
Abstract
The main aim of our long-term study on the at-sea distribution of the upper trophic levels in polar marine ecosystems is to deepen the understanding of the basic mechanisms affecting their distribution, i.e. hydrological factors such as water masses and fronts, pack ice and ice edge, eddies. A second goal consists in detecting spatial and temporal changes, with special attention to global climate changes, as well as possible consequences of (krill) fisheries. Seabirds and marine mammals were recorded during an autumn expedition of icebreaking RV Polarstern between Punta Arenas and the South Shetland Islands, Antarctica in March/ April 2012. During a total of 333 half-an-hour transect counts without width limitation, 113,500 seabirds were encountered, belonging to 40 species. The vast majority were southern fulmars and grey-headed albatross around the South Shetland Islands, with 76,800 and 7,000 individuals, respectively. Fin whale was by far the most abundant cetacean with 300 identified individuals in the same area, of which 100 in one count. These exceptional concentrations of fin whale seem to reflect an autumn pre-migration feeding aggregation. For most species, the majority was concentrated in very few counts, reflecting a very high patchiness and, as upper trophic levels, an important prey availability—mainly krill, nekton and small fish. Low biodiversity was reflected by both the low number of species and the fact that a few species represent the vast majority in numbers. It is suggested that the area deserves future biological studies, especially in autumn, and a special protection management.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In the frame of our long-term study of the quantitative distribution at sea of seabirds and marine mammals in polar marine ecosystems, the main aims are, on the one hand, to deepen the understanding of the mechanisms influencing their distribution (water masses, fronts, pack ice, ice edge and eddies) and on the other hand to try and detect spatial and temporal evolutions with special attention to possible consequences of global climatic changes, and of (krill) fisheries. As upper trophic levels, their distribution reflects prey abundance: zooplankton, krill, nekton, and small fish and so integrates the ecology and the biological production of the whole water column. They constitute thus the best approach to identify and localise areas of high biological production, and to detect temporal changes (Hunt 1990; Bost and Le Maho 1993; Furness and Camphuysen 1997; Joiris 2007; Joiris and Falck 2010).
This paper reports on data collected about the upper trophic levels in the south-western Atlantic sector, from South America to the South Shetland Islands, i.e. a poorly studied area in autumn.
Materials and methods
During the ANT XXVIII/4 expedition of icebreaking RV Polarstern from Punta Arenas to the South Shetland Islands and back, from March 14 to April 8, 2012, 333 transect counts were devoted to seabird and marine mammal observation. The transect counts were lasting half an hour, without width limitation from the bridge at 18 m above sea level on a continuous basis, light and visibility allowing (see description and discussion in Joiris 2007; Joiris and Falck 2010). The animals were detected with the naked eye, and observations confirmed and complemented with binoculars (10 × 42) when useful; photographic documents were also used, especially for species difficult to identify. Results are presented as basic unmodified data, i.e. numbers encountered per half-an-hour transect count. Density was also calculated on the basis of specific detection limits established by this team in function of size, colour and behaviour (jizz) (Joiris 2007; Joiris and Falck 2010) and mean ship’s speed: 10 knots in open water and 5 knots in ice covered areas.
Water temperature and salinity were continuously recorded on board Polarstern with a thermo-salinometer at subsurface sampling (keel −10 m). Ice cover was evaluated by us from the bridge and expressed as % coverage within an approximate range of 500 m around the ship. A navigation echo-sounder was continuously running, mainly in order to follow water depth, but also showing echo’s in the water column, such as zooplankton, nekton and fish (Simrad Kongsberg EK60 with multifrequency at 70 or 120 kHz and an angle of 7°).
