (77°30′S; Fig. 1). The Dry Valleys contain about 4,800 km2 of ice-free land, or ~10 % of total ice-free land on the continent, and are relatively well studied, with Taylor Valley (Fig. 1), being a Long-Term Ecological Research (LTER) site. However, only partial biodiversity surveys are available (e.g., Freckman and Virginia 1997; Powers et al. 1998). Taylor Valley runs east–west and has been previously categorized into three soil moisture zones determined largely by elevation and climate influences: subxerous, xerous, and ultraxerous (Campbell and Claridge 1982). The subxerous zone is the warmest and wettest and is nearest to the coast, while the other two areas lie inland and are much drier and colder. These inland (slope) parts of the Dry Valleys are often suggested to be the nearest equivalent on Earth to the environment on Mars (Wynn-Williams and Edwards 2000; Tamppari et al. 2012). The second site is Diamond Hill in the Brown Hills region (79°S) (Fig. 1), lying roughly 220 km south of the Dry Valleys and on the westernmost part of the Ross Sea coastline (Barrett et al. 2006; Storey et al. 2010).

Map of the western Ross Ice Shelf coast, showing the locations of Cape Hallett, Terra Nova Bay, Botany Bay, Taylor Valley, Diamond Hill, and the Queen Maud Mountains
Here, we add new biodiversity data for the lower Taylor Valley, Dry Valleys (77°37′S), and Diamond Hill, Brown Hills, Darwin region (79°50′S) as well as climate data obtained from automatic weather stations (AWS) at the survey sites. These sites are comparable as both are part of the coastal semidesert zones with mean monthly temperatures >0 °C for 0–2 month a year (Ochyra et al. 2008). We surveyed the species composition of vegetation and invertebrates at both sites and included a comparison with other regions based on published and unpublished data. Our aims were to (1) determine linkages between terrestrial biodiversity (mosses, lichens, algae, cyanobacteria, and arthropods) and the local climate; (2) compare the biodiversity at the two sites with others along the Ross Sea coastline and show if the additional data result in an even biodiversity gradient; and (3) consider the application of bioclimatic models and potential responses of the terrestrial biota to future environmental changes.
Materials and methods
Study areas
Both research sites are situated on the west coast of the Ross Sea (Fig. 1). The lower Taylor Valley research site (77°37′S, 162°59′E) is the eastern, lower-most part of the Valley from approximately Canada Glacier to the coast. For further details see Fountain et al. (1999). The Brown Hills site was at Diamond Hill (79°50′S, 159°19′E), which is close to the coast and appears, from our surveys, to be one of the richer areas for biodiversity in the area (Fig. 1). For further details see Colesie et al. (2013).
Biotic diversity
Data were collected at both sites using detailed visual surveys and collections spread across the accessible landscape, followed by curation and identification of specimens by traditional and molecular methodologies. Surveys were completed over five visits (total of 19 weeks) at Taylor Valley and two visits (total of 3 weeks) at Brown Hills and included 3–4 observers in each case. Invertebrates were identified using appropriate taxonomic keys (Wise 1964, 1967, 1971; Strandtmann 1967) and confirmed using mitochondrial DNA (COI) “barcoding” (Hebert et al. 2003). The moss species were determined by R. Seppelt (Tasmanian Herbarium, Hobart, Australia), and names are according to Ochyra et al. (2008). Lichen species were determined using the identification guide of lichens in Øvstedal and Smith (2001). Cyanobacteria and algae were identified by light microscopy of soil samples and appropriate taxonomic keys (e.g., Ettl and Gärtner 1995; Komárek and Anagnostitis 1999; Komárek and Anagnostitis 2005). Samples of lichens and mosses are archived at the private herbarium of Roman Türk and/or the University of Tasmania Herbarium, code HO (R. Seppelt). All species were categorized according to their substrate/ecology after Øvstedal and Smith (2001) into two life-history traits. Species that were confined to narrow habitats (sheltered, endolithic, hypolithic, associated with mosses, influenced by birds, growing in crevices, on perpetually flooded areas, on seal bones) were called conformants (C). All mosses, green algae, cyanobacteria, collembolans, and mites are classified as conformants, because of their strong link to high water availability. The majority of lichens were also conformants, but some were classified as tolerant (T) because they occupied broad, widely spread habitats and were tolerant to exceptionally cold, dry, and windy conditions. The tolerant group is roughly equivalent to the widespread group of lichens described in Øvstedal and Smith (2001).
