
Abstract
The Southern Ocean occupies about 10 % of the world’s oceans but has low species richness with only ~1.5 % of the marine fishes. Within the Southern Ocean, the Ross Sea region is one of the least exploited sea areas in the world, but is subject to commercial fishing. The fauna are not well known, and preliminary IPY molecular studies have indicated that species diversity has been underestimated in this region. DNA barcodes of fishes from the Ross Sea region were compared with barcodes of fishes from the Atlantic and Indian Ocean sectors of the Southern Ocean. Barcoding resolved 87.5 % of 112 species that typically exhibited high inter-specific divergences. Intra-specific divergence was usually low with shared haplotypes among regions. The Zoarcid Ophthalmolycus amberensis showed shallow divergences (0.1 %) within the Ross Sea and Australian Antarctic Territory but high inter-region divergence (2 %), indicative of cryptic species. Other potential cryptic species with high intra-specific divergences were found in Notolepis coatsi and Gymnoscopelus bolini. In contrast, several taxa showed low inter-specific divergences and shared haplotypes among morphological species. COI provided limited phylogenetic resolution of the genera Pogonophryne and Bathydraco. Trematomus loennbergii and T. lepidorhinus shared COI haplotypes, as previously noted in other regions, as did Cryodraco antarcticus and C. atkinsoni. There was a marked lack of congruence between morphological descriptions and COI divergences among the Ross Sea liparids with shallow or zero divergences among recently described species. Barcodes for the Ross Sea fishes highlighted several initial misidentifications that were corrected when specimens were re-examined.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The Antarctic fish fauna has low species richness, with only ~ 320 species (~1.5 % of all known marine fishes) described from the Southern Ocean that occupies about 10 % of the world’s ocean (Eastman and McCune 2000; Clarke and Johnston 2003; Møller et al. 2005; De Broyer and Danis 2010). The number of species is probably an underestimate as some taxa are not well known, either because they occupy little sampled deep-water habitats or because they are especially difficult to identify (Duhamel et al. 2010). The unique cold-adapted fauna date from the late Eocene (~ 38 million years ago) and over the past 10–30 million years have developed into the world’s most distinctive fish fauna (Clark 2000; Møller et al. 2005), with a very high level of endemism, 97 % in the perciform suborder Notothenioidei (Eastman 1993). The Artedidraconidae, Bathydraconidae, Nototheniidae, and Channichthyidae (all in the Notothenioidei) are the most speciose families and dominate the shelf fish fauna in terms of species diversity, abundance, and biomass (Eastman and McCune 2000; DeVries and Steffensen 2005). The notothenioids have evolved to fill a wide range of life history and ecological niches similar to those observed in taxonomically unrelated shelf fishes in warmer waters and provide one of the few examples of a species flock of marine fishes (Eastman 2000; Eastman and McCune 2000).
Recent (pre-IPY, International Polar Year) molecular studies of this region have revealed an underestimated species richness together with restricted gene flow among some populations of fishes and invertebrates, in particular along the Antarctic Peninsula (e.g. Brierley et al. 1993; Allcock et al. 1997; Held and Leese 2007; Hunter and Halanych 2008; Smith et al. 2008; Thornhill et al. 2008; Wilson et al. 2009). However, studies have been constrained by limited sampling opportunities in this often hostile environment, and by a lack of well-preserved specimens suitable for molecular and taxonomic studies.
The dominant feature in the Southern Ocean is the Antarctic Circumpolar Current (ACC), which isolates the waters around Antarctica and prevents the intrusion of warm water into high latitudes (DeVries and Steffensen 2005). The ACC forms a physical barrier to the northward dispersion of eggs and larvae and also acts as west–east transport system (Eastman 1993). Much of the Southern Ocean is between 3,000 and 5,000 m in depth with isolated plateaus and ridges; the continental shelf area is relatively narrow except for large embayments in the Ross and Weddell Seas, which are characterized by clockwise gyres. The physical heterogeneity and gyres provide a marked spatial variability and, when coupled with the age of the bathyal habitats, may have promoted evolutionary divergence in this unique cold marine environment (Møller et al. 2005). Habitat fragmentation can restrict dispersal among populations, limit gene flow, and lead to allopatric speciation (Avise 2000; Frankham et al. 2004). Thus, species may consist of one genetic population with gene flow via the circumpolar current (Smith et al. 2008; Duhamel et al. 2010; Lautrédou et al. 2010; Lecointre et al. 2011) or two, or more, isolated populations contained within major gyres in the Ross and Weddell Seas (Orsi et al. 1995). Molecular studies of the larger and more abundant fishes, especially those vulnerable to harvesting, have indicated a major genetic break north and south of the sub-Antarctic convergence zone (Smith and McVeagh 2000; Shaw et al. 2004), but weak population structure, genetic homogeneity, and low genetic diversity, in some circum-Antarctic species (Parker et al. 2002; Smith and Gaffney 2005; Zane et al. 2006; Kuhn and Gaffney 2008; Van de Putte et al. 2012). A cryptic species was detected in the grenadier bycatch in the toothfish longline fishery (Smith et al. 2011a).
While bathymetry is a major barrier to dispersal, historical processes, in particular the repeated cycles of glaciation, may have had a major influence on the patterns of genetic diversity of benthic fishes (Janko et al. 2007). During glacial periods, ice cover would have limited the amount of benthic habitat on the continental shelf, but during inter-glacial periods, more habitat and possibly altered sea connections were available. The collapse of the West Antarctic Ice Sheet during the Pleistocene inter-glacial periods potentially created a direct connection between the Ross and Weddell Seas (Pollard and DeConto 2009). The impacts of these historical processes on contemporary patterns of genetic differentiation of benthic Antarctic species have been largely unstudied until very recently (see for example Janosik and Halanych 2010; Janosik et al. 2011).
Molecular barcoding provides a valuable molecular tool for the reliable identification of specimens, for flagging of taxonomic problems, and for providing insights into gene flow in and around the Southern Ocean (e.g. Smith et al. 2008, Grant et al. 2011). The barcode identification system is based on diversity in a single region of the mitochondrial DNA, the cytochrome c oxidase I gene, COI (Hebert et al. 2003; Ratnasingham and Hebert 2007), and has aroused considerable debate (see for example Moritz and Cicero 2004; DeSalle 2006; Rubinoff et al. 2006; Buhay 2009). Species recognition via DNA barcoding relies on different species having unique COI sequences or different assemblages of closely related sequences. Intra-specific variation or genetic distance is usually much less than inter-specific variation, enabling species identification and, when high, flagging possible cryptic species (Waugh 2007). While substantial overlap in intra- and inter-specific variation has been reported in a few taxa [some marine gastropods (Meyer and Paulay 2005) and corals (Shearer and Coffroth 2008)], in marine fishes around 98 % of species tested can be distinguished by COI barcodes (Ward et al. 2009), with the Antarctic notothenioids an exception (Dettai et al. 2011a).