Results
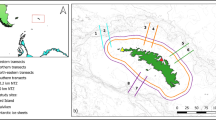
The route followed by Polarstern is shown in Fig. 1, as well as the position and numbering of the transect counts, the positions of the slope and ice edge (Online Resource 1). During the 333 counts devoted to the upper trophic levels, 113,000 birds were encountered belonging to 40 species, without taking into account the strictly coastal ones. The most numerous species was southern fulmar Fulmarus glacialoides with 77,000 individuals, i.e. 68 % of the total; next came grey-headed albatross Thalassarche chrysostoma with 7,000 individuals; and Antarctic prion Pachyptila desolata with 6,400 individuals (Table 1). Similarly, fin whale Balaenoptera physalus formed the vast majority of the identified large cetaceans: 308 individuals out of 330, i.e. 94 % (Table 1). Concentrations of these species were highest within the South Shetland Islands area, especially off Elephant Island (Fig. 2). Mean numbers for this area are out of 99,700 seabirds in 205 counts: 76,800 southern fulmars (77 %)—of which 57,600 in one count west of Elephant Island, 6,940 grey-headed albatrosses (Table 1) and 308 fin whales, corresponding to 105 sightings. These numbers of fin whales are probably an underestimate, considering that the majority of unidentified large whales (n = 87, Table 1) were actually fin. This is why data from this area were analysed and discussed in some detail (Table 2). Density was calculated in this limited area for fin whale and the most abundant seabird species (N ≥ 100): mean density was 0.03/km2 for fin whale; a mean density of more than 9/km2 was found for southern fulmar, of 6.2/km2 for grey-headed albatross, and 17/km2 for Antarctic prion, one of the most abundant species in the southern seas.

Route of RV Polarstern during expedition ANT XXVIII/4, March/April 2012; position and numbering of the transect counts for seabirds and marine mammals; position of the shelf slope indicated by a dotted line (top); pack ice northern limit in Bransfield Strait indicated by a dotted line (bottom): situation on April 4, 2012; based on a “Polar View” map (University of Bremen, Germany: http://iup.physik.uni-bremen.de) (online Resource 1)


Counting results for the most abundant species, totals per transect count in classes of abundance, birds: southern fulmar F. glacialoides (a), grey-headed albatross T. chrysostoma (b), Antarctic prion P. desolata (c); cetaceans: fin whale B. physalus (circles) and unidentified large whale (triangles) (d)
Moreover, these very high values were concentrated in few counts in a limited area north-west of Elephant Island, reflecting a huge patchiness in distribution: 100 fin whales in count 127 (March 21, 61°S, 55.50°W), 67,570 fulmars in three neighbouring counts, i.e. 1 % of the total (142, 143, 145, of which 57,600 in count 143, March 23, 61°S, 56°W), 457 Antarctic prions in count 143 and 1,480 grey-headed albatrosses in count 144. A colloquial summary becomes “it all happened there”. Translated into density (N/km2), these maximal numbers encountered correspond to 2.2 fin whales, 1,400 southern fulmars and 270 grey-headed albatrosses.
High prey availability was reflected by visual observation of large subsurface swarms of krill Euphausia superba, extending down to 50–100 m deep above the slope of the “continental” shelf of Elephant Island as seen on the echo-sounder screen (Online Resource 2).
Some observations deserve comments: 3 bird species were recorded more than 1,000 km south or southwest of their known range, according to Shirihai (2007); 2 sooty albatrosses Phoebetria fusca on March 16 at 58°S, 60°W, a few soft-plumaged petrels Pterodroma mollis in the Drake Passage, on the way to Elephant Island on March 17. A common diving-petrel Pelecanoides urinatrix was found alive as a shipwrecked bird on March 25 at about 50 km west of Elephant Island and released. Three hourglass dolphins Lagenorhynchus cruciger correspond to the group present far from the coast in open water (Santora 2011; Dellabianca et al. 2012). Temporal evolution was noted during the expedition. Southern fulmar, for instance, was close to absent from the south route and present in significant numbers during the return north transect (Fig. 2a).
This study area is covering the south-western Atlantic at the boundary of the Antarctic Circumpolar Current ACC and the Polar Front PF, i.e. at the limit between low and high Antarctic, as reflected by water temperature and salinity data (Fig. 3) and corresponding to the shelf slope. In the north and east of the South Shetland Islands, the bathymetry (slope of the continental shelf) might cause food-rich upwelling. On the other hand, the vicinity of a deep canyon strongly influences krill and top predator abundance (Santora and Reiss 2011). The biological production is high, as reflected for instance by high oxygen concentration (Orsi et al. 1995).