Diversity was compared using lists of taxa recorded at other coastal and slope sites within Southern Victoria Land. These species lists were generated from Castello (2003) for Terra Nova Bay (74°S), Seppelt et al. (2010) for Botany Bay (77°S) and Green et al. (2011b) and Sancho (unpubl. data) for the Queen Maud Mountains area (84°S). Lichen and bryophyte numbers for Cape Hallett (72°S) were obtained from unpublished data (Sancho and Green). Only one slope site was described, which was from Magalhães et al. (2012) for the Lake Wellman area (79°S). Correlation between latitude and species richness was computed with R 2 as coefficient of determination.
To assess the success of the sampling effort, we analyzed the relationship between the number of sampling locations and the number of lichen species recorded. A site-based rarefaction curve with 95 % confidence intervals was computed using the software EstimateS (Colwell 2006). The nonparametric estimator Chao2 was used to estimate total species richness of all the sites. The similarity of the lichen floristic composition was quantified using the Jaccard similarity index, J = a/(a + b + c) in which a is the number of shared species for the two sites, and b and c are the number of species unique to each of the individual sites. Mean similarity indices of each site were then compared with all other sites with Student’s t test (STATISTICA 10, StatSoft (Europe) GmbH, Hamburg).
To evaluate the similarity between the sites, we also performed nonmetric multidimensional scaling (NMDS) analysis on the resemblance matrix of Bray–Curtis distances with 500 random restarts (Primer E software). This ordination method has been frequently used in ecological studies (e.g., Minchin 1987; Peat et al. 2007) and has several advantages, such as minimizing the arch effect, releasing linearity constraints, and not requiring multivariate normality of data (Minchin 1987). As with other indirect gradient analyses, NMDS depicts the environment from the organism’s perspective (Clarke and Ainsworth 1993).
Climatic data
Data covering the 3-year period from 2005 to 2007 inclusive were available from an AWS in Taylor Valley at Lake Fryxell, 77°10.60′S 163°10.18′E) on the McMurdo LTER website, www.mcmlter.org (A. Fountain, Lake Fryxell Meteorological Station Measurements, knb-lter-mcm.7,010.7). The required approvals for use of the data were obtained from the LGP and McMurdo LTER coordinators. In the Darwin region, data were obtained from an AWS, located close to Diamond Hill for 3 years 2005–2007, 79°50.113′S, 159°19.12′E, as part of the New Zealand LGP, www.lgp.aq). Soil moisture and temperature were also available from November 16, 2004 to October 2, 2005 (Darwin Glacier region data products, www.lgp.aq). Monthly means of temperature, relative humidity (rH), and water vapor pressure deficit (VPD) were used as these were pre-calculated in the data sources and were compared for the summer (November–January) and winter (June–August) seasons using Student’s t test (STATISTICA 10, StatSoft (Europe) GmbH, Hamburg), with a Bonferroni adjustment of the error rate with p < 0.05 as the significance level. A Levene’s test was used to determine homoscedasticity of the variances, which was accepted (p > 0.05 in all cases, data not shown).
Results
Lichen patterns
A total of 77 lichen species were recorded along the Ross Sea Coast with the highest diversity at Cape Hallett (43 species) and the Queen Maud Mountains (42 species), the lowest number of species was found at Diamond Hill (9 species) (Table 1). Terra Nova Bay and Botany Bay had similar numbers of lichen species (26 and 29, respectively). Rarefaction curves confirmed that the sampling effort was sufficient with a Chao 2 estimator of 98.26 species, allowing further analysis.
The mean similarity index from all inter-site comparisons was 0.255 ± 0.07. However, Diamond Hill showed substantially lower similarity, 0.13–0.19, for all sites with the exception of Taylor Valley, 0.37. Almost all other similarity values were around 30 % (Fig. 2a). The percentage of conformant species was only 33 % at Diamond Hill, but over 50 % for all the other sites. The NMDS showed, with a very low stress level, that one end of the axis was marked by Cape Hallett and Terra Nova Bay, which grouped together, and the other by Diamond Hill (Fig. 2b). With the exception of Queen Maud Mountains, all sites were in the corresponding order along the stress axis as their latitudes. Queen Maud Mountains was placed at approximately the same position as Botany Bay. Life-history type was a major factor in the distribution of taxa with sites located to the left, being characterized by higher levels of conformant species and those to the right by higher levels of tolerant taxa (Fig. 2b).