Large-scale barcode projects with mostly temperate fishes have demonstrated the benefits of this approach (Ward et al. 2008, 2009), but there were relatively few barcode records for Antarctic fishes until the IPY, and these were restricted to the Scotia Sea (Rock et al. 2008). The IPY initiatives have provided more extensive geographical sampling and many more specimens of Southern Ocean fishes for molecular analyses (Lautrédou et al. 2010; Dettai et al. 2011a, b; Grant et al. 2011). The aims of this paper are to provide an overview of the barcode records for the Ross Sea fishes, collected during the IPY and other recent voyages, supplemented with specimens collected on commercial fishing vessels, and to provide a comparison of genetic divergence within the Ross Sea and between other regions of the Southern Ocean.
Materials and methods
Specimen and tissue collection
Antarctic fishes have been defined as those occurring south of the Antarctic Polar Front and so exclude those found around the sub-Antarctic Islands (Prince Edward, Crozet, Kerguelen, sub-Antarctic Islands south of New Zealand, and Burwood Bank) (Møller et al. 2005), although several species (e.g. Patagonian toothfish Dissostichus eleginoides) occur both north and south of the Antarctic Polar Front.
Specimens collected on research vessels during the IPY-CAML (Census of Antarctic Marine Life) surveys were tissue sampled at sea. Specimens caught on commercial vessels in the Southern Ocean were provisionally identified by fishery observers and the specimens frozen whole for onshore identification and processing. Muscle samples from fresh or thawed specimens were stored in 90 % ethanol and the specimens then fixed in 10 % formalin, prior to storage in 70 % isopropanol or 70 % ethanol. The majority of specimens were deposited into collections in the Muséum national d‘Histoire naturelle, Paris (567 specimens), Museum of New Zealand Te Papa Tongarewa (488), Australian Antarctic Division (215), CSIRO Australian National Fish Collection (94), the New Zealand National Institute of Water & Atmospheric Research (21), and the South African Institute for Aquatic Biodiversity (2). Provisional barcode results have been reported for specimens deposited in the Muséum national d‘Histoire naturelle collection (Dettai et al. 2011a, b) and for specimens in the genus Macrourus (Smith et al. 2011a). A further 371 pre-IPY specimens from the Scotia Sea were barcoded by Bangor University (Rock et al. 2008).
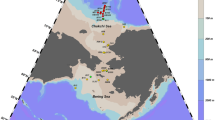
The majority of the specimens came from research voyages in three areas: the Ross Sea region (the Pacific sector); the Australian Antarctic Territory (the Indian Ocean sector); and the Scotia and Weddell Seas (the Atlantic sector) (Fig. 1). Specimens collected through Fisheries Observer Programmes covered the Ross Sea region, the Bellingshausen Sea (78–102oW), the Amundsen Sea (131–172oE), and the South Indian Ocean basin around Heard and McDonald Islands. Large species were not well sampled by IPY research voyages; for example, during the RV Tangaroa IPY-CAML survey, only two specimens of the Antarctic toothfish Dissostichus mawsoni and five specimens of Bathyraja were collected, yet these are the target and major bycatch species, respectively, in the toothfish longline fisheries in the same region (Smith et al. 2008).

Locations of fish specimens sampled in the Southern Ocean for DNA barcoding. AAA Australian Antarctic Territory, HAM Heard and McDonald Is., RS Ross Sea, SS Scotia Sea, WS Weddell Sea
Laboratory methods
DNA was extracted from a subsample of muscle tissue from each specimen using an automated Glass Fiber protocol (Ivanova et al. 2006). The 650-bp barcode region of COI was amplified under standard conditions using the primer cocktail FishF1t1 and FishR1t1 (Ivanova et al. 2007). PCR products were visualized on a 1.2 % agarose gel E-GelH (Invitrogen) and sequenced in both directions using the sequencing primers M13F and M13R (Ivanova et al. 2007) and the BigDyeH Terminator v.3.1 Cycle Sequencing Kit (Applied Biosystems, Inc.) on an ABI 3730 capillary sequencer following manufacturers instructions. Sequences were deposited in the Barcode of Life Data System, BOLD (Ratnasingham and Hebert 2007), in the public projects: Fishes of Antarctica (FNZB) and Fishes of the Australian Antarctic Territory (FOAAN). Further data sets, East Antarctic Teleost Fishes (EATF, Dettai et al. 2011a), Scotia Sea fish (ANTFI, Rock et al. 2008), and Rattails in the Southern Ocean (RATSO, Smith et al. 2011a), were included to provide a wide geographic coverage total of fishes barcoded from the Southern Ocean.
DNA analyses
Species with <3 specimens (n = 45) were deleted from the overall data set, as were specimens identified only to genus level, including species with provisional names. Sequence divergences within and among species were calculated using the Kimura two-parameter (K2P) distance model (Kimura 1981). A neighbour-joining (NJ) tree of K2P distances was created to provide a graphic representation of the patterning of divergence within and among species using MEGA v 4 (Tamura et al. 2007); support for each inter-node was evaluated by 1,000 bootstrap replications (Felsenstein 1985). These preliminary analyses of the Antarctic fishes COI data set revealed some instances of high intra-specific divergence (>2 %), highlighting a few possible misidentifications and also some potential cryptic species. A review of 1088 species of marine and freshwater fishes reported a mean K2P intra-specific distance of 0.343 ± 0.005, and a 2 % COI divergence had only a 3 % probability of conspecificity (Ward 2009). Specimens were re-checked to confirm or to revise the initial identifications. Tissue samples from some of the potential cryptic species were amplified for the nuclear rhodopsin gene using primers developed for fishes (Sevilla et al. 2007).
Subsequently maximum likelihood (ML) trees were built using PAUP 4.0b10 (Swofford 2003) with heuristic searches, employing tree bisection-reconnection branch swapping; support for each inter-node was evaluated by 1,000 bootstrap replications (Felsenstein 1985). Bayesian phylogenetic analyses were estimated with MrBayes version 3.0 (Huelsenbeck and Ronquist 2001). Four simultaneous Monte Carlo chains were run for 1 × 107 generations, saving the current tree every 1,000 generations. Consensus trees with posterior probabilities were created with a burn-in value equal to 1,000 (the first 1,000 trees were discarded). Nucleotide substitution models were selected in jModeltest version 0.1.1 (Posada 2008) using Akaike information criterion (AIC) and Bayesian information criterion (BIC).
COI sequences for the Ross Sea fishes (FNZ) have been deposited in GenBank, Accession Numbers JN640756–641178; additional COI sequences from the Australian Antarctic Division (FOAG) and the CSIRO Australian National Fish Collection (FOAG) were deposited in GenBank as JN640573–640754. Existing COI sequences were used from the Australian Antarctic Division (FOAG): GU805968–60005; Muséum national d‘Histoire naturelle (EATF): HQ712804–713372; Bangor University (ANTFI): EU326313–326436; and Rattails in the Southern Ocean (RATSO): JF265072–265128.
Results and discussion
The overall data set contained 1,445 specimens and 201 putative species of Southern Ocean fishes. Examination of the initial COI data revealed several instances of high intra-specific divergence (>2 %), suggesting possible identification errors or cryptic species (see Ward 2009), in particular from the Ross Sea region. Misidentifications are not surprising as some taxa are not well known or have been described from relatively few specimens from one region. Re-checking specimens confirmed that many of these instances did reflect initial misidentifications and most corrected identifications subsequently showed low intra-specific divergences.