Hydrological data collected on board Polarstern: salinity (a) and water temperature (°C) (b)
Discussion
Water masses and fronts, pack ice and ice edge are the main hydrological factors influencing the distribution of the upper trophic levels—seabirds and marine mammals—in the ocean. This is known for decades (Wynne-Edwards 1935; Joiris 1979; Pocklington 1979; Kinder et al. 1983) and was confirmed more recently by many studies (Elphick and Hunt 1993; Joiris and Falck 2010). Boundaries such as ice edge and slope are known to play a very important role, especially in Antarctic ecosystems (Ainley et al. 1998, 2007, 2012; Ribic et al. 1991, 2011; Jacobs 1991; Joiris 1991; van Franeker 1992; Bost et al. 2009; Branch 2011). Seabird and baleen whale distribution is depending on prey abundance, krill, nekton and fish (Heinemann et al. 1989; Ryan and Cooper 1989; Reid et al. 2000), mainly krill in this case. Few publications, however, simultaneously include birds and cetaceans (this team; Friedlaender 2006; Friedlaender et al. 2008; Santora et al. 2009a, 2010; Ballard et al. 2011). The South Shetland Islands, especially Elephant Island, are known for their high krill concentrations attracting seabird predators (Hunt et al. 1990; Hewitt and Demer 1993; Agnew 1997). They also support an important krill fishery, the main one together with South Georgia (Croxall and Nicol 2004) even if it strongly decreased during the last decades (Everson and Goss 1991; Jones and Ramm 2004). Important variations in krill density were recorded, directly influencing seabird foraging (Santora et al. 2009b). An exceptionally large “super krill swarm” was detected in 1981 (Tarling et al. 2009), then not any longer for decades (Lawson et al. 2008) and again detected in 2009 in the western Antarctic Peninsula in association with very high concentration of humpback whales (up to 5/km2) (Nowacek et al. 2011). The data presented here, even if semi-quantitative for krill (Online Resource 1), seem to reflect the presence of such a huge swarm again.
The high numbers of fin whales, southern fulmars and grey-headed albatrosses represent an exceptionally large aggregation and were concentrated in a very small subarea at the front, even if not strictly in the same counts, reflecting a huge patchiness. Most individuals of the three species were actively subsurface feeding and resting, both bird species mainly sitting on the water in “rafts”, an unusual observation for albatrosses. The quantitative importance of the area for the main species is to be considered in relation to their global population as well. Southern fulmar has one million breeding pairs, mainly on the islands of the Scotia Sea Arc (Creuwels et al. 2007). Our observed total of 7,000 grey-headed albatrosses, mainly adults, represents more than 1 % of the global population of about 600,000 individuals or 3 % of the estimated 250,000 adults, of which 90,000–95,000 breeding pairs (Shirihai 2007). Obviously, their absolute number in the area must be a lot higher than the observed ones. Many individuals are known to regularly visit this sector of the southern seas (ACAP 2009). This makes the South Shetland Islands area a major feeding ground for this species classified as “vulnerable and decreasing” (ACAP 2009; IUCN 2012). For fin whale, an endangered species as well (IUCN 2012), 100,000–120,000 individuals are present in the southern hemisphere, south of 50°S in summer (Gambell 1985; Shirihai 2007). They are usually encountered in groups of 3–10 or 20 with some concentrations of 100 (Gambell 1985). During previous studies in the area, they were noted in numbers varying between 9 and 30 sightings (Pankow and Kock 2000; Branch and Butterworth 2011; Sirovic et al. 2006; Scheidat et al. 2007). The large number we encountered was not mentioned before. Other species were noted in high numbers, but their relative abundance was low: e.g. Antarctic prion with 6,400 individuals out of a population estimated as up to 20 million breeding pairs (Shirihai 2007), probably the most numerous marine bird species in the southern hemisphere. Other species were concentrated in the same area on the slope of the “continental” shelf of Elephant Island: e.g. chinstrap penguin Pygoscolis antarctica and black-browed albatross Thalassarche [melanophrys] melanophrys, confirming the special importance of the shelf slope (Fig. 4).

Position of the peak abundance for selected seabird and cetacean species
Seasonal factors might be of importance as well, namely the probable autumn migration and accumulation of adult krill migrating before overwintering under pack ice (Lascara et al. 1999; Hewitt et al. 2004; Nicol 2006) on the one hand. On the other hand the seasonal migration of large cetaceans towards the tropics in autumn, fin whales were possibly aggregating in a pre-migration feeding situation. Concerning seabirds, the species composition was very different during the breeding season (Santora et al. 2009a, b); this difference probably reflects an important post-breeding movement for most bird species leaving their breeding grounds.