a Jaccard similarity indices from six independent sites along the Ross Sea coastline with CH Cape Hallett, TN Terra Nova Bay, BB Botany Bay, TV Taylor Valley, DH Diamond Hill, QM Queen Maud Mountains; b NMDS ordination of diversity at the six sites stress values in the right upper corner
Total terrestrial diversity
Total terrestrial (soil and rock, not including water-dominated ecosystems) diversity including lichens, mosses, and invertebrates was highest at Cape Hallett with 59 species and the lowest at Diamond Hill with only 11 species (Table 1). There was no overall cline (adjusted R 2 = 0.0, p = 0.537) as the total biodiversity at Queen Maud Mountains (51 species) was almost as high as at Cape Hallett (59 species) at the northern, end of the Ross Sea coastline. Queen Maud Mountains stand out as an exceptional site and if we remove it from the analysis, then total biodiversity at the remaining five sites showed a clear decline with latitude (adj. R 2 = 0.87, p = 0.013). Almost identical results are obtained if only lichens are considered.
The mean percentage of tolerant species across all sites was 31.6 ± 17.5 % with Diamond Hill standing out having almost doubled this value (55 %). The total number of conformant species ranged from 40 at Cape Hallett to only five at Diamond Hill, and 31 at Queen Maud Mountains. This was also reflected in the number of bryophyte species which was highest at Terra Nova Bay, with 16 species and the lowest at Diamond Hill with only one, Bryum argenteum.
Including cyanobacteria and green algae in a more detailed comparison of Taylor Valley and Diamond Hill (Table 2), the diversity of all taxonomic groups was lower at Diamond Hill. The only exception was the green algae with identical species number (4) at both sites albeit different species. The number of species of cyanobacteria ranged from 18 at Taylor Valley to two at Diamond Hill, with both Diamond Hill species living endolithically (Table 2). Only one mite species (Stereotydeus shoupi) was found at Diamond Hill. No Collembola were found at Diamond Hill, or indeed in the Brown Hills/Darwin Mountain region, despite extensive searches over two field seasons. This compares with one common collembolan species (Gomphiocephalus hodgsoni) found throughout Taylor Valley and two additional species (Neocryptopygus nivicolis, Antarcticinella monoculata) found immediately to the north of the Dry Valleys (≈77.10°S).
Climate
Mean summer temperatures (November–January) for the 3-year period, 2005–2007, were not significantly different between Diamond Hill of Brown Hills and Lake Fryxell of Taylor Valley. While winter temperatures (June–August) were warmer at the Brown Hills −24.6 ± 1.8 versus −30.33 ± 5.5 °C at Taylor Valley, respectively (Table 3). The annual cycle of mean monthly air temperatures over the 3 years (Fig. 3a) showed both sites to have the expected “bathtub” pattern with a long, relatively stable, cold winter period of about 6 months (April–September), this included a brief warm period around December and January, which resulted in a rapid rise and fall in temperature during this time.

Monthly mean temperatures (a) and relative Humidity (rH) (b) over 3 year from 2005 until 2007 (inclusive). Circles correspond with data from the Diamond Hill (Brown Hills), and triangles with Taylor Valley (Central Dry Valleys). Error bars show ±1 SE
Relative humidity of the air was lower at Diamond Hill with an overall 3 year mean of 50.35 % compared with 73.82 % at Lake Fryxell (Fig. 3b). Mean monthly values of rH over the 3 years showed that Diamond Hill always had lower rH and the air was rarely saturated (100 % rH) (Fig. 3b). The water vapor pressure of the atmosphere (VP) was always lower at Diamond Hill with mean VP for the 3 years being 0.096 kPA, which is 25 % of the level at Lake Fryxell. Water VPD, which is an indicator of drying pressure, was higher at Diamond Hill, about 30 % higher in the summer months and four times higher in winter (Table 3).