A reduced data set of 1,209 specimens and 112 species, based on 3 or more specimens per species and excluding specimens identified to genus level only, showed that the mean intra-specific divergence was 0.33 % and ranged from 0 to 3.95 %. This is similar to that reported in 294 species of mostly temperate fishes, including freshwater taxa, with a mean of 0.35 % and a range of 0–10.91 % (Ward et al. 2009; the higher values reflecting a few likely misidentifications and/or cryptic species). The mean intra-generic divergence among 58 genera of Southern Ocean fishes was 5.45 % and ranged from 0 to 13.11 %. This is lower than that observed among 103 genera of mostly temperate fishes, including freshwater taxa, with a mean of 8.11 % and a range of 0–24.18 % (Ward et al. 2009). A summary list of species, Specimen Reference Numbers, GenBank Accession Numbers, BOLD Process Numbers, and Institutes holding specimens is provided in Appendix 1 in Electronic supplementary material.
Sixty-four species were sampled in the Ross Sea and at least one other region. Most widely distributed species showed little or no sequence divergence among regions, with a common shared haplotype. However, high intra-specific divergence remained for several taxa after specimen re-examination. This is indicative of cryptic species and appeared more common within the Ross Sea than between regions. On the other hand, several taxa showed no COI divergence among morphological species, with only 87.5 % of species resolved by COI, appreciably less than the 98 % reported for marine fishes in general (Ward et al. 2009). It is possible that there has been insufficient time for COI differentiation to have occurred among these recently diverged species; alternatively, morphological descriptions of some species might have been premature and based on relatively few variable specimens. The overall COI data set revealed interesting taxonomic issues in several taxa, as detailed below.
Rajiformes
Skates are the dominant chondrichthyan group in Antarctic waters (Long 1994), but few specimens were captured during IPY voyages. Most specimens of Bathyraja and Amblyraja were taken as bycatch on toothfish longline vessels. Molecular analyses of cytochrome b and COI in the Southern Ocean Bathyraja have shown high sequence divergences (5.0–5.7 %) among specimens of B. eatonii from the Kerguelen Plateau (the type locality) and the Antarctic shelf, providing evidence for two allopatric species (Smith et al. 2008): B. eatonii from the Kerguelen Plateau and an undescribed species Bathyraja sp. (cf. eatonii) in the Southern Ocean. The DNA sequence divergences indicated significant differentiation among ocean basins in Bathyraja sp. (cf. eatonii) and in Bathyraja maccaini, but not within Bathyraja sp. (dwarf) (Smith et al. 2008). Specimens of the starry skate Amblyraja georgiana from the Ross Sea region (n = 6) and the Scotia Ridge (n = 6) had identical COI haplotypes.
Aulopiformes: Notolepis coatsi (Paralepididae: barracudinas)
Two species are recorded in the genus Notolepis in Antarctic waters: the Antarctic jonasfish Notolepis coatsi (circumpolar 50–73oS, with a black gill chamber) and the ringed barracudina Notolepis annulata (Western Atlantic 37°–72°S, possibly circumpolar, with a pale gill chamber, and distinctive light and dark stripes). Specimens from the Ross Sea region were identified as N. coatsi, with a dark gill chamber and inconspicuous scales, and did not have the distinctive rings and scale extensions of N. annulata. The COI data for N. coatsi indicated a high sequence divergence (2.8 %) with 2 clades (Fig. 2), with 12/643 parsimony informative sites, indicative of 2 species. Three specimens of N. coatsi from the Indian Ocean, Heard and McDonald region (76oE, 52oS, 300 m), and one specimen from the Australian Antarctic Territory (139oE, 61oS, 1,000 m) grouped with specimens from the Pacific North Scott seamounts (179oW, 68oS, 50 m depth), and not those in the second clade from the northern edge of Mawson Bank (177oE, 72oS, 1,403 m depth). Re-examination of the Ross Sea specimens in the light of the barcode results found no morphometric differences among representatives of the two clades, although the specimens were delicate and in poor condition. Specimens from the North Scott seamounts had 10–11 pectoral fin rays (mostly 10), while those from the northern edge of Mawson Bank had 10–11 pectoral fin rays (mostly 11). No specimens or COI sequences of N. annulata were available for comparison. Amplification of 456 base pairs of the nuclear rhodopsin gene showed similar sequence compositions, with no divergence parallel to the COI clades, among the 11 Ross Sea specimens of N. coatsi (GenBank Accession Numbers JN603596–JN603606). Further taxonomic study of this species is required as/when specimens are available.

COI relationships among specimens of the jonasfish Notolepis coatsi from the Ross Sea region (North Scott Seamounts, 52°S–68°S; and the northern edge of Mawson Bank, 72°S) and wider Southern Ocean (FNZB = Ross Sea, FOA and EATF = Australian Antarctic Territory). Unrooted ML tree; numbers at nodes are bootstrap percentage (>75 %) after 1,000 replicates based on ML and Bayesian inference posterior probability value (>0.9); scale bar is TIM1 +G distance. Sequence code numbers represent BOLD Process Numbers
Osmeriformes
Fifteen specimens of the deep-sea smelt Bathylagus antarcticus were sampled from the Ross Sea (10) and Australian Antarctic Territory (5) and showed high initial intra-specific divergence (mean 5.26 %) produced by one specimen (UM7142P48) representing a mislabelled tissue (of Cynomacrurus piriei); removal of this specimen resulted in low intra-specific divergence (0.63 %), with no evidence for regional haplotypes.
Myctophiformes
Gymnoscopelus (Myctophidae: lanternfishes)
Specimens from six species of lanternfishes of the genus Gymnoscopelus were sequenced for COI. Three species appeared in three well-supported clades: G. opisthopterus, G. braueri, and G. nicholsi, all with low K2P intra-specific divergences (Table 1). Among specimens of G. bolini, there were a high divergence (2.72 %, Table 1) and two well-supported clades, typical of species-level differentiation in this genus (Fig. 3), with 10/646 parsimony informative sites. Re-examination of the three Ross Sea specimens of G. bolini (Fig. 3) showed them to be correctly identified based on gill raker count, caudal photophore count, and photophore position; all 3 specimens were large (208–231 mm SL) and taken from the same tow. Unfortunately, no further samples of G. bolini were available for analysis; those available did not amplify for the nuclear rhodopsin gene. Specimens of two species Gymnoscopelus hintonoides and G. piabilis, and one specimen of G. microlampas appeared in a shallow, mixed species clade. Preserved specimens of Gymnoscopelus held at NMNZ were re-examined, and misidentifications highlighted by the barcode results were confirmed by morphometric and meristic re-examinations: specimens purportedly of both G. microlampas and G. piabilis were found to be G. hintonoides.

COI relationships among specimens of the lanternfish Gymnoscopelus bolini from the Ross Sea (FNZ and FNZB sequences) and Australian Antarctic Territory (FOAE and FOAG sequences). ML tree, rooted with Gymnoscopelus nicholsi. The scale bar represents an interval of the K2P +G model; numbers at nodes are bootstrap percentage (>75 %) after 1,000 replicates based on ML and Bayesian inference posterior probability value (>0.9). Sequence code numbers represent BOLD Process Numbers
Thirteen specimens of the lanternfish Nannobrachium achirus from the Ross Sea region (11) and Australian Antarctic Territory (2) showed shallow divergence (0.36 %) with a shared haplotype between regions (Table 1). One specimen initially misidentified as the deep-water lanternfish Taaningichthys bathyphilus had an identical haplotype to Nannobrachium achirus from the Ross Sea. Two other myctophids, Electrona antarctica and E. carlsbergi, also showed shallow intra-specific divergences among regions (Table 1) with no region-specific haplotypes.