Conclusions
Low biodiversity ecosystems such as the polar marine ones are characterised by low numbers of species, of which a few only form the majority of individuals. Moreover, their distribution is very patchy with huge aggregations in very limited zones, usually at the polar front in mixed Polar/Antarctic Water or Polar/Arctic Water, respectively. These areas thus show high biological production in the water column. If such concentrations were not covered by geographical studies, the numbers and densities are dramatically underestimated. Considering that upper trophic levels distribution basically reflects prey abundance, one might guess that, similarly, studies not covering these areas also dramatically underestimate prey numbers and density.
Present observations stress the international importance of the South Shetland Islands for several species, especially in autumn. This area therefore deserves a strong protection management, including interdiction of “scientific” whaling and krill fishery. More research outside summer or winter is needed in order to confirm and deepen the knowledge of such important autumn feeding aggregations.
References
ACAP (2009) Agreement on the Conservation of Albatrosses and Petrels. ACAP Species Assessments: www.acap.aq/acap-species
Agnew DJ (1997) The CCAMLR ecosystem monitoring programme. Antarct Sci 9:235–242
Ainley DG, Jacobs SS, Ribic CA, Gaffney I (1998) Seabird distribution and oceanic features of the Amundsen and southern Bellingshausen seas. Antarct Sci 10:111–123
Ainley DG, Dugger KM, Tonolio V, Gaffney I (2007) Cetacean occurrence patterns in the Amundsen and southern Bellingshausen Sea sector, southern ocean. Mar Mamm Sci 23:287–305
Ainley DG, Jongsomijt D, Ballard G, Thiele D, Fraser WR, Tynan T (2012) Modelling the relationship of Antarctic minke whale to major ocean boundaries. Polar Biol 35:281–290. doi:10.1007/s00300-011-1075-1
Ballard G, Jongsomjit D, Veloz SD, Ainley DG (2011) Coexistence of mesopredators in an intact polar ocean ecosystem: the basis for defining a Ross Sea marine protected area. Biol Conserv. doi:10.1016/j.biocon.2011.11.017 (in press)
Bost CA, Le Maho Y (1993) Seabirds as bio-indicators of changing marine ecosystems: new perspectives. Acta Oecol 14:463–470
Bost CA, Cotté C, Bailleul F, Cherel Y, Charassin JB, Guinet C, Ainley DG, Weimerskirch H (2009) The importance of oceanographic fronts to marine birds and mammals in the southern oceans. J Mar Syst 78:363–376
Branch TA (2011) Humpback abundance south of 60°S from three complete circumpolar sets of surveys. J Cetacean Res Manag 3:53–69
Branch TA, Butterworth DS (2011) Estimates of abundance south of 60° S for cetacean species sighted frequently on the 1978/79 to 1997/98 IWC/IDCR-SOWER sighting surveys. J Cetacean Res Manag 3:251–270
Creuwels JCS, Poncet S, Hodum PJ, van Franeker JA (2007) Distribution and abundance of the southern fulmar Fulmarus glacialoides. Polar Biol 30:1083–1097. doi:10.1007/s00300-007-0276-0
Croxall JP, Nicol S (2004) Management of Southern Ocean fisheries: global forces and future sustainability. Antarct Sci 16:569–584
Dellabianca N, Sciosa G, Schiavini A, Raya Rey AR (2012) Occurrence of hourglass dolphin (Lagenorhynchus cruciger) and habitat characteristics along the Patagonian Shelf and the Atlantic sector of the Southern Ocean. Polar Biol 35:1921–1927. doi:10.1007/s00300-012-1217-0
Elphick C, Hunt GL Jr (1993) Variations in the distributions of marine birds with water mass in the Northern Bering Sea. Condor 95:33–44
Everson I, Goss C (1991) Krill fishing activity in the southwest Atlantic. Antarct Sci 3:351–358
Friedlaender AS (2006) Distribution of whales in relation to prey and oceanographic processes in the inner shelf waters of the Western Antarctic Peninsula. Mar Ecol Progr Ser 317:297–310