Snowfalls were regular at Diamond Hill with heavier falls occurring approximately 10 times in the 3 years 2005–2008. However, most falls occurred in the winter months at subzero temperatures and the snow sublimated with no impact on the soil water content. One snow event occurred in mid-January 2005, indicated by the increased light reflectance (Fig. 4b). During this snow event, the air humidity was lower and the soil temperature (Fig. 4b) remained at 0 °C or slightly lower until the snow disappeared. Soil water content increased from about 2 % to about 4.5 % when the snow was present (Fig. 4a). However, soil water content declined again as soon as the snow had disappeared. The increase in soil water content most likely reflected a transfer of water vapor, rather than the formation of liquid water, as soil temperatures were always below 0 °C (Fig. 4a).

Changes in soil temperature (black) and moisture content (gray) (a), and soil reflectance (black) and air relative humidity (gray) (b), during a snow event in January 2005 at Diamond Hill, in the Brown Hills (79°S)
Discussion
For bioclimatic models of biodiversity distribution to be successful, a link between climate and the biota is necessary. Our detailed studies of the terrestrial biodiversity coupled with local climate data at two major regions in the central Ross Sea coast (Taylor Valley vs. Diamond Hill) certainly appear to show such a link. Total terrestrial biodiversity was substantially lower at Diamond Hill, 11 species, compared with 24 at lower Taylor Valley. This agrees with two recent studies showing a very low number of culturable bacteria in the soils of Diamond Hill (Aislabie et al. 2013), and a decline in bryophyte diversity at 80°S (Cannone et al. 2013). The Collembola are entirely absent at Diamond Hill although the area has been ice free, at least one million years (Storey et al. 2010) which has provided enough time for other ice-free areas (e.g., Ross Island) to be colonized (Stevens and Hogg 2002, 2003). The cyanobacteria, a group well known for its resilience, is also much reduced at Diamond Hill (2 species vs. 18 species at Taylor Valley). Another indicator of the more extreme nature of Diamond Hill is the high proportion of tolerant species (6 out of a total of 11), a significantly higher level than at other sites.
Temperature is often suggested to be the major controlling environmental factor in Antarctica. However, this does not appear to be the case in our analyses. Although the climatic data were limited to 3 years at both sites, it appears that temperatures particularly during the summer months were similar between the two sites. Winter temperatures (June–August) were actually warmer at the Brown Hills −24.6 versus −30.3 °C in Taylor Valley (Table 3). It seems that temperature alone is an unlikely explanation for the large differences in richness of terrestrial biota between the two sites. Instead, we propose that the lower biodiversity at Diamond Hill is a result of the low water availability, and in particular, a lack of liquid water. The presence of salts such as gypsum on rock surfaces in the vicinity of Diamond Hill indicates extreme dryness and absence of snow melt. Although there were at least ten snowfall events detected by increased ground reflectance in the period of February 2005–January 2007, the majority of these occurred between early February and the end of November when temperatures were at, or below −5 °C. Under these conditions, there is no liquid water. Only during one event, at the end of December 2005, were temperatures possibly warm enough (around −2 °C) to allow for snow melt. The lack of water will be exacerbated by the higher VPD in summer at Diamond Hill which will encourage snow loss by sublimation. rH of the air was always much lower at Diamond Hill throughout the year. Modeling of precipitation across Antarctica shows that there is an area of high sublimation and low net precipitation between Ross Island and Queen Maud Mountains, including the latitude of Diamond Hill (Bromwich and Guo 2004). The near absence of terrestrial cyanobacteria, declining from 18 species at Taylor Valley to two species at Diamond Hill, strongly supports a lack of liquid water. All of the photosynthetic species we found were poikilohydric and must be hydrated to become fully active. However, cyanobacteria are exceptional in that they have an absolute requirement for liquid water to become photosynthetically active, while other groups such as the algae and chlorolichens are able to reactivate from humid air (Lange et al. 1986).