Gadiformes
Three species of grenadier in the genus Macrourus have been recognized in the Southern Ocean, but DNA sequencing of the mitochondrial COI gene revealed four well-supported clades, suggesting the presence of an undescribed species, a conclusion subsequently supported by meristic and morphometric examination of specimens (Smith et al. 2011a). The four species were characterized by low intra-specific divergences (0.0–0.1 %) with no region-specific haplotypes (Smith et al. 2011a). Specimens of the undescribed species Macrourus sp had been misidentified as M. whitsoni, and the two species were sympatric in the Ross Sea region, being captured over the same depth range and on the same longline set in the toothfish fishery (Smith et al. 2011a). Most of the specimens captured in the CEAMARC cruises in the Australian Antarctic Territory (FAO 58) and provisionally identified as M. whitsoni (Dettai et al. 2011a) had identical COI sequences to Macrourus sp. Two specimens of M. whitsoni were identified from one deep-water station, where they were the only macrourids (depth 1,700 m, all the Macrourus sp. were taken from <850 m), indicating that the two species might not be sympatric throughout their ranges.
Another grenadier, the dogtooth grenadier Cynomacrurus piriei, had zero intra-specific divergence with a single haplotype in specimens from the Ross Sea (n = 8) and other regions (n = 4).
The slender codling Halargyreus johnsonii is a bentho-pelagic species found on the continental slope at about 450–3,000 m with an anti-tropical distribution in the Atlantic and Pacific Oceans, including the Southern Ocean. One COI haplotype was found among specimens from the Ross Sea region (n = 5) and the Australian Antarctic Territory (n = 6), but this haplotype was highly divergent from those from specimens collected around New Zealand (mean 4.2 %). Average sequence divergences were only 0 and 0.1 %, for specimens, respectively, from the pooled North Pacific, Atlantic, and Southern Oceans and from New Zealand (Smith et al. 2011b). The high sequence divergence is typical of congeneric fish species and indicates the presence of a cryptic species within what is currently recognized as H. johnsonii (Smith et al. 2011b). Meristic data for specimens from New Zealand and from the Southern Ocean north of the Ross Sea supported this conclusion (Smith et al. 2011b). DNA barcodes for another morid cod Antimora rostrata showed low intra-species divergence (0.1–0.2 %) among specimens from the Ross Sea region (17) and AAT (2), and low divergence (0.22 %) with the congeneric A. microlepis (Smith et al. 2011b).
Scorpaeniformes Liparidae (snailfishes)
A recent review of the taxonomy of the Ross Sea liparids has listed 34 species in three genera: six Careproctus, 27 Paraliparis, and one Genioliparis species (Stein 2011, submitted). Eighteen new species of Paraliparis were described, 11 from Mawson Bank alone, and many from single specimens (Stein 2011, submitted). In contrast to the morphological data, the COI data provided little resolution for many of the newly described species of Paraliparis; three single specimens of three of the new species from the Mawson Bank (P.043687, P.043691, and P.043719) shared a COI haplotype; a further six specimens of another six species (P.043718, P.043692, P.043689, P.043690, P.043688, and P.043721) shared a second haplotype. Two additional new species (P.043693 and P.045678) from the Ross Sea had the same COI haplotype as P. valentinae from the AAT. Unlike the Ross Sea data set, the AAT data set showed higher inter-specific divergences (mean = 4.27 %, range = 0.7–6.4 %) with distinct haplotypes among recognized taxa (P. antarcticus, P. charcoti, P. leobergi, and P. mawsoni) sampled with multiple specimens, and low (mean = 0.09 %, range = 0.0–0.3 %) intra-specific divergences (Table 2). Intra- and inter-clade divergences among Ross Sea specimens are similar to intra- and inter-specific divergences among the AAT taxa (Table 2). It is possible that there has been insufficient time for COI differentiation to occur among the newly described and recently diverged species in the Ross Sea region, but the lack of congruence between morphological and COI data is inconsistent. COI data for the circum-Antarctic Paraliparis antarcticus from the Ross Sea region and the AAT revealed two shallow clades with significant divergence (89 % bootstrap support, ML phylogenetic tree) of 0.5 %, with 3/648 phylogenetic informative sites; within-clade divergences were zero. However, two of the Ross Sea specimens appeared in the AAT clade, suggesting that the variation here reflects intra-specific rather than inter-specific differentiation.
A comparison of the Antarctic liparid COI sequences with northern hemisphere liparid COI sequences (available in GenBank, Steinke et al. 2009) showed a complex pattern of relationships with some northern taxa in sister clades to Ross Sea and AAT taxa (data not presented here). A similar case was reported for the liparids of the AAT Antarctic shelf and Kerguelen Islands (Duhamel et al. 2010). The COI results provide a very different pattern to that based on morphology and geography (Duhamel et al. 2010) and highlight the need for additional molecular and morphological studies of this family.
Perciformes
Artedidraconidae (barbeled plunderfishes)
The genus Pogonophryne contains many species that are difficult to identify morphologically, in part due to descriptions that were based on relatively few specimens from just one area (Balushkin and Eakin 1998; Eakin 1990; Eakin and Eastman 1998). A revised key to the Artedidraconidae noted that some species exhibited considerable intra-specific morphological variation (Eakin unpublished key 2006).
Currently, 19 species of Pogonophryne, in five species groups, are recognized (Eakin et al. 2009). The first phylogenetic analysis of the Pogonophryne, based on a single mitochondrial gene ND2 and including 11 of the 19 recognized species (sampled from the five proposed species groups), showed a reciprocal monophyly of the monotypic species groups (Eakin et al. 2009). Sequence divergence was shallow among the 11 species, with a maximum pairwise genetic distance of 1.4 % between P. scotti and P. marmorata, and with limited phylogenetic resolution among the five species groups (Eakin et al. 2009).
COI sequence data for Ross Sea specimens showed that the same species name had been applied to specimens with different COI sequences and that different names had been applied to specimens with similar COI sequences, highlighting a large number of initial misidentifications, and also questioning the resolution of COI among the closely related taxa. Specimens were re-examined by a specialist (J. Eastman, Ohio University) and some identifications revised. Specimens provisionally identified as P. immaculata, P. albipinna, and P. barsukovi appeared in the same clade, while specimens of P. eakini, P. mentella, P. squamibarbata, and P. orangiensis appeared in a second clade; specimens of P. scotti formed a third discrete clade, with a shared haplotype among specimens from the Ross Sea and Australian Antarctic Territory (Table 3, Appendix 2 in ESM).