Friedlaender AS, Fraser WR, Patterson D, Qian SS, Halpin PN (2008) The effects of prey demography on humpback whale abundance around Anvers Island, Antarctica. Polar Biol 31:1217–1224
Furness RW, Camphuysen CJK (1997) Seabirds as monitors of the marine environment. ICES J Mar Sc 54:726–737
Gambell R (1985) Fin whale Balaenoptera physalus (Linnaeus, 1758). In: Ridgway SH, Harrison R (eds) Handbook of marine mammals, vol 3: the sirenians and baleen whales. Academic Press, London, pp 171–192
Hammer WM, Hammer PP, Strand SW, Gilmer RW (1983) Behavior of Antarctic krill Euphausia superba: chemoreception, feeding, schooling, and molting. Science 220:433–435
Heinemann D, Hunt G, Everson I (1989) Relationship between the distribution of marine avian predators and their prey, Euphausia superba in Bransfield Strait and southern Drake Passage, Antarctica. Mar Ecol Prog Ser 58:3–16
Hewitt RP, Demer DA (1993) Dispersion and abundance of Antarctic krill in the vicinity of Elephant Island in the 1992 austral summer. Mar Ecol Prog Ser 58:29–39
Hewitt RP, Kimm S, Naganobou M et al (2004) Variation in the biomass and demography of Antarctic krill in the vicinity of the South Shetland Islands during the 1999/2000 austral summer. Deep-Sea Res 51:1411–1419
Hunt GL Jr (1990) The pelagic distribution of marine birds in a heterogeneous environment. Polar Res 8:43–54
Hunt GL Jr, Heinemann D, Veit RR, Heywood RB, Everson I (1990) The distribution, abundance and community structure of marine birds in southern Drake Passage and Bransfield Strait. Cont Shelf Res 10:243–257
IUCN (2012) Red list of threatened species, version 2012.1. www.iucnredlist.org
Jacobs S (1991) On the nature and significance of the Antarctic Slope Front. Mar Chem 35:9–24
Joiris CR (1979) Seabirds recorded in the northern North Sea in July: the ecological implications of their distribution. Gerfaut 68:419–440
Joiris CR (1991) Spring distribution and ecological role of seabirds and marine mammals in the Weddell Sea, Antarctica. Polar Biol 11:415–424. doi:10.1007/BF80233076
Joiris CR (2007) At-sea distribution of seabirds and marine mammals in the Greenland and Norwegian seas: impact of extremely low ice coverage. In: Symposium European research on polar environment and climate, Brussels, 5–6 March 2007 http://ec.europa.eu/research/environment/newsanddoc/agenda0307_en.htm
Joiris CR (2011) A major feeding ground for cetaceans and seabirds in the south-western Greenland Sea. Polar Biol 34:1597–1607. doi:10.1007/s00300-011-1022-1
Joiris CR, Falck E (2010) Summer at-sea distribution of little auks Alle alle and harp seals Pagophilus (Phoca) groenlandica in the Fram Strait and the Greenland Sea: impact of small-scale hydrological events. Polar Biol 34:541–548. doi:10.1007/s00300-010-0910-0
Jones CD, Ramm DC (2004) The commercial harvest of krill in the southwest Atlantic before and during the CCAMLR 2000 survey. Deep-Sea Res II 51:1421–1434
Kinder TH, Hunt GL Jr, Schneider D, Schumacher JD (1983) Correlations between seabirds and oceanic fronts around the Pribilof Islands, Alaska. Estuar Coast Shelf Sci 16:309–319
Lascara GL, Hofmann EE, Ross RM, Quentin LB (1999) Seasonal variability in the distribution of Antarctic krill, Euphausia superba, west of the Antarctic Peninsula. Deep-Sea Res I 46:951–984
Lawson GL, Wiebe PH, Ashjian CJ, Stanton TK (2008) Euphausiid distribution along the Western Antarctic Peninsula—Part B: distribution of euphausiid aggregations and biomass, and associations with environmental features. Deep-Sea Res II 55:432–454
Nicol S (2006) Krill, currents and sea ice: Euphausia superba and its changing environment. Bioscience 56:111–120
Nowacek DP, Friedlaender AS, Halpin PN, Hazen EL, Johnston DW et al (2011) Super-aggregations of krill and humpback whales in Wilhelmina Bay, Antarctic Peninsula. PLoS ONE 6(4):e19173. doi:10.1371/journal.pone.0019173