All studies, including those in Antarctica, linking climate and biodiversity are founded on the premise that the terrestrial biodiversity at any site is in equilibrium with the present climate. Any changes in climate will, therefore, lead to changes in biodiversity. Although bacterial and fungal communities certainly represent most of the diversity in the Dry Valley region (Cary et al. 2010), our focus was on primary producers, as the basis for trophic hierarchy. The comparison of Taylor Valley and Diamond Hill supports a role for climate in determining terrestrial biodiversity. However, despite the link between biodiversity and climate (via water availability), at these two sites, there are several reasons why application of bioclimatic modeling in the Ross Sea region could be problematic:
Data suggest a complex latitudinal gradient for lichen diversity
Lichens in continental Antarctica, especially in the Dry Valleys, are known to have extremely low growth rates, as low as 0.0036 mm/year, and some thalli may have existed for 1,000s of years (Green et al. 2011a, b, 2012). Their distribution and abundance, therefore, indicate climate conditions over much longer time scales, than the relatively short climate measurements, provided here. Furthermore, the number of lichen species appears to be spatially variable and site-dependent—both the Taylor Valley and Diamond Hill sites have lower total lichen species compared with other sites in the Ross Sea. The exceptional site is the Queen Maud Mountains region, around 84°S, where the lichen flora is actually one of the most diverse in the Ross Sea with around 42 species (Green et al. 2011b). This compares with sites much further north, including 26 species at Terra Nova Bay (Castello 2003), 29 species at Botany Bay (Seppelt et al. 2010) and 43 at Cape Hallett (Green, unpubl. data). The low number of lichen species at Diamond Hill suggests a more extreme habitat, and this is supported by the high percentage of tolerant species in Diamond Hill. We suggest that conformant species reach their ecological limits at Diamond Hill.
Micro- versus macro-habitats
The irregular distribution of terrestrial vegetation in the Ross Sea region is likely a reflection of low precipitation, so that vegetation is confined to microsites and is more a result of landscape and melt-water availability than of local macroclimate (Kennedy 1993; Green et al. 2011a). We also suggest from our detailed comparison between Diamond Hill and lower Taylor Valley that water availability was the dominant environmental factor as far as poikilohydric organisms are concerned (Kennedy 1993; Broady 1996, 2005). However, there is also a link with mean annual temperature, which acts by influencing the quantity of liquid water and the landscape dictating where the water will flow. Consequently, the location and abundance of terrestrial vegetation are dependent on local conditions rather than on macro-environmental factors such as temperature or light. The net result is that there is no clear gradient in the number of lichen species, or for terrestrial biodiversity in general, along the Ross Sea coast.
Biodiversity may reflect history rather than climate
One reason for the high diversity in the Queen Maud Mountains may be the presence of a relict group of species, such as lichens and invertebrates that have survived at least since the last major ice-free period (Stevens et al. 2006). This has been linked with collapses of the Western Antarctic Ice Sheet and corresponding open seaways resulting in the present biodiversity being, in part, a consequence of history rather than climate (Green et al. 2011b).
Dispersal limitations
Dispersal among ice-free areas may also have a stronger influence on species occurrence than local macroclimate. For example, the distributions of mites and Collembola in the Ross Sea region seem to be restricted by ice barriers (e.g., Demetras et al. 2010). For bryophytes along the western Ross Sea coastline, 80°S stands out as an area of low diversity (Cannone et al. 2013). The Brown Hills vicinity, close to the center of the Ross Sea latitudinal gradient, appears to be the most extreme coastal site and is likely a barrier to dispersal. Conditions inland is even more extreme. The fact that species of lichen to the south in the Queen Maud Mountains (Green et al. 2011b) have not moved northward supports Brown Hills as a dispersal barrier. This is further supported by the unique arthropod (e.g., Demetras et al. 2010) and lichen (Sancho, unpubl. data) species in the Queen Maud Mountains that have evolved in the absence of gene flow from the north.
Conclusions
The above arguments suggest that terrestrial biota along the Ross Sea coastline is not a result of the macroclimate and associated parameters used in climatic models. Instead, biodiversity at any site is largely a product of local microclimate conditions. Accordingly, forecasting the effects of climate change on vegetation distribution becomes more challenging for the Ross Sea region of Antarctica. A previous study comparing climate data with vegetation distribution in South Australia (Traill et al. 2013) suggested that arid species’ diversity was largely driven by environmental parameters at the localized scale. Thus, it appears that when vegetation becomes confined to microhabitats, such as in arid zones, then macroclimate is much less successful in forecasting vegetation distribution.