Three species, Artedidraco skottsbergi, Dolloidraco longedorsalis, and Histiodraco velifer, showed no evidence for region-specific haplotypes (Table 3). However, the low inter- and intra-specific divergences noted in genus Pogonophryne were also present in the other artedidraconids: although there are species-specific clusters, they are only separated by a few base pairs (Dettai et al. 2011a; Lecointre et al. 2011). Consequently, the lack of region-specific haplotypes might reflect generally low variability of COI. Low genetic divergence among artedidraconid species has been reported with three mitochondrial and one nuclear DNA marker, indicating very recent divergence of small species “subflocks” within the notothenioid flock (Lecointre et al. 2011).
Nototheniidae (cod icefishes)
The Nototheniidae family has a relatively long history of phylogenetic studies employing a range of mitochondrial and nuclear DNA markers (Ritchie et al. 1996; Near et al. 2004; Janko et al. 2007; Sanchez et al. 2007; Kuhn and Near 2009; Lautrédou et al. 2010). The recent COI study by Lautrédou et al. (2010) included the greatest number of specimens (220) and species (12), but few (4 specimens, 3 species) were from the Ross Sea region.
Two species of Trematomus were sequenced from both the Ross Sea and other regions and appeared in two well-supported clades. Trematomus scotti showed shallow K2P divergence (D = 0.82 %, Table 4) and shared haplotypes among Ross Sea and Indian Ocean regions, as did T. eulepidotus (D = 0.68 %, Table 4). Several Ross Sea specimens initially identified as T. eulepidotus were shown to be Lepidonotothen squamifrons based on COI sequence data and specimen re-examination.
Ross Sea specimens identified as T. loennbergii and T. lepidorhinus appeared in the same COI clade (Appendix 3 in ESM). Lautrédou et al. (2010) also reported a lack of COI divergence among T. loennbergii and T. lepidorhinus from the Indian Ocean sector and commented that although they are currently considered valid species, the morphological distinctions present problems and that no conclusion can be drawn on the status of these species until morphologies and more variable nuclear markers are investigated (Lautrédou et al. 2010). Certainly, the two species are morphologically similar, although T. lepidorhinus has a scaled snout to upper jaw, and scaled lower jaw and branchiostegal membrane, while T. loennbergii lacks scales at the tip of the snout and lacks scales on lower jaw and branchiostegal membrane.
Two species of Lepidonotothen sampled from Ross Sea and Kerguelen Islands showed a K2P inter-specific divergence of 0.19 % and low intra-specific sequence divergences (L. larseni 0.22 %, L. squamifrons 0.18 %) with shared intra-specific haplotypes between regions (Table 4). Several Ross Sea specimens were identified as L. kempi, a synonym of L. squamifrons (Eschmeyer and Fricke 2009). The Antarctic silverfish Pleuragramma antarctica also showed low intra-specific sequence divergence (D = 0.21 %) with one common haplotype among specimens from the Ross Sea and AAT (Table 4).
Channichthyidae (crocodile icefishes)
Fifteen species of crocodile icefishes are found around Antarctica and off South America (Gon and Heemstra 1990); 11 were sequenced for COI, 10 from two regions. Nine species with specimens from the Ross Sea and Australian Antarctic Territory had shallow K2P intra-specific divergences (0.0–0.3 %) with shared intra-specific haplotypes among the two regions (Table 5). Chionobathyscus dewitti is probably circumpolar occurring on the continental shelf and slope between 500 and 2000 m. Specimens of C. dewitti from the Ross Sea region revealed two clades with shallow divergence 0.3 % (<65 % bootstrap support ML, K2P distance) with specimens in the two clades differing by two base substitutions, both at third codon positions with no change in amino acids. There was no evidence for geographical structure with specimens in both clades sampled between 64 and 72oS, and some specimens from the same tow appearing in both clades. Two specimens from the Australian Antarctic Territory had identical sequences to those from the Ross Sea in clade one. It is likely that this shallow sequence divergence represents intra-specific variation. Other crocodile icefishes sampled in two regions (Chaenodraco wilsoni, Chionodraco myersi, Chionodraco hamatus, Dacodraco hunteri, Neopagetopsis ionah, Pagetopsis macropterus, and Pagetopsis maculatus) showed shallow (non-robust < 50 % bootstrap support) intra-specific divergences (<0.31 %, Table 5), as did Channichthys rhinoceratus sampled only in the AAT (mean D = 0.05 %, range = 0–0.31).
Ross Sea specimens identified as Cryodraco antarcticus and C. atkinsoni appeared in one well-supported clade with two shallow subclades, one C. antarcticus clade and one mixed C. antarcticus and C. atkinsoni clade that differed by one base-pair, at a third codon position, and had weak bootstrap support (ML K2P distance 62 %, out-group Chionodraco myseri). A combined Ross Sea–EATF data set showed that the same two common haplotypes were shared among specimens of C. antarcticus from the Ross Sea and Australian Antarctic Territory, with two additional haplotypes in the AAT, that differed at a single base position (Fig. 4). Re-examination of the Ross Sea region specimens based on the COI results highlighted several initial misidentifications, but overall, there remained specimens attributed to each species, based on a combination of characters: pelvic fin length, head length, number of second dorsal fin rays, and origin of lower lateral line, after La Mesa et al. (2002). Ross Sea specimens identified as C. atkinsoni shared haplotypes with C. antarcticus from the Ross Sea and AAT. Gon and Heemstra (1990) regarded C. atkinsoni as a junior synonym of C. antarcticus and suggested that the taxonomic status of C. antarcticus should be re-evaluated (Gon and Heemstra 1990). Subsequently, a taxonomic study of Cryodraco, based on more specimens, recognized Cryodraco antarcticus and C. atkinsoni as good species, both with a circum-Antarctic distribution (La Mesa et al. 2002) (see also Eschmeyer and Fricke 2009). An early molecular phylogeny of the Channichthyidae did not include C. atkinsoni (Chen et al. 1998), while a DNA phylogeny based on three mitochondrial genes, including COI, showed shallow divergence between 2 specimens of C. antarcticus and C. atkinsoni (Near et al. 2003), but COI sequences were not provided.

COI relationships among the Cryodraco antarcticus and C. atkinsoni specimens from the Ross Sea (FNZB sequence code numbers) and AAT (EATF sequence code numbers); ML tree, rooted with Chionodraco myersi. The scale bar represents an interval of the K2P model; the nodes were not well supported with bootstrap percentages <75 %. Sequence code numbers represent BOLD Process Numbers
Bathydraconidae (Antarctic dragonfishes)
Three circum-Antarctic species, Vomeridens infuscipinnis, Prionodraco evansii, and Gerlachea australis, each showed shallow K2P intra-specific divergences and shared intra-specific haplotypes among regions (Table 6 and Appendix 4 in ESM). Two Ross Sea specimens of V. infuscipinnis were initially misidentified as Bathydraco macrolepis. Specimens identified as Gymnodraco victori and G. acuticeps from the Australian Antarctic Territory shared haplotypes (Appendix 4).