Orsi AH, Whithworth T III, Nowlin WD Jr (1995) On the meridional extent and fronts of the Antarctic Circumpolar Current. Deep Sea Res 42:641–673
Pankow H, Kock KH (2000) Results of a cetacean sighting survey in the Antarctic Peninsula region in November–December 1996. IWC SC/52/E23
Pocklington R (1979) An oceanographic interpretation of seabird distribution in the Indian Ocean. Mar Biol 51:9–21
Reid K, Brierley AS, Nevitt GA (2000) An initial examination of relationships between the distribution of whales and Antarctic krill Euphausia superba at South Georgia. J Cetacean Res Manag 2:143–149
Ribic CA, Ainley DG, Fraser WR (1991) Habitat selection by marine mammals in the marginal ice zone. Antarct Sci 3:181–186
Ribic CA, Ainley DG, Ford RG, Fraser WR, Tynan CT, Woehler E (2011) Water masses, ocean fronts, and the structure of Antarctic seabird communities: putting the eastern Bellingshausen Sea in perspective. Deep-Sea Res II 58:1695–1709
Ryan PG, Cooper J (1989) The distribution and abundance of aerial seabirds in relation to Antarctic krill in the Prydz Bay region, Antarctica during late summer. Polar Biol 10:199–209
Santora JA (2011) Habitat use of hourglass dolphins near the South Shetland Islands, Antarctica. Polar Biol. doi:10.1007/s00300-011-1133-8
Santora JA, Reiss CS (2011) Geopatial variability of krill and top predators within an Antarctic submarine canyon system. Mar Biol 158:2527–2540
Santora JA, Force MP, Ampela K, Van Cise AM (2009a) Chapter 8: Distribution abundance and behavior of seabirds and marine mammals. In: Van Cise AM (ed) Antarctic marine living resources program. 2008–2009 field station report, p 5
Santora JA, Reiss CS, Cossio AM, Veiti RR (2009b) Interannual spatial variability of krill (Euphausia superba) influences seabird foraging behavior near Elephant Island, Antarctica. Fish Oceanogr 18:20–35
Santora JA, Reiss CS, Loeb VJ, Veit RR (2010) Spatial association between hotspots of baleen whales and demographic patterns of Antarctic krill Euphausia superba suggests size-dependent predation. Mar Ecol Prog Ser 405:255–269
Scheidat M, Kock KH, Friedlaender A, Lehnert L, Williams R (2007) Preliminary results of aerial surveys around Elephant Island and the South Shetland Islands. IWC SC/59/IA21
Shirihai H (2007) A complete guide to Antarctic wildlife. The birds and marine mammals of the Antarctic continent and the Southern Ocean. A & C Black, London, p 544
Sirovic A, Hildebrand J, Thiele D (2006) Baleen whales in the Scotia Sea during January and February 2003. J Cetacean Res Manage 8:161–171
Tarling GA, Klevjer T, Fielding S, Watkins J, Atkinson A et al (2009) Variability and predictability of Antarctic krill swarm structure. Deep-Sea Res I 56:1994–2012
van Franeker JA (1992) Top predators as indicators for ecosystem events in the confluence zone and marginal ice zone of the Weddell and Scotia seas, Antarctica, November 1988 to January 1989 (EPOS leg 2). Polar Biol 12:93–102
Wynne-Edwards VC (1935) On the habits and distribution of birds of the North Atlantic. Proc Boston Soc Nat Hist 40:233–346
Acknowledgments
We are very grateful to AWI (Bremerhaven), late coordinator E. Fahrbach and chief scientist M. Lucassen for invitation on board RV Polarstern; observers were OD and C. Gruwier. Reviewers S. Geelhoed and D. Ainley kindly helped improving a first version of the ms.
Conflict of interest
None.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
About this article
Cite this article
Joiris, C.R., Dochy, O. A major autumn feeding ground for fin whales, southern fulmars and grey-headed albatrosses around the South Shetland Islands, Antarctica. Polar Biol 36, 1649–1658 (2013). https://doi.org/10.1007/s00300-013-1383-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-013-1383-8