It is probable that local changes in distribution will occur as a result of changes in location and duration of snow melt, and abundance of snow. However, predicted changes in precipitation are less certain than those for temperature (Pachauri and Reisinger 2008). Faster growth rates for lichens from the wet maritime Antarctic (e.g., Sancho and Pintado 2004) relative to those of the Ross Sea coastline (e.g., Schroeter et al. 2011) may foreshadow potential responses of the continental taxa as well as provide a sensitive indicator of climate changes (Sancho et al. 2007). It is clear that further, intensive surveying of the terrestrial biota including bacterial and fungal diversity is required along the Ross Sea coastline, particularly to the south of Ross Island. Molecular approaches such as DNA barcoding (sensu Hebert et al. 2003) will also be beneficial. While, these data will be critical for biogeographical studies and conservation efforts, application to bioclimatic models may be more challenging.
References
Adams BJ, Bardgett RD, Ayres E et al (2006) Diversity and distribution of Victoria Land biota. Soil Biol Biochem 38:3003–3018
Aislabie JM, Lau A, Dsouza M, Sheperd C, Rhodes P, Turner SJ (2013) Bacterial composition of soils of Lake Wellman area, Darwin Mountains, Antarctica. Extremophiles 17:775–786
Barrett JE, Virginia RA, Hopkins DW et al (2006) Terrestrial ecosystem processes of Victoria Land, Antarctica. Soil Biol Biochem 38:3019–3034
Broady PA (1996) Diversity, distribution and dispersal of Antarctic terrestrial algae. Biodivers Conserv 5:1307–1335
Broady PA (2005) The distribution of terrestrial and hydro-terrestrial algal associations at three contrasting locations in southern Victoria Land, Antarctica. Algol Stud 38:95–112
Bromwich DH, Guo Z (2004) Modelled Antarctic precipitation. Part I: spatial and temporal variability. J Clim 17:427–447
Campbell IB, Claridge GGC (1982) The influence of moisture on the development of soils of the cold deserts of Antarctica. Geoderma 28:221–238
Cannone N, Convey P, Guglielmin M (2013) Diversity trends of bryophytes in continental Antarctica. Polar Biol 36:259–271
Cary SC, McDonald R, Barrett JE, Cowan DA (2010) On the rocks: the microbiology of Antarctic Dry Valley soils. Nat Rev Microbiol 8:129–138
Castello C (2003) Lichens of the Terra Nova Bay area, Northern Victoria Land (continental Antarctica). Stud Geobot 22:3–54
Clarke KR, Ainsworth M (1993) A method of linking multivariate community structure to environmental variables. Mar Ecol Prog Ser 92:205–219
Colesie C, Gommeaux M, Green TGA, Büdel B (2013) Biological soil crusts in continental Antarctica: Garwood Valley, southern Victoria Land, and Diamond Hill, Darwin Mountains region. Antarct Sci 26:115–123
Colwell RK (2006) EstimateS: statistical estimation of species richness and shared species from samples. Version 8. Persistent. purl.oclc.org/estimates. Accessed 25 July 2013
De Frenne P, Graae BJ, Rodriguez-Sanchez F, Kolb A, Chaberrie O, Decocq G, De Kort H, De Schrijver A, Diekmann M, Eriksson O, Gruwez R, Hermy M, Lenoir J, Plue J, Coomes DA, Verheyen K (2013) Latitudinal gradients as natural laboratories to infer species’ responses to temperature. J Ecol 101:784–795
Demetras NJ, Hogg ID, Banks JC, Adams BJ (2010) Latitudinal distribution and mitochondrial DNA (COI) variability of Stereotydeus spp. (Acari: prostigmata) in Victoria Land and the central Transantarctic Mountains. Antarct Sci 22:749–756
Ettl H, Gärtner G (1995) Syllabus der Boden-, Luft- und Flechtenalgen. Gustav Fischer, Stuttgart
Fountain AG, Lyons WB, Burkins MB, Dana GL, Doran PT, Lewis KJ, McKnight DM, Moorhead DL, Parsons AN, Priscu JC, Wall DH, Wharton RA, Virginia RA (1999) Physical controls on the Taylor Valley ecosystem, Antarctica. Bioscience 49:961–971