Four Bathydraco species showed shallow divergence (Table 6), with shared haplotypes among B. marri and B. macrolepis and among B. antarcticus and B. joannae (Appendix 3). Rock et al. (2008) also reported a lack of species resolution within the Bathydraco in the Scotia Sea with both COI (B. macrolepis, B. antarcticus, and B. joannae) and cytochrome b (B. macrolepis, B. marri, B. antarcticus, and B. joannae). An earlier molecular study utilizing the mitochondrial control region and cytochrome b indicated a rapid diversification in the bathydraconids with just three clades across nine genera (Derome et al. 2002). Given that morphological discrimination between bathydraconids is relatively straightforward, and assuming that the morphological variants do indeed equate to true biological species, additional faster-evolving nuclear DNA markers will be required for molecular identification of species of Bathydraco (Rock et al. 2008).
Zoarcidae (Eelpouts)
The bathydemersal circum-Antarctic Ophthalmolycus amberensis occurs between 60 and 78°S at depths 140–826 m. Five specimens of O. amberenesis from the Ross Sea region had very different haplotypes to 8 specimens from the Australian Antarctic Territory (Fig. 5); within-region divergences were 0.1 %, and the between-region divergence was 2.0 %, with 11/651 parsimony informative sites. The high between-region divergence is indicative of species-level divergence. Additional specimens and tissue samples are needed from the type locality (Isla Amberes, Antarctic Peninsula, 64° 40′S, 63° 20′W) to resolve the taxonomic status of specimens from the Ross Sea and AAT, with specimens from at least one region representing an undescribed species.

COI relationships among the Zoacidae Ophthalmolycus amberensis and Pachycara specimens from the Ross Sea (FNZ and FNZB sequence code numbers) and AAT (EATF sequence code numbers); ML tree, rooted with Pachycara brachycephalum (FNZB567-06) from the Ross Sea. The scale bar represents an interval of the TN93 +G model; numbers at nodes are bootstrap percentages (>75 %). Sequence code numbers represent BOLD Process Numbers
Ross Sea specimens identified as Pachycara, including Pachycara brachycephalum, appeared in a discrete clade from P. brachycephalum from the Australian Antarctic Territory with high sequence divergence (1.5 %) and appeared to represent an undescribed species (Fig. 5).
Conclusions
Comparison of the 64 species of Southern Ocean fishes considered here has shown that most show low intra-specific divergence with shared haplotypes between Ocean basins in the Southern Ocean. A few taxa with deep intra-specific divergences, indicative of cryptic species, were found in Notolepis coatsi, Gymnoscopelus bolini, and Ophthalmolycus amberensis and need to be validated with additional specimens and with additional DNA markers and detailed morphometric analyses. Two species of Macrourus, M. whitsoni, and Macrourus sp. are sympatric in the Ross Sea region but not in the AAT, and additional specimens from different areas and depths are required to establish the distribution of these two species and their vulnerabilities in the toothfish fisheries in the Southern Ocean. COI provided limited phylogenetic resolution of species in the genera Pogonophryne, Bathydraco, and Cryodraco; overall COI separated only 87 % of 112 currently recognized Southern Ocean fishes, much less than the 98 % observed for temperate fishes (Ward et al. 2009). This reduced success rate is consistent with recent divergence within these genera, and the categorization of this Southern Ocean assemblage as a species swarm (Eastman 2000; Eastman and McCune 2000). In the liparid genus Paraliparis, there was a lack of congruence between morphological descriptions and COI diversity with little or no COI divergence among species recently described from the Ross Sea. Additional specimens and both mitochondrial and nuclear DNA markers are needed to resolve the taxonomic status of these species.
References
Allcock I, Brierley AS, Thorpe J, Rodhouse PG (1997) Restricted geneflow and evolutionary divergence between geographically separated populations of the Antarctic octopus Pareledone turqueti. Mar Biol 129:97–102
Avise JC (2000) Phylogeography: the history and formation of species. Harvard University Press, Cambridge, p 477
Balushkin A, Eakin R (1998) A new toad plunderfish Pogonophryne fusca sp. nova (Fam. Artedidraconidae: Notothenioidei) with notes on species composition and species groups in the genus Pogonophryne Regan. J Ichthyol 38:574–579
Brierley AS, Rodhouse PG, Thorpe JP, Clarke MR (1993) Genetic evidence of population heterogeneity and cryptic speciation in the ommastrephid squid Martialia hyadesi from the Patagonian Shelf and Antarctic Polar Front Zone. Mar Biol 16:593–602
Buhay JE (2009) “COI-like” sequences are becoming problematic in molecular systematic and DNA barcoding studies. J Crust Biol 29:96–110
Chen WJ, Bonillo C, Lecointre G (1998) Phylogeny of the Channichthyidae (Notothenioidei, Teleostei) based on two mitochondrial genes In: di Prisco G, Pisano E, Clarke A (eds) Fishes of Antarctica: a biological overview. Springer-Verlag Italia, Milano, 1998. ISBN 88 470 0028 9
Clark AM (2000) Benthic organisms and environmental variability in Antarctica: responses to seasonal, decadal and long-term change. Ocean Polar Res 23:433–440
Clarke A, Johnston N (2003) Antarctic marine benthic diversity. Oceanogr Mar Biol Ann Rev 41:47–114
De Broyer C, Danis B (2010) How many species in the Southern Ocean? Towards a dynamic inventory of the Antarctic marine species. Deep Sea Res II 58:5–17
Derome N, Chen WJ, Dettai A, Bonillo C, Lecointre G (2002) Phylogeny of Antarctic dragonfishes (Bathydraconidae, Notothenioidei, Teleostei) and related families based on their anatomy and two mitochondrial genes. Mol Phylogenet Evol 24:139–152
DeSalle R (2006) Species discovery versus species identification in DNA barcoding efforts: response to Rubinoff. Cons Biol 20:1545–1547
Dettai A et al (2011a) The actinopterygian diversity of the CEAMARCcruises: barcoding and molecular taxonomy as a multi level tool for new findings. Deep Sea Res II 58:250–263
Dettai A et al (2011b) Barcoding and molecular systematics of the benthic and demersal organisms of the CEAMARC survey. Polar Sci 5:298–312
DeVries A, Steffensen J (2005) The Arctic and Antarctic polar marine environments. Physiol Fish 22:1–24
Duhamel G, Hautecoeur M, Dettai A, Causse R, Pruvost P, Busson F, Couloux, Koubbi P, Williams R, Ozouf-Costaz C, Nowara G (2010) Liparids from the Eastern sector of Southern Ocean and first information from molecular studies. Cybium 34:319–343
Eakin R (1990) Artedidraconidae. In: Gon O, Heemstra P (eds) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, pp 332–356