Freckman DW, Virginia RA (1997) Low diversity Antarctic soil nematode communities: distribution and response to disturbance. Ecology 78:363–369
Green TGA, Sancho LG, Pintado A, Schroeter B (2011a) Functional and spatial pressures on terrestrial vegetation in Antarctica forced by global warming. Polar Biol 34:1643–1656
Green TGA, Sancho LG, Türk R, Seppelt RD, Hogg ID (2011b) High diversity of lichens at 84°S, Queen Maud Mountains, suggests preglacial survival of species in the Ross Sea region, Antarctica. Polar Biol 34:1211–1220
Green TGA, Brabyn L, Beard C, Sancho LG (2012) Extremely low lichen growth rates in Taylor Valley, Dry Valleys, continental Antarctica. Polar Biol 35:535–541
Hebert PDN, Cywinska A, Ball SL, deWaard JR (2003) Biological identifications through DNA barcodes. Proc R Soc Lond B 270:313–322
Heinkinnen RK, Luoto M, Araujo MB, Virkkala R, Thuiller W, Sykes MT (2006) Methods and uncertainties in bioclimatic envelope modelling under climate change. Prog Phys Geogr 30:751–777
Hogg ID, Cary CS, Convey P, Newsham KK, O’Donnell AG, Adams BJ, Aislabie J, Frati F, Stevens MI, Wall DH (2006) Biotic interactions in Antarctic terrestrial ecosystems: are they a factor? Soil Biol Biochem 38:3035–3040
Howard-Williams C, Peterson D, Lyons WB, Cattaneo-Vietti R, Gordon S (2006) Measuring ecosystem response in a rapidliy changing environment: the Latitudinal Gradient Project. Antarct Sci 18:465–471
Howard-Williams C, Hawes I, Gordon S (2010) The environmental basis of ecosystem variability in Antarctica: research in the Latitudinal Gradient Project. Antarct Sci 22:591–602
Kennedy AD (1993) Water as a limiting factor in the Antarctic terrestrial environment: a biogeographical synthesis. Arc Antarct Alp Res 25:308–315
Komárek J, Anagnostitis K (1999) Cyanoprokaryota-1. Teil/part 1: Chroococcales. In: Ettl H, Gerloff J, Heynig H, Mollenbauer D (eds) Süßwasserflora von Mitteleuropa 19/1. Elsevier, Heidelberg
Komárek J, Anagnostitis K (2005) Cyanoprokaryota—2. Teil/2nd part: Oscillatoriales. In: Büdel B, Krienitz L, Gärtner G, Schagerl M (eds) Süsswasserflora von Mitteleuropa 19/2. Elsevier, Heidelberg
Lange OL, Kilian E, Ziegler H (1986) Water vapor uptake and photosynthesis of lichens: performance differences in species with green and blue-green algae as phycobionts. Oecologia 71:104–110
Magalhães C, Stevens MI, Cary SC, Ball BA, Storey BC, Wall DH, Türk R, Ruprecht U (2012) At limits of life: multidisciplinary insights reveal environmental constraints in biotic diversity in continental Antarctica. PLoS One 7:1–10
Minchin PR (1987) An evaluation of the relative robustness of techniques for ecological ordination. Vegetatio 69:89–107
Ochyra R, Lewis Smith RI, Bednarek-Ochyra H (2008) The illustrated moss flora of Antarctica. Cambridge University Press, Cambridge
Øvstedal DO, Smith RIL (2001) Lichens of Antarctica and South Georgia. A guide to their identification and ecology. Cambridge University Press, Cambridge
Pachauri RK, Reisinger A (2008) Climate change 2007. Synthesis report. Contribution of working groups 1, 2 and 3 to the fourth assessment report. IPCC, Geneva
Pearson RG, Dawson TP (2003) Predicting the impacts of climate change on the distribution of species: are bioclimate envelope models useful? Glob Ecol Biogr 12:361–371
Peat HJ, Clarke A, Convey P (2007) Diversity and biogeography of the Antarctic flora. J Biogeogr 34:132–146
Powers LE, Ho MC, Freckman DW, Virginia RA (1998) Distribution, community structure, and microhabitats of soil invertebrates along an elevational gradient in Taylor Valley, Antarctica. Arct Alp Res 30:133–141
Sancho LG, Pintado A (2004) Evidence of high annual growth rate for lichens in the maritime Antarctic. Polar Biol 27:312–319
Sancho LG, Green TGA, Pintado A (2007) Slowest to fastest: extreme range in lichen growth rates supports their use as an indicator of climate change in Antarctica. Flora 202:667–673
Schroeter B, Green TGA, Pannewitz S, Schlensog M, Sancho LG (2011) Summer variability, winter dormancy: lichen activity over 3 years at Botany Bay, 77°S latitude, continental Antarctica. Polar Biol 34:13–22
Seppelt RD, Türk R, Green TGA, Moser G, Pannewitz S, Sancho LG, Schroeter B (2010) Lichen and moss communities of Botany Bay, Granite Harbour, Ross Sea, Antarctica. Antarct Sci 22:691–702
Stevens MI, Hogg ID (2002) Expanded distributional records of Collembola and Acari in southern Victoria Land, Antarctica. Pedobiologia 46:485–496
Stevens MI, Hogg ID (2003) Long-term isolation and recent range expansion revealed for the endemic springtail Gomphiocephalus hodgsoni from southern Victoria Land, Antarctica. Mol Ecol 12:2357–2369
Stevens MI, Greenslade P, Hogg ID, Sunnucks P (2006) Southern hemisphere springtails: could any have survived glaciation of Antarctica? Mol Biol Evol 23:874–882
Storey BC, Fink D, Hood D, Joy K, Shulmeister J, Riger-Kusk M, Stevens MI (2010) Cosmogenic nuclide exposure age constraints on the glacial history of the Lake Wellman area, Darwin Mountains. Antarct Sci 22:603–618
Strandtmann RW (1967) Terrestrial Prostigmata (trombidiform mites). In: Gressitt JL (ed) Entomology of Antarctica. American Geophysical Union, Antarctic Research Series 10, pp 51–80
Tamppari LK, Anderson RM, Archer PD et al (2012) Effects of extreme cold and aridity on soils and habitability: McMurdo dry valleys as an analogue for the Mars Phoenix landing site. Antarct Sci 24:211–228
Traill LW, Wanger TC, de Little SC, Brook BW (2013) Rainfall and temperature variation does not explain arid species diversity in outback Australia. Res Rep Biodivers Stud 3:1–8
Wise KAJ (1964) New records of Collembola and Acarina in Antarctica. Pac Insects 6:522–523
Wise KAJ (1967) Collembola (springtails). In: Gressitt JL (ed) Entomology of Antarctica. American Geophysical Union, Antarctic Research Series 10, pp 123–148
Wise KAJ (1971) The Collembola of Antarctica. Pac Insect Monogr 25:57–74
Wynn-Williams DD, Edwards HGM (2000) Antarctic ecosystems as models for extraterrestrial surface habitats. Planet Space Sci 48:1065–1075
Acknowledgments
We are grateful to Antarctica New Zealand (AntNZ) for logistical support over several years as part of the LGP coordinated by Shulamit Gordon. Logistics support was also provided by the Australian Antarctic Programme, the Spanish National Antarctic Program and the US Coastguard Reserve. The University of Waikato Vice Chancellor’s Fund and the Department of Biological Sciences, University of Waikato provided financial assistance with field costs. The research was supported by the New Zealand MBIE grant, “Understanding, valuing and protecting Antarctica’s unique terrestrial ecosystems: predicting biocomplexity in Dry Valley ecosystems,” and TGAG by the Spanish Education Ministry Grants Nos. POL2006-08405 and CTM2009-12838-C04-01. BB and CC acknowledge the DFG Schwerpunktprogramm 1158 (BU 666/11-1). Thanks to O. Breuss for the identification of Verrucaria species, to U. Ruprecht, and to H. Reichenberger for field support. We thank Dr. A. Fountain for making available the microclimate data from Lake Fryxell (Lake Fryxell Meteorological Station Measurements, knb-lter-mcm. 7010.7). Special thanks to Dr. R. Wirth for support with the statistical analyses and to two anonymous reviewers for their helpful and constructive comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Colesie, C., Green, T.G.A., Türk, R. et al. Terrestrial biodiversity along the Ross Sea coastline, Antarctica: lack of a latitudinal gradient and potential limits of bioclimatic modeling. Polar Biol 37, 1197–1208 (2014). https://doi.org/10.1007/s00300-014-1513-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-014-1513-y