Eakin R, Eastman J (1998) New species of Pogonophryne (Pisces, Artedidraconidae) from the Ross Sea, Antarctica. Copeia 1998:1005–1009
Eakin RR, Eastman JT, Near TJ (2009) A new species and a molecular phylogenetic analysis of the Antarctic fish genus Pogonophryne (Notothenioidei: Artedidraconidae). Copeia 2009:705–713
Eastman J (1993) Antarctic fish biology: evolution in a unique environment. Academic Press, San Diego, p 322
Eastman JT (2000) Antarctic notothenioid fishes as subjects for research in evolutionary biology. Antarct Sci 12:276–287
Eastman J, McCune A (2000) Fishes on the Antarctic continental shelf: evolution of a marine species flock? J Fish Biol 57:84–102
Eschmeyer WN, Fricke R (2009) Catalog of Fishes electronic version (9 September 2009). http://research.calacademy.org/ichthyology/catalog/fishcatmain.asp
Felsenstein J (1985) Confidence limits on phylogenies: an approach using the bootstrap. Evolution 39:783–791
Frankham R, Ballou J, Briscoe D (2004) A primer of conservation genetics. Cambridge University Press, Cambridge, p 234
Gon O, Heemstra P (1990) Fishes of the Southern Ocean. JLB Smith Institute of Ichthyology, Grahamstown, p 462
Grant RA, Griffiths H, Steinke D, Linse K (2011) Antarctic DNA barcoding; a drop in the ocean? Polar Biol 34:775–780
Hebert PDN, Ratnasingham S, deWaard JR (2003) Barcoding animal life: cytochrome c oxidase subunit 1 divergences among closely related species. Proc R Soc Lond Biol Sci 270:S96–S99
Held C, Leese F (2007) The utility of fast evolving molecular markers for studying speciation in the Antarctic benthos. Polar Biol 30:513–521
Huelsenbeck JP, Ronquist F (2001) MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755
Hunter RL, Halanych KM (2008) Evaluating connectivity in the brooding brittle star Astrotoma agassizii across the drake passage in the Southern Ocean. J Hered 99:137–148
Ivanova NV, deWaard JR, Heber PDN (2006) An inexpensive, automation-friendly protocol for recovering high-quality DNA. Mol Ecol Notes 6:998–1002
Ivanova NV, Zemlak TS, Hanner RH, Hebert PDN (2007) Universal primer cocktails for fish DNA barcoding. Mol Ecol Notes 7:544–548
Janko K, Lecointre G, DeVries D, Couloux A, Cruaud C, Marshall C (2007) Did glacial advances during the Pleistocene influence differently the demographic histories of benthic and pelagic Antarctic shelf fishes? Inferences from intraspecific mitochondrial and nuclear DNA sequence diversity. BMC Evol Biol 7:220
Janosik AM, Halanych KM (2010) Unrecognized Antarctic biodiversity: a case study with Odontaster (Odontasteridae; Asteroidea). J Int Comp Biol 50:981–992
Janosik AM, Mahon AR, Halanych KM (2011) Evolutionary history of Southern Ocean seastar species across the Drake Passage, and the discovery of 2 species of Odontaster (Odontasteridae; Asteroidea). Polar Biol 34:575–586
Kimura M (1981) Estimation of evolutionary distances between homologous nucleotide sequences. Proc Nat Acad Sci 78:454–458
Kuhn KL, Gaffney PM (2008) Population subdivision in the Antarctic toothfish (Dissostichus mawsoni) revealed by mitochondrial and nuclear single nucleotide polymorphisms (SNPs). Antarct Sci 20:327–338
Kuhn KL, Near J (2009) Phylogeny of trematomus (Notothenioidei: Nototheniidae) inferred from mitochondrial and nuclear gene sequences. Antarct Sci 21:565–570
La Mesa M, Vacchi M, Iwami T, Eastman JT (2002) Taxonomic studies of the Antarctic icefish genus Cryodraco Dollo, 1900 (Nototheniodei: Channichthidae). Polar Biol 25:384–390
Lautrédou A-C, Bonillo C, Denis G, Cruaud C, Ozouf-Costaz C, Lecointre G, Dettai A (2010) Molecular taxonomy and identification within the Antarctic genus Trematomus (Notothenioidei, Teleostei): how valuable is barcoding with COI? Polar Sci 4:333–352
Lecointre G, Gallut C, Bonillo C, Couloux A, Ozouf-Costaz C, Dettai A (2011) The Antarctic Fish Genus Artedidraco is paraphyletic (Teleostei, Notothenioidei, Artedidraconidae). Polar Biol 34:1135–1145
Long DJ (1994) Quaternary colonization or Paleogene persistence?: historical biogeography of skates (Chondrichthys: Rajidae) in the Antarctic ichthyofauna. Palaeobiology 20:215–228
Meyer C, Paulay G (2005) DNA Barcoding: error rates based on comprehensive sampling. PLoS Biol 3:2229–2238
Møller P, Nielsen, Anderson M (2005) Systematics of polar fishes. Fish Physiol 22:25–78
Moritz C, Cicero C (2004) DNA barcoding: promises and pitfalls. PLoS Biol 2:1529–1531
Near TJ, Pesavento JJ, Cheng CH (2003) Mitochondrial DNA, morphology, and the phylogenetic relationships of Antarctic icefishes (Notothenioidei: Channichthyidae). Mol Phylogenet Evol 28:87–98
Near TJ, Pesavento JJ, Cheng CC (2004) Phylogenetic investigations of Antarctic notothenioid fishes (Perciformes: Notothenioidei) using complete gene sequences of the mitochondrial encoded 16S rRNA. Mol Phylogenet Evol 32:881–891
Orsi AH, Whitworth T, Nowlin WD (1995) On the meridional extent and fronts of the Antarctic circumpolar current. Deep Sea Res 42:641–673
Parker RW, Paige KN, DeVries AL (2002) Genetic variation among populations of the Antarctic toothfish: evolutionary insights and implications for conservation. Polar Biol 25:256–261
Pollard D, DeConto RM (2009) Modelling West Antarctic ice sheet growth and collapse through the past five million years. Nature 458:329–333
Posada D (2008) jModelTest: phylogenetic model averaging. Mol Biol Evol 25:1253–1256
Ratnasingham S, Hebert PDN (2007) The barcode of life database. Mol Ecol Notes 7:355–364
Ritchie PA, Bargelloni L, Meyer A, Taylor JA, MacDonald JA, Lambert DM (1996) Mitochondrial phylogeny of trematomid fishes (Nototheniidae, Perciformes) and the evolution of antarctic fish. Mol Phylogenet Evol 5:383–390
Rock J, Costa FO, Walker DI, North AW, Hutchinson WF, Carvalho GR (2008) DNA barcodes of fish of the Scotia Sea, Antarctica indicate priority groups for taxonomic and systematics focus. Antarct Sci 20:253–262
Rubinoff D, Cameron S, Will K (2006) A genomic perspective on the shortcomings of mitochondrial DNA for “barcoding” identification. J Hered 97:581–594
Sanchez S, Dettai A, Bonillo C, Ozouf-Costaz C, Detrich B, Lecointre G (2007) Molecular and morphological phylogenies of the Antarctic teleostean family Nototheniidae, with emphasis on the Trematominae. Polar Biol 30:155–166
Sevilla RG, Diez A, Noren M, Mouchel O, Jerome M, Verrez-Bagnis V, Van Pelt H, Favre KL, Krey G, Bautista JM (2007) Primers and polymerase chain reaction conditions for DNA barcoding teleost fish based on the mitochondrial cytochrome b and nuclear rhodopsin genes. Mol Ecol Notes 7:730–734
Shaw P, Arkhipkin A, Al-Khairulla H (2004) Genetic structuring of Patagonian toothfish populations in the Southwest Atlantic Ocean: the effect of the Antarctic Polar Front and deep-water troughs as barriers to genetic exchange. Mol Ecol 13:3293–3303
Shearer T, Coffroth M (2008) Barcoding corals: limited by interspecific divergence, not intraspecific variation. Mol Ecol Res 8:247–255
Smith PJ, Gaffney P (2005) Low genetic diversity in the Antarctic toothfish Dissostichus mawsoni observed with mitochondrial and intron DNA markers. CCAMLR Sci 12:43–51
Smith PJ, McVeagh SM (2000) Allozyme and microsatellite DNA markers of toothfish population structure in the Southern Ocean. J Fish Biol 57:72–83
Smith PJ, Steinke D, McVeagh SM, Stewart AL, Struthers CD, Roberts CD (2008) Molecular analysis of Southern Ocean skates (Bathyraja) reveals a new species of Antarctic skate. J Fish Biol 73:1170–1182
Smith PJ, Steinke D, McMillan PJ, Stewart AL, McVeagh SM, Diaz de Astarloa JM, Welsford D, Ward RD (2011a) DNA barcoding highlights a cryptic species of grenadier (genus Macrourus) in the Southern Ocean. J Fish Biol 78:355–365
Smith PJ, Steinke D, McMillan PJ, Stewart AL, Ward RD (2011b) DNA barcoding of morids (Actinopterygii, Moridae) reveals deep divergence in the anti tropical Halargyreus johnsonii but little distinction between Antimora rostrata and A. microlepis. Mitochondrial DNA 22(S1):21–26
Steinke D, Zemlak TS, Boutillier JA, Hebert PDN (2009) DNA barcoding of Pacific Canada’s fishes. Mar Biol 156:2641–2647
Swofford DL (2003) PAUP*: phylogenetic analysis using parsimony (*and other methods). Version 4. Sinauer Associates, Sunderland, MA, United States
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecular evolutionary genetics analysis (MEGA) software version 4.0. Mol Biol Evol 24:1596–1599
Thornhill DJ, Mahon AR, Norenburg JL, Halanych KM (2008) Open-ocean barriers to dispersal: a test case with the Antarctic Polar Front and the ribbon worm Parborlasia corrugatus (Nemertea: Lineidae). Mol Ecol 17:5104–5117
Van de Putte AP, Van Houdt JKJ, Maes GE, Hellemans B, Collins MA, Volckaert FAM (2012) High genetic diversity and connectivity in a common mesopelagic fish of the Southern Ocean: the myctophid Electrona antarctica. Deep Sea Res II Top Stud Oceanogr 59–60:199–207
Ward RD (2009) DNA barcode divergence among species and genera of birds and fishes. Mol Ecol Res 9:1077–1085
Ward RD, Holmes B, O’Hara TD (2008) DNA barcoding discriminates echinoderm species. Mol Ecol Notes 8:1202–1211
Ward RD, Hanner R, Hebert PDN (2009) The campaign to DNA barcode all fishes, FISH-BOL. J Fish Biol 74:329–356
Waugh J (2007) DNA barcoding in animal species: progress, potential and pitfalls. BioEssays 29:188–197
Wilson NG, Schrödl M, Halanych KM (2009) Ocean barriers and glaciation: explosive radiation of Pleistocene lineages in the Antarctic sea slug Doris kerguelenensis (Mollusca, Nudibranchia). Mol Ecol 18:965–984
Zane L, Marcato S, Bargelloni L, Bortolotto E, Papetti C, Simonato M, Varotto V, Patarnello T (2006) Demographic history and population structure of the Antarctic silverfish Pleuragramma antarcticum. Mol Ecol 15:4499–4511
Acknowledgments
We are grateful to the New Zealand Ministry of Fisheries and CCAMLR Scientific Observers, and to the Australian Fisheries Management Authority observers for the collection of specimens in the commercial fisheries in the Ross Sea region, the Australian Antarctic Territory, and the Indian Ocean. Additional Ross Sea IPY specimens were collected and identified by Peter McMillan and Andrew Stewart aboard the NIWA Research vessel Tangaroa. Peter Smith, Peter McMillan, and Andrew Stewart were supported by the New Zealand Government under the NZ International Polar Year-Census of Antarctic Marine Life Project and acknowledge the Ministry of Fisheries Science Team and Ocean Survey 20/20 CAML Advisory Group (Land Information New Zealand, Ministry of Fisheries, Antarctica New Zealand, Ministry of Foreign Affairs and Trade, and National Institute of Water and Atmospheric Research Ltd.). D. Steinke and laboratory analyses of sequences generated at the Canadian Centre for DNA Barcoding (CCDB) were supported by funding of the Alfred P. Sloan Foundation to MarBOL and by the Government of Canada through Genome Canada, through the Ontario Genomics Institute (2008-0GI-ICI-03). Preservation and storage costs for voucher specimens from the Ross Sea registered into the Museum of New Zealand Te Papa Tongarewa were provided by the Te Papa Collection Development programme. The collection and study of the East Antarctic specimens from the CAML-CEAMARC cruises of the RSV Aurora Australis and the TRV Umitaka Maru (IPY project no. 53) were supported by the Australian Antarctic Division, the Japanese Science Foundation, the French polar institute IPEV (ICOTA and REVOLTA programs), the CNRS, the MNHN, and the ANR (White Project ANTFLOCKs USAR no. 07-BLAN-0213-01 directed by Guillaume Lecointre). We are grateful to Anton Van de Putte (Leuven, Belgium), Olly Berry, and Sharon Appleyard (Hobart, Australia), and two anonymous referees for constructive comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
300_2012_1173_MOESM1_ESM.xls
Appendix 1 Southern Ocean fishes barcoded listed by Species, Specimen Reference Number, GenBank Accession Number, Bold Process Number, and Institute storing specimens. The list excludes published data on Macrourus, available in Smith et al. (2011) (XLS 240 kb)
300_2012_1173_MOESM2_ESM.pdf
Appendix 2 COI relationships among the Artedidraconidae (154 specimens, including those identified to genus level only) from the Southern Ocean; unrooted ML tree. The scale bar represents an interval of the K2P model; numbers at nodes are bootstrap percentages (>75%). Sequence numbers represent BOLD Process Number, GenBank Accession Number, followed by Registration Number, and species name (PDF 37 kb)
300_2012_1173_MOESM3_ESM.pdf
Appendix 3 COI relationships among Trematomus specimens (181 specimens, including those identified to genus level only); unrooted ML tree. The scale bar represents an interval of the K2P model; numbers at nodes are bootstrap percentages (>75%). Sequence numbers represent BOLD Process Number, GenBank Accession Number, followed by Registration Number, and species name (PDF 42 kb)
300_2012_1173_MOESM4_ESM.pdf
Appendix 4 COI relationships among the Bathydraconidae (168 specimens, including those identified to genus level only) from the Southern Ocean; unrooted ML tree. The scale bar represents an interval of the K2P +G model; numbers at nodes are bootstrap percentages (>75%). Sequence numbers represent BOLD Process Number, GenBank Accession Number, followed by Registration Number, and species name (PDF 39 kb)
Rights and permissions
About this article
Cite this article
Smith, P.J., Steinke, D., Dettai, A. et al. DNA barcodes and species identifications in Ross Sea and Southern Ocean fishes. Polar Biol 35, 1297–1310 (2012). https://doi.org/10.1007/s00300-012-1173-8
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00300-012-1173-8